Chicken eEF1α is a Critical Factor for the Polymerase Complex Activity of Very Virulent Infectious Bursal Disease Virus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, Antibodies, and Regents
2.2. Construction of Plasmids
2.3. RNAi and Transfection
2.4. Virus Infection and ELD50 Titration
2.5. Immunoprecipitation, SDS-PAGE, and Silver Staining
2.6. Co-Immunoprecipitation and Western Blot
2.7. RNA Extraction and RT-qPCR
2.8. Confocal Microscopy
2.9. IBDV Minigenome System for Detection of Polymerase Activity
2.10. Statistical Analysis
3. Results
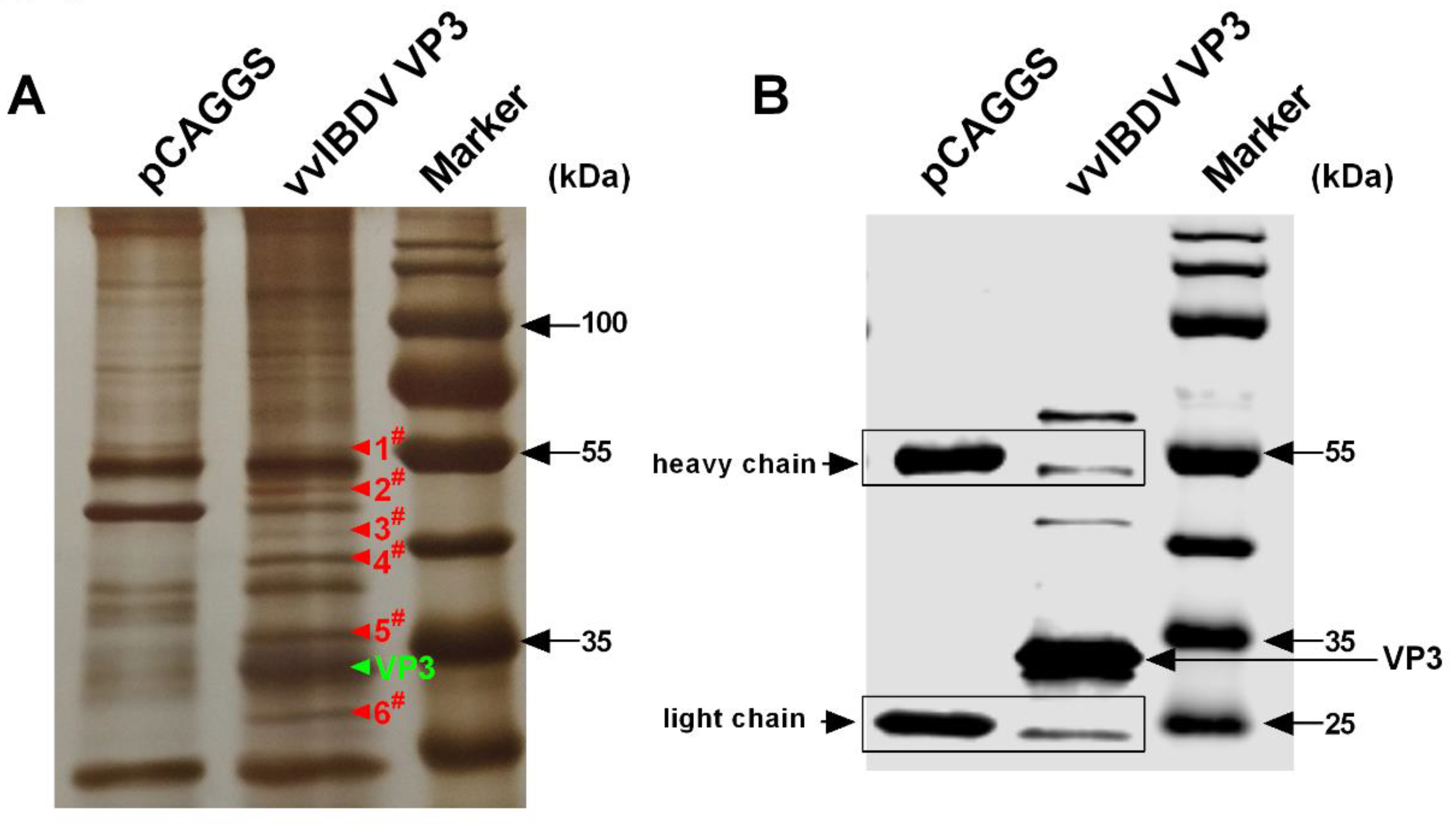
3.1. Host Proteins Immunoprecipitated with vvIBDV VP3
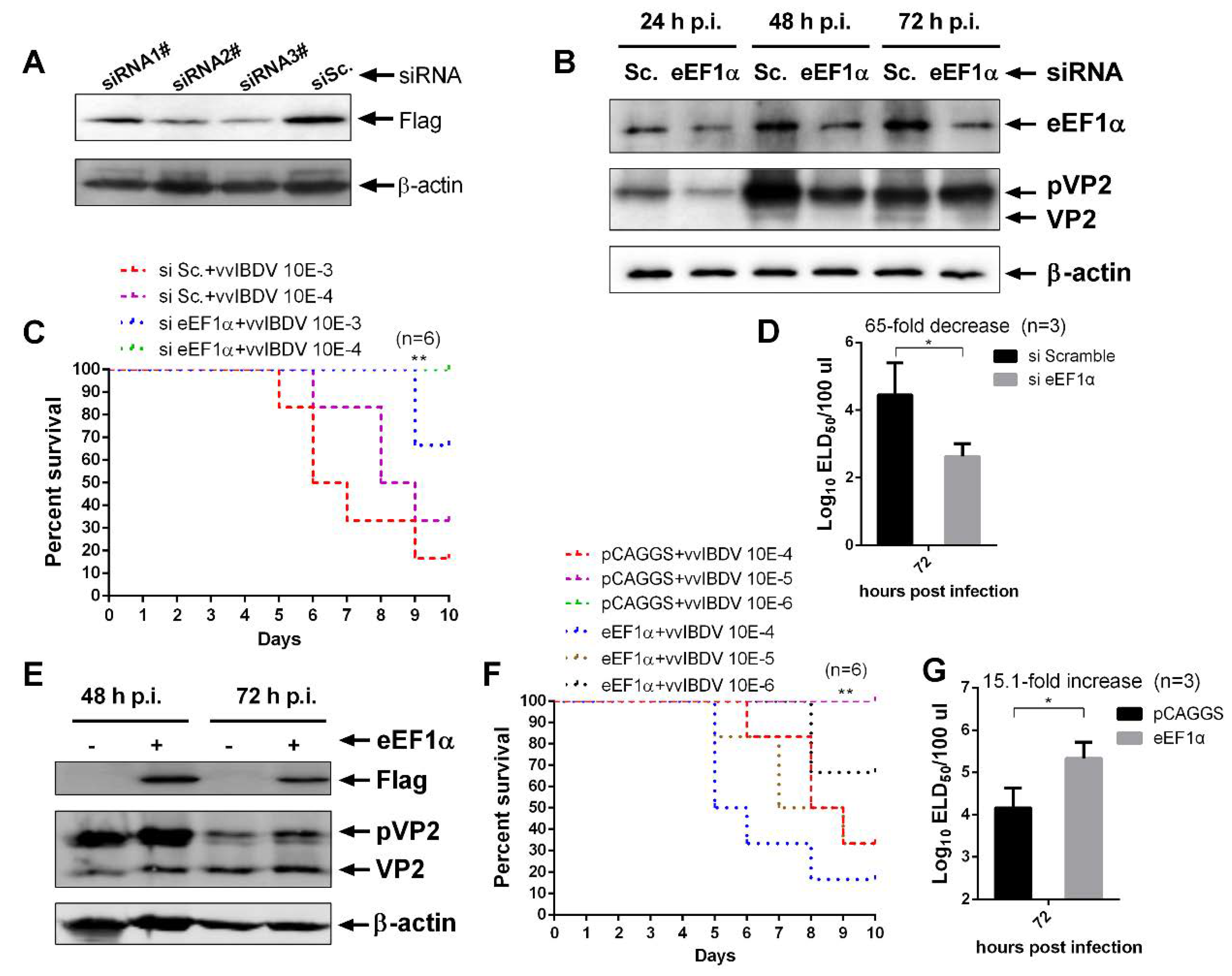
3.2. cheEF1α is Critical for vvIBDV Replication
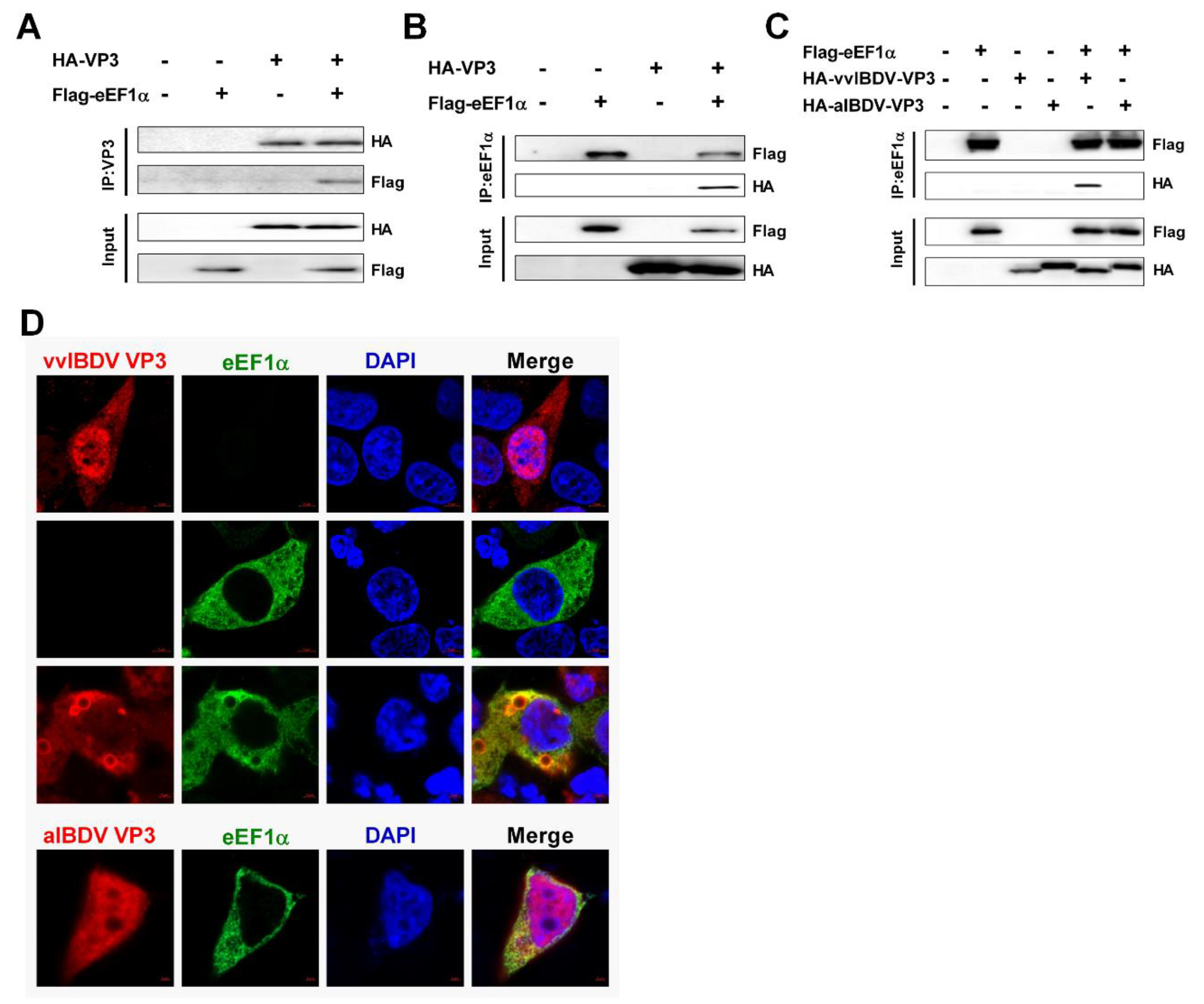
3.3. cheEF1α Interacts with vvIBDV VP3, but not with aIBDV VP3
3.4. Both N- and C-Terminal Amino Acids of VP3 Are Essential in the Interaction Between vvIBDV VP3 and cheEF1α
3.5. The Third Domain of cheEF1α Interacted with vvIBDV VP3
3.6. cheEF1α Promotes the Activity of vvIBDV Polymerase
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cosgrove, A.S. An Apparently New Disease of Chickens: Avian Nephrosis. Avian Dis. 1962, 6, 385–389. [Google Scholar] [CrossRef]
- Luque, D.; Rivas, G.; Alfonso, C.; Carrascosa, J.L.; Rodriguez, J.F.; Caston, J.R. Infectious bursal disease virus is an icosahedral polyploid dsRNA virus. Proc. Natl. Acad. Sci. USA 2009, 106, 2148–2152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, H.; Islam, M.R.; Raue, R. Research on infectious bursal disease—The past, the present and the future. Vet. Microbiol. 2003, 97, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Gao, H.; Fu, C.; Gao, Y.; Ju, Y. Changes in VP3 and VP5 genes during the attenuation of the very virulent infectious bursal disease virus strain Gx isolated in China. Virus Genes 2007, 34, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.H.; Saif, Y.M. Antigenic diversity of infectious bursal disease viruses. Avian Dis. 1987, 31, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Chettle, N.; Stuart, J.C.; Wyeth, P.J. Outbreak of virulent infectious bursal disease in East Anglia. Vet. Rec. 1989, 125, 271–272. [Google Scholar] [CrossRef]
