Characterization of Unknown Orthobunya-Like Viruses from India


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. NGS of Unknown Infectious Agents
2.2. Bioinformatic Analysis
2.3. Reverse Transcription PCR and Sanger Sequencing
2.4. Isolation and Characterization of Cat Que Virus Strains JM1 and PB1
3. Results
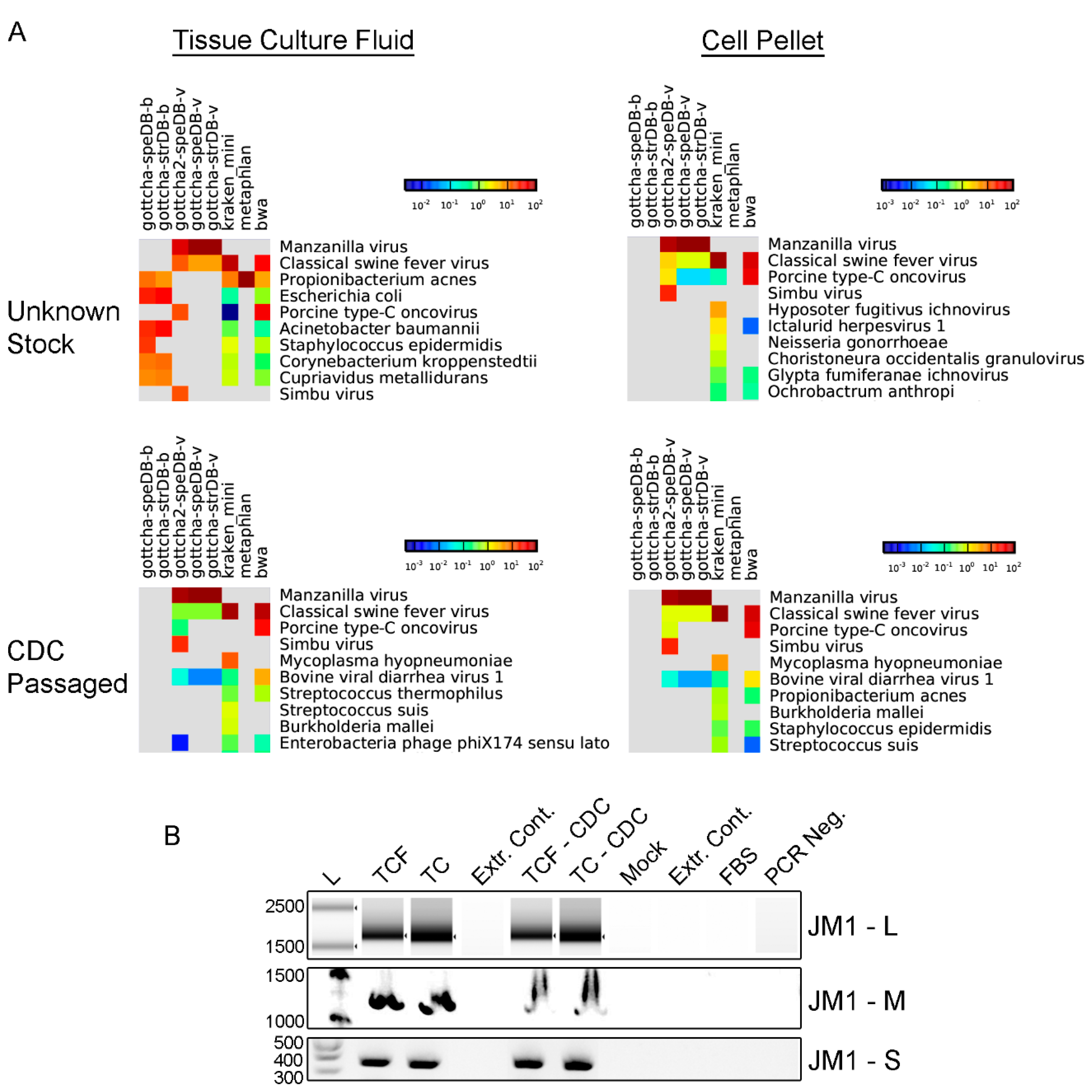
3.1. Identification of Novel Infectious Agents by Sequencing
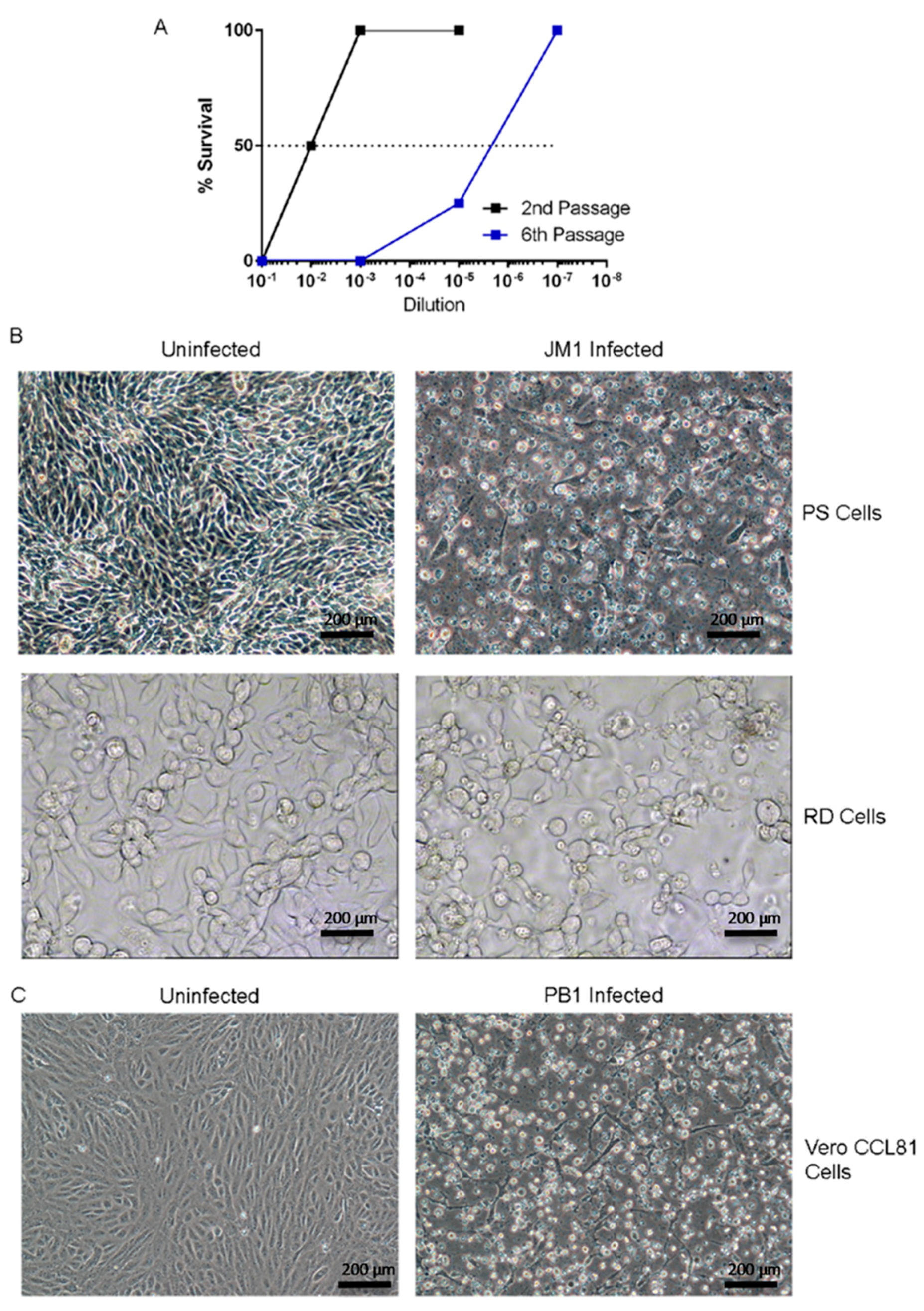
3.2. Isolation and Characterization of an Unknown Infectious Agent
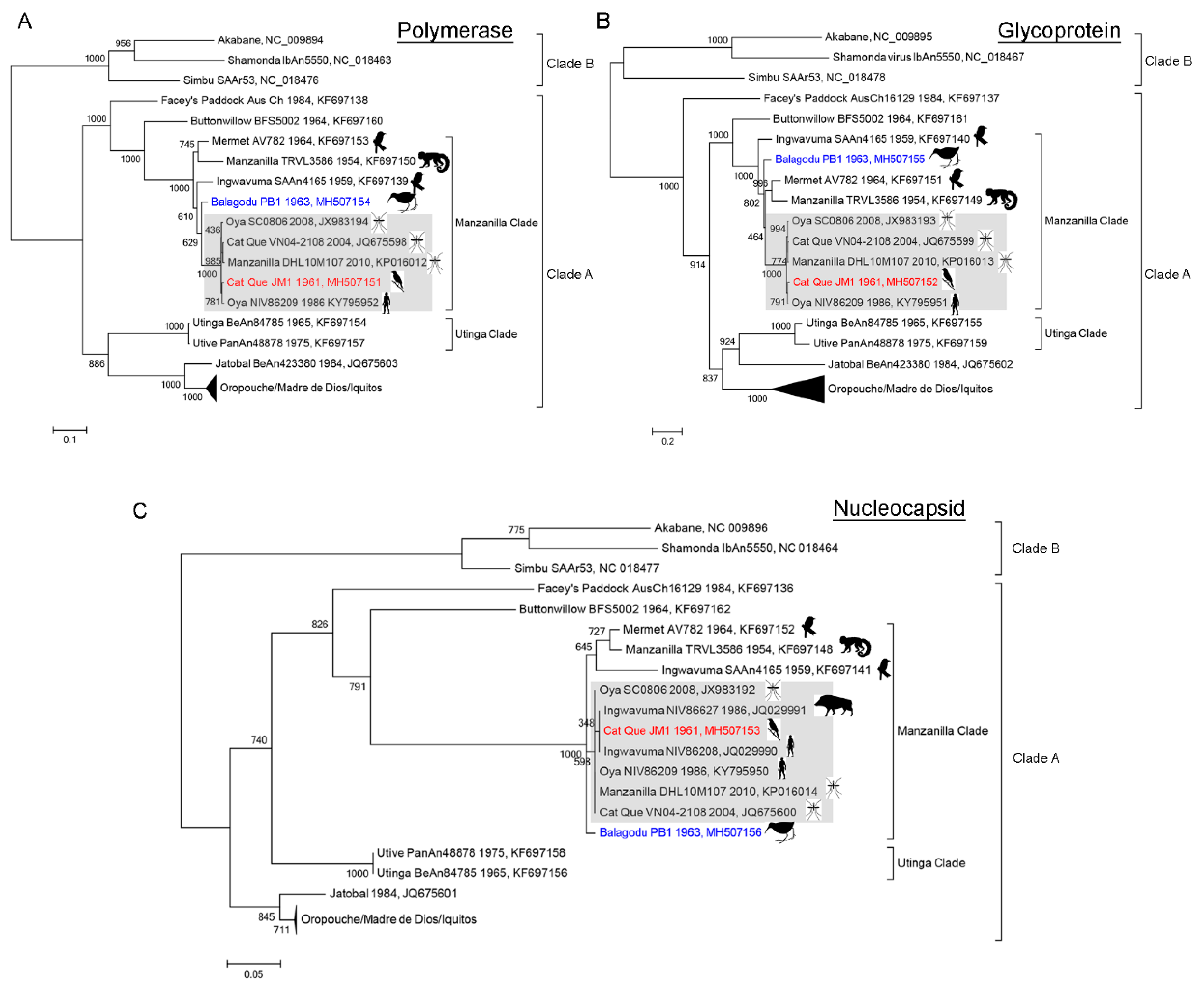
3.3. Inference of Phylogenetic Relationships
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rivers, T.M. Viruses and Koch’s Postulates. J. Bacteriol. 1937, 33, 1–12. [Google Scholar] [PubMed]
- Lipkin, W.I. Microbe hunting. Microbiol. Mol. Biol. Rev. 2010, 74, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Fredricks, D.N.; Relman, D.A. Sequence-based identification of microbial pathogens: A reconsideration of Koch’s postulates. Clin. Microbiol. Rev. 1996, 9, 18–33. [Google Scholar] [PubMed]
- Piantadosi, A.; Mukerji, S.S.; Chitneni, P.; Cho, T.A.; Cosimi, L.A.; Hung, D.T.; Goldberg, M.B.; Sabeti, P.C.; Kuritzkes, D.R.; Grad, Y.H. Metagenomic Sequencing of an Echovirus 30 Genome From Cerebrospinal Fluid of a Patient With Aseptic Meningitis and Orchitis. Open Forum. Infect. Dis. 2017, 4, ofx138. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, O.I.; Lambert, A.J.; Hawkinson, D.J.; Pastula, D.M.; Goldsmith, C.S.; Hunt, D.C.; Staples, J.E. Novel thogotovirus associated with febrile illness and death, United States, 2014. Emerg. Infect. Dis. 2015, 21, 760–764. [Google Scholar] [CrossRef] [PubMed]
