Co-Expression of Chicken IL-2 and IL-7 Enhances the Immunogenicity and Protective Efficacy of a VP2-Expressing DNA Vaccine against IBDV in Chickens

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids, Cells, Viruses, and Chickens
2.2. Vaccine, Antibodies, and Proteins
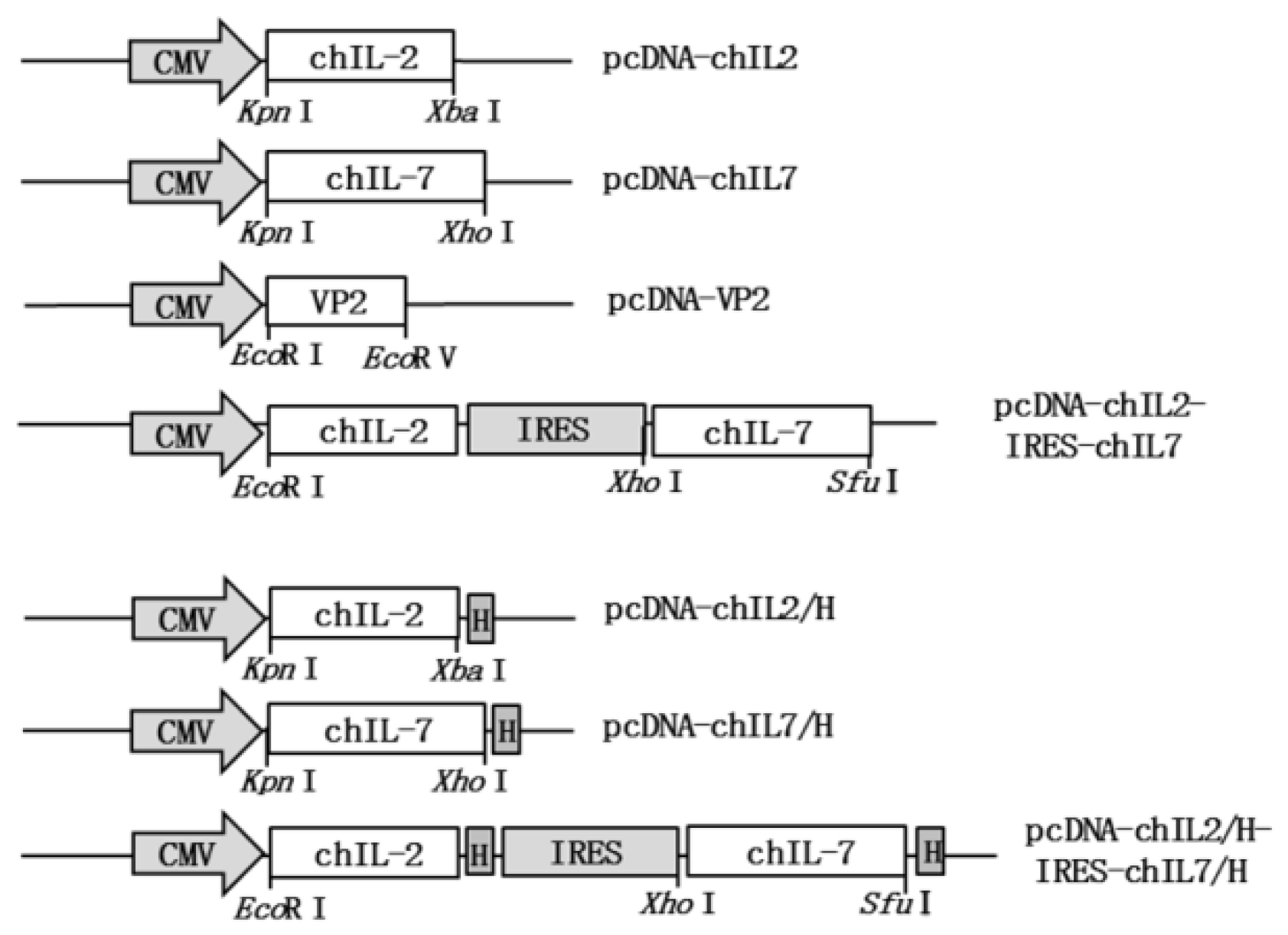
2.3. Construction of chIL-2 Expression Vector and chIL-2/chIL-7 Bicistronic Expression Vectors
2.4. Expression in HEK293T Cells
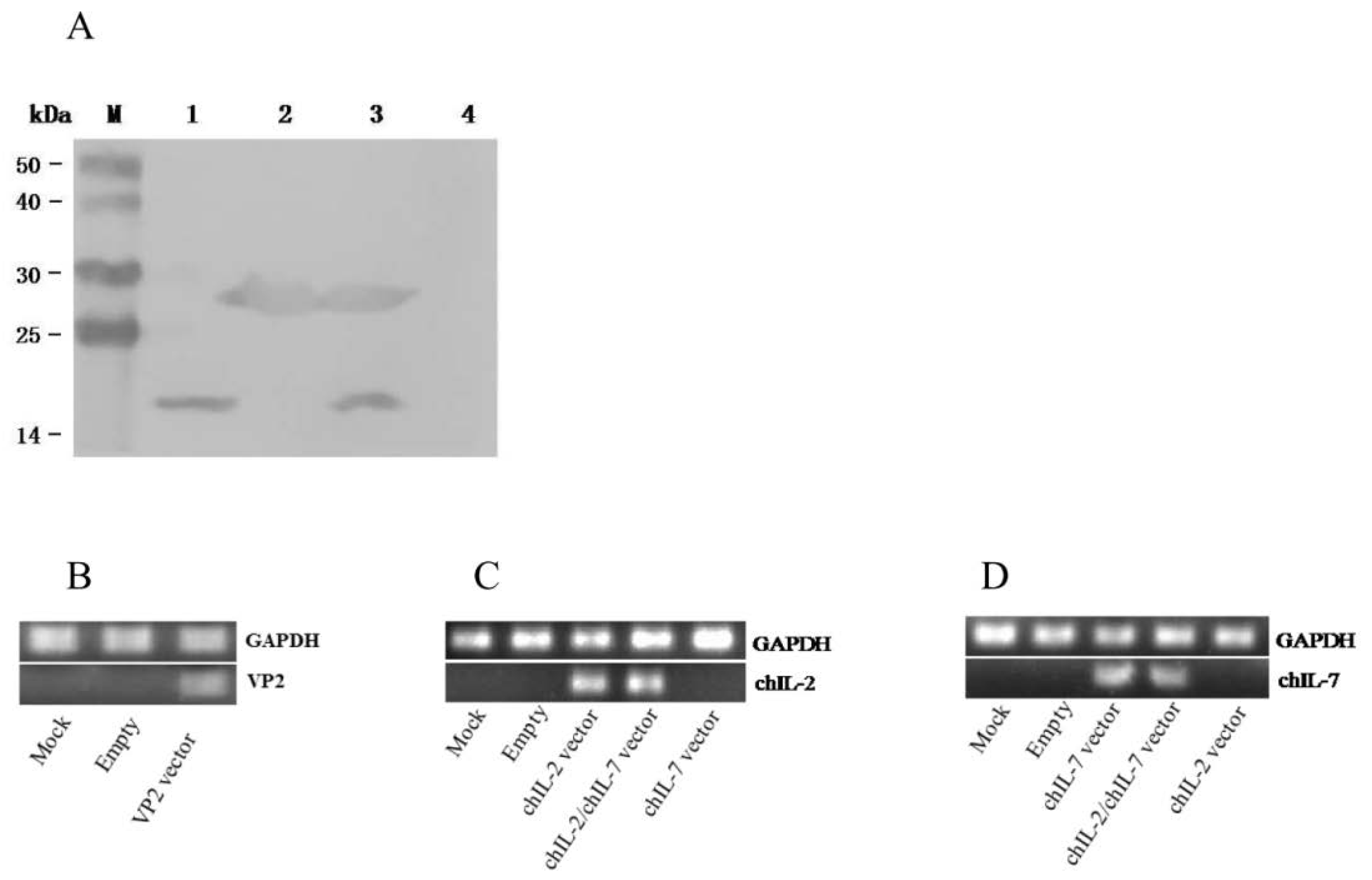
2.5. Western Blot
2.6. Detections of VP2, chIL-2 and chIL-7 Expressions In Vivo by RT-PCR
2.7. IBDV Propagation and Titer Determination
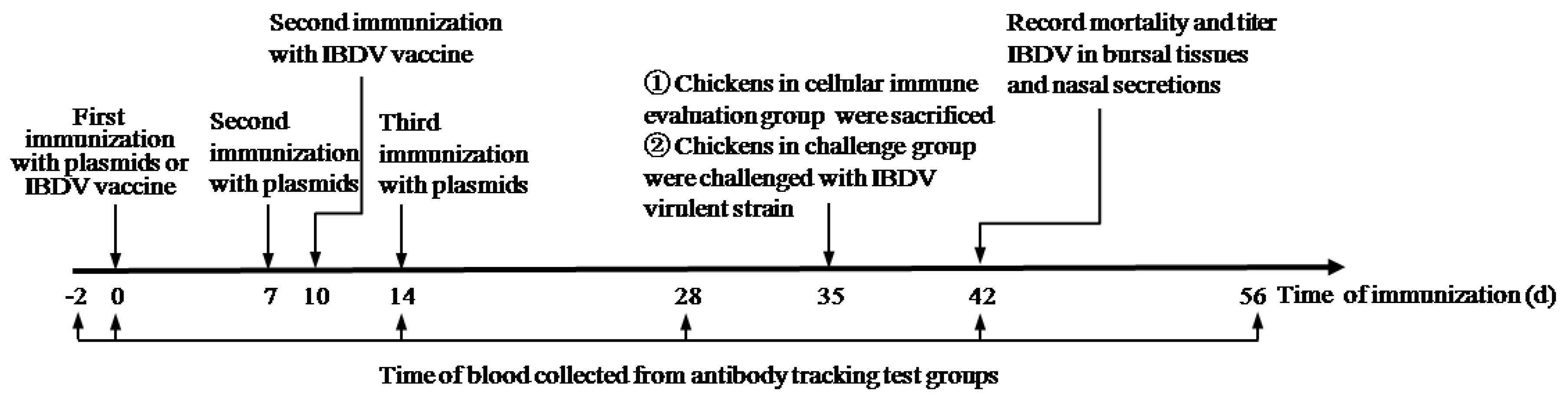
2.8. Animal Experiment
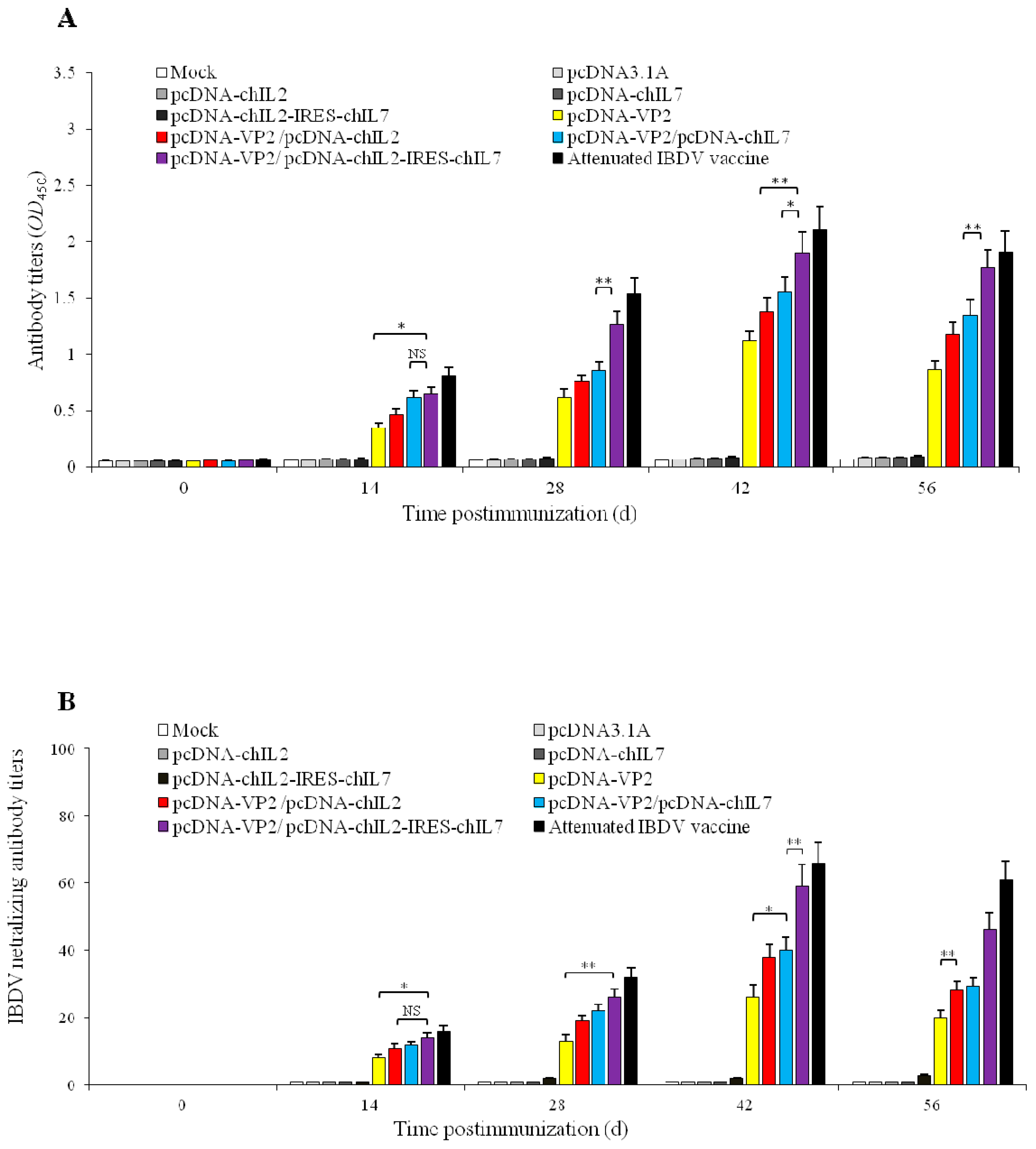
2.9. Detection of Serum Antibody Titers
2.10. Detection of Serum Neutralization Titers
2.11. Lymphocyte Proliferation Assay
2.12. Detection of Cytokine Production
2.13. Viral Challenge Study
2.14. Statistics
3. Results
3.1. Construction of chIL-2 and chIL-7 Expression Vectors
3.2. In Vitro and In Vivo Expression of chIL-2 and chIL-7
3.3. ChckenIL-2 and chIL-7 Genes Mutually Enhance Humoral Immune Response to IBDV VP2 DNA Vaccine in Immunized Chickens
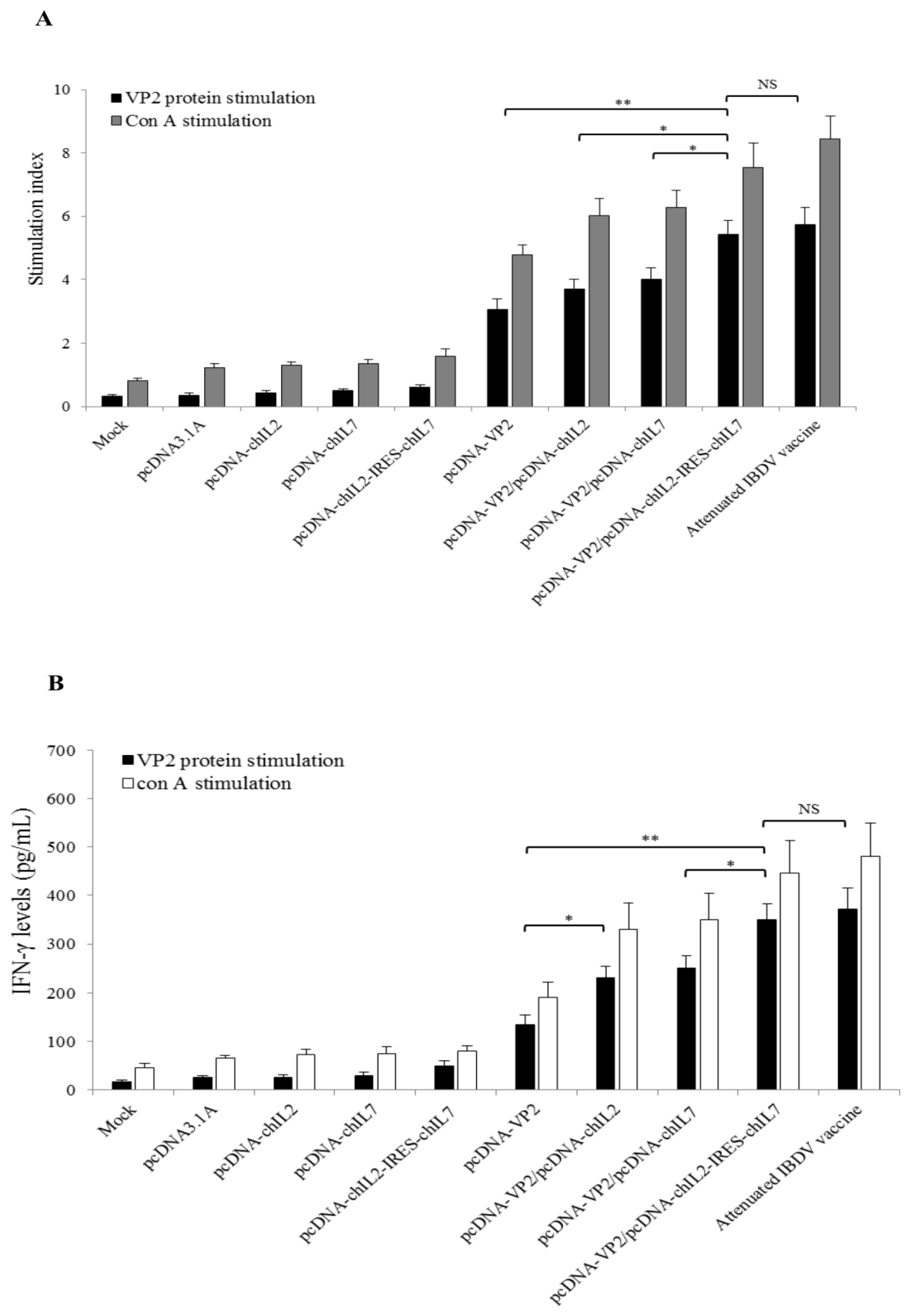
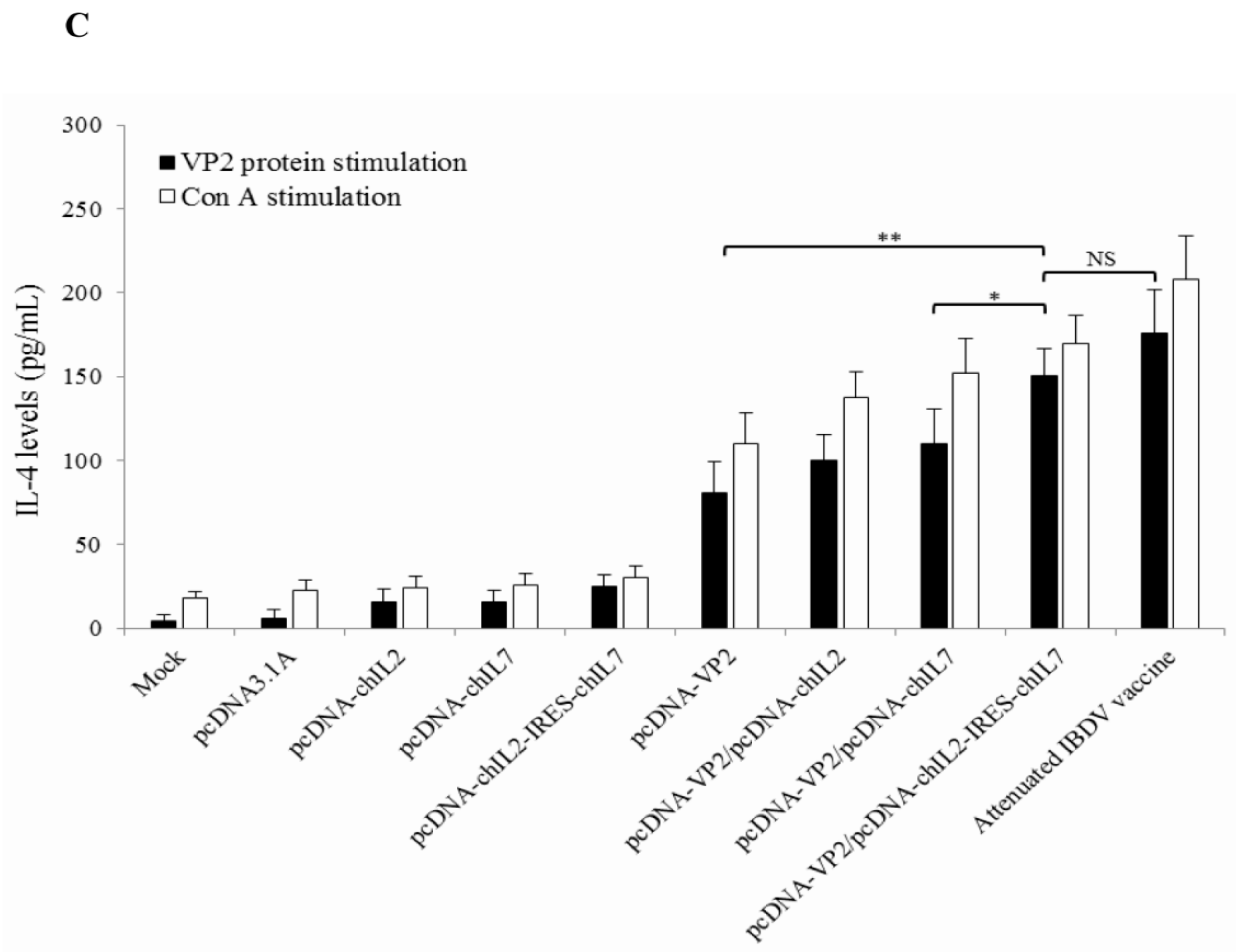
3.4. Chicken IL-2 and chIL-7 Genes Mutually Enhance VP2 DNA Vaccine-Induced Cellular Immune Response in Immunized Chickens
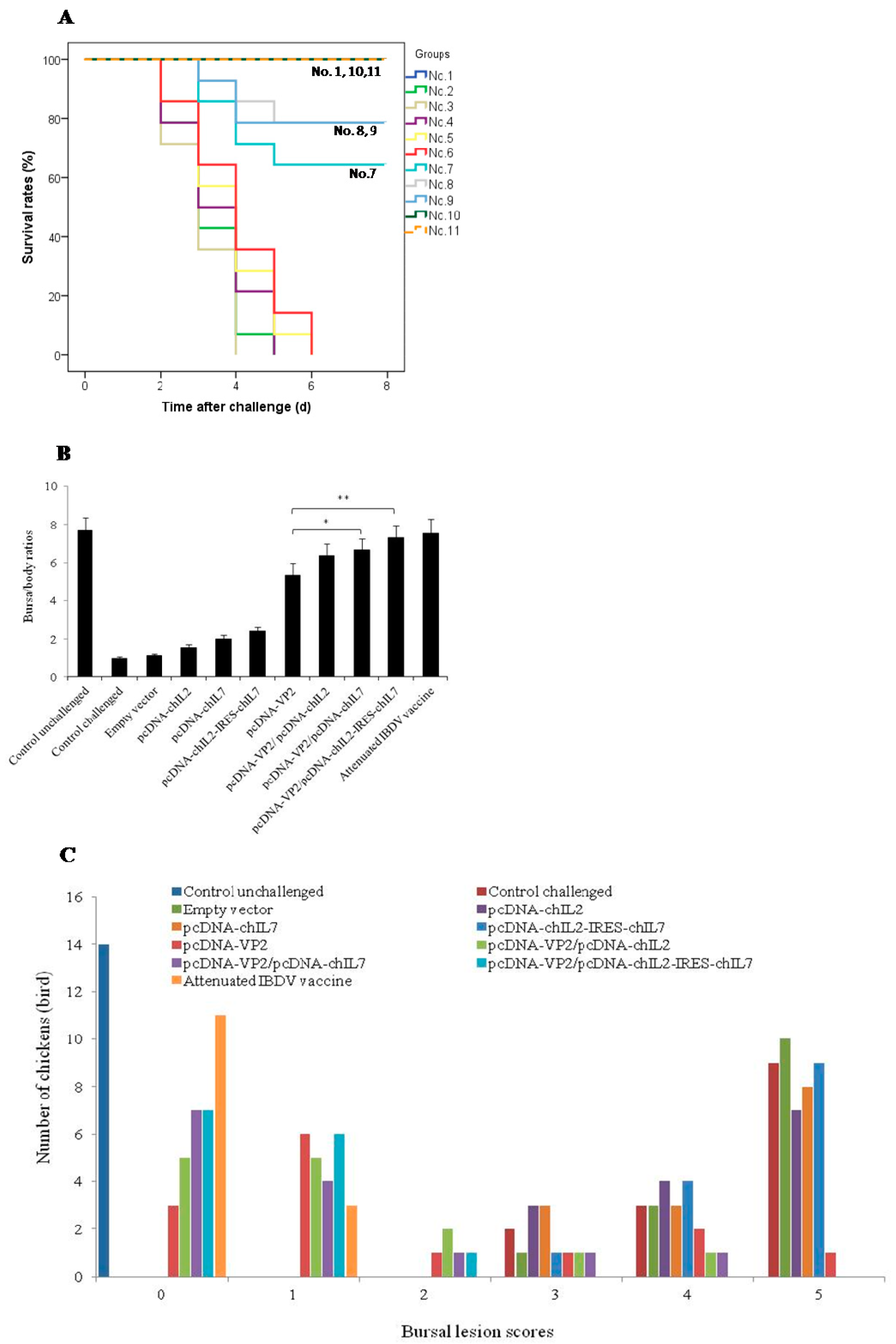
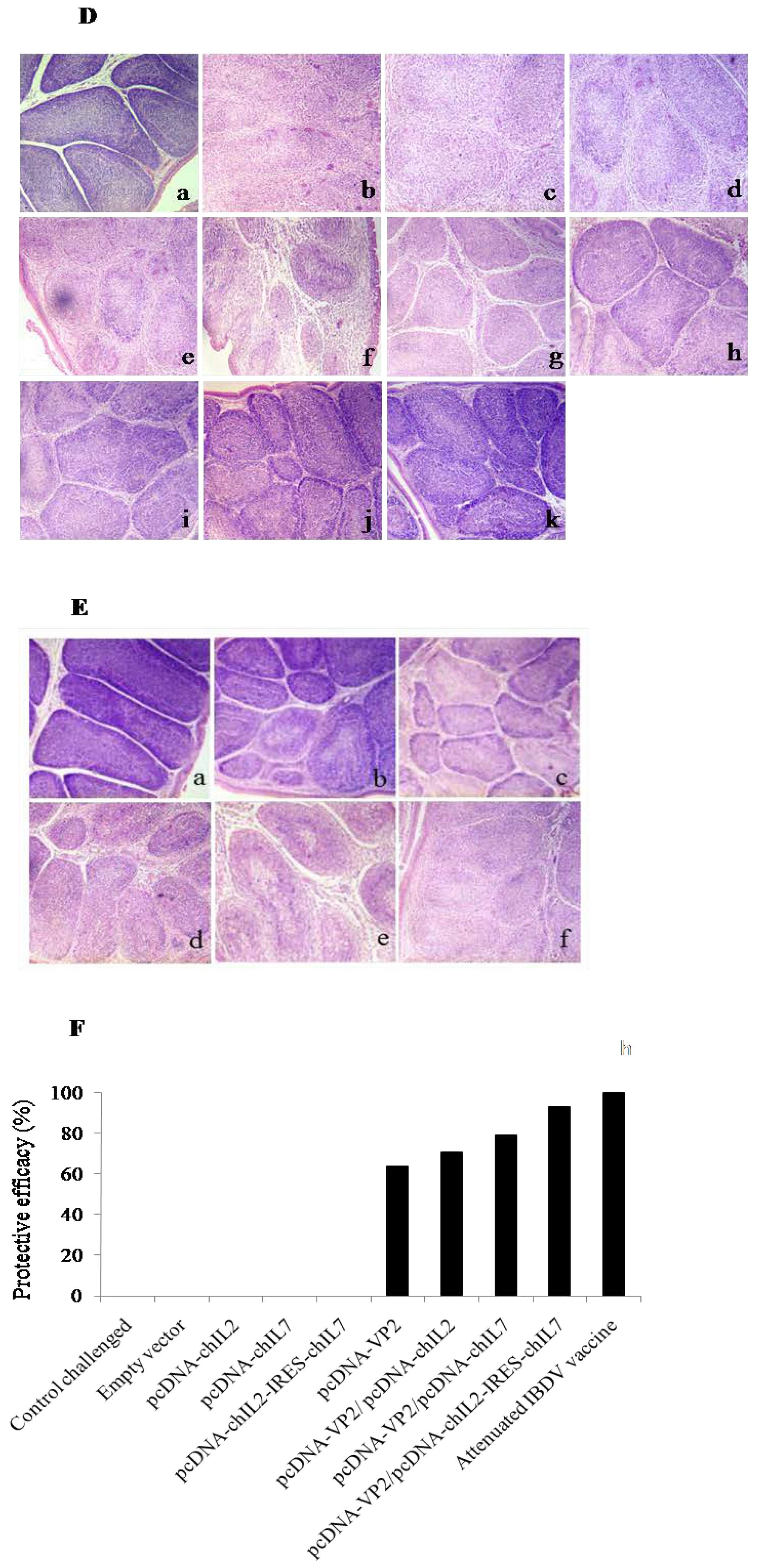
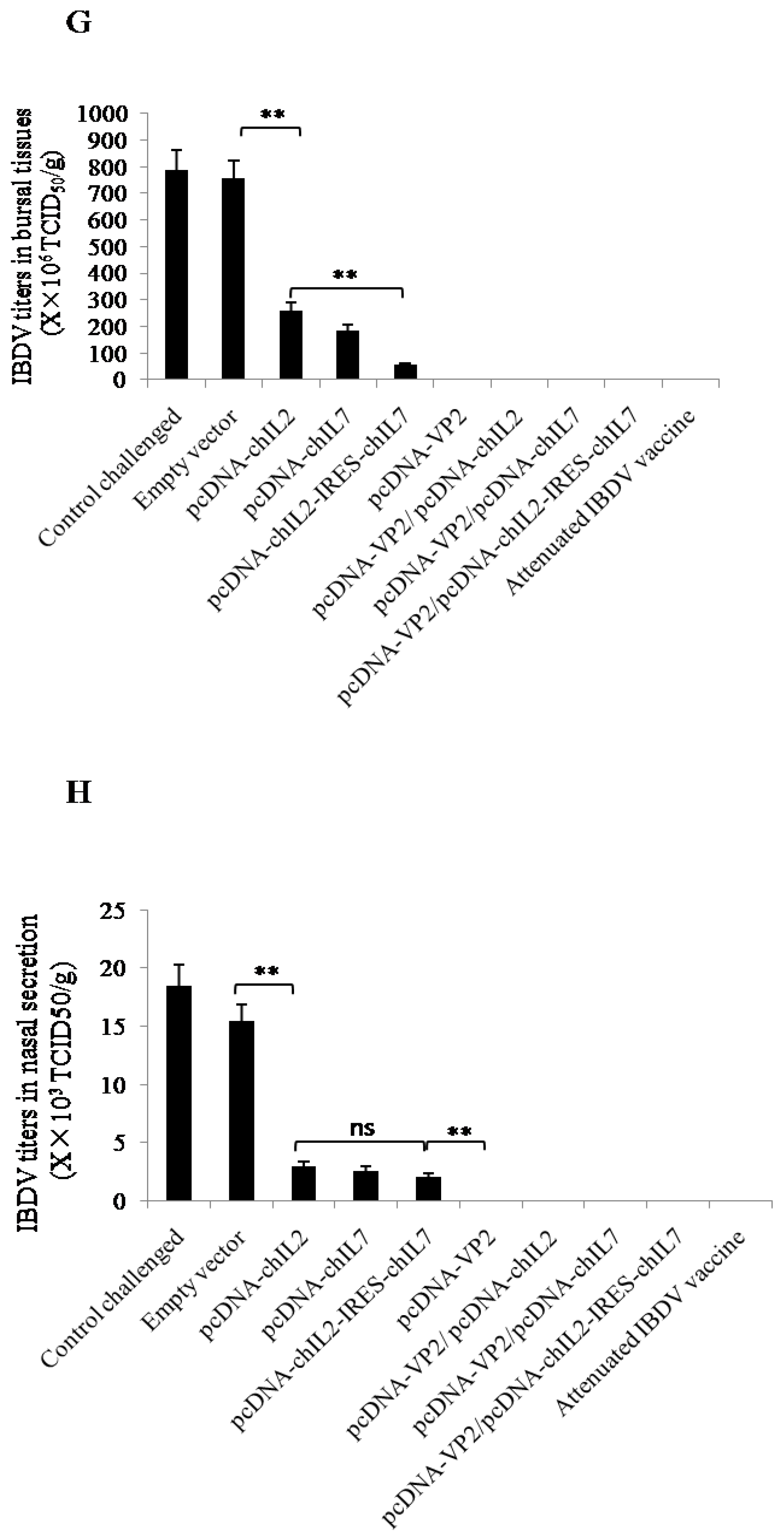
