
Evolution of Chemical Diversity in a Group of Non-Reduced Polyketide Gene Clusters: Using Phylogenetics to Inform the Search for Novel Fungal Natural Products



Abstract
:
1. Introduction

2. Group V NR-PKS Phylogeny


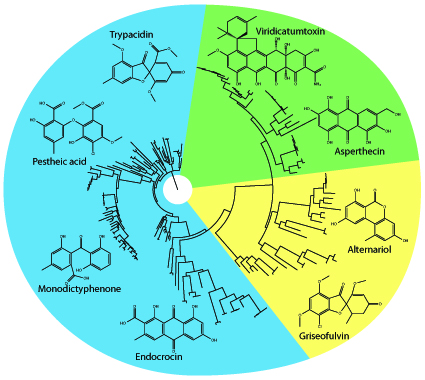{kind=link}
{kind=link}
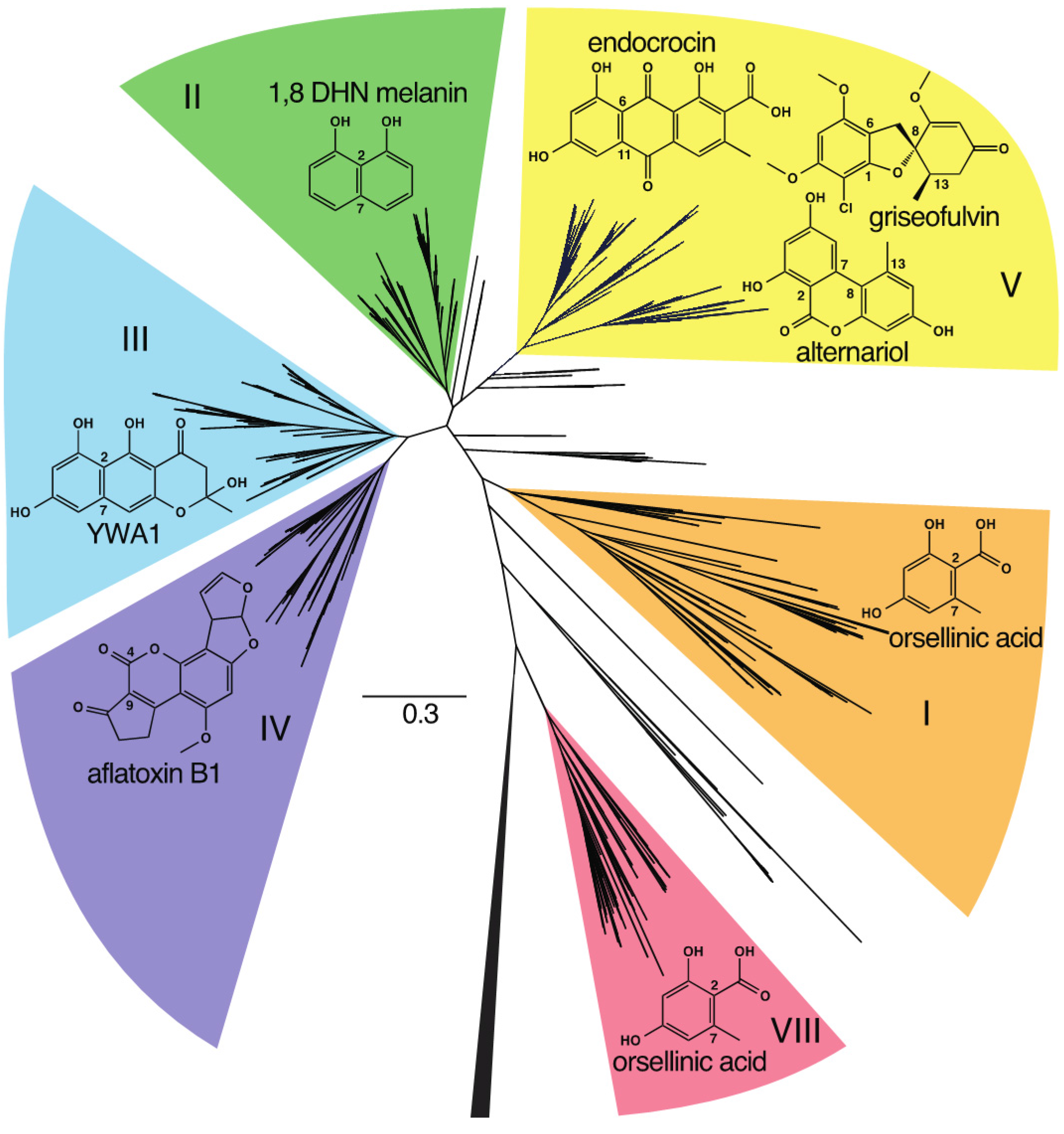{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Genus | Species | PKS | Sub-group | Characterized product | Predicted product | Reference |
|---|---|---|---|---|---|---|---|
| Dothidiomycete | Alternaria | alternata | AFN68300 | V3 | Alternariol | Characterized | [61] |
| Aureobasidium | melanogenum | KEQ66138 | V1 | None | Emodin anthrone | This study | |
| Bipolaris | maydis | EMD89515 | V3 | None | No data | This study | |
| ENH99769 | V3 | None | No data | This study | |||
| ENI07798 | V1 | None | Other anthrone | This study | |||
| AAR90273 | V1 | None | No data | This study | |||
| AAR90274 | V3 | None | No data | This study | |||
| oryzae | EUC41199 | V1 | None | Benzophenone | This study | ||
| EUC45162 | V1 | None | Other anthrone | This study | |||
| sorokiniana | EMD62925 | V1 | None | Other anthrone | This study | ||
| EMD66882 | V1 | None | Benzophenone | This study | |||
| victoriae | EUN25734 | V1 | None | Other anthrone | This study | ||
| zeicola | EUC29913 | V1 | None | Other anthrone | This study | ||
| Macrophomina | phaseolina | EKG11397 | V1 | None | No product | This study | |
| EKG18431 | V1 | None | No data | This study | |||
| Phaeosphaeria | nodorum | EAT76667 | V3 | Alternariol | Characterized | [62] | |
| Pseudocercospora | fijiensis | EME79056 | V1 | None | Emodin | This study | |
| Pseudogymnoascus | destructans | ELR08155 | V1 | None | Endocrocin anthrone | This study | |
| pannorum | KFY01830 | V1 | None | Endocrocin anthrone | This study | ||
| KFY04191 | V1 | None | Other anthrone | This study | |||
| KFY04767 | V1 | None | Endocrocin anthrone | This study | |||
| KFY14212 | V1 | None | Emodin anthrone | This study | |||
| KFY24933 | V1 | None | Endocrocin anthrone | This study | |||
| KFY28347 | V1 | None | Endocrocin anthrone | This study | |||
| KFY28376 | V1 | None | Endocrocin anthrone | This study | |||
| KFY41668 | V1 | None | Emodin anthrone | This study | |||
| KFY50141 | V1 | None | Grisandiene | This study | |||
| KFY61750 | V1 | None | Endocrocin anthrone | This study | |||
| KFY73941 | V1 | None | Endocrocin anthrone | This study | |||
| KFY81274 | V1 | None | Other anthrone | This study | |||
| KFY90954 | V1 | None | Other anthrone | This study | |||
| KFY97098 | V1 | None | Endocrocin anthrone | This study | |||
| KFZ02364 | V1 | None | Endocrocin anthrone | This study | |||
| KFZ03027 | V1 | None | Other anthrone | This study | |||
| KFZ03785 | V1 | None | Endocrocin anthrone | This study | |||
| KFZ09857 | V1 | None | Endocrocin anthrone | This study | |||
| Pyrenophora | teres | EFQ95560 | V1 | None | Endocrocin anthrone | This study | |
| tritici | EDU45231 | V1 | None | Endocrocin anthrone | This study | ||
| Setosphaeria | turcica | EOA88807 | V1 | None | Other anthrone | This study | |
| Sphaerulina | musiva | EMF17386 | V1 | None | Other anthrone | This study | |
| Verruconis | gallopava | KIW05310 | V1 | None | No product | This study | |
| Eurotiomycete | Arthroderma | benhamiae | EFE32713 | V2 | None | Neosartoricin, fumicycline | This study |
| otae | EEQ30779 | V2 | None | Neosartoricin, fumicycline | This study | ||
| EEQ31623 | V3 | None | Alternariol | This study | |||
| Aspergillus | acidus | Aspfo1_0068040 | V2 | None | TAN-1612 | This study | |
| Aspfo1_0069798 | V1 | None | No data | This study | |||
| aculeatus | Aacu16872_063282 | V1 | None | No data | This study | ||
| Aacu16872_063333 | V1 | None | No data | This study | |||
| brasiliensis | Aspbr1_0070836 | V1 | None | No data | This study | ||
| Aspbr1_0071307 | V2 | None | TAN-1612 | This study | |||
| clavatus | EAW13612 | V1 | None | No data | This study | ||
| flavus | EED53479 | V1 | None | No data | This study | ||
| KJJ30826 | V3 | None | No data | This study | |||
| fumigatus | EAL84397 | V1 | Endocrocin | Characterized | [47] | ||
| EAL84875 | V2 | Neosartoricin, fumicycline | Characterized | [49,63] | |||
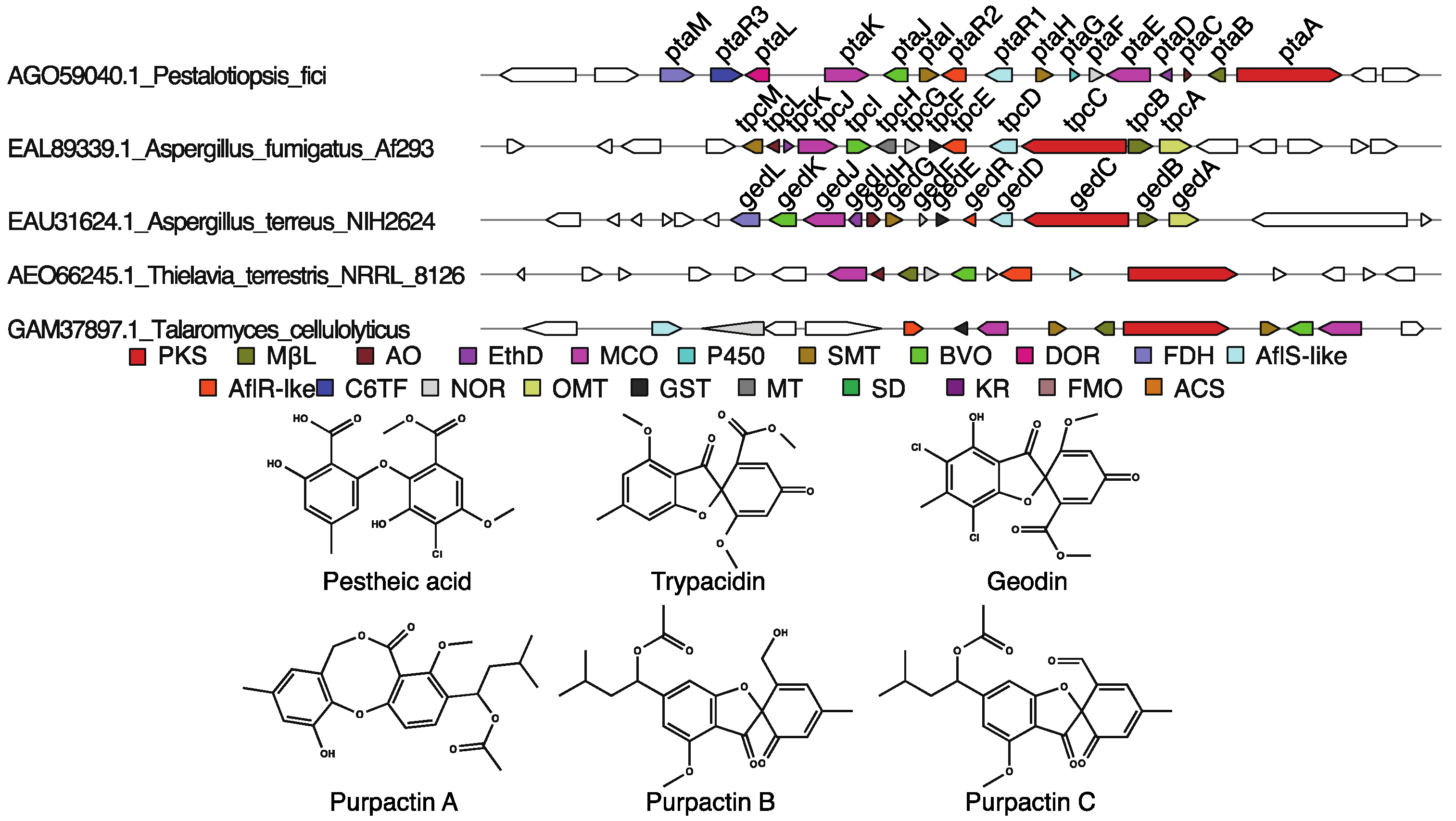
| EAL89339 | V1 | Trypacidin | Characterized | [53] | |||
| EDP47078 | V1 | Endocrocin | Characterized | [47] | |||
| EDP47964 | V2 | None | Neosartoricin, fumicycline | This study | |||
| EDP50840 | V1 | None | Trypacidin | This study | |||
| KEY78897 | V1 | None | Endocrocin | This study | |||
| KEY82310 | V1 | None | Other anthraquinone | This study | |||
| KEY82351 | V2 | None | Neosartoricin, fumicycline | This study | |||
| glaucus | Aspgl1_0045725 | V1 | None | No data | This study | ||
| kawachii | GAA85937 | V2 | None | TAN-1612 | This study | ||
| GAA88581 | V1 | None | No data | This study | |||
| nidulans | CBF70387 | V2 | Asperthecin | Characterized | [46] | ||
| CBF79143 | V3 | Alternariol | Characterized | [54] | |||
| CBF90097 | V1 | Monodictyphenone | Characterized | [51] | |||
| niger | EHA20150 | V1 | None | No data | This study | ||
| XP_001402309 | V1 | None | No product | This study | |||
| AEN83889 | V2 | TAN-1612 | Characterized | [64] | |||
| CAK40778 | V2 | None | TAN-1612 | This study | |||
| CAK47960 | V1 | None | No data | This study | |||
| ochraceoroseus | KKK15179 | V1 | None | Emodin | This study | ||
| KKK17199 | V1 | None | No data | This study | |||