- Nunoya, T.; Otaki, Y.; Tajima, M.; Hiraga, M.; Saito, T. Occurrence of acute infectious bursal disease with high mortality in Japan and pathogenicity of field isolates in specific-pathogen-free chickens. Avian Dis. 1992, 36, 597–609. [Google Scholar] [CrossRef]
- Berg, T.P.; Gonze, M.; Meulemans, G. Acute infectious bursal disease in poultry: Isolation and characterisation of a highly virulent strain. Avian Pathol. 1991, 20, 133–143. [Google Scholar] [CrossRef]
- Yuwen, Y.; Gao, Y.; Gao, H.; Qi, X.; Li, T.; Liu, W.; Wang, X. Sequence analysis of the VP2 hypervariable region of eight very virulent infectious bursal disease virus isolates from the northeast of China. Avian Dis. 2008, 52, 284–290. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, X.; Kang, Z.; Yu, F.; Qin, L.; Gao, H.; Gao, Y.; Wang, X. A single amino acid in the C-terminus of VP3 protein influences the replication of attenuated infectious bursal disease virus in vitro and in vivo. Antiviral Res. 2010, 87, 223–229. [Google Scholar] [CrossRef]
- Li, K.; Courtillon, C.; Guionie, O.; Allee, C.; Amelot, M.; Qi, X.; Gao, Y.; Wang, X.; Eterradossi, N. Genetic, antigenic and pathogenic characterization of four infectious bursal disease virus isolates from China suggests continued evolution of very virulent viruses. Infect. Genet. Evol. 2015, 30, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Qi, X.; Ren, X.; Cui, L.; Wang, X.; Zhu, P. Molecular characteristics and evolutionary analysis of a very virulent infectious bursal disease virus. Sci. China Life Sci. 2015, 58, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, R.; Zhu, S.; Wei, L.; Wang, J.; Yan, X.; Li, Z.; Liu, J. Transcriptional profiles in bursal B-lymphoid DT40 cells infected with very virulent infectious bursal disease virus. Virol. J. 2017, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulwich, K.L.; Giotis, E.S.; Gray, A.; Nair, V.; Skinner, M.A.; Broadbent, A.J. Differential gene expression in chicken primary B cells infected ex vivo with attenuated and very virulent strains of infectious bursal disease virus (IBDV). J. Gen. Virol. 