Deciphering the Hazardous Effects of AFB1 and T-2 Toxins: Unveiling Toxicity and Oxidative Stress Mechanisms in PK15 Cells and Mouse Kidneys


Abstract
:1. Introduction
2. Results
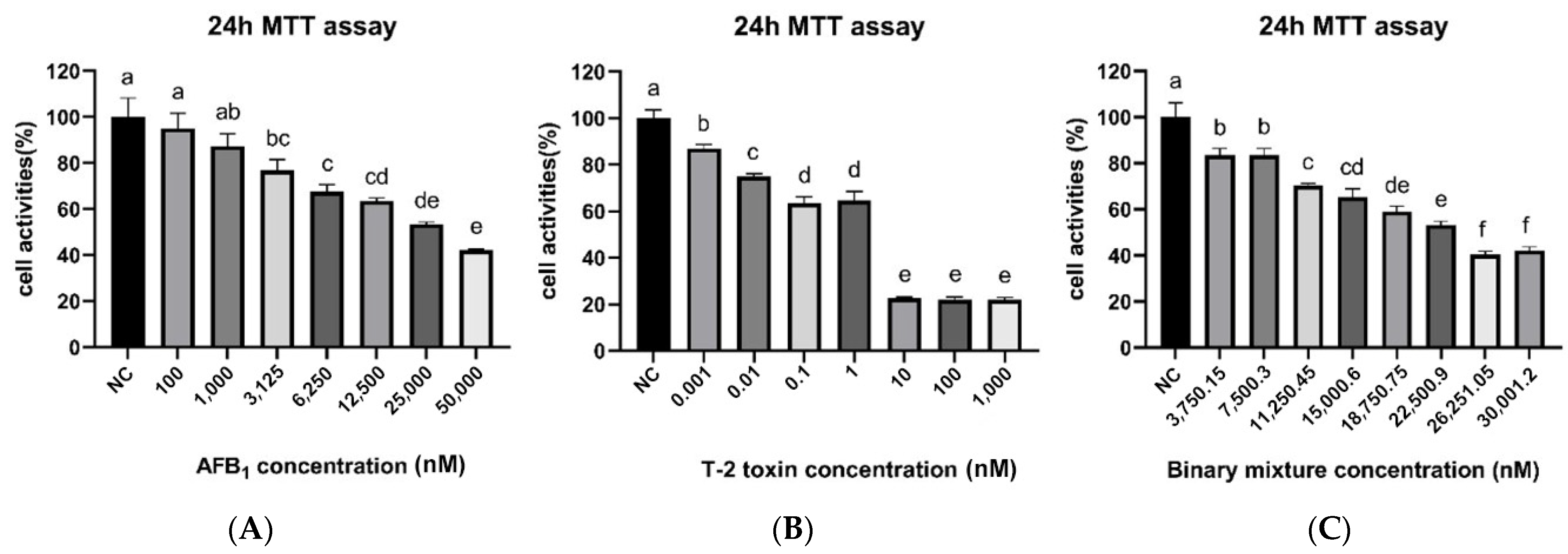
2.1. Cellular Activity and IC50