| oryzae | BAE58990 | V1 | None | No data | This study | ||
| BAE62229 | V3 | None | Alternariol | This study | |||
| KDE80734 | V3 | None | Alternariol | This study | |||
| KDE81226 | V1 | None | No data | This study | |||
| XP_001820992 | V1 | None | No data | This study | |||
| parasiticus | KJK64046 | V3 | None | No data | This study | ||
| rambellii | KKK15908 | V1 | None | No data | This study | ||
| KKK27047 | V1 | None | Other anthraquinone | This study | |||
| ruber | EYE98259 | V1 | None | Endocrocin anthrone | This study | ||
| sydowii | Aspsy1_0090693 | V1 | None | No data | This study | ||
| Aspsy1_0144848 | V1 | None | No data | This study | |||
| Aspsy1_0151845 | V2 | None | Asperthecin | This study | |||
| Aspsy1_0157033 | V2 | None | Neosartoricin, fumicycline | This study | |||
| Aspsy1_1049255 | V1 | None | No data | This study | |||
| terreus | EAU31624 | V1 | Geodin | Characterized | [65,66] | ||
| EAU37396 | V1 | None | No data | This study | |||
| BAB88752 | V1 | None | No data | This study | |||
| tubingensis | Asptu1_0059858 | V2 | None | No data | This study | ||
| Asptu1_0123892 | V2 | None | TAN-1612 | This study | |||
| ustus | KIA75323 | V1 | None | Benzophenone | This study | ||
| KIA75530 | V2 | None | Asperthecin | This study | |||
| KIA75835 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | |||
| versicolor | Aspve1_0089706 | V2 | None | Neosartoricin, fumicycline | This study | ||
| Aspve1_0122449 | V1 | None | No data | This study | |||
| Aspve1_0886277 | V2 | None | Asperthecin | This study | |||
| wentii | Aspwe1_0034272 | V1 | None | No data | This study | ||
| zonatus | Aspzo1_2112764 | V1 | None | No data | This study | ||
| Capronia | epimyces | EXJ89638 | V1 | None | Other anthrone | This study | |
| Cladophialophora | carrionii | ETI19899 | V1 | None | Benzophenone | This study | |
| yegresii | EXJ61970 | V1 | None | Benzophenone | This study | ||
| Endocarpon | pusillum | ERF75912 | V1 | None | No product | This study | |
| Microsporum | gypseum | EFR03594 | V2 | None | Neosartoricin, fumicycline | This study | |
| Neosartorya | fischeri | EAW20700 | V2 | Neosartoricin, fumicycline | Characterized | [49,63] | |
| EAW24682 | V1 | None | Benzophenone | This study | |||
| EAW24697 | V1 | None | Grisandiene | This study | |||
| EAW25724 | V1 | None | No data | This study | |||
| Penicillium | aethiopicum | ADI24926 | V2 | Viridicatumtoxin | Characterized | [48] | |
| ADI24953 | V3 | Griseofulvin | Characterized | [48] | |||
| expansum | KGO43750 | V1 | None | Benzophenone | This study | ||
| KGO65172 | V1 | None | Emodin | This study | |||
| oxalicum | EPS34273 | V1 | None | Benzophenone | This study | ||
| Talaromyces | cellulolyticu | GAM33809 | V1 | None | No data | This study | |
| GAM37897 | V1 | None | Grisandiene | This study | |||
| GAM40075 | V3 | None | No data | This study | |||
| GAM42425 | V1 | None | No data | This study | |||
| GAM43179 | V1 | None | No data | This study | |||
| islandicus | CRG83532 | V1 | None | No data | This study | ||
| CRG86674 | V3 | None | No data | This study | |||
| CRG92129 | V1 | None | No data | This study | |||
| marneffei | KFX46552 | V1 | None | Grisandiene | This study | ||
| KFX52365 | V1 | None | Other anthrone | This study | |||
| ADH01670 | V1 | None | No data | This study | |||
| ADH01674 | V1 | None | No data | This study | |||
| stipitatus | EED18910 | V1 | None | Emodin anthrone | This study | ||
| EED18976 | V1 | None | Benzophenone | This study | |||
| Trichophyton | equinum | EGE06343 | V2 | None | Neosartoricin, fumicycline | This study | |
| interdigitale | KDB28089 | V2 | None | Neosartoricin, fumicycline | This study | ||
| rubrum | EZG11077 | V2 | None | Neosartoricin, fumicycline | This study | ||
| KDB38561 | V2 | None | Neosartoricin, fumicycline | This study | |||
| soudanense | EZF78756 | V2 | None | Neosartoricin, fumicycline | This study | ||
| tonsurans | EGD99348 | V2 | None | Neosartoricin, fumicycline | This study | ||
| verrucosum | EFE44835 | V2 | None | Neosartoricin, fumicycline | This study | ||
| Lecanoromycete | Usnea | longissima | AGI60157 | V1 | None | No data | This study |
| Leotiomycete | Blumeria | graminis | CCU75801 | V3 | None | No data | This study |
| EPQ66189 | V3 | None | No data | This study | |||
| Botrytis | cinerea | AAR90250 | V3 | None | No data | This study | |
| EMR83380 | V3 | None | No data | This study | |||
| XP_001553397 | V3 | None | No data | This study | |||
| Oidiodendron | maius | KIM92894 | V1 | None | Grisandiene | This study | |
| KIM93459 | V2 | None | No data | This study | |||
| KIM96903 | V2 | None | No data | This study | |||
| KIM99919 | V3 | None | No data | This study | |||
| Sclerotinia | borealis | ESZ98980 | V1 | None | Benzophenone | This study | |
| Sordariomycete | Claviceps | purpurea | CCE31584 | V1 | None | Benzophenone | This study |
| Colletotrichum | graminicola | EFQ33703 | V3 | None | Alternariol | This study | |
| sublineola | KDN60962 | V3 | None | No data | This study | ||
| KDN62802 | V3 | None | No product | This study | |||
| Diaporthe | ampelina | KKY32371 | V1 | None | Benzophenone | This study | |
| KKY34489 | V1 | None | Endocrocin anthrone | This study | |||
| KKY37364 | V1 | None | No product | This study | |||
| Eutypa | lata | EMR67234 | V1 | None | Benzophenone | This study | |
| Gaeumannomyces | graminis | EJT69423 | V3 | None | Alternariol | This study | |
| Grosmannia | clavigera | EFX02748 | V1 | None | Benzophenone | This study | |