2017, 98, 2918–2930. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.R.; Wang, M.Y.; Liao, J.H.; Hsiao, Y.P.; Lai, S.Y. Endopeptidase activity characterization of E. coli-derived infectious bursal disease virus protein 4 tubules. Protein Eng. Des. Sel. 2012, 25, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Mendez, F.; Romero, N.; Cubas, L.L.; Delgui, L.R.; Rodriguez, D.; Rodriguez, J.F. Non-Lytic Egression of Infectious Bursal Disease Virus (IBDV) Particles from Infected Cells. PLoS ONE 2017, 12, e0170080. [Google Scholar] [CrossRef]
- Lombardo, E.; Maraver, A.; Espinosa, I.; Fernandez-Arias, A.; Rodriguez, J.F. VP5, the nonstructural polypeptide of infectious bursal disease virus, accumulates within the host plasma membrane and induces cell lysis. Virology 2000, 277, 345–357. [Google Scholar] [CrossRef] [Green Version]
- Lombardo, E.; Maraver, A.; Caston, J.R.; Rivera, J.; Fernandez-Arias, A.; Serrano, A.; Carrascosa, J.L.; Rodriguez, J.F. VP1, the putative RNA-dependent RNA polymerase of infectious bursal disease virus, forms complexes with the capsid protein VP3, leading to efficient encapsidation into virus-like particles. J. Virol. 1999, 73, 6973–6983. [Google Scholar] [CrossRef] [Green Version]
- von Einem, U.I.; Gorbalenya, A.E.; Schirrmeier, H.; Behrens, S.E.; Letzel, T.; Mundt, E. VP1 of infectious bursal disease virus is an RNA-dependent RNA polymerase. J. Gen. Virol. 2004, 85, 2221–2229. [Google Scholar] [CrossRef]
- Mata, C.P.; Mertens, J.; Fontana, J.; Luque, D.; Allende-Ballestero, C.; Reguera, D.; Trus, B.L.; Steven, A.C.; Carrascosa, J.L.; Caston, J.R. The RNA-Binding Protein of a Double-Stranded RNA Virus Acts like a Scaffold Protein. J. Virol. 2018, 92, 19. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, D.; Rodriguez, J.F.; Abaitua, F. Intracellular interference of infectious bursal disease virus. J. Virol. 2005, 79, 14437–14441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacken, M.G.; Peeters, B.P.; Thomas, A.A.; Rottier, P.J.; Boot, H.J. Infectious bursal disease virus capsid protein VP3 interacts both with VP1, the RNA-dependent RNA polymerase, and with viral double-stranded RNA. J. Virol. 