Genomic, Morphological and Biological Traits of the Viruses Infecting Major Fruit Trees

 , and
, and 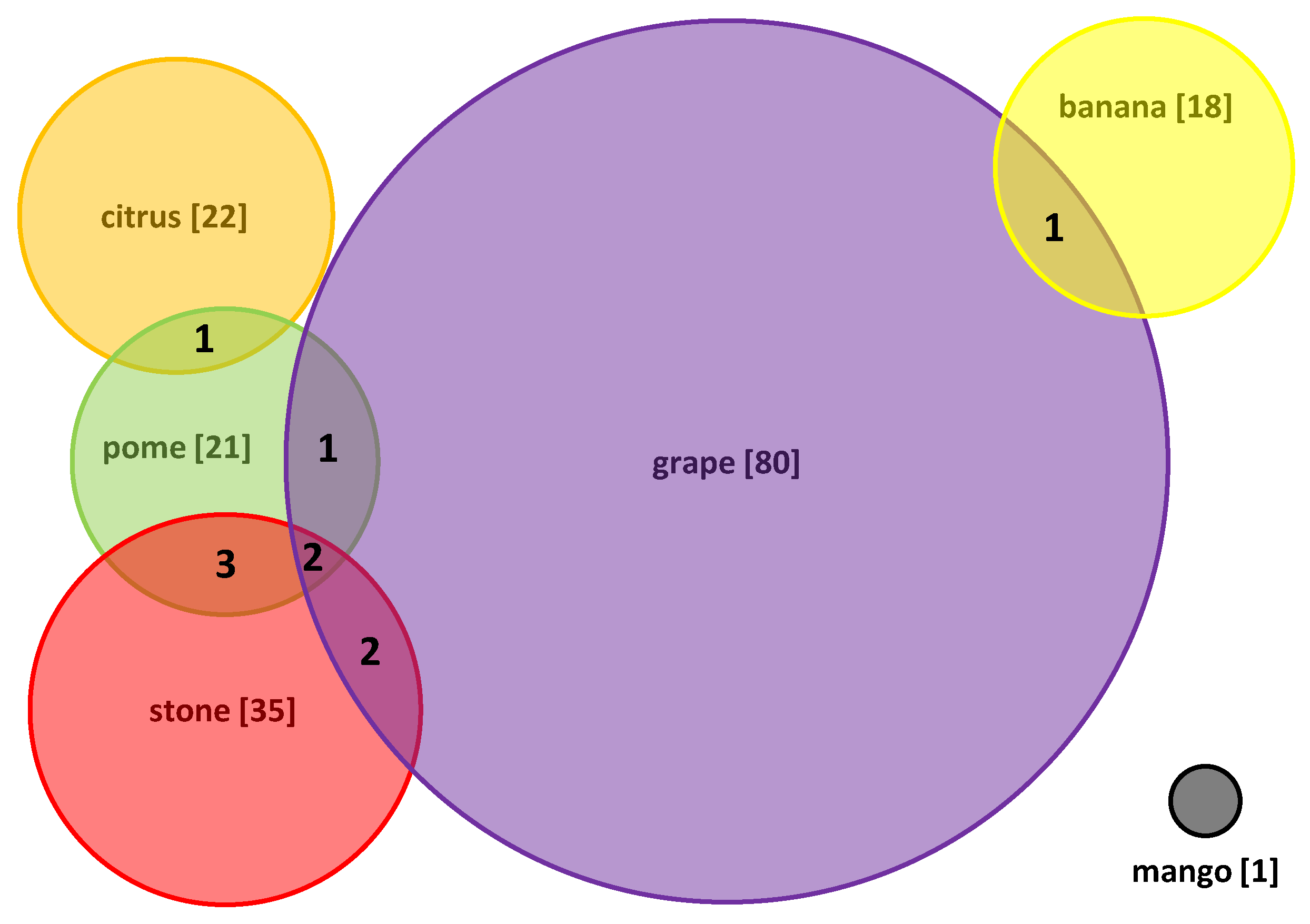{kind=link}
Abstract
:1. Introduction
2. Viruses Infecting Fruit Trees
2.1. Viruses Infecting Citrus
2.2. Viruses Infecting Pome Fruits
2.3. Viruses Infecting Stone Fruits
2.4. Viruses Infecting Grapevine
2.5. Viruses Infecting Mango
2.6. Viruses Infecting Banana
3. Summary
Supplementary Materials
Acknowledgments
Conflicts of Interest
References
- Janick, J. The origins of fruits, fruit growing, and fruit breeding. Plant Breeding Rev. 2005, 25, 255–320. [Google Scholar]
- Lee, R.F. Control of virus diseases of citrus. In Advances in Virus Research; Elsevier BV: London, UK, 2015; Volume 91, pp. 143–173. [Google Scholar]
- Barba, M.; Ilardi, V.; Pasquini, G. Control of Pome and Stone Fruit Virus Diseases; Elsevier BV: London, UK, 2015; Volume 91, pp. 47–83. [Google Scholar]
- Dawson, W.O. Citrus tristeza virus: Complex and tristeza disease. In Molecular Genetics of Citrus Tristeza Virus; Karasev, A.V., Hilf, M.E., Eds.; APS Press: St. Paul, MN, USA, 2010; pp. 53–72. [Google Scholar]
- Milne, R.G.; Garcia, M.L.; Grau, O. Genus Ophiovirus. In Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses; van Regenmortel, M.H.V., Fauquest, C.M., Bishop, D.H.L., Carstens, E.B., Estes, M.K., Lemon, S.M., Maniloff, J., Moyo, M.A., McGeoch, D.J., Pringle, C.R., et al., Eds.; Academic Press: San Diego, CA, USA, 2000. [Google Scholar]
- Loconsole, G.; Önelge, N.; Potere, O.; Giampetruzzi, A.; Bozan, O.; Satar, S.; De Stradis, A.; Savino, V.N.; Yokomi, R.K.; Saponari, M. Identification and characterization of citrus yellow vein clearing virus, a putative new member of the genus Mandarivirus. Phytopathology 2012, 102, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Choudhary, N.; Guillermo, L.M.; Shao, J.; Govindarajulu, A.; Achor, D.; Wei, G.; Picton, D.D.; Levy, L.; Nakhla, M.K.; et al. A novel virus of the genus Cilevirus causing symptoms similar to citrus leprosis. Phytopathology 2013, 103, 488–500. [Google Scholar] [CrossRef]
- Rodrigues, J.C.V.; Kitajima, E.; Childers, C.; Chagas, C. Citrus leprosis virus vectored by Brevipalpus phoenicis (Acari: Tenuipalpidae) on citrus in Brazil. Exp. Appl. Acarol. 2003, 30, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Asmae, A.; Essaїd, A.B.; Mohammed, I. Recent advances in citrus psorosis virus. Virusdisease 2014, 25, 261–276. [Google Scholar]
- Vives, M.C.; Velázquez, K.; Pina, J.A.; Moreno, P.; Guerri, J.; Navarro, L. Identification of a new enamovirus associated with citrus vein enation disease by deep sequencing of small RNAs. Phytopathology 2013, 103, 1077–1086. [Google Scholar] [CrossRef] [PubMed]