3.5. Chicken IL-2 and chIL-7 Genes Mutually Improve the Protective Efficacy of VP2 DNA Vacccine-Immunized Chickens against Virulent IBDV Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, J.M.; Kim, I.J.; Rautenschlein, S.; Yeh, H.Y. Infectious bursal disease virus of chickens: pathogenesis and immunosuppression. Dev. Comp. Immunol. 2000, 24, 223–235. [Google Scholar] [CrossRef]
- Brown, M.D.; Skinner, M.A. Coding sequences of both genome segments of a European ‘very virulent’ infectious bursal disease virus. Virus. Res. 1996, 40, 1–15. [Google Scholar] [CrossRef]
- Lombardo, E.; Maraver, A.; Espinosa, I.; Fernandez-Aris, A.; Rodriguez, J.F. VP5, the nonstructural polypeptide of infectious bursal disease virus, accumulates within the host plasma membrane and induces cell lysis. Virol. 2000, 277, 345–357. [Google Scholar] [CrossRef]
- Müller, H.; Becht, H. Biosynthesis of virus specific proteins in cells infected with infectious bursal disease virus and their significance as structural elements for infectious and incomplete particles. J. Virol. 1982, 44, 384–392. [Google Scholar] [PubMed]
- Balamurugan, V.; Kataria, J.M. Economically important non-oncogenic immunosuppressive viral diseases of chicken-current status. Vet. Res. Commun. 2006, 30, 541–566. [Google Scholar] [CrossRef] [PubMed]
- Azad, A.A.; McKern, N.M.; Macreadie, I.G.; Failla, P.; Heine, H.G.; Chapman, A.; Ward, C.W.; Fahey, K.J. Physicochemical and immunological characterization of recombinant host-protective antigen (VP2) of infectious bursal disease virus. Vaccine 1991, 9, 715–722. [Google Scholar] [CrossRef]
- Spies, U.; Müller, H.; Becht, H. Properties of RNA polymerase activity associated with infectious bursal disease virus and characterization of its reaction products. Virus. Res. 1987, 8, 127–140. [Google Scholar] [CrossRef]
