
Prevalence and Association of Trypanosomes and Sodalis glossinidius in Tsetse Flies from the Kafue National Park in Zambia

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
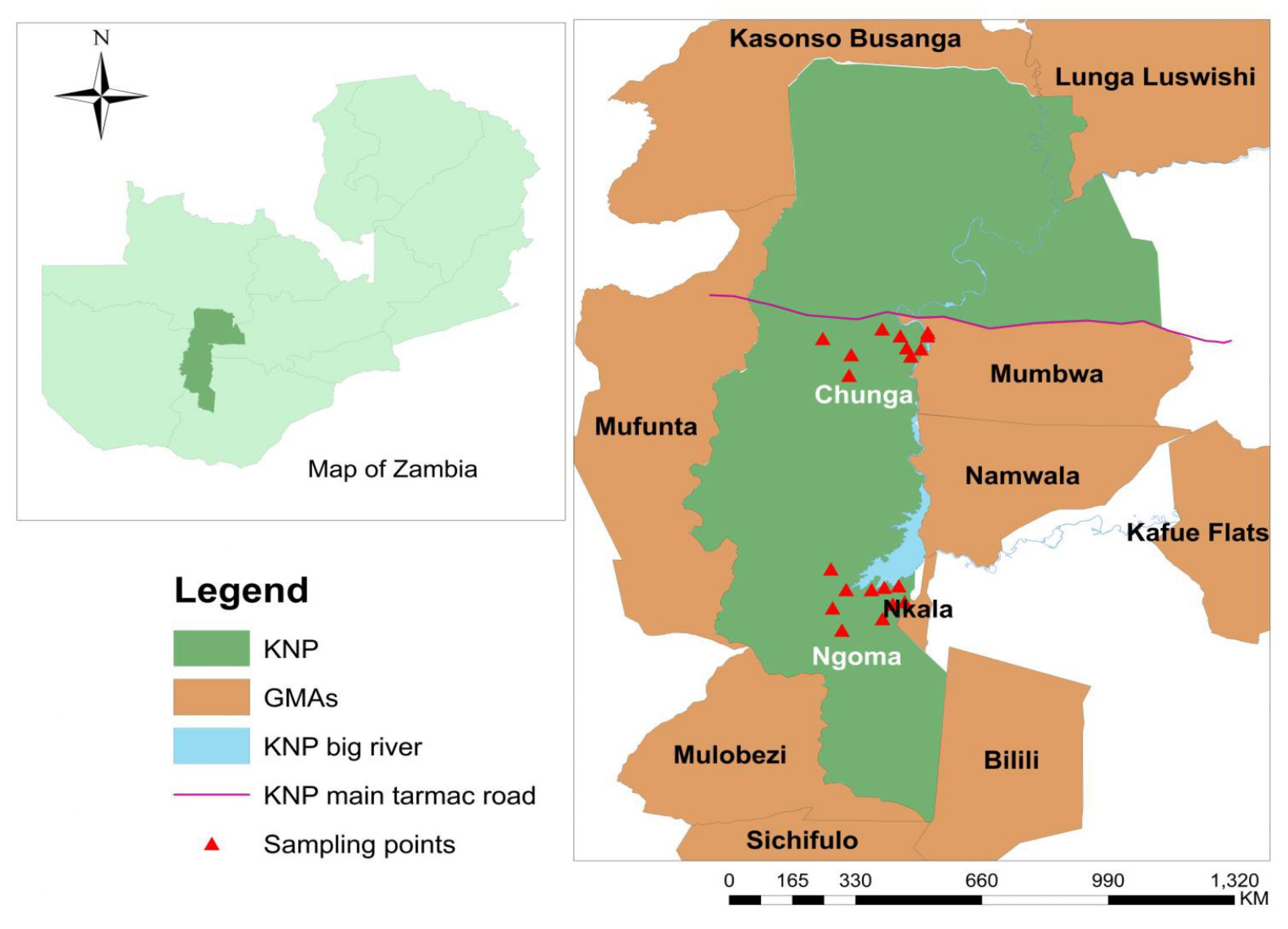
2.1. Study Area and Tsetse Fly Collection
2.2. Sex and Species Determination of Tsetse Flies
2.3. DNA Extraction
2.4. PCR for the Identification of African Trypanosomes and Sodalis glossinidius
2.5. Data Analysis
3. Results
3.1. Tsetse Fly Survey
3.2. Prevalence of Trypanosomes in Tsetse Flies
3.3. Prevalence of S. glossinidius in Tsetse Flies
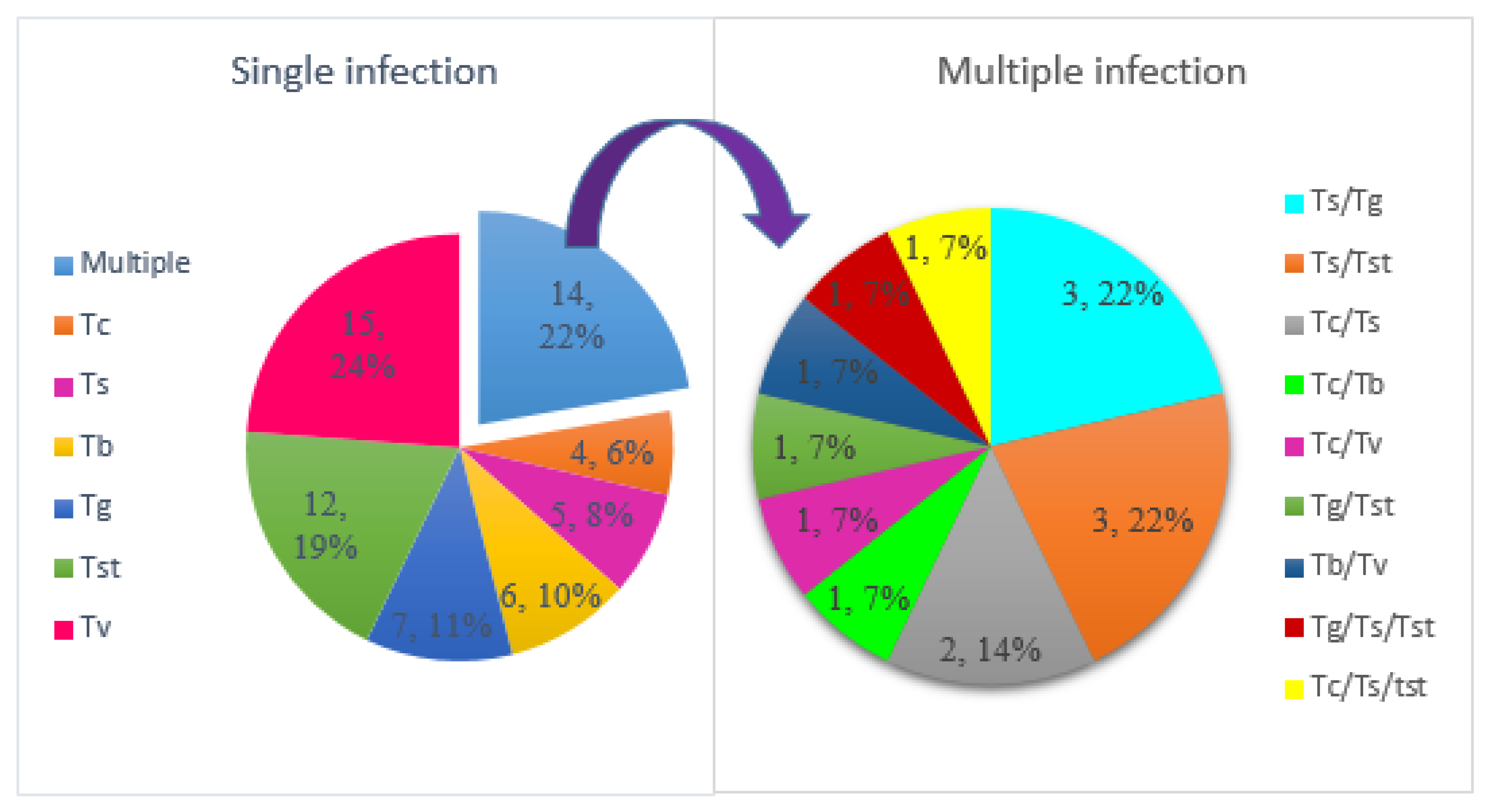
3.4. Association between S. glossinidius and Presence of African Trypanosomes
4. Discussion
4.1. Prevalence of Trypanosome Species
4.2. Prevalence of S. glossinidius
4.3. Association between S. glossinidius and Trypanosome Infections in Tsetse Flies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geiger, A.; Ponton, F.; Simo, G. Adult blood-feeding tsetse flies, trypanosomes, microbiota and the fluctuating environment in sub-Saharan Africa. ISME J. 2015, 9, 1496–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristjanson, P.M.; Swallow, B.M.; Rowlands, G.J.; Kruska, R.L.; de Leeuw, P.N. Measuring the costs of African animal trypanosomosis, the potential benefits of control and returns to research. Agric. Syst. 1999, 59, 79–98. [Google Scholar] [CrossRef]