| EFX04268 | V3 | None | No product | This study | |||
| Metarhizium | acridum | EFY89907 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | |
| album | KHN97625 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | ||
| anisopliae | KID64953 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | ||
| KID65337 | V1 | None | Grisandiene | This study | |||
| KJK89454 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | |||
| KJK95825 | V1 | None | Grisandiene | This study | |||
| brunneum | KID65403 | V1 | None | Grisandiene | This study | ||
| KID75074 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | |||
| guizhouense | KID84413 | V1 | None | Grisandiene | This study | ||
| KID87428 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | |||
| majus | KID93731 | V1 | None | Grisandiene | This study | ||
| KID97686 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | |||
| robertsii | EXU97310 | V2 | None | Viridicatumtoxin-like naphthacenedione | This study | ||
| EXV00352 | V1 | None | Grisandiene | This study | |||
| Myceliophthora | thermophila | AEO58356 | V1 | None | Benzophenone | This study | |
| Pestalotiopsis | fici | AGO59040 | V1 | Pestheic acid | Characterized | [52] | |
| ETS76950 | V1 | Pestheic acid | Characterized | This study | |||
| Phaeomoniella | chlamydospora | KKY14863 | V3 | None | No data | This study | |
| Stachybotrys | chartarum | KEY72288 | V1 | None | Benzophenone | This study | |
| KFA45466 | V3 | None | No data | This study | |||
| KFA54249 | V1 | None | Benzophenone | This study | |||
| KFA70942 | V3 | None | No data | This study | |||
| KFA71118 | V1 | None | Benzophenone | This study | |||
| Thielavia | terrestris | AEO66245 | V1 | None | Grisandiene | This study | |
| Trichoderma | atroviride | EHK46042 | V1 | None | No data | This study | |
| harzianum | KKO99957 | V1 | None | Other anthraquinone | This study | ||
| KKP03797 | V3 | None | No data | This study | |||
| reesei | ETR97748 | V3 | None | Alternariol | This study | ||
| virens | EHK20655 | V3 | None | Alternariol | This study |

3. Phylogenetics Directs Product Prediction
3.1. Group V1
3.1.1. Endocrocin-Like Clusters

3.1.2. Monodictyphenone-Like Clusters
3.1.3. Trypacidin, Geodin, and Pestheic Acid-Like Clusters

3.2. Group V2
3.2.1. Asperthecin-Like Clusters

3.2.2. TAN-1612-Like Clusters
3.2.3. Neosartoricin-Like Clusters
3.2.4. Viridicatumtoxin-Like Clusters
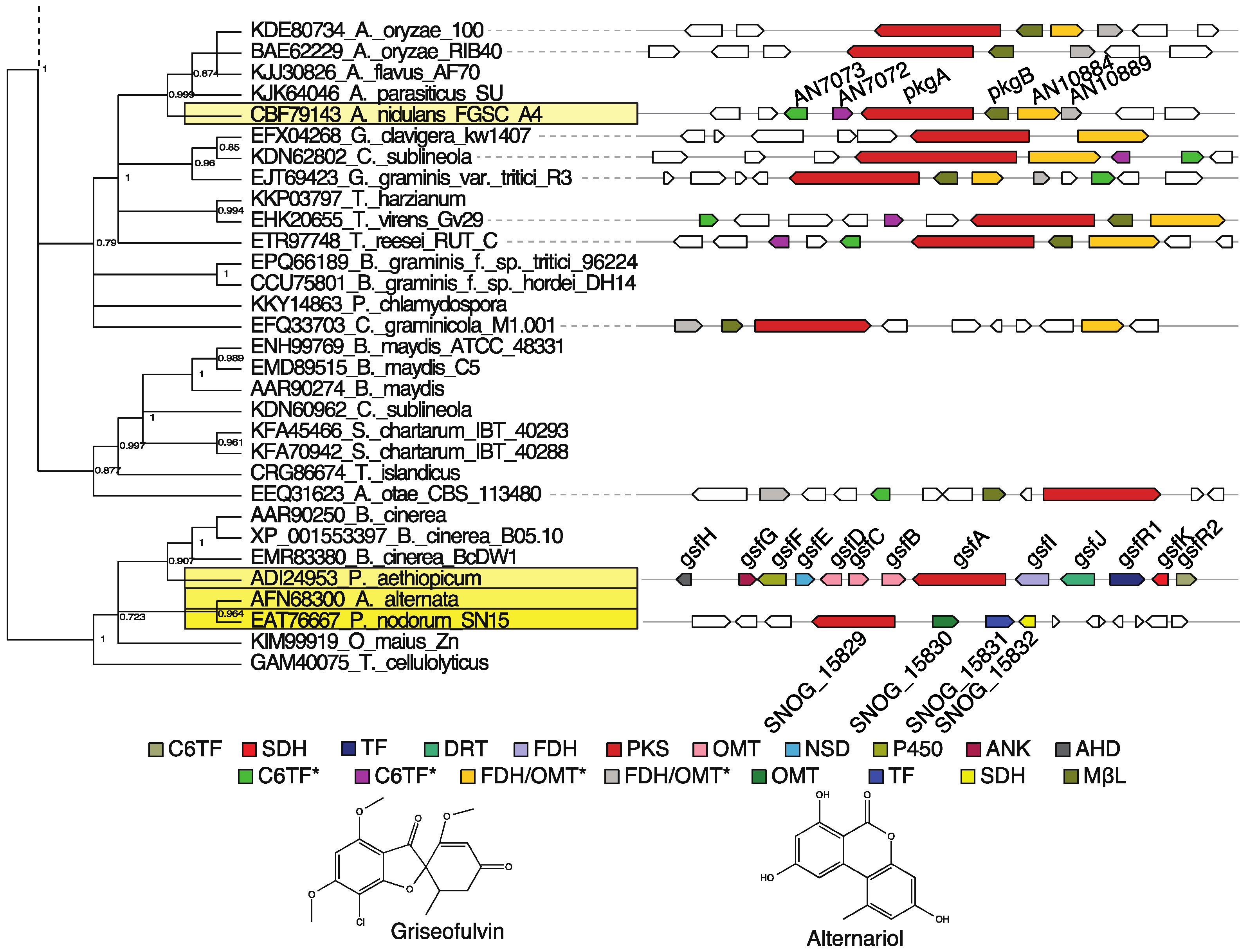
3.3. Group V3: Griseofulvin and Various Alternariol-Like Pathways

4. Uncharacterized Clusters in Symbionts and Pathogens
4.1. Plant Pathogens
4.2. Mycorrhizal Fungi
4.3. Fungal Pathogens
4.4. Animal Pathogens
5. Experimental Section
6. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Perez-Nadales, E.; Nogueira, M.F.; Baldin, C.; Castanheira, S.; el Ghalid, M.; Grund, E.; Lengeler, K.; Marchegiani, E.; Mehrotra, P.V.; Moretti, M.; et al. Fungal model systems and the elucidation of pathogenicity determinants. Fungal Genet. Biol. 2014, 70, 42–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohlfs, M.; Churchill, A.C. Fungal secondary metabolites as modulators of interactions with insects and other arthropods. Fungal Genet. Biol. 2011, 48, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Scherlach, K.; Graupner, K.; Hertweck, C. Molecular bacteria-fungi interactions: Effects on environment, food, and medicine. Annu. Rev. Microbiol. 2013, 67, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Allam, N.G.; Abd el-Zaher, E.H.F. Protective role of Aspergillus fumigatus melanin against ultraviolet (UV) irradiation and Bjerkandera adusta melanin as a candidate vaccine against systemic candidiasis. Afr. J. Biotech. 2012, 11, 6566–6577. [Google Scholar]
- Chiang, Y.M.; Oakley, B.R.; Keller, N.P.; Wang, C.C. Unraveling polyketide synthesis in members of the genus Aspergillus. Appl. Microbiol. Biotechnol. 2010, 86, 1719–1736. [Google Scholar] [CrossRef] [PubMed]
- Bushley, K.E.; Turgeon, B.G. Phylogenomics reveals subfamilies of fungal nonribosomal peptide synthetases and their evolutionary relationships. BMC Evol. Biol. 2010, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Wawrzyn, G.T.; Bloch, S.E.; Schmidt-Dannert, C. Discovery and characterization of terpenoid biosynthetic pathways of fungi. Methods Enzymol. 2012, 515, 83–105. [Google Scholar] [PubMed]
- Umemura, M.; Nagano, N.; Koike, H.; Kawano, J.; Ishii, T.; Miyamura, Y.; Kikuchi, M.; Tamano, K.; Yu, J.; Shin-ya, K.; et al. Characterization of the biosynthetic gene cluster for the ribosomally synthesized cyclic peptide ustiloxin B in Aspergillus flavus. Fungal Genet. Biol. 2014, 68, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, N.D.; Moktali, V.; Medema, M.H. Bioinformatics approaches and software for detection of secondary metabolic gene clusters. Methods Mol. Biol. 2012, 944, 23–45. [Google Scholar] [PubMed]