- Mardassi, H.; Khabouchi, N.; Ghram, A.; Namouchi, A.; Karboul, A. A very virulent genotype of infectious bursal disease virus predominantly associated with recurrent infectious bursal disease outbreaks in Tunisian vaccinated flocks. Avian Dis. 2004, 48, 829–840. [Google Scholar] [CrossRef]
- Snyder, D.B. Changes in the field status of infectious bursal disease virus. Avian Pathol. 1990, 19, 419–423. [Google Scholar] [CrossRef] [Green Version]
- Fussell, L.W. Poultry industry strategies for control of immunosuppressive diseases. Poult. Sci. 1998, 77, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Li, K.; Gao, L.; Qi, X.; Gao, Y.; Qin, L.; Wang, Y.; Wang, X. DNA prime-protein boost vaccination enhances protective immunity against infectious bursal disease virus in chickens. Vet. Microbiol. 2013, 64, 9–17. [Google Scholar] [CrossRef]
- Pradhan, S.N.; Prince, P.R.; Madhumathi, J.; Arunkumar, C.; Roy, P.; Narayanan, R.B.; Antony, U. DNA vaccination with VP2 gene fragment confers protection against infectious bursal disease virus in chickens. Vet. Microbiol. 2014, 171, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Hulse, D.J.; Romero, C.H. Partial protection against infectious bursal disease virus through DNA-mediated vaccination with the VP2 capsid protein and chicken IL-2 genes. Vaccine 2004, 22, 1249–1259. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.H.; Yan, Y.X.; Jiang, J.; Lu, P. DNA immunization against very virulent infectious bursal disease virus with VP2-4-3 gene and chicken IL-6 gene. J. Vet. Med. B Infect. Dis. Vet. Public Health. 