- Steverding, D. The history of African trypanosomiasis. Parasites Vectors 2008, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.P.M.; Cecchi, G.; Wint, G.R.W.; Mattioli, R.C.; Robinson, T.P. Mapping the economic benefits to livestock keepers from intervening against bovine trypanosomosis in Eastern Africa. Prev. Vet. Med. 2014, 113, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Swallow, B.S. Impacts of Trypanosomiasis on African Agriculture; ILRI/FAO: Rome, Italy, 2000. [Google Scholar]
- Holmes, P. Tsetse-transmitted trypanosomes—Their biology, disease impact and control. J. Invertebr. Pathol. 2013, 112, S11–S14. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.R.; Hamilton, P.B.; Gibson, W.C. African trypanosomes: Celebrating diversity. Trends Parasitol. 2010, 26, 324–328. [Google Scholar] [CrossRef]
- Cecchi, G.; Paone, M.; Feldmann, U.; Vreysen, M.J.B.; Diall, O.; Mattioli, R.C. Assembling a geospatial database of tsetse-transmitted animal trypanosomosis for Africa. Parasit Vectors 2014, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Büscher, P.; Cecchi, G.; Jamonneau, V.; Priotto, G. Human African trypanosomiasis. Lancet 2017, 390, 2397–2409. [Google Scholar] [CrossRef]
- WHO. Trypanosomiasis, Human African (Sleeping Sickness). 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 8 February 2021).
