
Stem Cell-Derived Viral Antigen-Specific T Cells Suppress HIV Replication and PD-1 Expression on CD4+ T Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
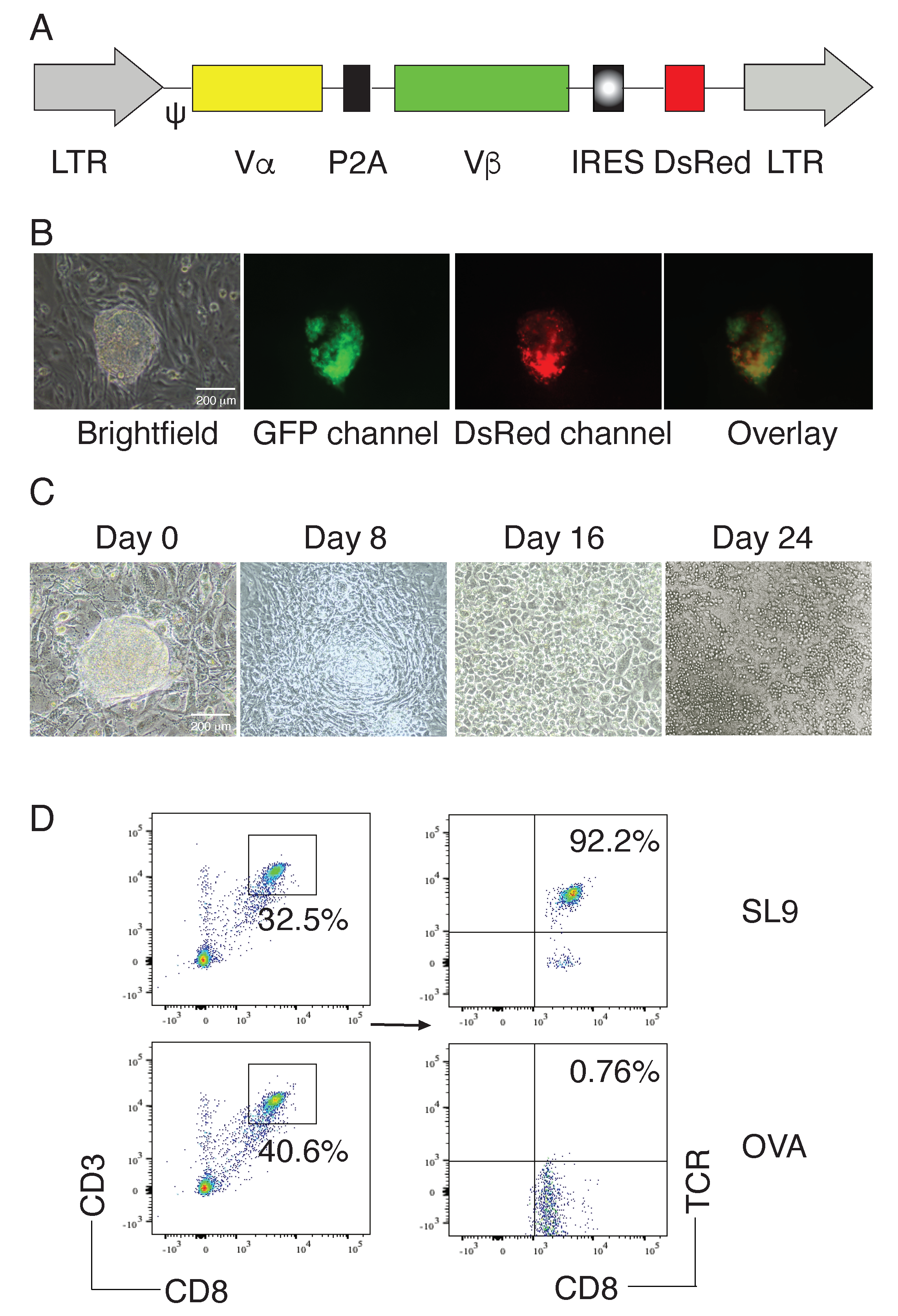
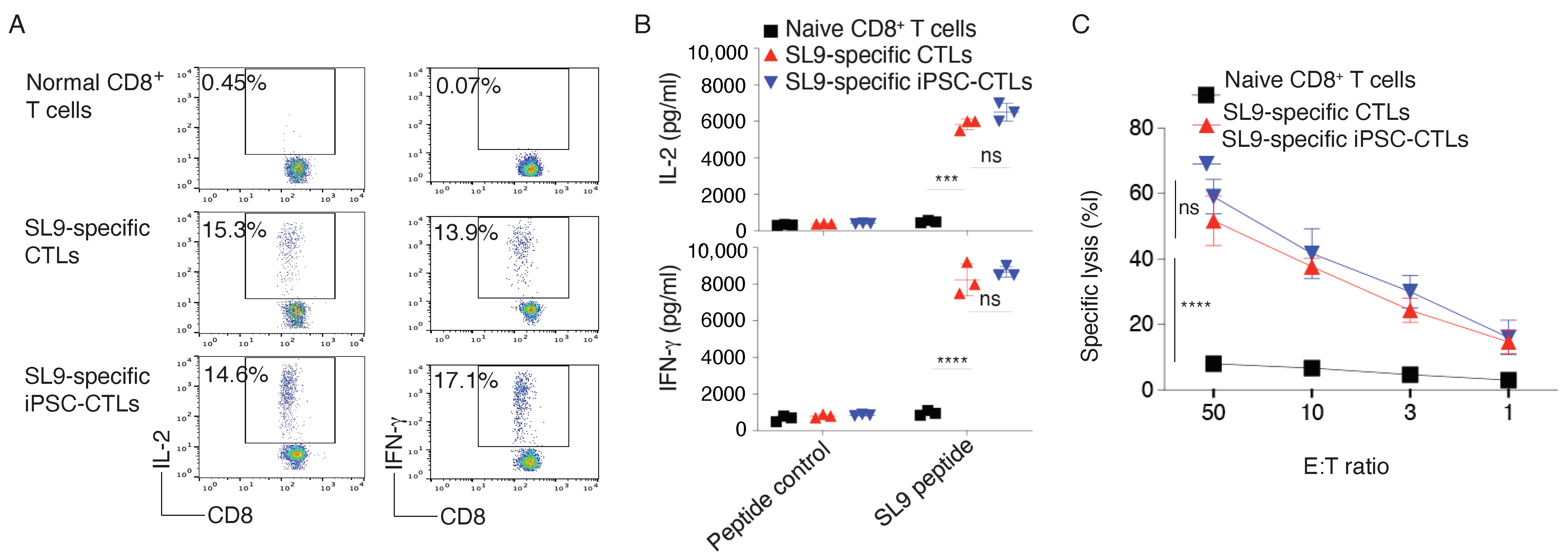
2.1. Generation of HIV-Specific iPSC-CTLs
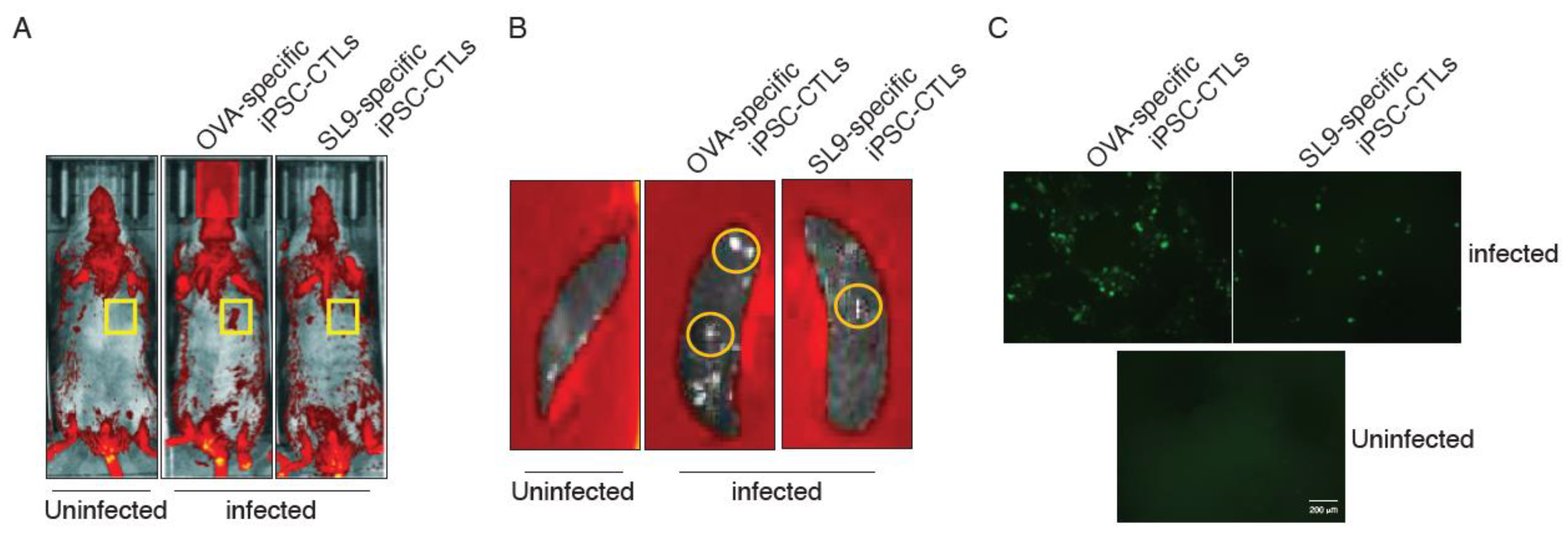
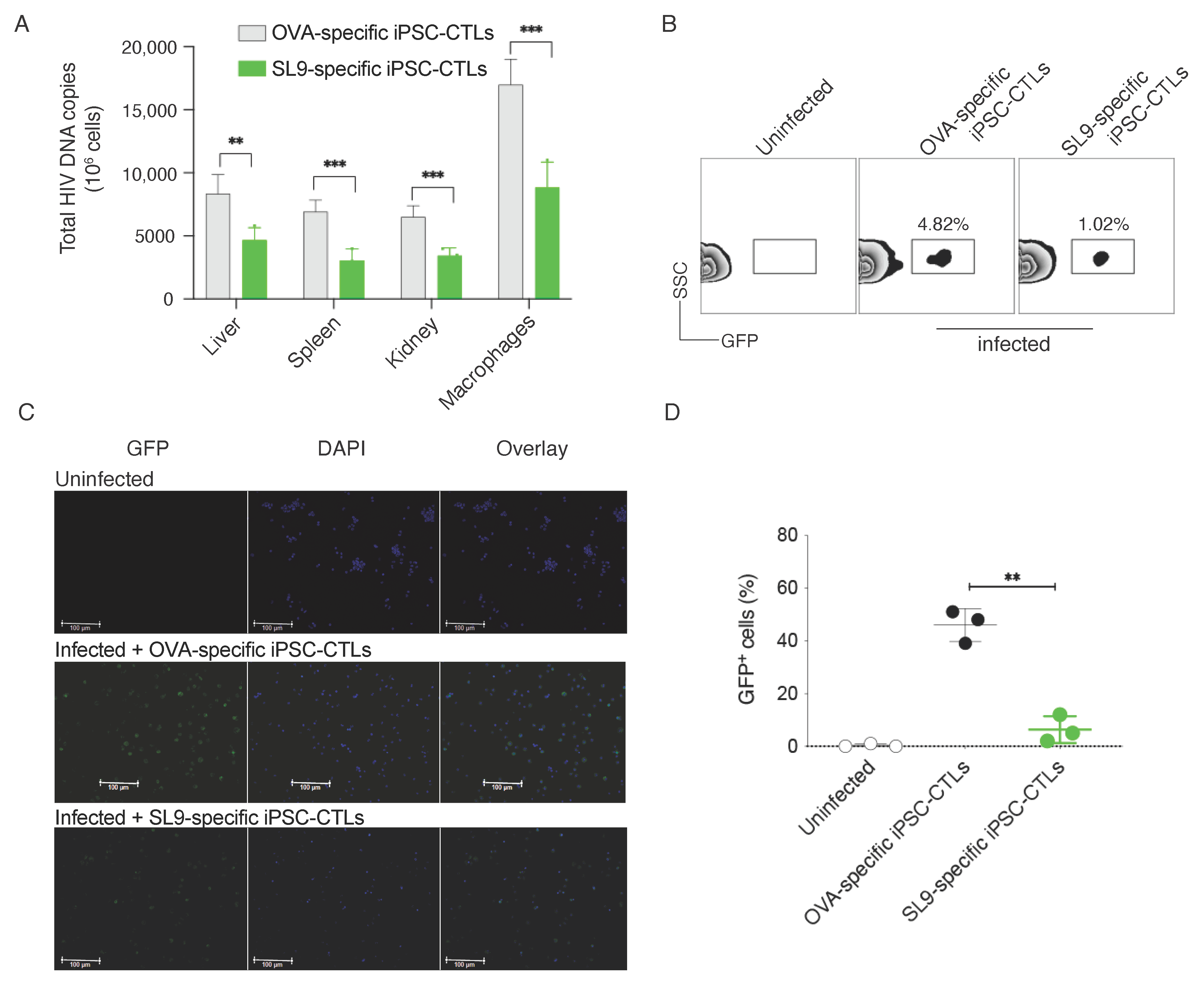
2.2. EcoHIV Replication in Mice
2.3. Adoptive Transfer of SL9-Specific iPSC-CTLs Reduced EcoHIV Replication
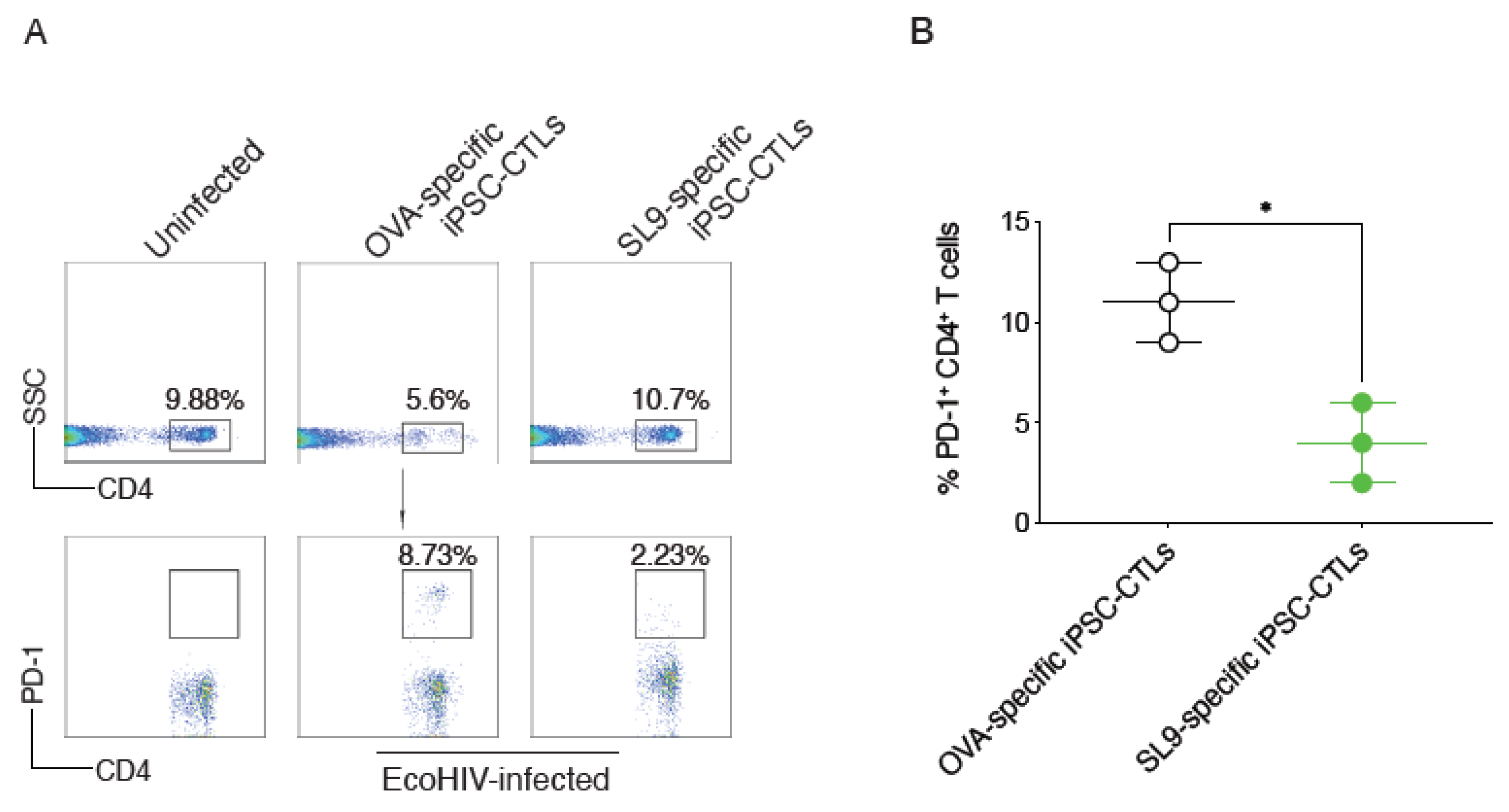
2.4. SL9-Specific iPSC-CTLs Suppressed PD-1 Expression on CD4+ T Cells
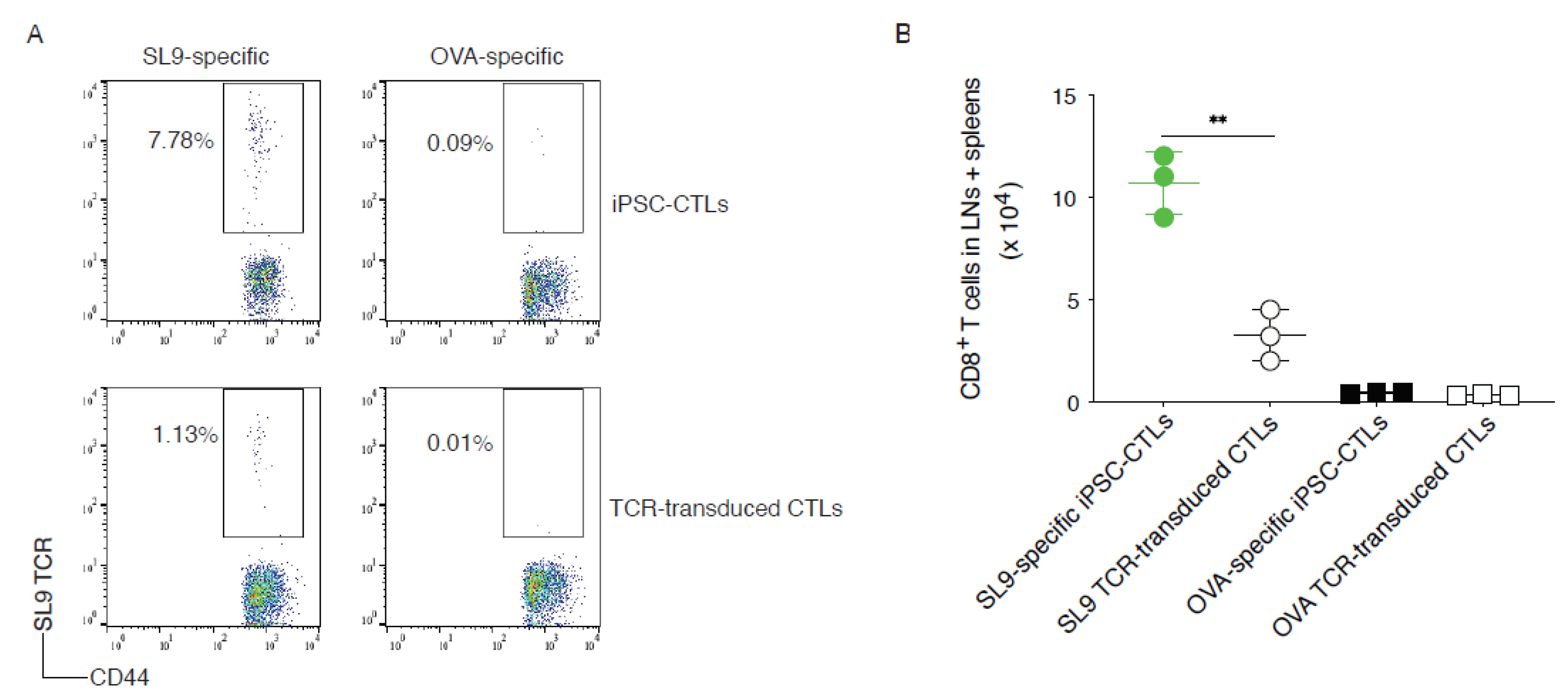
2.5. SL9-Specific iPSC-CTLs Persist In Vivo
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Cell Lines and Mouse
4.3. Cell Culture
4.4. EcoHIV Infection
4.5. Retroviral Transduction and Generation of Ag-Specific iPSC-CTLs
4.6. Antibodies
4.7. Flow Cytometric Analysis
4.8. Adoptive Cell Transfer
4.9. QPCR
4.10. Confocal Microscopy
4.11. Live Mice Imaging
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maina, E.K.; Adan, A.A.; Mureithi, H.; Muriuki, J.; Lwembe, R.M. A review of current strategies towards the elimination of latent hiv-1 and subsequent hiv-1 cure. Curr. HIV Res. 2021, 19, 14–26. [Google Scholar] [CrossRef]
- Ambinder, R.F.; Capoferri, A.A.; Durand, C.M. Haemopoietic cell transplantation in patients living with hiv. Lancet HIV 2020, 7, e652–e660. [Google Scholar] [CrossRef]
- Korencak, M.; Byrne, M.; Richter, E.; Schultz, B.T.; Juszczak, P.; Ake, J.A.; Ganesan, A.; Okulicz, J.F.; Robb, M.L.; de Los Reyes, B.; et al. Effect of hiv infection and antiretroviral therapy on immune cellular functios. JCI Insight 2019, 4, e126675. [Google Scholar] [CrossRef] [PubMed]