2002, 76, 11301–11311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrero, D.; Garriga, D.; Navarro, A.; Rodriguez, J.F.; Verdaguer, N. Infectious Bursal Disease Virus VP3 Upregulates VP1-Mediated RNA-Dependent RNA Replication. J. Virol. 2015, 89, 11165–11168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luque, D.; Saugar, I.; Rodriguez, J.F.; Verdaguer, N.; Garriga, D.; Martin, C.S.; Velazquez-Muriel, J.A.; Trus, B.L.; Carrascosa, J.L.; Caston, J.R. Infectious bursal disease virus capsid assembly and maturation by structural rearrangements of a transient molecular switch. J. Virol. 2007, 81, 6869–6878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraver, A.; Clemente, R.; Rodriguez, J.F.; Lombardo, E. Identification and molecular characterization of the RNA polymerase-binding motif of infectious bursal disease virus inner capsid protein VP3. J. Virol. 2003, 77, 2459–2468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevalier, C.; Lepault, J.; Erk, I.; Da Costa, B.; Delmas, B. The maturation process of pVP2 requires assembly of infectious bursal disease virus capsids. J. Virol. 2002, 76, 2384–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraver, A.; Ona, A.; Abaitua, F.; Gonzalez, D.; Clemente, R.; Ruiz-Diaz, J.A.; Caston, J.R.; Pazos, F.; Rodriguez, J.F. The oligomerization domain of VP3, the scaffolding protein of infectious bursal disease virus, plays a critical role in capsid assembly. J. Virol. 2003, 77, 6438–6449. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, C.; Lepault, J.; Da Costa, B.; Delmas, B. The last C-terminal residue of VP3, glutamic acid 257, controls capsid assembly of infectious bursal disease virus. J. Virol. 2004, 78, 3296–3303. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Wang, Y.; Zhang, E.; Han, X.; Yu, Z.; Liu, H. VP1 and VP3 Are Required and Sufficient for Translation Initiation of Uncapped Infectious Bursal Disease Virus Genomic Double-Stranded RNA. J. Virol. 2018, 92, 2. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Jia, L.; Sun, Y.; Hu, B.; Wang, L.; Lu, X.; Zhou, J. Inhibition of antiviral innate immunity by birnavirus VP3 protein via blockage of viral double-stranded RNA binding to the host cytoplasmic RNA detector MDA5. J. Virol. 2014, 88, 11154–11165. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Duan, X.; Fu, M.; Liu, Y.; Wang, Y.; Li, X.; Cao, H.; Zheng, S.J. The association of ribosomal protein L18 (RPL18) with infectious bursal disease virus viral protein VP3 enhances viral replication. Virus Res. 2018, 245, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Mateyak, M.K.; Kinzy, T.G. eEF1A: Thinking outside the ribosome. J. Biol. Chem. 2010, 285, 21209–21213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, Y.H.; Chou, S.M.; Wang, Y.M.; Chang, Y.T.; Huang, S.Y.; Jung, M.Y.; Huang, Y.H.; Chen, M.R.; Chang, M.F.; Chang, S.C. Hepatitis C virus NS4A inhibits cap-dependent and the viral IRES-mediated translation through interacting with eukaryotic elongation factor 1A. J. Biomed. Sci. 2006, 13, 861–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, K.; Wei, T.; Li, D.; Qin, F.; Warrilow, D.; Lin, M.H.; Sivakumaran, H.; Apolloni, A.; Abbott, C.M.; Jones, A.; et al. Eukaryotic elongation factor 1 complex subunits are critical HIV-1 reverse transcription cofactors. Proc. Natl. Acad. Sci. USA 2012, 109, 9587–9592. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wei, T.; Rawle, D.J.; Qin, F.; Wang, R.; Soares, D.C.; Jin, H.; Sivakumaran, H.; Lin, M.H.; Spann, K.; et al. Specific Interaction between eEF1A and HIV RT Is Critical for HIV-1 Reverse Transcription and a Potential Anti-HIV Target. PLoS Pathog. 2015, 11, e1005289. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Yang, D.; Gao, R.; Liang, T.; Wang, H.; Zhou, G.; Yu, L. Modification of the internal ribosome entry site element impairs the growth of foot-and-mouth disease virus in porcine-derived cells. J. Gen. Virol. 2016, 97, 901–911. [Google Scholar] [CrossRef]