- Rock, K.S.; Stone, C.M.; Hastings, I.M.; Keeling, M.J.; Torr, S.J.; Chitnis, N. Mathematical models of human African trypanosomiasis epidemiology. Adv. Parasitol. 2015, 87, 53–133. [Google Scholar] [CrossRef]
- WHO. Control and Surveillance of Human African Trypanosomiasis: Report of a WHO Expert Committee; World Health Organization: Geneva, Switzerland, 2013. Available online: https://apps.who.int/iris/handle/10665/95732 (accessed on 8 February 2021).
- Wang, J.; Weiss, B.L.; Aksoy, S. Tsetse fly microbiota: Form and function. Front. Cell. Infect. Microbiol. 2013, 3, 69. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Gooding, R.H.; Aksoy, S. Phylogenetically distant symbiotic microorganisms reside in Glossina midgut and ovary tissues. Med. Vet. Entomol. 1993, 7, 377–383. [Google Scholar] [CrossRef]
- Cheng, Q.; Aksoy, S. Tissue tropism, transmission and expression of foreign genes in vivo in midgut symbionts of tsetse flies. Insect Mol. Biol. 1999, 8, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Welburn, S.C.; Maudlin, I. Tsetse-trypanosome interactions: Rites of passage. Parasitol. Today 1999, 15, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Maudlin, I.; Welburn, S.C. Lectin mediated establishment of midgut infections of Trypanosoma congolense and Trypanosoma brucei in Glossina morsitans. Trop. Med. Parasitol. 1987, 38, 167–170. [Google Scholar] [PubMed]
- Attardo, G.M.; Guz, N.; Strickler-Dinglasan, P.; Aksoy, S. Molecular aspects of viviparous reproductive biology of the tsetse fly (Glossina morsitans morsitans): Regulation of yolk and milk gland protein synthesis. J. Insect Physiol. 2006, 52, 1128–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beard, C.B.; O’Neill, S.L.; Mason, P.; Mandelco, L.; Woese, C.R.; Tesh, R.B.; Richards, F.F.; Aksoy, S. Genetic transformation and phylogeny of bacterial symbionts from tsetse. Insect Mol. Biol. 1993, 1, 123–131. [Google Scholar] [CrossRef]
- De Vooght, L.; Van Keer, S.; Van Den Abbeele, J. Towards improving tsetse fly paratransgenesis: Stable colonization of Glossina morsitans morsitans with genetically modified Sodalis. BMC Microbiol. 2018, 18, 31–38. [Google Scholar] [CrossRef]
- Makhulu, E.E.; Villinger, J.; Adunga, V.O.; Jeneby, M.M.; Kimathi, E.M.; Mararo, E.; Oundo, J.W.; Musa, A.A.; Wambua, L. Tsetse blood-meal sources, endosymbionts and trypanosome-associations in the Maasai Mara National Reserve, a wildlife-human-livestock interface. PLoS Negl. Trop. Dis. 2021, 15, e0008267. [Google Scholar] [CrossRef]
- Dennis, J.W.; Durkin, S.M.; Horsley Downie, J.E.; Hamill, L.C.; Anderson, N.E.; MacLeod, E.T. Sodalis glossinidius prevalence and trypanosome presence in tsetse from Luambe National Park, Zambia. Parasit Vectors 2014, 7, 378. [Google Scholar] [CrossRef] [Green Version]
- Mbewe, N.J.; Mweempwa, C.; Guya, S.; Wamwiri, F.N. Microbiome frequency and their association with trypanosome infection in male Glossina morsitans centralis of Western Zambia. Vet. Parasitol. 2015, 211, 93–98. [Google Scholar] [CrossRef]