- Gallo, A.; Ferrara, M.; Perrone, G. Phylogenetic study of polyketide synthases and nonribosomal peptide synthetases involved in the biosynthesis of mycotoxins. Toxins 2013, 5, 717–742. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. Hmmer web server: 2015 update. Nucleic Acids Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Wang, I.K.; Reeves, C.; Gaucher, G.M. Isolation and sequencing of a genomic DNA clone containing the 3’ terminus of the 6-methylsalicylic acid polyketide synthetase gene of Penicillium urticae. Can. J. Microbiol. 1991, 37, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Fujii, I.; Ono, Y.; Tada, H.; Gomi, K.; Ebizuka, Y.; Sankawa, U. Cloning of the polyketide synthase gene atX from Aspergillus terreus and its identification as the 6-methylsalicylic acid synthase gene by heterologous expression. Mol. Gen. Genet. 1996, 253, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Ripka, S.; Siegner, A.; Schiltz, E.; Schweizer, E. The multifunctional 6-methylsalicylic acid synthase gene of Penicillium patulum. Its gene structure relative to that of other polyketide synthases. Eur. J. Biochem. 1990, 192, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Mayorga, M.E.; Timberlake, W.E. The developmentally regulated Aspergillus nidulans wA gene encodes a polypeptide homologous to polyketide and fatty acid synthases. Mol. Gen. Genet. 1992, 235, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chang, P.K.; Cary, J.W.; Wright, M.; Bhatnagar, D.; Cleveland, T.E.; Payne, G.A.; Linz, J.E. Comparative mapping of aflatoxin pathway gene clusters in Aspergillus parasiticus and Aspergillus flavus. Appl. Environ. Microbiol. 1995, 61, 2365–2371. [Google Scholar] [PubMed]
- Trail, F.; Mahanti, N.; Rarick, M.; Mehigh, R.; Liang, S.H.; Zhou, R.; Linz, J.E. Physical and transcriptional map of an aflatoxin gene cluster in Aspergillus parasiticus and functional disruption of a gene involved early in the aflatoxin pathway. Appl. Environ. Microbiol. 1995, 61, 2665–2673. [Google Scholar] [PubMed]
- Yu, J.H.; Leonard, T.J. Sterigmatocystin biosynthesis in Aspergillus nidulans requires a novel type I polyketide synthase. J. Bacteriol. 1995, 177, 4792–4800. [Google Scholar] [PubMed]
- Yang, G.; Rose, M.S.; Turgeon, B.G.; Yoder, O.C. A polyketide synthase is required for fungal virulence and production of the polyketide T-toxin. Plant Cell 1996, 8, 2139–2150. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.E.; Kroken, S.; Inderbitzin, P.; Asvarak, T.; Li, B.Y.; Shi, L.; Yoder, O.C.; Turgeon, B.G. Two polyketide synthase-encoding genes are required for biosynthesis of the polyketide virulence factor, T-toxin, by Cochliobolus heterostrophus. Mol. Plant Microbe Interact. 2006, 19, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sánchez, V.; Avalos, J.; Limón, M.C. Identification and regulation of fusA, the polyketide synthase gene responsible for fusarin production in Fusarium fujikuroi. Appl. Environ. Microbiol. 2012, 78, 7258–7266. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Cox, R.J.; Lazarus, C.M.; Simpson, T.J. Fusarin C biosynthesis in Fusarium moniliforme and Fusarium venenatum. ChemBioChem 2004, 5, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Gaffoor, I.; Trail, F. Characterization of two polyketide synthase genes involved in zearalenone biosynthesis in Gibberella zeae. Appl. Environ. Microbiol. 2006, 72, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.T.; Lee, Y.R.; Jin, J.; Han, K.H.; Kim, H.; Kim, J.C.; Lee, T.; Yun, S.H.; Lee, Y.W. Two different polyketide synthase genes are required for synthesis of zearalenone in Gibberella zeae. Mol. Microbiol. 2005, 58, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.H.; Desjardins, A.E.; Plattner, R.D.; Hohn, T.M. A polyketide synthase gene required for biosynthesis of fumonisin mycotoxins in Gibberella fujikuroi mating population A. Fungal Genet. Biol. 1999, 27, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Zhu, X.; Du, L. Developing a genetic system for functional manipulations of FUM1, a polyketide synthase gene for the biosynthesis of fumonisins in Fusarium verticillioides. FEMS Microbiol. Lett. 2005, 248, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Kinoshita, H.; Ishihara, S.; Sakai, K.; Nagai, S.; Nihira, T. Polyketide synthase gene responsible for citrinin biosynthesis in Monascus purpureus. Appl. Environ. Microbiol. 2005, 71, 3453–3457. [Google Scholar] [CrossRef] [PubMed]
- Awakawa, T.; Kaji, T.; Wakimoto, T.; Abe, I. A heptaketide naphthaldehyde produced by a polyketide synthase from Nectria haematococca. Bioorg. Med. Chem. Lett. 2012, 22, 4338–4340. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Harris-Coward, P.Y.; Ehrlich, K.C.; Di Mavungu, J.D.; Malysheva, S.V.; De Saeger, S.; Dowd, P.F.; Shantappa, S.; Martens, S.L.; Calvo, A.M. Functional characterization of a veA-dependent polyketide synthase gene in Aspergillus flavus necessary for the synthesis of asparasone, a sclerotium-specific pigment. Fungal Genet. Biol. 2014, 64, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Uka, V.; Han, Z.; Buyst, D.; Harris-Coward, P.Y.; Ehrlich, K.C.; Wei, Q.; Bhatnagar, D.; Dowd, P.F.; Martens, S.L.; et al. An Aspergillus flavus secondary metabolic gene cluster containing a hybrid PKS-NRPS is necessary for synthesis of the 2-pyridones, leporins. Fungal Genet. Biol. 2015, 81, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.; Nowrousian, M. The polyketide synthase gene pks4 is essential for sexual development and regulates fruiting body morphology in Sordaria macrospora. Fungal Genet. Biol. 2014, 68, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Nonaka, T.; Fujii, I. Fungal type III polyketide synthases. Nat. Prod. Rep. 2014, 31, 1306–1317. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Keller, N.; Oakley, B.R.; Wang, C.C. A gene cluster containing two fungal polyketide synthases encodes the biosynthetic pathway for a polyketide, asperfuranone, in Aspergillus nidulans. J. Am. Chem. Soc. 2009, 131, 2965–2970. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Butchko, R.A.; Baker, S.E.; Proctor, R.H. Phylogenomic and functional domain analysis of polyketide synthases in Fusarium. Fungal Biol. 2012, 116, 318–331. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.M.; Dancy, B.C.; Hill, E.A.; Udwary, D.W.; Townsend, C.A. Identification of a starter unit acyl-carrier protein transacylase domain in an iterative type I polyketide synthase. Proc. Natl. Acad. Sci. USA 2006, 103, 16728–16733. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, C.M.; Townsend, C.A. Initial characterization of a type I fatty acid synthase and polyketide synthase multienzyme complex NorS in the biosynthesis of aflatoxin B1. Chem. Biol. 2002, 9, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Qiao, K.; Gao, Z.; Meehan, M.J.; Li, J.W.; Zhao, X.; Dorrestein, P.C.; Vederas, J.C.; Tang, Y. Enzymatic synthesis of resorcylic acid lactones by cooperation of fungal iterative polyketide synthases involved in hypothemycin biosynthesis. J. Am. Chem. Soc. 2010, 132, 4530–4531. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sanchez, J.F.; Chiang, Y.M.; Oakley, B.R.; Wang, C.C. Rational domain swaps reveal insights about chain length control by ketosynthase domains in fungal nonreducing polyketide synthases. Org. Lett. 2014, 16, 1676–1679. [Google Scholar] [CrossRef] [PubMed]
- Crawford, J.M.; Thomas, P.M.; Scheerer, J.R.; Vagstad, A.L.; Kelleher, N.L.; Townsend, C.A. Deconstruction of iterative multidomain polyketide synthase function. Science 2008, 320, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Gerber, R.; Lou, L.; Du, L. A PLP-dependent polyketide chain releasing mechanism in the biosynthesis of mycotoxin fumonisins in Fusarium verticillioides. J. Am. Chem. Soc. 2009, 131, 3148–3149. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Chiang, Y.M.; Szewczyk, E.; Reyes-Dominguez, Y.; Davidson, A.D.; Sanchez, J.F.; Lo, H.C.; Watanabe, K.; Strauss, J.; Oakley, B.R.; et al. Chromatin-level regulation of biosynthetic gene clusters. Nat. Chem. Biol. 2009, 5, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Schroeckh, V.; Scherlach, K.; Nützmann, H.W.; Shelest, E.; Schmidt-Heck, W.; Schuemann, J.; Martin, K.; Hertweck, C.; Brakhage, A.A. Intimate bacterial-fungal interaction triggers biosynthesis of archetypal polyketides in Aspergillus nidulans. Proc. Natl. Acad. Sci. USA 2009, 106, 14558–14563. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.F.; Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Ahuja, M.; Elizabeth Oakley, C.; Woo Bok, J.; Keller, N.; Oakley, B.R.; Wang, C.C. Molecular genetic analysis of the orsellinic acid/F9775 gene cluster of Aspergillus nidulans. Mol. Biosyst. 2010, 6, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Fujii, I.; Watanabe, A.; Sankawa, U.; Ebizuka, Y. Identification of claisen cyclase domain in fungal polyketide synthase wA, a naphthopyrone synthase of Aspergillus nidulans. Chem. Biol. 2001, 8, 189–197. [Google Scholar] [CrossRef]
- Korman, T.P.; Crawford, J.M.; Labonte, J.W.; Newman, A.G.; Wong, J.; Townsend, C.A.; Tsai, S.C. Structure and function of an iterative polyketide synthase thioesterase domain catalyzing claisen cyclization in aflatoxin biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 6246–6251. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, E.; Chiang, Y.M.; Oakley, C.E.; Davidson, A.D.; Wang, C.C.; Oakley, B.R. Identification and characterization of the asperthecin gene cluster of Aspergillus nidulans. Appl. Environ. Microbiol. 2008, 74, 7607–7612. [Google Scholar] [CrossRef] [PubMed]
- Lim, F.Y.; Hou, Y.; Chen, Y.; Oh, J.H.; Lee, I.; Bugni, T.S.; Keller, N.P. Genome-based cluster deletion reveals an endocrocin biosynthetic pathway in Aspergillus fumigatus. Appl. Environ. Microbiol. 2012, 78, 4117–4125. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Cacho, R.; Tang, Y. Identification of the viridicatumtoxin and griseofulvin gene clusters from Penicillium aethiopicum. Chem. Biol. 2010, 17, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Fang, J.; Liu, H.; Filler, S.G.; Wang, P.; Tang, Y. Genome mining of a prenylated and immunosuppressive polyketide from pathogenic fungi. Org. Lett. 2013, 15, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Awakawa, T.; Yokota, K.; Funa, N.; Doi, F.; Mori, N.; Watanabe, H.; Horinouchi, S. Physically discrete beta-lactamase-type thioesterase catalyzes product release in atrochrysone synthesis by iterative type I polyketide synthase. Chem. Biol. 2009, 16, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Chiang, Y.M.; Szewczyk, E.; Davidson, A.D.; Entwistle, R.; Keller, N.P.; Wang, C.C.; Oakley, B.R. Characterization of the Aspergillus nidulans monodictyphenone gene cluster. Appl. Environ. Microbiol. 2010, 76, 2067–2074. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, L.; Zhang, F.; Wang, W.; Li, J.; Guo, L.; Che, Y.; Liu, G. Identification of the first diphenyl ether gene cluster for pestheic acid biosynthesis in plant endophyte Pestalotiopsis fici. ChemBioChem 2014, 15, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Throckmorton, K.; Lim, F.Y.; Kontoyiannis, D.; Zheng, W.; Keller, N.P. Redundant synthesis of a conidial polyketide by two distinct secondary metabolite clusters in Aspergillus fumigatus. Environ. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, M.; Chiang, Y.M.; Chang, S.L.; Praseuth, M.B.; Entwistle, R.; Sanchez, J.F.; Lo, H.C.; Yeh, H.H.; Oakley, B.R.; Wang, C.C. Illuminating the diversity of aromatic polyketide synthases in Aspergillus nidulans. J. Am. Chem. Soc. 2012, 134, 8212–8221. [Google Scholar] [CrossRef] [PubMed]