- Tenore, S.B.; Avelino-Silva, V.I.; Costa, P.R.; Franco, L.M.; Sabino, E.C.; Kalil, J.; Cerqueira, N.B.; Nakagawa, Z.; Kallas, E.G. Immune effects of lactobacillus casei shirota in treated hiv-infected patients with poor cd4+ t-cell recovery. AIDS 2020, 34, 381–389. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Hou, S.; Liu, C.; Zhang, A.; Bai, Q.; Han, M.; Yang, Y.; Wei, G.; Shen, T.; Yang, X.; et al. Follicular cxcr5- expressing cd8(+) t cells curtail chronic viral infection. Nature 2016, 537, 412–428. [Google Scholar] [CrossRef]
- Collins, D.R.; Gaiha, G.D.; Walker, B.D. Cd8(+) t cells in hiv control, cure and prevention. Nat. Rev. Immunol. 2020, 20, 471–482. [Google Scholar] [CrossRef]
- Abram, M.E.; Ferris, A.L.; Das, K.; Quinones, O.; Shao, W.; Tuske, S.; Alvord, W.G.; Arnold, E.; Hughes, S.H. Mutations in hiv-1 reverse transcriptase affect the errors made in a single cycle of viral replication. J. Virol. 2014, 88, 7589–7601. [Google Scholar] [CrossRef] [Green Version]
- Dalel, J.; Ung, S.K.; Hayes, P.; Black, S.L.; Joseph, S.; King, D.F.; Makinde, J.; Gilmour, J. Hiv-1 infection and the lack of viral control are associated with greater expression of interleukin-21 receptor on cd8+ t cells. AIDS 2021. [Google Scholar] [CrossRef]
- Gomez, C.E.; Perdiguero, B.; Falqui, M.; Marin, M.Q.; Becares, M.; Sorzano, C.O.S.; Garcia-Arriaza, J.; Esteban, M.; Guerra, S. Enhancement of hiv-1 env-specific cd8 t cell responses using interferon-stimulated gene 15 as an immune adjuvant. J. Virol. 2020, 95. [Google Scholar] [CrossRef] [PubMed]