- Geiger, A.; Ravel, S.; Mateille, T.; Janelle, J.; Patrel, D.; Cuny, G.; Frutos, R. Vector competence of Glossina palpalis gambiensis for Trypanosoma brucei s.l. and genetic diversity of the symbiont Sodalis glossinidius. Mol. Biol. Evol. 2007, 24, 102–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farikou, O.; Njiokou, F.; Mbida Mbida, J.A.; Njitchouang, G.R.; Djeunga, H.N.; Asonganyi, T.; Simarro, P.P.; Cuny, G.; Geiger, A. Tripartite interactions between tsetse flies, Sodalis glossinidius and trypanosomes--an epidemiological approach in two historical human African trypanosomiasis foci in Cameroon. Infect. Genet. Evol. 2010, 10, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Thapa, B.; Child, B.; Parent, G.; Mupeta, P. Tourism Demand Assessment-Kafue National Park, Zambia. Center for African Studies Research Report; University of Florida: Gainesville, FL, USA, 2011; pp. 58–59. [Google Scholar]
- Mwima, H. A Brief history of Kafue National Park, Zambia. Koedoe Afr. Prot. Area Conserv. Sci. 2001, 44, 57–72. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Hayashida, K.; Delesalle, V.; Qiu, Y.; Omori, R.; Simuunza, M.; Sugimoto, C.; Namangala, B.; Yamagishi, J. Genetic diversity of African trypanosomes in tsetse flies and cattle from the Kafue ecosystem. Front. Vet. Sci. 2021, 8, 599815. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.N. Training Manual for Tsetse Control Personnel: Tsetse Biology, Systematics and Distribution; Techniques; FAO: Rome, Italy, 1982; Volume 1. [Google Scholar]
- Njiru, Z.K.; Constantine, C.C.; Guya, S.; Crowther, J.; Kiragu, J.M.; Thompson, R.C.A.; Dávila, A.M.R. The use of ITS1 rDNA PCR in detecting pathogenic African trypanosomes. Parasitol. Res. 2005, 95, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Desquesnes, M.; McLaughlin, G.; Zoungrana, A.; Dávila, A.M. Detection and identification of Trypanosoma of African livestock through a single PCR based on internal transcribed spacer 1 of rDNA. Int. J. Parasitol. 2001, 31, 610–614. [Google Scholar] [CrossRef]
- Adams, E.R.; Malele, I.I.; Msangi, A.R.; Gibson, W.C. Trypanosome identification in wild tsetse populations in Tanzania using generic primers to amplify the ribosomal RNA ITS-1 region. Acta Trop. 2006, 100, 103–109. [Google Scholar] [CrossRef]
- Masiga, D.K.; Smyth, A.J.; Hayes, P.; Bromidge, T.J.; Gibson, W.C. Sensitive detection of trypanosomes in tsetse flies by DNA amplification. Int. J. Parasitol. 1992, 22, 909–918. [Google Scholar] [CrossRef]
- Radwanska, M.; Chamekh, M.; Vanhamme, L.; Claes, F.; Magez, S.; Magnus, E.; de Baetselier, P.; Büscher, P.; Pays, E. The serum resistance-associated gene as a diagnostic tool for the detection of Trypanosoma brucei rhodesiense. Am. J. Trop. Med. Hyg. 2002, 67, 684–690. [Google Scholar] [CrossRef] [Green Version]
- Majiwa, P.A.; Thatthi, R.; Moloo, S.K.; Nyeko, J.H.; Otieno, L.H.; Maloo, S. Detection of trypanosome infections in the saliva of tsetse flies and buffy-coat samples from antigenaemic but aparasitaemic cattle. Parasitology 1994, 108 Pt 3, 313–322. [Google Scholar] [CrossRef]
- Masiga, D.K.; McNamara, J.J.; Laveissière, C.; Truc, P.; Gibson, W.C. A high prevalence of mixed trypanosome infections in tsetse flies in Sinfra, Côte d’Ivoire, detected by DNA amplification. Parasitology 1996, 112 Pt 1, 75–80. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 7 November 2021).