- McMullan, L.K.; Folk, S.M.; Kelly, A.J.; MacNeil, A.; Goldsmith, C.S.; Metcalfe, M.G.; Batten, B.C.; Albarino, C.G.; Zaki, S.R.; Rollin, P.E.; et al. A new phlebovirus associated with severe febrile illness in Missouri. N. Engl. J. Med. 2012, 367, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y. Viral pathogen discovery. Curr. Opin. Microbiol. 2013, 16, 468–478. [Google Scholar] [CrossRef] [PubMed]
- Pavri, K.; Sheikh, B.H.; Singh, K.R.; Rajagopalan, P.K.; Casals, J. Balagodu virus, a new arbovirus isolated from Ardeola grayii (Sykes) in Mysore State, South India. Indian J. Med. Res. 1969, 57, 758–764. [Google Scholar] [PubMed]
- Bhatt, P.N.; Work, T.H.; Varma, M.G.; Trapido, H.; Murthy, D.P.; Rodrigues, F.M. Kyasanur forest diseases. IV. Isolation of Kyasanur forest disease virus from infected humans and monkeys of Shimogadistrict, Mysore state. Indian J. Med. Sci. 1966, 20, 316–320. [Google Scholar] [PubMed]
- Work, T.H.; Trapido, H.; Murthy, D.P.; Rao, R.L.; Bhatt, P.N.; Kulkarni, K.G. Kyasanur forest disease. III. A preliminary report on the nature of the infection and clinical manifestations in human beings. Indian J. Med. Sci. 1957, 11, 619–645. [Google Scholar] [PubMed]
- Go, Y.Y.; Balasuriya, U.B.; Lee, C.K. Zoonotic encephalitides caused by arboviruses: Transmission and epidemiology of alphaviruses and flaviviruses. Clin. Exp. Vaccine Res. 2014, 3, 58–77. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Kuiken, T. Changing Role of Wild Birds in the Epidemiology of Avian Influenza A Viruses. Adv. Virus Res. 2018, 100, 279–307. [Google Scholar] [PubMed]