- Iwanami, T.; Kano, T.; Koizumi, M. Pathogenic diversity of citrus tatter leaf virus isolates. Ann. Phytopathol. Soc. Jpn. 1991, 45, 712–720. [Google Scholar]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Carstens, E.B.; Lefkowitz, E. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2014). Arch. Virol. 2014, 159, 2831–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, M.; Wu, Q.; Atta, S.; Su, H.; Yu, Y.; Chen, H.; Zhou, C.Y. First molecular evidence of citrus yellow vein clearing virus from citrus in Punjab, Pakistan. Plant Dis. 2016, 100, 540. [Google Scholar] [CrossRef]
- Hartung, J.S.; Huang, Q. Cloning and sequence analysis of an infectious clone of citrus yellow mosaic virus that can infect sweet orange via Agrobacterium-mediated inoculation. J. Virol. 2001, 82, 2549–2558. [Google Scholar]
- Karasev, A.V.; Han, S.S.; Iwanami, T. Satsuma dwarf and related viruses belong to a new lineage of plant picorna-like viruses. Virus Genes 2001, 23, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H. Mechanical transmission of viruses of Satsuma dwarf and Natsudaidai dwarf from citrus to citrus. Jpn. J. Phytopathol. 1972, 38, 156–160. [Google Scholar] [CrossRef]
- Bennani, B.; Mendes, C.; Zemzami, M.; Azeddoug, H.; Nolasco, G. Citrus variegation virus: molecular variability of a portion of the RNA 3 containing the coat protein gene and design of primers for RT-PCR detection. Eur. J. Plant Pathol. 2002, 108, 155–162. [Google Scholar] [CrossRef]
- Matsumura, E.E.; Coletta-Filho, H.D.; Nouri, S.; Falk, B.W.; Nerva, L.; Oliveira, T.S.; Dorta, S.O.; Machado, M.A.; White, K.A. Deep sequencing analysis of RNAs from citrus plants grown in a citrus sudden death-affected area reveals diverse known and putative novel viruses. Viruses 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Navarro, B.; Minutolo, M.; De Stradis, A.; Palmisano, F.; Alioto, D.; Di Serio, F. The first phlebo-like virus infecting plants: a case study on the adaptation of negative-stranded RNA viruses to new hosts. Mol. Plant Pathol. 2018, 19, 1075–1089. [Google Scholar] [CrossRef] [PubMed]
- Yaegashi, H.; Yoshikawa, N.; Candresse, T. Apple chlorotic leaf spot virus in pome fruits. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; pp. 17–22. [Google Scholar]
- Massart, S.; Jijakli, M.H.; Kummert, J. Apple stem grooving virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; pp. 85–90. [Google Scholar]
- Digiaro, M.; Savino, V.; Di Terlizzi, B. Ilarvirus in apricot and plum pollen. Acta Hortic. 1992, 309, 93–98. [Google Scholar] [CrossRef]
- Barba, M.; Pasquini, G.; Quacquarelli, A. Role of seeds in the epidemiology of two almond viruses. Acta Hortic. 1986, 193, 127–130. [Google Scholar] [CrossRef]