- Nakayima, J.; Nakao, R.; Alhassan, A.; Mahama, C.; Afakye, K.; Sugimoto, C. Molecular epidemiological studies on animal trypanosomiases in Ghana. Parasit Vectors 2012, 5, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musaya, J.; Chisi, J.; Senga, E.; Nambala, P.; Maganga, E.; Matovu, E.; Enyaru, J. Polymerase chain reaction identification of Trypanosoma brucei rhodesiense in wild tsetse flies from Nkhotakota Wildlife Reserve, Malawi. Malawi Med. J. 2017, 29, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Nayupe, S.F.; Simwela, N.V.; Kamanga, P.M.; Chisi, J.E.; Senga, E.; Musaya, J.; Maganga, E. The use of molecular technology to investigate trypanosome infections in tsetse flies at Liwonde Wild Life Reserve. Malawi Med. J. 2019, 31, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Laohasinnarong, D.; Goto, Y.; Goto, Y.; Asada, M.; Nakao, R.; Hayashida, K.; Kajino, K.; Kawazu, S.; Sugimoto, C.; Inoue, N.; et al. Studies of trypanosomiasis in the Luangwa valley, north-eastern Zambia. Parasit Vectors 2015, 8, 497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, C.M.F.; Garcia, H.A.; Sheferaw, D.; Rodrigues, A.C.; Pereira, C.L.; Camargo, E.P.; Teixeira, M.M.G. Genetic diversity of trypanosomes pathogenic to livestock in tsetse flies from the Nech Sar National Park in Ethiopia: A concern for tsetse suppressed area in Southern Rift Valley? Infect. Genet. Evol. 2019, 69, 38–47. [Google Scholar] [CrossRef]
- Channumsin, M.; Ciosi, M.; Masiga, D.; Turner, C.M.R.; Mable, B.K. Sodalis glossinidius presence in wild tsetse is only associated with presence of trypanosomes in complex interactions with other tsetse-specific factors. BMC Microbiol. 2018, 18, 163. [Google Scholar] [CrossRef] [Green Version]
- Kamdem, C.N.; Tiofack, A.A.Z.; Mewamba, E.M.; Ofon, E.A.; Gomseu, E.B.D.; Simo, G. Molecular identification of different trypanosome species in tsetse flies caught in the wildlife reserve of Santchou in the western region of Cameroon. Parasitol. Res. 2020, 119, 805–813. [Google Scholar] [CrossRef]
- Simwango, M.; Ngonyoka, A.; Nnko, H.J.; Salekwa, L.P.; Ole-Neselle, M.; Kimera, S.I.; Gwakisa, P.S. Molecular prevalence of trypanosome infections in cattle and tsetse flies in the Maasai Steppe, northern Tanzania. Parasit Vectors 2017, 10, 507. [Google Scholar] [CrossRef] [Green Version]
- Shereni, W.; Anderson, N.E.; Nyakupinda, L.; Cecchi, G. Spatial distribution and trypanosome infection of tsetse flies in the sleeping sickness focus of Zimbabwe in Hurungwe District. Parasit Vectors 2016, 9, 605. [Google Scholar] [CrossRef]
- Opiro, R.; Opoke, R.; Angwech, H.; Nakafu, E.; Oloya, F.A.; Openy, G.; Njahira, M.; Macharia, M.; Echodu, R.; Malinga, G.M.; et al. Apparent density, trypanosome infection rates and host preference of tsetse flies in the sleeping sickness endemic focus of northwestern Uganda. BMC Vet. Res. 2021, 17, 365. [Google Scholar] [CrossRef]
- Wamwiri, F.N.; Alam, U.; Thande, P.C.; Aksoy, E.; Ngure, R.M.; Aksoy, S.; Ouma, J.O.; Murilla, G.A. Wolbachia, Sodalis and trypanosome co-infections in natural populations of Glossina austeni and Glossina pallidipes. Parasit Vectors 2013, 6, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, N.A.; Rose, C.; Ejeh, N.O.; Acosta-Serrano, A. Flying tryps: Survival and maturation of trypanosomes in tsetse flies. Trends Parasitol. 2013, 29, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Rotureau, B.; Van Den Abbeele, J. Through the dark continent: African trypanosome development in the tsetse fly. Front. Cell. Infect. Microbiol. 2013, 3, 53. [Google Scholar] [CrossRef] [Green Version]