- Kroken, S.; Glass, N.L.; Taylor, J.W.; Yoder, O.C.; Turgeon, B.G. Phylogenomic analysis of type I polyketide synthase genes in pathogenic and saprobic ascomycetes. Proc. Natl. Acad. Sci. USA 2003, 100, 15670–15675. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Image, I.I.; Xu, W.; Image, I.; Tang, Y. Classification, prediction, and verification of the regioselectivity of fungal polyketide synthase product template domains. J. Biol. Chem. 2010, 285, 22764–22773. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zhang, Z.; Shao, C.L.; Wang, J.L.; Bai, H.; Wang, C.Y. Bioinformatical analysis of the sequences, structures and functions of fungal polyketide synthase product template domains. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Lackner, G.; Misiek, M.; Braesel, J.; Hoffmeister, D. Genome mining reveals the evolutionary origin and biosynthetic potential of basidiomycete polyketide synthases. Fungal Genet. Biol. 2012, 49, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Hansen, F.T.; Gardiner, D.M.; Lysøe, E.; Fuertes, P.R.; Tudzynski, B.; Wiemann, P.; Sondergaard, T.E.; Giese, H.; Brodersen, D.E.; Sørensen, J.L. An update to polyketide synthase and non-ribosomal synthetase genes and nomenclature in Fusarium. Fungal Genet. Biol. 2015, 75, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.; Fetzner, R.; Burkhardt, B.; Podlech, J.; Metzler, M.; Dang, H.; Lawrence, C.; Fischer, R. Identification of a polyketide synthase required for alternariol (AOH) and alternariol-9-methyl ether (AME) formation in Alternaria alternata. PLoS ONE 2012, 7, e40564. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Muria-Gonzalez, J.M.; Mead, O.L.; Solomon, P.S. Snpks19 encodes the polyketide synthase for alternariol mycotoxin biosynthesis in the wheat pathogen Parastagonospora nodorum. Appl. Environ. Microbiol. 2015. [Google Scholar] [CrossRef] [PubMed]
- König, C.C.; Scherlach, K.; Schroeckh, V.; Horn, F.; Nietzsche, S.; Brakhage, A.A.; Hertweck, C. Bacterium induces cryptic meroterpenoid pathway in the pathogenic fungus Aspergillus fumigatus. ChemBioChem 2013, 14, 938–942. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chooi, Y.H.; Sheng, Y.; Valentine, J.S.; Tang, Y. Comparative characterization of fungal anthracenone and naphthacenedione biosynthetic pathways reveals an α-hydroxylation-dependent claisen-like cyclization catalyzed by a dimanganese thioesterase. J. Am. Chem. Soc. 2011, 133, 15773–15785. [Google Scholar] [CrossRef] [PubMed]
- Couch, R.D.; Gaucher, G.M. Rational elimination of Aspergillus terreus sulochrin production. J. Biotechnol. 2004, 108, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.T.; Nielsen, J.B.; Anyaogu, D.C.; Holm, D.K.; Nielsen, K.F.; Larsen, T.O.; Mortensen, U.H. Heterologous reconstitution of the intact geodin gene cluster in Aspergillus nidulans through a simple and versatile pcr based approach. PLoS ONE 2013, 8, e72871. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.M.; Townsend, C.A. Ordering the reductive and cytochrome p450 oxidative steps in demethylsterigmatocystin formation yields general insights into the biosynthesis of aflatoxin and related fungal metabolites. J. Am. Chem. Soc. 2005, 127, 3724–3733. [Google Scholar] [CrossRef] [PubMed]
- Henry, K.M.; Townsend, C.A. Synthesis and fate of o-carboxybenzophenones in the biosynthesis of aflatoxin. J. Am. Chem. Soc. 2005, 127, 3300–3309. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Li, P.; Scharfenstein, L.; Chang, P.K. HypC, the anthrone oxidase involved in aflatoxin biosynthesis. Appl. Environ. Microbiol. 2010, 76, 3374–3377. [Google Scholar] [CrossRef] [PubMed]
- Cary, J.W.; Ehrlich, K.C.; Bland, J.M.; Montalbano, B.G. The aflatoxin biosynthesis cluster gene, aflX, encodes an oxidoreductase involved in conversion of versicolorin A to demethylsterigmatocystin. Appl. Environ. Microbiol. 2006, 72, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Montalbano, B.; Boué, S.M.; Bhatnagar, D. An aflatoxin biosynthesis cluster gene encodes a novel oxidase required for conversion of versicolorin A to sterigmatocystin. Appl. Environ. Microbiol. 2005, 71, 8963–8965. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.X.; Fujii, I.; Ebizuka, Y.; Gomi, K.; Sankawa, U. Molecular cloning and heterologous expression of the gene encoding dihydrogeodin oxidase, a multicopper blue enzyme from Aspergillus terreus. J. Biol. Chem. 1995, 270, 21495–21502. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Ramirez-Prado, J.H.; Jakobek, J.L.; Horn, B.W. Gene duplication, modularity and adaptation in the evolution of the aflatoxin gene cluster. BMC Evol. Biol. 2007, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Medema, M.H.; Takano, E.; Breitling, R. Detecting sequence homology at the gene cluster level with multigeneblast. Mol. Biol. Evol. 2013, 30, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Berthier, E.; Lim, F.Y.; Deng, Q.; Guo, C.J.; Kontoyiannis, D.P.; Wang, C.C.; Rindy, J.; Beebe, D.J.; Huttenlocher, A.; Keller, N.P. Low-volume toolbox for the discovery of immunosuppressive fungal secondary metabolites. PLoS Pathog. 2013, 9, e1003289. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, K.C.; Mack, B.M.; Wei, Q.; Li, P.; Roze, L.V.; Dazzo, F.; Cary, J.W.; Bhatnagar, D.; Linz, J.E. Association with AflR in endosomes reveals new functions for AflJ in aflatoxin biosynthesis. Toxins 2012, 4, 1582–1600. [Google Scholar] [CrossRef] [PubMed]