- Elliott, R.M. Bunyaviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Wolters Kluwer Health: Philadelphia, PA, USA, 2013; Volume 1. [Google Scholar]
- Naveca, F.G.; Nascimento, V.A.; Souza, V.C.; de Figueiredo, R.M.P. Human Orthobunyavirus Infections, Tefe, Amazonas, Brazil. PLoS Curr. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Webster, D.; Dimitrova, K.; Holloway, K.; Makowski, K.; Safronetz, D.; Drebot, M.A. California Serogroup Virus Infection Associated with Encephalitis and Cognitive Decline, Canada, 2015. Emerg. Infect. Dis. 2017, 23, 1423–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teleron, A.L.; Rose, B.K.; Williams, D.M.; Kemper, S.E.; McJunkin, J.E. La Crosse Encephalitis: An Adult Case Series. Am. J. Med. 2016, 129, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.D.; Chaubal, G.Y.; Shete, A.M.; Mourya, D.T. A mini-review of Bunyaviruses recorded in India. Indian J. Med. Res. 2017, 145, 601–610. [Google Scholar] [PubMed]
- Sakkas, H.; Bozidis, P.; Franks, A.; Papadopoulou, C. Oropouche Fever: A Review. Viruses 2018, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Travassos da Rosa, J.F.; de Souza, W.M.; Pinheiro, F.P.; Figueiredo, M.L.; Cardoso, J.F.; Acrani, G.O.; Nunes, M.R.T. Oropouche Virus: Clinical, Epidemiological, and Molecular Aspects of a Neglected Orthobunyavirus. Am. J. Trop. Med. Hyg. 2017, 96, 1019–1030. [Google Scholar] [PubMed]