- Signaboubo, D.; Payne, V.K.; Moussa, I.M.A.; Hassane, H.M.; Berger, P.; Kelm, S.; Simo, G. Diversity of tsetse flies and trypanosome species circulating in the area of Lake Iro in southeastern Chad. Parasites Vectors 2021, 14, 293. [Google Scholar] [CrossRef]
- Welburn, S.C.; Maudlin, I. The nature of the teneral state in Glossina and its role in the acquisition of trypanosome infection in tsetse. Ann. Trop. Med. Parasitol. 1992, 86, 529–536. [Google Scholar] [CrossRef]
- Isaac, C.; Ciosi, M.; Hamilton, A.; Scullion, K.M.; Dede, P.; Igbinosa, I.B.; Nmorsi, O.P.G.; Masiga, D.; Turner, C.M.R. Molecular identification of different trypanosome species and subspecies in tsetse flies of northern Nigeria. Parasit Vectors 2016, 9, 301. [Google Scholar] [CrossRef] [Green Version]
- Jamonneau, V.; Ravel, S.; Koffi, M.; Kaba, D.; Zeze, D.G.; Ndri, L.; Sane, B.; Coulibaly, B.; Cuny, G.; Solano, P. Mixed infections of trypanosomes in tsetse and pigs and their epidemiological significance in a sleeping sickness focus of Côte d’Ivoire. Parasitology 2004, 129 Pt 6, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Moloo, S.K.; Sabwa, C.L.; Kabata, J.M. Vector competence of Glossina pallidipes and G. morsitans centralis for Trypanosoma vivax, T. congolense and T. b. brucei. Acta Trop. 1992, 51, 271–280. [Google Scholar] [CrossRef]
- Squarre, D.; Kabongo, I.; Munyeme, M.; Mumba, C.; Mwasinga, W.; Hachaambwa, L.; Sugimoto, C.; Namangala, B. Human African trypanosomiasis in the Kafue National Park, Zambia. PLoS Negl. Trop. Dis. 2016, 10, e0004567. [Google Scholar] [CrossRef]
- Squarre, D.; Hayashida, K.; Gaithuma, A.; Chambaro, H.; Kawai, N.; Moonga, L.; Namangala, B.; Sugimoto, C.; Yamagishi, J. Diversity of trypanosomes in wildlife of the Kafue ecosystem, Zambia. Int. J. Parasitol. Parasites Wildl. 2020, 12, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Odeniran, P.O.; Macleod, E.T.; Ademola, I.O.; Welburn, S.C. Endosymbionts interaction with trypanosomes in Palpalis group of Glossina captured in southwest Nigeria. Parasitol. Int. 2019, 70, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Mathew, C.Z. Biological and Molecular Aspects of Sodalis Glossinidius. Ph.D. Thesis, The University of Edinburgh, College of Medicine and Veterinary Medicine, Edinburgh, UK, 2007. [Google Scholar]
- Kame-Ngasse, G.I.; Njiokou, F.; Melachio-Tanekou, T.T.; Farikou, O.; Simo, G.; Geiger, A. Prevalence of symbionts and trypanosome infections in tsetse flies of two villages of the “Faro and Déo” division of the Adamawa region of Cameroon. BMC Microbiol. 2018, 18 (Suppl. 1), 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farikou, O.; Njiokou, F.; Cuny, G.; Geiger, A. Microsatellite genotyping reveals diversity within populations of Sodalis glossinidius, the secondary symbiont of tsetse flies. Vet. Microbiol. 2011, 150, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, A.; Ravel, S.; Frutos, R.; Cuny, G. Sodalis glossinidius (Enterobacteriaceae) and vectorial competence of Glossina palpalis gambiensis and Glossina morsitans morsitans for Trypanosoma congolense savannah type. Curr. Microbiol. 2005, 51, 35–40. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Organism | Target Gene | Primer Name | Primer Sequence (5’ to 3’) | Amplicon Size (bp) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|---|---|