- Yu, F.; Ren, X.; Wang, Y.; Qi, X.; Song, J.; Gao, Y.; Qin, L.; Gao, H.; Wang, X. A single amino acid V4I substitution in VP1 attenuates virulence of very virulent infectious bursal disease virus (vvIBDV) in SPF chickens and increases replication in CEF cells. Virology 2013, 440, 204–209. [Google Scholar] [CrossRef] [Green Version]
- Mertens, J.; Casado, S.; Mata, C.P.; Hernando-Perez, M.; de Pablo, P.J.; Carrascosa, J.L.; Caston, J.R. A protein with simultaneous capsid scaffolding and dsRNA-binding activities enhances the birnavirus capsid mechanical stability. Sci. Rep. 2015, 5, 13486. [Google Scholar] [CrossRef] [Green Version]
- Stricker, R.L.; Behrens, S.E.; Mundt, E. Nuclear factor NF45 interacts with viral proteins of infectious bursal disease virus and inhibits viral replication. J. Virol. 2010, 84, 10592–10605. [Google Scholar] [CrossRef] [Green Version]
- Andersen, G.R.; Pedersen, L.; Valente, L.; Chatterjee, I.; Kinzy, T.G.; Kjeldgaard, M.; Nyborg, J. Structural basis for nucleotide exchange and competition with tRNA in the yeast elongation factor complex eEF1A:eEF1Balpha. Mol. Cell 2000, 6, 1261–1266. [Google Scholar] [CrossRef]
- Liu, G.; Tang, J.; Edmonds, B.T.; Murray, J.; Levin, S.; Condeelis, J. F-actin sequesters elongation factor 1alpha from interaction with aminoacyl-tRNA in a pH-dependent reaction. J. Cell Biol. 1996, 135, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.; Crosby, S.R.; Burton, D.R.; Lilley, F.; Murphy, M.F. Actin bundling and polymerisation properties of eukaryotic elongation factor 1 alpha (eEF1A), histone H2A-H2B and lysozyme in vitro. J. Struct. Biol. 2011, 176, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Friis, M.B.; Rasmussen, T.B.; Belsham, G.J. Modulation of translation initiation efficiency in classical swine fever virus. J. Virol. 2012, 86, 8681–8692. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Zeng, X.; Yao, S.; Gao, L.; Zhang, L.; Qi, X.; Duan, Y.; Yang, B.; Gao, Y.; Liu, C.; et al. Voltage-Dependent Anion Channel 1 Interacts with Ribonucleoprotein Complexes To Enhance Infectious Bursal Disease Virus Polymerase Activity. J. Virol. 