- Skory, C.D.; Chang, P.K.; Cary, J.; Linz, J.E. Isolation and characterization of a gene from aspergillus parasiticus associated with the conversion of versicolorin a to sterigmatocystin in aflatoxin biosynthesis. Appl. Environ. Microbiol. 1992, 58, 3527–3537. [Google Scholar] [PubMed]
- Simpson, T.J. Genetic and biosynthetic studies of the fungal prenylated xanthone shamixanthone and related metabolites in Aspergillus spp. Revisited. ChemBioChem 2012, 13, 1680–1688. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.F.; Entwistle, R.; Hung, J.H.; Yaegashi, J.; Jain, S.; Chiang, Y.M.; Wang, C.C.; Oakley, B.R. Genome-based deletion analysis reveals the prenyl xanthone biosynthesis pathway in Aspergillus nidulans. J. Am. Chem. Soc. 2011, 133, 4010–4017. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P.; Kantz, N.J.; Adams, T.H. Aspergillus nidulans verA is required for production of the mycotoxin sterigmatocystin. Appl. Environ. Microbiol. 1994, 60, 1444–1450. [Google Scholar] [PubMed]
- Studt, L.; Wiemann, P.; Kleigrewe, K.; Humpf, H.U.; Tudzynski, B. Biosynthesis of fusarubins accounts for pigmentation of Fusarium fujikuroi perithecia. Appl. Environ. Microbiol. 2012, 78, 4468–4480. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Willmann, A.; Straeten, M.; Kleigrewe, K.; Beyer, M.; Humpf, H.U.; Tudzynski, B. Biosynthesis of the red pigment bikaverin in Fusarium fujikuroi: Genes, their function and regulation. Mol. Microbiol. 2009, 72, 931–946. [Google Scholar] [CrossRef] [PubMed]
- Pi, B.; Yu, D.; Dai, F.; Song, X.; Zhu, C.; Li, H.; Yu, Y. A genomics based discovery of secondary metabolite biosynthetic gene clusters in Aspergillus ustus. PLoS ONE 2015, 10, e0116089. [Google Scholar] [CrossRef] [PubMed]
- Cacho, R.A.; Chooi, Y.H.; Zhou, H.; Tang, Y. Complexity generation in fungal polyketide biosynthesis: A spirocycle-forming P450 in the concise pathway to the antifungal drug griseofulvin. ACS Chem. Biol. 2013, 8, 2322–2330. [Google Scholar] [CrossRef] [PubMed]
- Liu, D. Myceliophthora and Thielavia. In Molecular Detection of Human Fungal Pathogens; Liu, D., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Boca Raton, FL, USA, 2011; pp. 445–448. [Google Scholar]
- Gauthier, T.; Wang, X.; Sifuentes Dos Santos, J.; Fysikopoulos, A.; Tadrist, S.; Canlet, C.; Artigot, M.P.; Loiseau, N.; Oswald, I.P.; Puel, O. Trypacidin, a spore-borne toxin from Aspergillus fumigatus, is cytotoxic to lung cells. PLoS ONE 2012, 7, e29906. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeboya, M.O.; Edwards, R.L.; Lassoe, T.; Maitland, D.J.; Shields, L.; Whalley, A.J.S. Metabolites of the higher fungi. Part 29. Maldoxin, maldoxone, dihydromaldoxin, isodihydromaldoxin and dechlorodihydromaldoxin. A spirocyclohexadienone, a depsidone and three diphenyl ethers: Keys in the depsidone biosynthetic pathway from a member of the fungus genus Xylaria. J. Chem. Soc. 1996, 12, 1419–1425. [Google Scholar]
- Chooi, Y.H.; Wang, P.; Fang, J.; Li, Y.; Wu, K.; Tang, Y. Discovery and characterization of a group of fungal polycyclic polyketide prenyltransferases. J. Am. Chem. Soc. 2012, 134, 9428–9437. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.H.; Tang, Y. Navigating the fungal polyketide chemical space: From genes to molecules. J. Org. Chem. 2012, 77, 9933–9953. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Guo, C.J.; Palmer, J.M.; Sekonyela, R.; Wang, C.C.; Keller, N.P. Prototype of an intertwined secondary-metabolite supercluster. Proc. Natl. Acad. Sci. USA 2013, 110, 17065–17070. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.M.; Donzelli, B.G.; Krasnoff, S.B.; Keyhani, N.O. Discovering the secondary metabolite potential encoded within entomopathogenic fungi. Nat. Prod. Rep. 2014, 31, 1287–1305. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.R.; Nielsen, J.B.; Klitgaard, A.; Petersen, L.M.; Zachariasen, M.; Hansen, T.J.; Blicher, L.H.; Gotfredsen, C.H.; Larsen, T.O.; Nielsen, K.F.; et al. Accurate prediction of secondary metabolite gene clusters in filamentous fungi. Proc. Natl. Acad. Sci. USA 2013, 110, E99–E107. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, E.; Amichai, B.; Grunwald, M.H. Griseofulvin and its uses. Int. J. Antimicrob. Agents 1996, 6, 189–194. [Google Scholar] [CrossRef]
- Aggarwal, N.; Goindi, S. Preparation and in vivo evaluation of solid lipid nanoparticles of griseofulvin for dermal use. J. Biomed. Nanotechnol. 2013, 9, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Margarido, L.A.C. Oral treatments for fungal infections of the skin of the foot. Sao Paulo Med. J. 2014, 132, CD003584. [Google Scholar]
- Cox, R. Oxidative rearrangements during fungal biosynthesis. Nat. Prod. Rep. 2014, 31, 1405–1424. [Google Scholar] [CrossRef] [PubMed]
- Wicker, T.; Oberhaensli, S.; Parlange, F.; Buchmann, J.P.; Shatalina, M.; Roffler, S.; Ben-David, R.; Doležel, J.; Šimková, H.; Schulze-Lefert, P.; et al. The wheat powdery mildew genome shows the unique evolution of an obligate biotroph. Nat. Genet. 2013, 45, 1092–1096. [Google Scholar] [CrossRef] [PubMed]
- Duplessis, S.; Cuomo, C.A.; Lin, Y.C.; Aerts, A.; Tisserant, E.; Veneault-Fourrey, C.; Joly, D.L.; Hacquard, S.; Amselem, J.; Cantarel, B.L.; et al. Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. USA 2011, 108, 9166–9171. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Young, C.A.; Hesse, U.; Amyotte, S.G.; Andreeva, K.; Calie, P.J.; Fleetwood, D.J.; Haws, D.C.; Moore, N.; Oeser, B.; et al. Plant-symbiotic fungi as chemical engineers: Multi-genome analysis of the Clavicipitaceae reveals dynamics of alkaloid loci. PLoS Genet. 2013, 9, e1003323. [Google Scholar] [CrossRef] [PubMed]