| Trypanosoma spp. | ITS1 rDNA | ITS1 CF | CCGGAAGTTCACCGATATTG | Variable | 58 | [30] |
| ITS1 BR | TTGCTGCGTTCTTCAACGAA | |||||
| T. congolense Kilifi | Satellite DNA monomer | TCK 1 | GTGCCCAAATTTGAAGTGAT | 294 | 55 | [33] |
| TCK 2 | ACTCAAAATCGTGCACCTCG | |||||
| T. congolense Forest | Satellite DNA monomer | TCF 1 | GGACACGCCAGAAGGTACTT | 350 | 55 | [33] |
| TCF 2 | GTTCTCGCACCAAATCCAAC | |||||
| T. congolense Savannah | Satellite DNA monomer | TCS 1 | CGAGAACGGGCACTTTGCGA | 316 | 55 | [33] |
| TCS 2 | GGACAAAGAAATCCCGCACA | |||||
| T.b. rhodesiense | SRA gene | SRA284 F | ATAGTGACAAGATGCGTACCAACGC | 284 | 68 | [34] |
| SRA284 R | AATGTGTTCGAGTACTTCGGTCACCT | |||||
| T. vivax | TVIV-F | CTGAGTGCTCCATGTCCCAC | 142 | 60 | [32] | |
| TVIV-R | CCACCAGAACACCAACCTGA | |||||
| T. brucei s. l. | TBR 1 | GAATATTAAACAATGCGCAG | 164 | 58 | [33] | |
| TBR 2 | CCATTTATTAGCTTTGTTGC | |||||
| T. simiae | TSM1 | CCGGTCAAAAACGCATT | 437 | 58 | [33] | |
| TSM2 | AGTCGCCCGGAGTCGAT | |||||
| T. simiae Tsavo | TST1 | GTCCTGCCACCGAGTATGC | 450 | 58 | [35] | |
| TST2 | CGAGCATGCAGGATGGCCG | |||||
| T. godfreyi | DGG1 | CTGAGGCTGAACAGCGACTC | 149 | 58 | [36] | |
| DGG2 | GGCGTATTGGCATAGCGTAC | |||||
| S. glossinidius | GPO1 | GPO1F | TGAGAGGTTCGTCAATGA | 1200 | 55 | [14] |
| GPO1R | ACGCTGCGTGACCATTC |
| Variable | Categories | n | Prevalence (%) | 95% CI | Statistical Analysis |
|---|---|---|---|---|---|
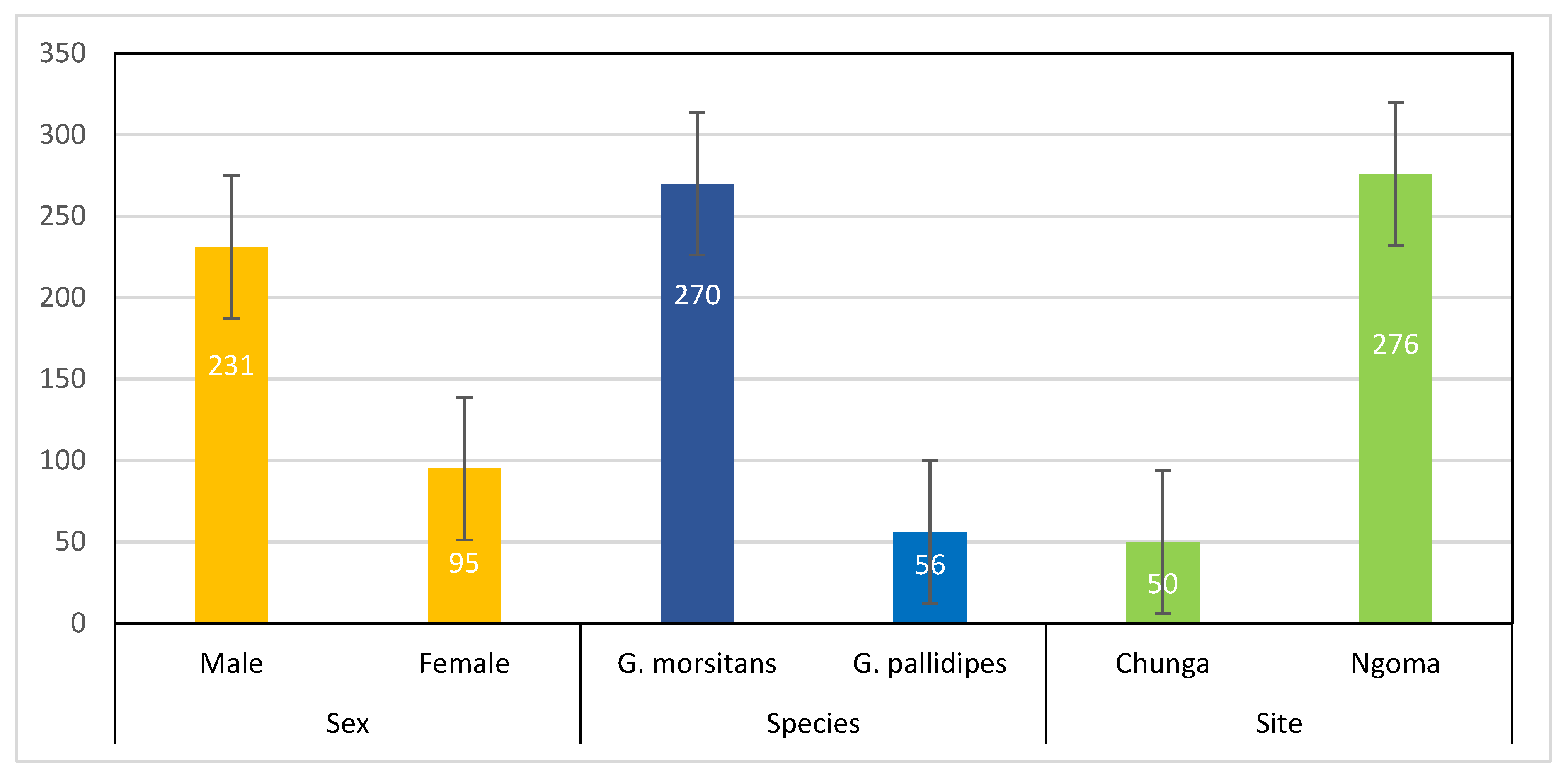
| Sampling site | Chunga | 50 | 6.0 | 2.06–16.22 | χ2 = 6.73, p = 0.009 * |
| Ngoma | 276 | 21.7 | 17.28–26.98 | ||
| Sex | Male | 231 | 20.4 | 15.66–26.00 | χ2 = 0.53, p = 0.467 |
| Female | 95 | 16.8 | 10.64–25.62 | ||
| Species | G. morsitans | 270 | 18.9 | 14.67–23.98 | χ2 = 0.19, p = 0.661 |
| G. pallidipes | 56 | 21.4 | 12.71–33.82 | ||
| G. morsitans | Male | 190 | 20.0 | 14.93–26.26 | χ2 = 0.30, p = 0.583 |
| Female | 80 | 16.3 | 9.75–25.84 | ||
| G. pallidipes | Male | 41 | 22.0 | 12.00–36.71 | p = 1.000 a |
| Female | 15 | 20.0 | 7.05–45.19 | ||
| Chunga | G. morsitans | 47 | 6.4 | 2.19–17.16 | p = 1.000 a |
| G. pallidipes | 3 | 0.0 | 0.00–56.15 | ||
| Ngoma | G. morsitans | 223 | 21.5 | 16.64–27.38 | χ2 = 0.03, p = 0.859 |
| G. pallidipes | 53 | 22.6 | 13.45–35.53 |
| Trypanosome Species | Overall (n = 326) | G. morsitans (n = 270) | G. pallidipes (n = 56) |
|---|---|---|---|
| T. congolense | 4.0% (2.35–6.70) | 4.4% (2.56–7.61) | 1.8% (0.32–9.45) |
| p = 0.705 a | |||
| T. vivax | 6.4% (4.25–9.65) | 6.3% (3.97–9.85) | 7.1% (2.81–16.98) |
| p = 0.768 a | |||
| T. b. brucei | 2.5% (1.25–4.77) | 1.9% (0.79–4.26) | 5.4% (1.84–14.61) |
| p = 0.142 a | |||
| T. simiae | 4.6% (2.81–7.45) | 4.8% (2.84–8.06) | 3.6% (0.98–12.12) |
| p = 1.000 a | |||
| T. godfreyi | 3.7% (2.12–6.32) | 3.3% (1.76–6.21) | 5.4% (1.84–14.61) |
| p = 0.440 a | |||
| T. s. Tsavo | 3.1% (1.67–5.55) | 3.7% (2.02–6.68) | 0.0% (0.0–6.42) |
| p = 0.221 a | |||
| Trypanosome Species | Sex | Sampling Site | ||
|---|---|---|---|---|
| Male (n = 231) | Female (n = 95) | Chunga (n = 50) | Ngoma (n = 276) | |
| T. congolense | 4.8% (2.68–8.32) | 2.1% (0.58–7.35) | 2.0% (0.35–10.50) | 4.4% (2.50–7.44) |