2005, 52, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Gao, H.; Gao, L.; Qi, X.; Gao, Y.; Qin, L.; Wang, Y.; Wang, X. Adjuvant effects of interleukin-18 in DNA vaccination against infectious bursal disease virus in chickens. Vaccine 2013, 31, 1799–1805. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Sung, H.W.; Yoon, B.I.; Kwon, H.M. Protection of chicken against very virulent IBDV provided by in ovo priming with DNA vaccine and boosting with killed vaccine and the adjuvant effects of plasmid-encoded chicken interleukin-2 and interferon-gamma. J. Vet. Sci. 2009, 10, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Huo, S.; Zuo, Y.; Li, N.; Li, X.; Zhang, Y.; Wang, L.; Liu, H.; Zhang, J.; Cui, D.; He, P.; et al. Chicken IL-7 as a potent adjuvant enhances IBDV VP2 DNA vaccine immunogenicity and protective efficacy. Vet. Microbiol. 2016, 193, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Zhang, J.; Zuo, Y.; Huo, S.; Zhang, Y.; Wang, L.; Li, X.; Zhong, F. Recombinant chicken interleukin-7 as a potent adjuvant increases the immunogenicity and protection of inactivated infectious bursal disease vaccine. Vet. Res. 2018, 49, 10. [Google Scholar] [CrossRef] [Green Version]
- Rochman, Y.; Spolski, R.; Leonard, W.J. New insights into the regulation of T cells by gamma (c) family cytokines. Nat. Rev. Immunol. 2009, 9, 480–490. [Google Scholar] [CrossRef]
- Guimond, M.; Veenstra, R.G.; Grindler, D.J.; Zhang, H.; Cui, Y.; Murphy, R.D.; Kim, S.Y.; Na, R.; Hennighausen, L.; Kurtulus, S.; et al. Interleukin 7 signaling in dendritic cells regulates the homeostatic proliferation and niche size of CD4+ T cells. Nat. Immunol. 2009, 10, 149–157. [Google Scholar] [CrossRef]