- De Regge, N. Akabane, Aino and Schmallenberg virus-where do we stand and what do we know about the role of domestic ruminant hosts and Culicoides vectors in virus transmission and overwintering? Curr. Opin. Virol. 2017, 27, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Plyusnin, A.; Beaty, B.J.; Elliott, R.M.; Goldbach, R.; Kormelink, R.; Lundkvist, Å.; Schmaljohn, C.S.; Tesh, R.B. Bunyaviridae; International Committee on Taxonomy of Viruses: San Diego, CA, USA, 2017. [Google Scholar]
- Briese, T.; Calisher, C.H.; Higgs, S. Viruses of the family Bunyaviridae: Are all available isolates reassortants? Virology 2013, 446, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladner, J.T.; Savji, N.; Lofts, L.; Travassos da Rosa, A.; Wiley, M.R.; Gestole, M.C.; Rosen, G.E.; Guzman, H.; Vasconcelos, P.F.; Nunes, M.R.; et al. Genomic and phylogenetic characterization of viruses included in the Manzanilla and Oropouche species complexes of the genus Orthobunyavirus, family Bunyaviridae. J. Gen. Virol. 2014, 95, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, J.C.; Giambalvo, D.; Hernandez, R.; Auguste, A.J.; Tesh, R.B.; Weaver, S.C.; Montanez, H.; Liria, J.; Lima, A.; Travassos da Rosa, J.F.; et al. Isolation of Madre de Dios Virus (Orthobunyavirus; Bunyaviridae), an Oropouche Virus Species Reassortant, from a Monkey in Venezuela. Am. J. Trop. Med. Hyg. 2016, 95, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Palacios, G.; da Rosa, A.T.; Savji, N.; Sze, W.; Wick, I.; Guzman, H.; Hutchison, S.; Tesh, R.; Lipkin, W.I. Aguacate virus, a new antigenic complex of the genus Phlebovirus (family Bunyaviridae). J. Gen. Virol. 