- Sweet, J.B. Fruit tree virus infections of woody exotic and indigenous plants in Britain. Acta Phytopathol. 1980, 15, 231–238. [Google Scholar] [CrossRef]
- Fernando, B.M.; da Silva, J.C.F.; Martins, F.T.V.; Batista, F.E.P.; Murilo, Z.F. A novel, highly divergent ssdna virus identified in Brazil infecting apple, pear and grapevine. Virus Res. 2015, 210, 27–33. [Google Scholar]
- Capote, N.; Gorris, M.T.; Martínez, M.C.; Asensio, M.; Olmos, A.; Cambra, M. Inference between D and M types of plum pox virus in Japanese plum assessed by specific monoclonal antibodies and quantitative real-time reverse transcription polymerase chain reaction. Phytopathology 2006, 96, 320–325. [Google Scholar] [CrossRef]
- Barba, M.; Hadidi, A.; Candresse, T.; Cambra, M. Plum pox virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; pp. 185–198. [Google Scholar]
- Garcia, J.A.; Glasa, M.; Cambra, M.; Candresse, T. Plum pox virus and sharka: a model potyvirus and a major disease. Mol. Plant Pathol. 2013, 15, 226–241. [Google Scholar] [CrossRef] [PubMed]
- James, D.; Varga, A.; Sanderson, D. Genetic diversity of plum pox virus: strains, disease and related challenges for control. Can. J. Plant Pathol. 2013, 35, 431–441. [Google Scholar] [CrossRef]
- Pasquini, G.; Barba, M. The question of seed transmissibility of plum pox virus. EPPO Bull. 2006, 36, 287–292. [Google Scholar] [CrossRef]
- Caglayan, K.; Ulubas Serce, C.; Gazel, M.; Varveri, C. Prune dwarf virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; pp. 199–206. [Google Scholar]
- Rampitsch, C.; Eastwell, K.C.; Hall, J. Setting confidence limits for the detection of prune dwarf virus in Prunus avium with a monoclonal antibody-based triple antibody-sandwich ELISA. Ann. Appl. Boil. 1995, 126, 485–491. [Google Scholar] [CrossRef]
- Uyemoto, J.K. Important diseases of prunus caused by viruses and other graft-transmissible pathogens in California and South Carolina. Plant Dis. 1992, 76, 5. [Google Scholar] [CrossRef]
- Nemeth, M. Virus, Mycoplasmas and Rickettsia Diseases of Fruit Trees; Akademiai Kiado: Budapest, Hungary, 1986. [Google Scholar]
- Posnette, A.F.; Ellenberger, C.E. The line-pattern virus disease of plums. Ann. Appl. Boil. 1957, 45, 74–80. [Google Scholar] [CrossRef]
- Jelkmann, W.; Rott, M.; Uyemoto, J.K. Cherry green ring mottle virus. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T., Jelkmann, W., Eds.; APS Press: St. Paul, MN, USA, 2011; pp. 115–118. [Google Scholar]
- Hadidi, A.; Barba, M. Economic Impact of Pome and Stone Fruit Viruses and Viroids; Scientific Societies: St. Paul, MN, USA, 2011; pp. 1–7. [Google Scholar]