- Boyman, O.; Sprent, J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat. Rev. Immunol. 2012, 12, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Erard, F.; Corthesy, P.; Nabholz, M.; Lowenthal, J.W.; Zaech, P.; Plaetinck, G.; MacDonald, H.R. Interleukin 2 is both necessary and sufficient for the growth and differentiation of lectin-stimulated cytolytic T lymphocyte precursors. J. Immunol. 1985, 134, 1644–1652. [Google Scholar] [PubMed]
- Namen, A.E.; Schmierer, A.E.; March, C.J.; Overell, R.W.; Park, L.S.; Urdal, D.L.; Mochizuki, D.Y. B cell precursor growth-promoting activity. Purification and characterization of a growth factor active on lymphocyte precursors. J. Exp. Med. 1998, 167, 988–1002. [Google Scholar] [CrossRef] [PubMed]
- Fry, T.J.; Mackall, C.L. Interleukin-7: master regulator of peripheral T-cell homeostasis? Trends Immunol. 2001, 22, 564–571. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, S.; Li, X.; Wang, L.; Zhang, J.; Xu, J.; Huo, S.; Cao, X.; Zhong, Z.; Zhong, F. Mutual enhancement of IL-2 and IL-7 on DNA vaccine immunogenicity mainly involves regulations on their receptor expression and receptor-expressing lymphocyte generation. Vaccine 2015, 33, 3480–3487. [Google Scholar] [CrossRef]
- Yu, X.; Rui, L.; Shao, Q.; Liu, H.; Lu, Y.; Zhang, Y.; Li, Z. Changes of CD4+CD25+ cells ratio in immune organs from chickens challenged with infectious bursal disease virus strains with varying virulences. Viruses 2015, 7, 1357–1372. [Google Scholar] [CrossRef]
- Kibenge, F.S.; Qian, B.; Cleghorn, J.R.; Martin, C.K. Infectious bursal disease virus polyprotein processing does not involve cellular proteases. Arch. Virol. 1997, 142, 2401–2419. [Google Scholar] [CrossRef]