2011, 92, 1445–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerrard, S.R.; Li, L.; Barrett, A.D.; Nichol, S.T. Ngari virus is a Bunyamwera virus reassortant that can be associated with large outbreaks of hemorrhagic fever in Africa. J. Virol. 2004, 78, 8922–8926. [Google Scholar] [CrossRef] [PubMed]
- Matranga, C.B.; Andersen, K.G.; Winnicki, S.; Busby, M.; Gladden, A.D.; Tewhey, R.; Stremlau, M.; Berlin, A.; Gire, S.K.; England, E.; et al. Enhanced methods for unbiased deep sequencing of Lassa and Ebola RNA viruses from clinical and biological samples. Genome Biol. 2014, 15, 519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiconis, X.; Borges-Rivera, D.; Satija, R.; DeLuca, D.S.; Busby, M.A.; Berlin, A.M.; Sivachenko, A.; Thompson, D.A.; Wysoker, A.; Fennell, T.; et al. Comparative analysis of RNA sequencing methods for degraded or low-input samples. Nat. Methods 2013, 10, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matranga, C.B.; Gladden-Young, A.; Qu, J.; Winnicki, S.; Nosamiefan, D.; Levin, J.Z.; Sabeti, P.C. Unbiased Deep Sequencing of RNA Viruses from Clinical Samples. J. Vis. Exp. 2016, 113, 54117. [Google Scholar] [CrossRef] [PubMed]
- Li, P.E.; Lo, C.C.; Anderson, J.J.; Davenport, K.W.; Bishop-Lilly, K.A.; Xu, Y.; Ahmed, S.; Feng, S.; Mokashi, V.P.; Chain, P.S. Enabling the democratization of the genomics revolution with a fully integrated web-based bioinformatics platform. Nucleic Acids Res. 2017, 45, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Viral-ngs: Genomics Analysis Pipelines for Viral Sequencing. Available online: https://viral-ngs.readthedocs.io/en/latest/ (accessed on 30 January 2018).