- Mekouria, T.A.; Smith, T.J.; Beers, E.; Eastwell, K.C. Little cherry virus 2 is transmitted to sweet cherry by Pseudococcus maritimus (ehrhorn), a new vector of this virus. Plant Dis. 2013, 97, 851. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cai, L.; Zhou, L.; Yang, Z.; Hong, N.; Wang, G.; Li, S.; Xu, W. Deep sequencing reveals the first fabavirus infecting peach. Sci. Rep. 2017, 7, 11329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent advances on detection and characterization of fruit tree viruses using high-throughput sequencing technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. Directory of virus and virus-like diseases of the grapevine and their agents. J. Plant Pathol. 2014, 96, 1–136. [Google Scholar]
- Martelli, G.P.; Boudon-Padieu, E. Directory of infectious diseases of grapevines. In Options Méditerranéennes Série b; Martelli, G.P., Boudon-Padieu, E., Eds.; CIHEAM-IAMB: Bari, Italy, 2006; Volume 55. [Google Scholar]
- Martelli, G.P.; Abou Ghanem-Sabanadzovic, N.; Agranovsky, A.A.; Al Rwahnih, M.; Dolja, V.V.; Dovas, C.I.; Fuchs, M.; Gugerli, P.; Hu, J.S.; Jelkmann, W.; et al. Taxonomic revision of the family Closteroviridae with special reference to the grapevine leafroll-associated members of the genus Ampelovirus and the putative species unassigned to the family. J. Plant Pathol. 2012, 94, 7–19. [Google Scholar]
- Herrbach, E.; Alliaume, A.; Prator, C.A.; Daane, K.M.; Cooper, M.L.; Almeida, R.P.P. Vector transmission of grapevine leafroll-associated viruses. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer Nature: Cham, Switzerland, 2017; pp. 483–503. [Google Scholar]
- Martelli, G.P. An overview on grapevine viruses, viroids, and the diseases they cause. In Grapevine Viruses: Molecular Biology, Diagnostics and Management; Springer Nature: Cham, Switzerland, 2017; pp. 31–46. [Google Scholar]
- Lima, M.F.; Alkowni, R.; Uyemoto, J.K.; Golino, D.; Osman, F.; Rowhani, A. Molecular analysis of a California strain of Rupestris stem pitting-associated virus isolated from declining Syrah grapevines. Arch. Virol. 2006, 151, 1889–1894. [Google Scholar] [CrossRef] [PubMed]
- Maliogka, V.I.; Martelli, G.P.; Fuchs, M.; Katis, N.I. Control of viruses infecting grapevine. In Advances in Virus Research; Elsevier BV: London, UK, 2015; Volume 91, pp. 175–227. [Google Scholar]
- Andret-Link, P.; Laporte, C.; Valat, L.; Ritzenthaler, C.; Demangeat, G.; Vigne, E. Grapevine fanleaf virus: still a major threat to the grapevine industry. J. Plant Pathol. 2004, 86, 183–195. [Google Scholar]
- Raski, D.J.; Goheen, A.C.; Lider, L.A.; Meredith, C.P. Strategies against grape vine fanleaf virus and its nematode vector. Plant Dis. 1983, 67, 335–339. [Google Scholar] [CrossRef]