- Hsieh, M.K.; Wu, C.C.; Lin, T.L. The effect of co-administration of DNA carrying chicken interferon-gamma gene on protection of chickens against infectious bursal disease by DNA-mediated vaccination. Vaccine 2006, 24, 6955–6965. [Google Scholar] [CrossRef]
- Rodriguez-Chavez, I.R.; Rosenberger, J.K.; Cloud, S.S. Characterization of the antigenic, immunogenic, and pathogenic variation of infectious bursal disease virus due to propagation in different host systems (bursa, embryo, and cell culture). I. Antigenicity and immunogenicity. Avian Pathol. 2002, 31, 463–471. [Google Scholar] [CrossRef]
- Kumar, S.; Ahi, Y.S.; Salunkhe, S.S.; Koul, M.; Tiwari, A.K.; Gupta, P.K.; Rai, A. Effective protection by high efficiency bicistronic DNA vaccine against infectious bursal disease virus expressing VP2 protein and chicken IL-2. Vaccine 2009, 27, 864–869. [Google Scholar] [CrossRef]
- Maity, H.K.; Dey, S.; Mohan, C.M.; Khulape, S.A.; Pathak, D.C.; Vakharia, V.N. Protective efficacy of a DNA vaccine construct encoding the VP2 gene of infectious bursal disease and a truncated HSP70 of Mycobacterium tuberculosis in chickens. Vaccine 2015, 33, 1033–1039. [Google Scholar] [CrossRef]
- Mahmood, M.S.; Siddique, M.; Hussain, I.; Khan, A.; Mansoor, M.K. Protection capability of recombinant plasmid DNA vaccine containing VP2 gene of very virulent infectious bursal disease virus in chickens adjuvanted with CpG oligodeoxynucleotide. Vaccine 2006, 24, 4838–4846. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Yang, X.M.; Xie, Q.M.; Ma, J.Y.; Luo, Y.N.; Cao, Y.C.; Chen, F.; Bi, Y.Z. The potent adjuvant effects of chicken beta-defensin-1 when genetically fused with infectious bursal disease virus VP2 gene. Vet. Immunol. Immunopathol. 2010, 136, 92–97. [Google Scholar] [CrossRef]