| p = 0.360 a | p = 0.700 a | |||
| T. vivax | 6.5% (3.97–10.44) | 6.3% (2.93–13.10) | 2.0% (0.35–10.50) | 7.3% (4.74–10.93) |
| X2 = 0.004, p = 0.953 | p = 0.220 a | |||
| T. b. brucei | 2.6% (1.20–5.55) | 2.1% (0.58–7.35) | 2.0% (0.35–10.50) | 2.5% (1.23–5.14) |
| p = 1.000 a | p = 1.000 a | |||
| T. simiae | 4.3% (2.37–7.78) | 5.3% (2.27–11.73) | 2.0% (0.35–10.50) | 5.1% (3.05–8.33) |
| p = 0.773 a | p = 0.483 a | |||
| T. godfreyi | 3.9% (2.06–7.24) | 3.2% (1.08–8.88) | 2.0% (0.35–10.50) | 4.0% (2.24–6.99) |
| p = 1.000 a | p = 0.700 a | |||
| T. s. Tsavo | 3.0% (1.48–6.12) | 3.2% (1.08–8.88) | 0.0% (0.0–7.13) | 3.6% (1.98–6.54) |
| p = 1.000 a | p = 0.371 a | |||
| Species | Location | n | Prevalence (95% CI) | |||||
|---|---|---|---|---|---|---|---|---|
| M | F | Total | M | F | Overall | p-Value | ||
| G. morsitans | Chunga | 22 | 25 | 47 | 27.3% (13.15–48.15) | 28.0% (14.28–47.58) | 27.7% (16.94–41.76) | χ2 = 0.97, p = 0.324 |
| Ngoma | 168 | 55 | 223 | 21.4% (15.90–28.24) | 20.0% (11.55–32.36) | 21.1% (16.24–26.90) | ||
| Total | 190 | 80 | 270 | 25.3% (19.62–31.89) | 22.5% (14.73–32.79) | 22.2% (17.67–27.55) | ||
| χ2 = 0.01, p = 0.943 | ||||||||
| G. pallidipes | Chunga | 2 | 1 | 3 | 0.0 | 0.0 | 0.0 | p = 1.000 a |
| Ngoma | 39 | 14 | 53 | 15.4% (7.25–29.73) | 35.7% (16.34–61.24) | 20.8% (12.00–33.46) | ||
| Total | 41 | 15 | 56 | 14.6% (6.88–28.44) | 33.3% (15.18–58.29) | 19.6% (11.34–31.84) | ||
| p = 0.142 a | ||||||||
| Overall | G. morsitans | G. pallidipes | Male | Female | Chunga | Ngoma | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T+ | T− | T+ | T− | T+ | T− | T+ | T− | T+ | T− | T+ | T− | T+ | T− | |
| S+ | 30 | 41 | 25 | 35 | 5 | 6 | 23 | 26 | 7 | 16 | 2 | 11 | 28 | 30 |
| S− | 33 | 222 | 26 | 184 | 7 | 38 | 24 | 159 | 9 | 63 | 1 | 36 | 32 | 190 |
| χ2 = 30.61, p < 0.001 | χ2 = 26.12, p < 0.001 | p = 0.045 a | χ2 = 28.42, p < 0.001 | p = 0.058 a | p = 0.162 a | χ2 = 30.39, p < 0.001 | ||||||||
| T. congolense | T. vivax | T. b. brucei | T. simiae | T. simiae Tsavo | T. godfreyi | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tc+ | Tc− | Tv+ | Tv− | Tbb+ | Tbb− | Ts+ | Ts− | Tst+ | Tst− | Tg+ | Tg− | |
| S+ | 4 | 67 | 10 | 61 | 4 | 67 | 7 | 64 | 6 | 65 | 6 | 66 |
| S− | 9 | 247 | 11 | 244 | 4 | 251 | 8 | 247 | 4 | 251 | 6 | 249 |
| p = 0.491 a | p = 0.006 a | p = 0.072 a | p = 0.025 a | p = 0.009 a | p = 0.027 a | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kallu, S.A.; Ndebe, J.; Qiu, Y.; Nakao, R.; Simuunza, M.C. Prevalence and Association of Trypanosomes and Sodalis glossinidius in Tsetse Flies from the Kafue National Park in Zambia. Trop. Med. Infect. Dis. 2023, 8, 80. https://doi.org/10.3390/tropicalmed8020080
Kallu SA, Ndebe J, Qiu Y, Nakao R, Simuunza MC. Prevalence and Association of Trypanosomes and Sodalis glossinidius in Tsetse Flies from the Kafue National Park in Zambia. Tropical Medicine and Infectious Disease. 2023; 8(2):80. https://doi.org/10.3390/tropicalmed8020080
Chicago/Turabian StyleKallu, Simegnew Adugna, Joseph Ndebe, Yongjin Qiu, Ryo Nakao, and Martin C. Simuunza. 2023. "Prevalence and Association of Trypanosomes and Sodalis glossinidius in Tsetse Flies from the Kafue National Park in Zambia" Tropical Medicine and Infectious Disease 8, no. 2: 80. https://doi.org/10.3390/tropicalmed8020080