- Khouja, H.R.; Daghino, S.; Abbà, S.; Boutaraa, F.; Chalot, M.; Blaudez, D.; Martino, E.; Perotto, S. OmGOGAT-disruption in the ericoid mycorrhizal fungus Oidiodendron maius induces reorganization of the n pathway and reduces tolerance to heavy-metals. Fungal Genet. Biol. 2014, 71, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Elmore, M.H.; McGary, K.L.; Wisecaver, J.H.; Slot, J.C.; Geiser, D.M.; Sink, S.; O’Donnell, K.; Rokas, A. Clustering of two genes putatively involved in cyanate detoxification evolved recently and independently in multiple fungal lineages. Genome Biol. Evol. 2015, 7, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ying, S.H.; Li, J.G.; Tian, C.G.; Feng, M.G. Insight into the transcriptional regulation of Msn2 required for conidiation, multi-stress responses and virulence of two entomopathogenic fungi. Fungal Genet. Biol. 2013, 54, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Punya, J.; Swangmaneecharern, P.; Pinsupa, S.; Nitistaporn, P.; Phonghanpot, S.; Kunathigan, V.; Cheevadhanarak, S.; Tanticharoen, M.; Amnuaykanjanasin, A. Phylogeny of type I polyketide synthases (PKSs) in fungal entomopathogens and expression analysis of PKS genes in Beauveria bassiana BCC 2660. Fungal Biol. 2015, 119, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Chibucos, M.C.; Crabtree, J.; Nagaraj, S.; Chaturvedi, S.; Chaturvedi, V. Draft genome sequences of human pathogenic fungus Geomyces pannorum sensu lato and bat white nose syndrome pathogen Geomyces (Pseudogymnoascus) destructans. Genome Announc. 2013, 1. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Yadav, G.; Gokhale, R.S.; Mohanty, D. Towards prediction of metabolic products of polyketide synthases: An in silico analysis. PLoS Comput. Biol. 2009, 5, e1000351. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. Trimal: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. Figtree. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 6 April 2015).
- Bendele, A.M.; Carlton, W.W.; Nelson, G.E.; Peterson, R.E.; Grove, M.D. Viridicatumtoxin mycotoxicosis in mice and rats. Toxicol. Lett. 1984, 22, 287–291. [Google Scholar] [CrossRef]
- Hutchison, R.D.; Steyn, P.S.; Van Rensburg, S.J. Viridicatumtoxin, a new mycotoxin from Penicillium viridicatum westling. Toxicol. Appl. Pharmacol. 1973, 24, 507–509. [Google Scholar] [CrossRef]
- Pero, R.W.; Posner, H.; Blois, M.; Harvan, D.; Spalding, J.W. Toxicity of metabolites produced by the “Alternaria”. Environ. Health Perspect. 1973, 4, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Demuner, A.J.; Almeida Barbosa, L.C.; Mendes Miranda, A.C.; Geraldo, G.C.; da Silva, C.M.; Giberti, S.; Bertazzini, M.; Forlani, G. The fungal phytotoxin alternariol 9-methyl ether and some of its synthetic analogues inhibit the photosynthetic electron transport chain. J. Nat. Prod. 2013, 76, 2234–2245. [Google Scholar] [CrossRef] [PubMed]
- Esbelin, J.; Mallea, S.; Ram, A.F.; Carlin, F. Role of pigmentation in protecting Aspergillus niger conidiospores against pulsed light radiation. Photochem. Photobiol. 2013, 89, 758–761. [Google Scholar] [CrossRef] [PubMed]
- Romero-Martinez, R.; Wheeler, M.; Guerrero-Plata, A.; Rico, G.; Torres-Guerrero, H. Biosynthesis and functions of melanin in Sporothrix schenckii. Infect. Immun. 2000, 68, 3696–3703. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Casadevall, A. Decreased susceptibility of melanized Cryptococcus neoformans to UV light. Appl. Environ. Microbiol. 1994, 60, 3864–3866. [Google Scholar] [PubMed]
- Tsai, H.F.; Chang, Y.C.; Washburn, R.G.; Wheeler, M.H.; Kwon-Chung, K.J. The developmentally regulated alb1 gene of Aspergillus fumigatus: Its role in modulation of conidial morphology and virulence. J. Bacteriol. 1998, 180, 3031–3038. [Google Scholar] [PubMed]
- Fourie, P.; Schutte, T.; Serfontein, S.; Swart, F. Modeling the effect of temperature and wetness on Guignardia pseudothecium maturation and ascospore release in citrus orchards. Phytopathology 2013, 103, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Guenther, J.C.; Trail, F. The development and differentiation of Gibberella zeae (anamorph: Fusarium graminearum) during colonization of wheat. Mycologia 2005, 97, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Salvo, J.J. Isolation and characterization of sexual spore pigments from Aspergillus nidulans. Appl. Environ. Microbiol. 1994, 60, 979–983. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Throckmorton, K.; Wiemann, P.; Keller, N.P. Evolution of Chemical Diversity in a Group of Non-Reduced Polyketide Gene Clusters: Using Phylogenetics to Inform the Search for Novel Fungal Natural Products. Toxins 2015, 7, 3572-3607. https://doi.org/10.3390/toxins7093572
Throckmorton K, Wiemann P, Keller NP. Evolution of Chemical Diversity in a Group of Non-Reduced Polyketide Gene Clusters: Using Phylogenetics to Inform the Search for Novel Fungal Natural Products. Toxins. 2015; 7(9):3572-3607. https://doi.org/10.3390/toxins7093572
Chicago/Turabian StyleThrockmorton, Kurt, Philipp Wiemann, and Nancy P. Keller. 2015. "Evolution of Chemical Diversity in a Group of Non-Reduced Polyketide Gene Clusters: Using Phylogenetics to Inform the Search for Novel Fungal Natural Products" Toxins 7, no. 9: 3572-3607. https://doi.org/10.3390/toxins7093572