- Qasim, W.; Brunetto, M.; Gehring, A.J.; Xue, S.A.; Schurich, A.; Khakpoor, A.; Zhan, H.; Ciccorossi, P.; Gilmour, K.; Cavallone, D.; et al. Immunotherapy of hcc metastases with autologous t cell receptor redirected t cells, targeting hbsag in a liver transplant patient. J. Hepatol. 2014, 62, 486–491. [Google Scholar] [CrossRef]
- Migueles, S.A.; Chairez, C.; Lin, S.; Gavil, N.V.; Rosenthal, D.M.; Pooran, M.; Natarajan, V.; Rupert, A.; Dewar, R.; Rehman, T.; et al. Adoptive lymphocyte transfer to an hiv-infected progressor from an elite controller. JCI Insight 2019, 4, e130664. [Google Scholar] [CrossRef] [Green Version]
- Hinrichs, C.S.; Borman, Z.A.; Gattinoni, L.; Yu, Z.; Burns, W.R.; Huang, J.; Klebanoff, C.A.; Johnson, L.A.; Kerkar, S.P.; Yang, S.; et al. Human effector cd8+ t cells derived from naive rather than memory subsets possess superior traits for adoptive immunotherapy. Blood 2011, 117, 808–814. [Google Scholar] [CrossRef] [Green Version]
- Kerkar, S.P.; Sanchez-Perez, L.; Yang, S.; Borman, Z.A.; Muranski, P.; Ji, Y.; Chinnasamy, D.; Kaiser, A.D.; Hinrichs, C.S.; Klebanoff, C.A.; et al. Genetic engineering of murine cd8+ and cd4+ t cells for preclinical adoptive immunotherapy studies. J. Immunother. 2011, 34, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Maldini, C.R.; Gayout, K.; Leibman, R.S.; Dopkin, D.L.; Mills, J.P.; Shan, X.; Glover, J.A.; Riley, J.L. Hiv-resistant and hiv-specific car-modified cd4(+) t cells mitigate hiv disease progression and confer cd4(+) t cell help in vivo. Mol. Ther. 2020, 28, 1585–1599. [Google Scholar] [CrossRef]