2017, 91, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, T.; Mathur, M.; Gupta, A.K.; Janssen, G.M.; Banerjee, A.K. RNA polymerase of vesicular stomatitis virus specifically associates with translation elongation factor-1 alphabetagamma for its activity. Proc. Natl. Acad. Sci. USA 1998, 95, 1449–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackwell, J.L.; Brinton, M.A. Translation elongation factor-1 alpha interacts with the 3’ stem-loop region of West Nile virus genomic RNA. J. Virol. 1997, 71, 6433–6444. [Google Scholar] [CrossRef] [Green Version]
- Pflug, A.; Lukarska, M.; Resa-Infante, P.; Reich, S.; Cusack, S. Structural insights into RNA synthesis by the influenza virus transcription-replication machine. Virus Res. 2017, 234, 103–117. [Google Scholar] [CrossRef]
- Ishikawa, M. Studies on the mechanism of tobacco mosaic virus RNA replication. J. Gen. Plant Pathol. 2017, 83, 410–413. [Google Scholar] [CrossRef]
- Luan, H.; Shine, M.B.; Cui, X.; Chen, X.; Ma, N.; Kachroo, P.; Zhi, H.; Kachroo, A. The Potyviral P3 Protein Targets Eukaryotic Elongation Factor 1A to Promote the Unfolded Protein Response and Viral Pathogenesis. Plant Physiol. 2016, 172, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Cimarelli, A.; Luban, J. Translation elongation factor 1-alpha interacts specifically with the human immunodeficiency virus type 1 Gag polyprotein. J. Virol. 1999, 73, 5388–5401. [Google Scholar] [CrossRef] [Green Version]
- Accola, M.A.; Huang, B.; Al Masri, A.; McNiven, M.A. The antiviral dynamin family member, MxA, tubulates lipids and localizes to the smooth endoplasmic reticulum. J. Biol. Chem. 2002, 277, 21829–21835. [Google Scholar] [CrossRef] [Green Version]
- Stranden, A.M.; Staeheli, P.; Pavlovic, J. Function of the mouse Mx1 protein is inhibited by overexpression of the PB2 protein of influenza virus. Virology 1993, 197, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Qu, L.J.; Yao, J.F.; Yang, N. Skewed allele frequencies of an Mx gene mutation with potential resistance to avian influenza virus in different chicken populations. Poult. Sci. 2006, 85, 1327–1329. [Google Scholar] [CrossRef] [PubMed]