- Sela, N.; Luria, N.; Yaari, M.; Prusky, D.; Dombrovsky, A. Genome sequence of a potential new benyvirus isolated from mango RNA-seq data. Genome Announc. 2016, 4, e01250-16. [Google Scholar] [CrossRef]
- Kumar, P.L.; Selvarajan, R.; Iskra-Caruana, M.-L.; Chabannes, M.; Hanna, R. Biology, etiology, and control of virus diseases of banana and plantain. In Advances in Virus Research; Elsevier BV: London, UK, 2015; Volume 91, pp. 229–269. [Google Scholar]
- Nelson, S.C. Banana Bunchy Top: Detailed Signs and Symptom. Available online: https://www.ctahr.hawaii.edu/bbtd/downloads/BBTV-details.pdf (accessed on 1 April 2019).
- Thangavelu, R.; Selvarajan, R.; Singh, H.P. Status of banana streak virus and banana bract mosaic virus diseases in India. In Banana: Improvement, Production and Utilization; AIPUB NRCB: Trichy, India, 2000; pp. 364–376. [Google Scholar]
- Dahal, G.; Ortiz, R.; Tenkouano, A.; Hughes, J.A.; Thottappilly, G.; Vuylsteke, D. Relationship between natural occurrence of banana streak badnavirus and symptom expression, relative concentration of viral antigen. Plant Pathol. 2000, 49, 68–79. [Google Scholar] [CrossRef]
- Kubiriba, J.; Legg, J.P.; Tushemereirwe, W.; Adipala, E. Vector transmission of banana streak virus in the screenhouse in Uganda. Ann. Appl. Biol. 2001, 139, 37–43. [Google Scholar] [CrossRef]
- Selvarajan, R.; Jeyabaskaran, K.J. Effect of banana bract mosaic virus (BbrMV) on growth and yield of cultivar Nendran (Alantain, AAB). Indian Phytopathol. 2006, 59, 496–500. [Google Scholar]
- Szittya, G.; Silhavy, D.; Molnár, A.; Havelda, Z.; Lovas, A.; Lakatos, L.; Bánfalvi, Z.; Burgyán, J. Low temperature inhibits RNA silencing-mediated defence by the control of siRNA generation. EMBO J. 2003, 22, 633–640. [Google Scholar] [CrossRef] [Green Version]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umer, M.; Liu, J.; You, H.; Xu, C.; Dong, K.; Luo, N.; Kong, L.; Li, X.; Hong, N.; Wang, G.; et al. Genomic, Morphological and Biological Traits of the Viruses Infecting Major Fruit Trees. Viruses 2019, 11, 515. https://doi.org/10.3390/v11060515
Umer M, Liu J, You H, Xu C, Dong K, Luo N, Kong L, Li X, Hong N, Wang G, et al. Genomic, Morphological and Biological Traits of the Viruses Infecting Major Fruit Trees. Viruses. 2019; 11(6):515. https://doi.org/10.3390/v11060515
Chicago/Turabian StyleUmer, Muhammad, Jiwen Liu, Huafeng You, Chuan Xu, Kaili Dong, Ni Luo, Linghong Kong, Xuepei Li, Ni Hong, Guoping Wang, and et al. 2019. "Genomic, Morphological and Biological Traits of the Viruses Infecting Major Fruit Trees" Viruses 11, no. 6: 515. https://doi.org/10.3390/v11060515