- Ayala, V.I.; Trivett, M.T.; Barsov, E.V.; Jain, S.; Piatak, M., Jr.; Trubey, C.M.; Alvord, W.G.; Chertova, E.; Roser, J.D.; Smedley, J.; et al. Adoptive transfer of engineered rhesus simian immunodeficiency virus-specific cd8+ t cells reduces the number of transmitted/founder viruses established in rhesus macaques. J. Virol. 2016, 90, 9942–9952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, F.; Zhao, B.; Haque, R.; Xiong, X.; Budgeon, L.; Christensen, N.D.; Wu, Y.; Song, J. In vivo programming of tumor antigen-specific t lymphocytes from pluripotent stem cells to promote cancer immunosurveillance. Cancer Res. 2011, 71, 4742–4747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, A.; Zheng, J.H.; Follenzi, A.; Dilorenzo, T.; Sango, K.; Hyman, J.; Chen, K.; Piechocka-Trocha, A.; Brander, C.; Hooijberg, E.; et al. Lentiviral vectors encoding human immunodeficiency virus type 1 (hiv-1)-specific t-cell receptor genes efficiently convert peripheral blood cd8 t lymphocytes into cytotoxic t lymphocytes with potent in vitro and in vivo hiv-1-specific inhibitory activity. J. Virol. 2008, 82, 3078–3089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potash, M.J.; Chao, W.; Bentsman, G.; Paris, N.; Saini, M.; Nitkiewicz, J.; Belem, P.; Sharer, L.; Brooks, A.I.; Volsky, D.J. A mouse model for study of systemic hiv-1 infection, antiviral immune responses, and neuroinvasiveness. Proc. Natl. Acad. Sci. USA 2005, 102, 3760–3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.J.; Borjabad, A.; Hadas, E.; Kelschenbach, J.; Kim, B.H.; Chao, W.; Arancio, O.; Suh, J.; Polsky, B.; McMillan, J.; et