- Balkissoon, D.; Staines, K.; McCauley, J.; Wood, J.; Young, J.; Kaufman, J.; Butter, C. Low frequency of the Mx allele for viral resistance predates recent intensive selection in domestic chickens. Immunogenetics 2007, 59, 687–691. [Google Scholar] [CrossRef] [PubMed]
- Rawle, D.J.; Li, D.; Swedberg, J.E.; Wang, L.; Soares, D.C.; Harrich, D. HIV-1 Uncoating and Reverse Transcription Require eEF1A Binding to Surface-Exposed Acidic Residues of the Reverse Transcriptase Thumb Domain. MBio 2018, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Duan, Y.; Han, C.; Yao, S.; Qi, X.; Gao, Y.; Maier, H.J.; Britton, P.; Chen, L.; Zhang, L.; et al. Infectious Bursal Disease Virus Subverts Autophagic Vacuoles To Promote Viral Maturation and Release. J. Virol. 2017, 91, 5. [Google Scholar] [CrossRef] [Green Version]
- Delgui, L.R.; Rodriguez, J.F.; Colombo, M.I. The endosomal pathway and the Golgi complex are involved in the infectious bursal disease virus life cycle. J. Virol. 2013, 87, 8993–9007. [Google Scholar] [CrossRef] [Green Version]
- Gimenez, M.C.; Zanetti, F.A.; Terebiznik, M.R.; Colombo, M.I.; Delgui, L.R. Infectious Bursal Disease Virus Hijacks Endosomal Membranes as the Scaffolding Structure for Viral Replication. J. Virol. 2018, 92, 11. [Google Scholar] [CrossRef] [Green Version]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Yan, N.; Liu, A.; Li, Y.; Chen, Z.; Gao, L.; Qi, X.; Gao, Y.; Liu, C.; Zhang, Y.; et al. Chicken eEF1α is a Critical Factor for the Polymerase Complex Activity of Very Virulent Infectious Bursal Disease Virus. Viruses 2020, 12, 249. https://doi.org/10.3390/v12020249
Yang B, Yan N, Liu A, Li Y, Chen Z, Gao L, Qi X, Gao Y, Liu C, Zhang Y, et al. Chicken eEF1α is a Critical Factor for the Polymerase Complex Activity of Very Virulent Infectious Bursal Disease Virus. Viruses. 2020; 12(2):249. https://doi.org/10.3390/v12020249
Chicago/Turabian StyleYang, Bo, Nana Yan, Aijing Liu, Yue Li, Zehua Chen, Li Gao, Xiaole Qi, Yulong Gao, Changjun Liu, Yanping Zhang, and et al. 2020. "Chicken eEF1α is a Critical Factor for the Polymerase Complex Activity of Very Virulent Infectious Bursal Disease Virus" Viruses 12, no. 2: 249. https://doi.org/10.3390/v12020249