- Ridpath, J.F.; Bolin, S.R. Differentiation of types 1a, 1b and 2 bovine viral diarrhoea virus (BVDV) by PCR. Mol. Cell. Probes. 1998, 12, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Patience, C.; Takeuchi, Y.; Weiss, R.A. Infection of human cells by an endogenous retrovirus of pigs. Nat. Med. 1997, 3, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Van Kuppeveld, F.J.; van der Logt, J.T.; Angulo, A.F.; van Zoest, M.J.; Quint, W.G.; Niesters, H.G.; Galama, J.M.; Melchers, W.J. Genus- and species-specific identification of mycoplasmas by 16S rRNA amplification. Appl. Environ. Microbiol. 1992, 58, 2606–2615. [Google Scholar] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Jerome, K.R.; Lennette, E.H. Lennette’s Laboratory Diagnosis of Viral Infections; CRC Press: Boca Raton, FL, USA, 2010; Volume 50. [Google Scholar]
- Pawar, S.D.; Singh, A.; Gangodkar, S.V.; Rao, B.L. Propagation of Chandipura virus in chick embryos. Indian J. Exp. Biol. 2005, 43, 930–932. [Google Scholar] [PubMed]
- Gagnieur, L.; Cheval, J.; Gratigny, M.; Hebert, C.; Muth, E.; Dumarest, M.; Eloit, M. Unbiased analysis by high throughput sequencing of the viral diversity in fetal bovine serum and trypsin used in cell culture. Biologicals 2014, 42, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro de Oliveira, T.F.; Fonseca, A.A., Jr.; Camargos, M.F.; de Oliveira, A.M.; Pinto Cottorello, A.C.; Souza Ados, R.; de Almeida, I.G.; Heinemann, M.B. Detection of contaminants in cell cultures, sera and trypsin. Biologicals 2013, 41, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Kaden, V.; Lange, E.; Steyer, H.; Bruer, W.; Langner, C.H. Role of birds in transmission of classical swine fever virus. J. Vet. Med. B Infect. Dis. Vet. Public Health 2003, 50, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Shete, A.; Bondre, V.; Patil, D.; Kokate, P.; Chaudhari, S.; Srivastava, S.; Jadhav, S.; Mourya, D. Isolation and characterization of Oya virus a member of Simbu serogroup, family Bunyaviridae, isolated from Karnataka, India. Infect. Genet. Evol. 2016, 44, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, J.; Wang, L.; Fu, S.; Li, M.; Zhao, G.; Zhu, W.; Wang, D.; Liang, G. Molecular Characterization and Seroprevalence in Pigs of SC0806, a Cat Que Virus Isolated from Mosquitoes in Sichuan Province, China. Vector Borne Zoonotic Dis. 2015, 15, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kono, Y.; Yusnita, Y.; Mohd Ali, A.R.; Maizan, M.; Sharifah, S.H.; Fauzia, O.; Kubo, M.; Aziz, A.J. Characterization and identification of Oya virus, a Simbu serogroup virus of the genus Bunyavirus, isolated from a pig suspected of Nipah virus infection. Arch. Virol. 