- Stepaniak, J.A.; Shuster, J.E.; Hu, W.; Sundick, R.S. Production and in vitro characterization of recombinant chicken interleukin-2. J. Interferon. Cytokine Res. 1999, 19, 515–526. [Google Scholar]
- Leone, A.; Picker, L.J.; Sodora, D.L. IL-2, IL-7 and IL-15 as immune-modulators during SIV/HIV vaccination and treatment. Curr. HIV Res. 2009, 7, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Jie, Z.; Liang, Y.; Desai, M.; Soong, L.; Sun, J. Type 1 interferon-induced IL-7 maintains CD8+ T-cell responses and homeostasis by suppressing PD-1 expression in viral hepatitis. Cell. Mol. Immunol. 2015, 12, 213–221. [Google Scholar] [CrossRef]
- Park, S.H.; Song, M.Y.; Nam, H.J.; Im, S.J.; Sung, Y.C. Codelivery of IL-7 Augments Multigenic HCV DNA Vaccine-induced Antibody as well as broad T cell responses in cynomolgus monkeys. Immune Netw. 2010, 10, 198–205. [Google Scholar] [CrossRef]
- Chen, J.; Li, Z.Y.; Petersen, E.; Liu, W.G.; Zhu, X.Q. Co-administration of interleukins 7 and 15 with DNA vaccine improves protective immunity against Toxoplasma gondii. Exp. Parasitol. 2016, 162, 18–23. [Google Scholar] [CrossRef]
- Sun, Y.; Zhong, F.; Li, X.; Wang, X.; Wang, L.; Jia, Q.; Han, D.; Li, Z.; Zhang, F.; Pan, H. Immune enhancing effects of canine interleukin-7 gene on canine parvovirus DNA vaccine. Sci. Agric. Sin. 2012, 45, 2058–2066. [Google Scholar]
- Chen, H.; Zhong, F.; Li, X.; Wang, L.; Sun, Y.; Neng, C.; Zhang, K.; Li, W.; Wen, J. Effects of canine IL-2 and IL-7 genes on enhancing immunogenicity of canine parvovirus VP2 gene vaccine in mice. Wei Sheng Wu Xue Bao 2012, 52, 1392–1399. [Google Scholar] [PubMed]