al. Ecohiv infection of mice establishes latent viral reservoirs in t cells and active viral reservoirs in macrophages that are sufficient for induction of neurocognitive impairment. PLoS Pathog. 2018, 14, e1007061. [Google Scholar] [CrossRef] [Green Version]
- Porichis, F.; Hart, M.G.; Zupkosky, J.; Barblu, L.; Kwon, D.S.; McMullen, A.; Brennan, T.; Ahmed, R.; Freeman, G.J.; Kavanagh, D.G.; et al. Differential impact of pd-1 and/or interleukin-10 blockade on hiv-1-specific cd4 t cell and antigen-presenting cell functions. J. Virol. 2014, 88, 2508–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leibman, R.S.; Riley, J.L. Engineering t cells to functionally cure hiv-1 infection. Mol. Ther. J. Am. Soc. Gene Ther. 2015, 23, 1149–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Loenen, M.M.; de Boer, R.; Amir, A.L.; Hagedoorn, R.S.; Volbeda, G.L.; Willemze, R.; van Rood, J.J.; Falkenburg, J.H.; Heemskerk, M.H. Mixed t cell receptor dimers harbor potentially harmful neoreactivity. Proc. Natl. Acad. Sci. USA 2010, 107, 10972–10977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stadtmauer, E.A.; Fraietta, J.A.; Davis, M.M.; Cohen, A.D.; Weber, K.L.; Lancaster, E.; Mangan, P.A.; Kulikovskaya, I.; Gupta, M.; Chen, F.; et al. Crispr-engineered t cells in patients with refractory cancer. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Echchannaoui, H.; Petschenka, J.; Ferreira, E.A.; Hauptrock, B.; Lotz-Jenne, C.; Voss, R.H.; Theobald, M. A potent tumor-reactive p53-specific single-chain tcr without on- or off-target autoimmunity in vivo. Mol. Ther. 2019, 27, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgiadis, C.; Preece, R.; Nickolay, L.; Etuk, A.; Petrova, A.; Ladon, D.; Danyi, A.; Humphryes-Kirilov, N.; Ajetunmobi, A.; Kim, D.; et al. Long terminal repeat crispr-car-coupled “universal” t cells mediate potent anti-leukemic effects. Mol. Ther. 2018, 26, 1215–1227. [Google Scholar] [CrossRef] [Green Version]
- Gudiol, C.; Lewis, R.E.; Strati, P.; Kontoyiannis, D.P. Chimeric antigen receptor t-cell therapy for the treatment of lymphoid malignancies: Is there an excess risk for infection? Lancet Haematol. 2021, 8, e216–e228. [Google Scholar] [CrossRef]
- Haque, R.; Lei, F.; Xiong, X.; Bian, Y.; Zhao, B.; Wu, Y.; Song, J. Programming of regulatory t cells from pluripotent stem cells and prevention of autoimmunity. J. Immunol. 2012, 189, 1228–1236. [Google Scholar] [CrossRef] [Green Version]
- Vizcardo, R.; Masuda, K.; Yamada, D.; Ikawa, T.; Shimizu, K.; Fujii, S.; Koseki, H.; Kawamoto, H. Regeneration of human tumor antigen-specific t cells from ipscs derived from mature cd8(+) t cells. Cell Stem Cell 2013, 12, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Kaneko, S.; Kawana-Tachikawa, A.; Tajima, Y.; Goto, H.; Zhu, D.; Nakayama-Hosoya, K.; Iriguchi, S.; Uemura, Y.; Shimizu, T.; et al. Generation of rejuvenated antigen-specific t cells by reprogramming to pluripotency and redifferentiation. Cell Stem Cell 2013, 12, 114–126. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wen, R.; Yang, Z.; Chen, Z. Genome editing using crispr/cas9 to treat hereditary hematological disorders. Gene Ther. 2021, 1–10. [Google Scholar] [CrossRef]