2002, 147, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.E.; Crabtree, M.B.; Nam, V.S.; Yen, N.T.; Duc, H.M.; Miller, B.R. Isolation of arboviruses from mosquitoes collected in northern Vietnam. Am. J. Trop. Med. Hyg. 2005, 73, 470–473. [Google Scholar] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antivir. Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
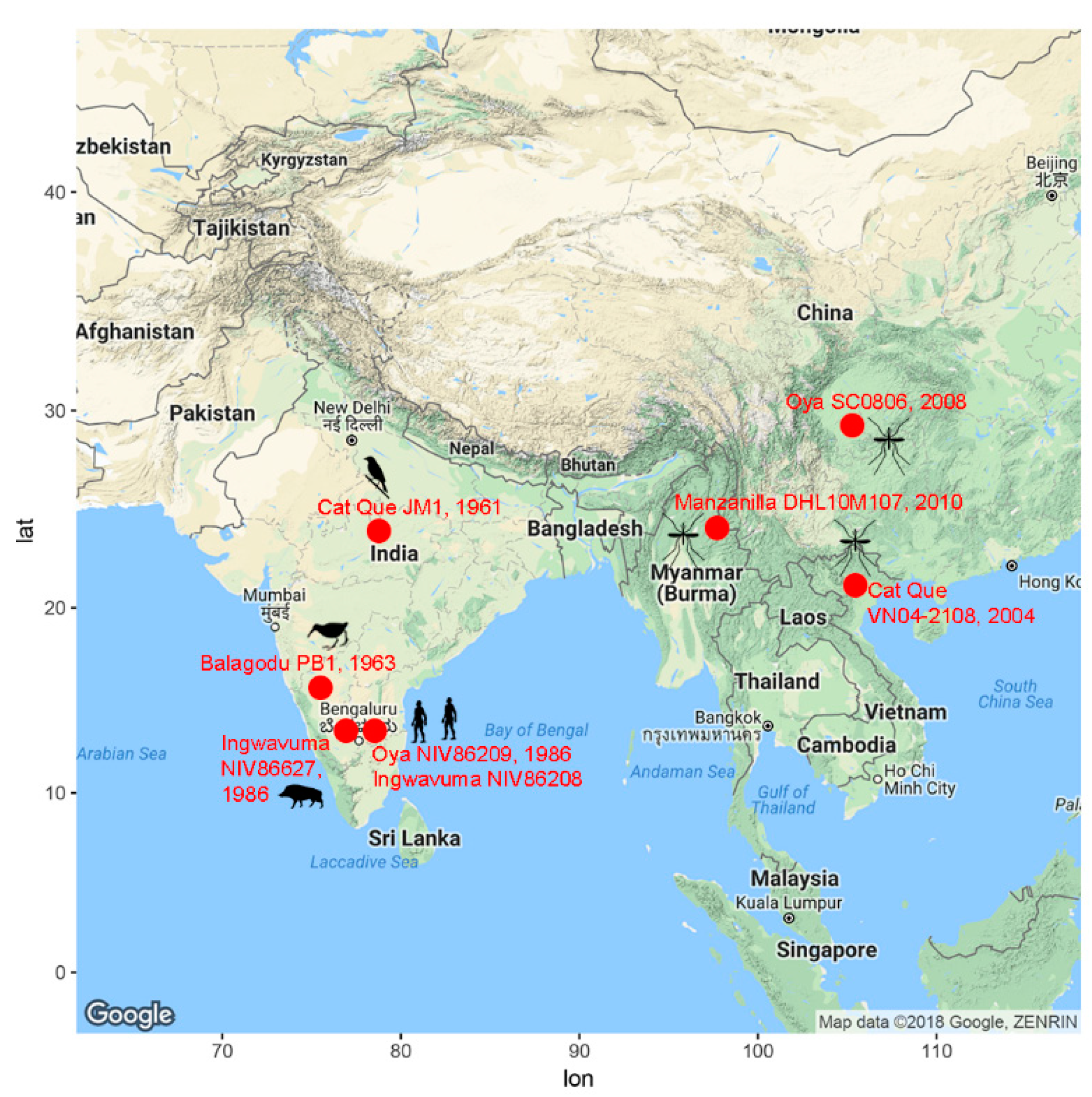{kind=link}
| Inoculum | Organism | Route | Age | Mortality | Mortality at Day Post Infection (DPI) | Remarks |
|---|---|---|---|---|---|---|
| Mouse brain suspension, Neat | Swiss mice | intraperitoneal | 1 do | 8/8 | 4 | |
| Mouse brain suspension, Neat | Swiss mice | intracranial | 10 do | 8/8 | 3 | |
| Mouse brain suspension, Neat | Swiss mice | intraperitoneal | 10 do | 4 */8 | 6 | * Paralyzed Mice died on 7 dpi, Left leg paralysis noted in a survivor. |
| Mouse brain suspension, Neat | Swiss mice | intraperitoneal | 17 do | 1 */8 | 7 | * Paralyzed mouse died on 8 dpi. |
| Mouse brain suspension, Neat | Chicken | Allantoic | est. 8–10 do | 2/3 | 2, 3 | Hemorrhages on head, frank oozing of blood from skull, generalized edema, body hemorrhages. |
| Tissue Culture Fluid (<3 passages) | Chicken | Allantoic | est. 8–10 do | 1/7 | 6 | Hemorrhages on heart and liver. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Whitmer, S.L.M.; Yadav, P.D.; Sarkale, P.; Chaubal, G.Y.; Francis, A.; Klena, J.; Nichol, S.T.; Ströher, U.; Mourya, D.T. Characterization of Unknown Orthobunya-Like Viruses from India. Viruses 2018, 10, 451. https://doi.org/10.3390/v10090451
Whitmer SLM, Yadav PD, Sarkale P, Chaubal GY, Francis A, Klena J, Nichol ST, Ströher U, Mourya DT. Characterization of Unknown Orthobunya-Like Viruses from India. Viruses. 2018; 10(9):451. https://doi.org/10.3390/v10090451
Chicago/Turabian StyleWhitmer, Shannon L. M., Pragya D. Yadav, Prasad Sarkale, Gouri Y. Chaubal, Alicia Francis, John Klena, Stuart T. Nichol, Ute Ströher, and Devendra T. Mourya. 2018. "Characterization of Unknown Orthobunya-Like Viruses from India" Viruses 10, no. 9: 451. https://doi.org/10.3390/v10090451