- Huo, S.; Wang, L.; Zhang, Y.; Zhang, J.; Zuo, Y.; Xu, J.; Cui, D.; Li, X.; Zhong, F. Molecular cloning of chicken IL-7 and characterization of its antiviral activity against IBDV in vivo. Poult. Sci. 2016, 95, 2647–2654. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Number of Chicken | Inoculum | Dose (µg) |
|---|---|---|---|
| 1 | 30 | Mock | 0 |
| 2 | 30 | pcDNA3.1A | 200 |
| 3 | 30 | pcDNA-chIL2 | 200 |
| 4 | 30 | pcDNA-chIL7 | 200 |
| 5 | 30 | pcDNA-chIL2-IRES-chIL7 | 200 |
| 6 | 30 | pcDNA-VP2 | 200 |
| 7 | 30 | pcDNA-VP2 + pcDNA-chIL2 | 200 + 200 |
| 8 | 30 | pcDNA-VP2 + pcDNA-chIL7 | 200 + 200 |
| 9 | 30 | pcDNA-VP2+ pcDNA-chIL2-IRES-chIL7 | 200 + 100 |
| 10 | 30 | Attenuated IBDV vaccine (Ringpu) | 2 × 103 TCID50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, S.; Zhang, J.; Fan, J.; Wang, X.; Wu, F.; Zuo, Y.; Zhong, F. Co-Expression of Chicken IL-2 and IL-7 Enhances the Immunogenicity and Protective Efficacy of a VP2-Expressing DNA Vaccine against IBDV in Chickens. Viruses 2019, 11, 476. https://doi.org/10.3390/v11050476
Huo S, Zhang J, Fan J, Wang X, Wu F, Zuo Y, Zhong F. Co-Expression of Chicken IL-2 and IL-7 Enhances the Immunogenicity and Protective Efficacy of a VP2-Expressing DNA Vaccine against IBDV in Chickens. Viruses. 2019; 11(5):476. https://doi.org/10.3390/v11050476
Chicago/Turabian StyleHuo, Shanshan, Jianlou Zhang, Jinghui Fan, Xing Wang, Fengyang Wu, Yuzhu Zuo, and Fei Zhong. 2019. "Co-Expression of Chicken IL-2 and IL-7 Enhances the Immunogenicity and Protective Efficacy of a VP2-Expressing DNA Vaccine against IBDV in Chickens" Viruses 11, no. 5: 476. https://doi.org/10.3390/v11050476