- Haque, M.; Lei, F.; Xiong, X.; Ren, Y.; Kumar, A.; Das, J.K.; Ren, X.; Fang, D.; de Figueiredo, P.; Yang, J.M.; et al. Stem cell-derived viral antigen-specific t cells suppress hbv replication through production of ifn-gamma and tnf. iScience 2020, 23, 101333. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.; Lei, F.; Xiong, X.; Das, J.K.; Ren, X.; Fang, D.; Salek-Ardakani, S.; Yang, J.M.; Song, J. Stem cell-derived tissue-associated regulatory t cells suppress the activity of pathogenic cells in autoimmune diabetes. JCI Insight 2019, 4, e126471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony-Gonda, K.; Bardhi, A.; Ray, A.; Flerin, N.; Li, M.; Chen, W.; Ochsenbauer, C.; Kappes, J.C.; Krueger, W.; Worden, A.; et al. Multispecific anti-hiv duocar-t cells display broad in vitro antiviral activity and potent in vivo elimination of hiv-infected cells in a humanized mouse model. Sci. Transl. Med. 2019, 11, eaav5685. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.; Song, J.; Fino, K.; Sandhu, P.; Song, X.; Lei, F.; Zheng, S.; Ni, B.; Fang, D.; Song, J. Stem cell-derived tissue-associated regulatory t cells ameliorate the development of autoimmunity. Sci. Rep. 2016, 6, 20588. [Google Scholar] [CrossRef] [Green Version]
- Kah, J.; Koh, S.; Volz, T.; Ceccarello, E.; Allweiss, L.; Lutgehetmann, M.; Bertoletti, A.; Dandri, M. Lymphocytes transiently expressing virus-specific t cell receptors reduce hepatitis b virus infection. J. Clin. Investig. 2017, 127, 3177–3188. [Google Scholar] [CrossRef]
- Khakpoor, A.; Ni, Y.; Chen, A.; Ho, Z.Z.; Oei, V.; Yang, N.; Giri, R.; Chow, J.X.; Tan, A.T.; Kennedy, P.T.; et al. Spatiotemporal differences in presentation of cd8 t cell epitopes during hepatitis b virus infection. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Song, A.; Haque, R.; Lei, F.; Weiler, L.; Xiong, X.; Wu, Y.; Croft, M.; Song, J. Cooperation between molecular targets of costimulation in promoting t cell persistence and tumor regression. J. Immunol. 2009, 182, 6744–6752. [Google Scholar] [CrossRef] [Green Version]
- Hadas, E.; Borjabad, A.; Chao, W.; Saini, M.; Ichiyama, K.; Potash, M.J.; Volsky, D.J. Testing antiretroviral drug efficacy in conventional mice infected with chimeric hiv-1. AIDS 2007, 21, 905–909. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haque, M.; Lei, F.; Xiong, X.; Ren, Y.; Peng, H.-Y.; Wang, L.; Kumar, A.; Das, J.K.; Song, J. Stem Cell-Derived Viral Antigen-Specific T Cells Suppress HIV Replication and PD-1 Expression on CD4+ T Cells. Viruses 2021, 13, 753. https://doi.org/10.3390/v13050753
Haque M, Lei F, Xiong X, Ren Y, Peng H-Y, Wang L, Kumar A, Das JK, Song J. Stem Cell-Derived Viral Antigen-Specific T Cells Suppress HIV Replication and PD-1 Expression on CD4+ T Cells. Viruses. 2021; 13(5):753. https://doi.org/10.3390/v13050753
Chicago/Turabian StyleHaque, Mohammad, Fengyang Lei, Xiaofang Xiong, Yijie Ren, Hao-Yun Peng, Liqing Wang, Anil Kumar, Jugal Kishore Das, and Jianxun Song. 2021. "Stem Cell-Derived Viral Antigen-Specific T Cells Suppress HIV Replication and PD-1 Expression on CD4+ T Cells" Viruses 13, no. 5: 753. https://doi.org/10.3390/v13050753