2.2. Cellular Toxicity Induced by the Synergistic Action of AFB1 and T-2
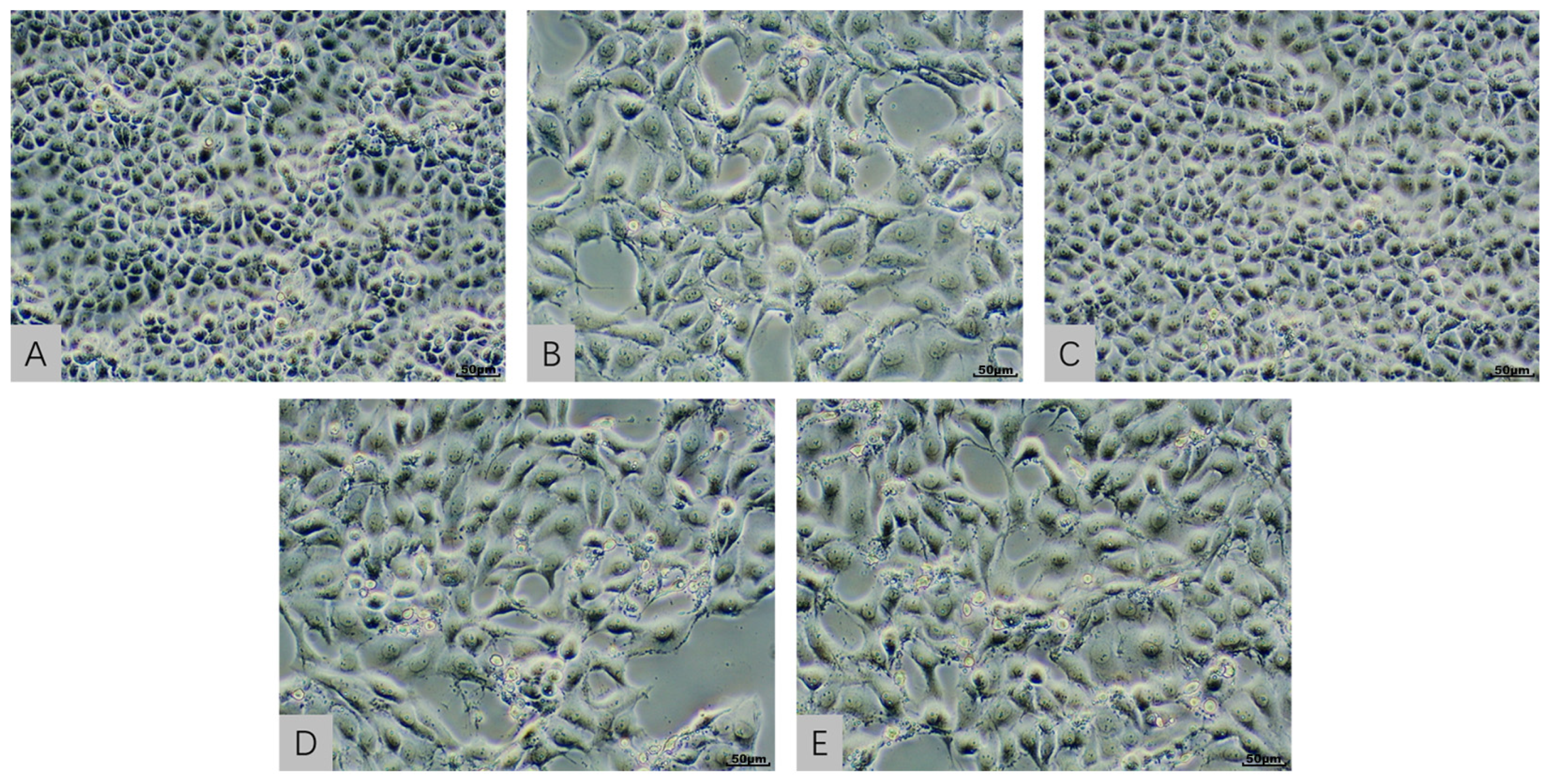
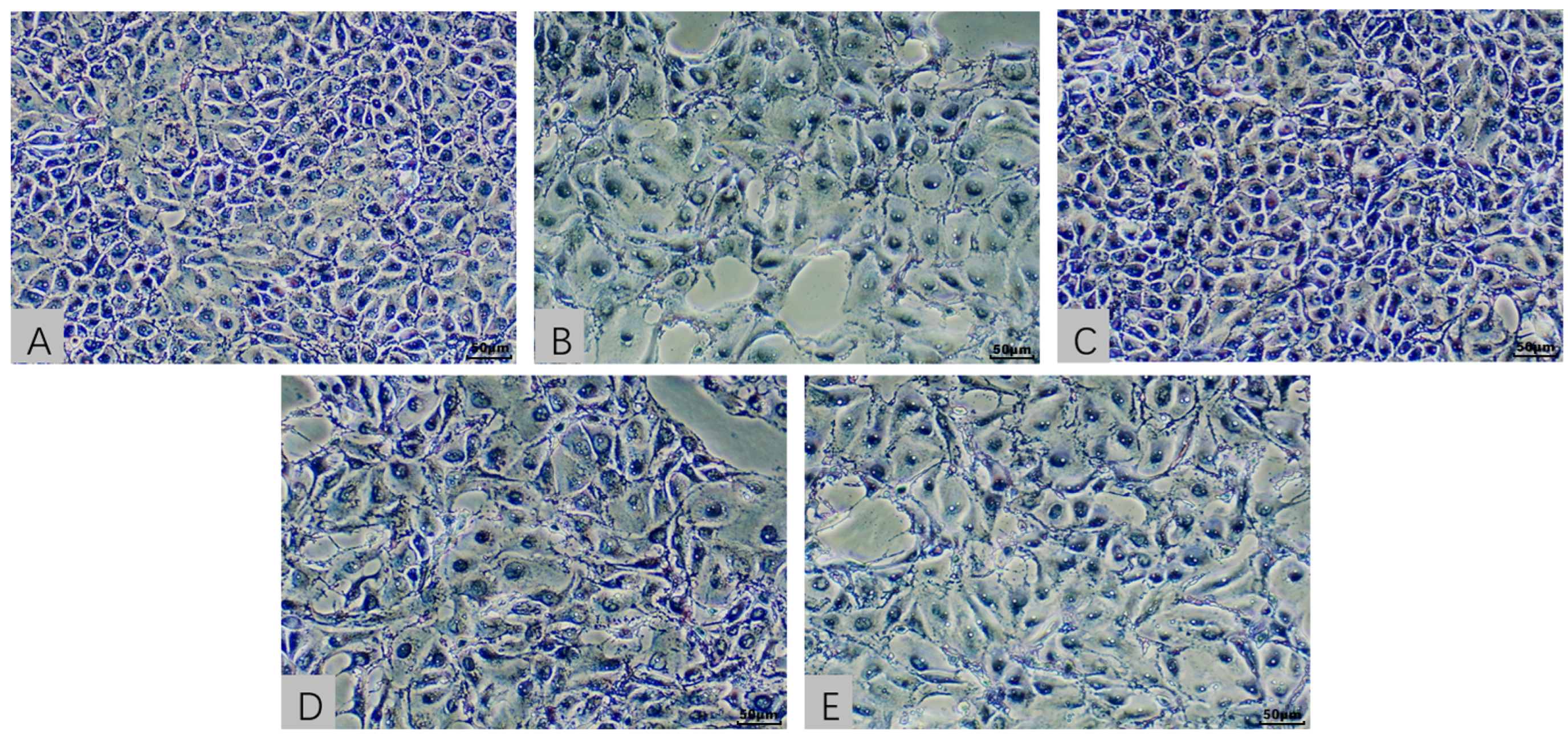
2.3. Cellular Morphological Evaluation in an In Vitro Model Exposed to AFB1 and T-2 Alone and in Combination
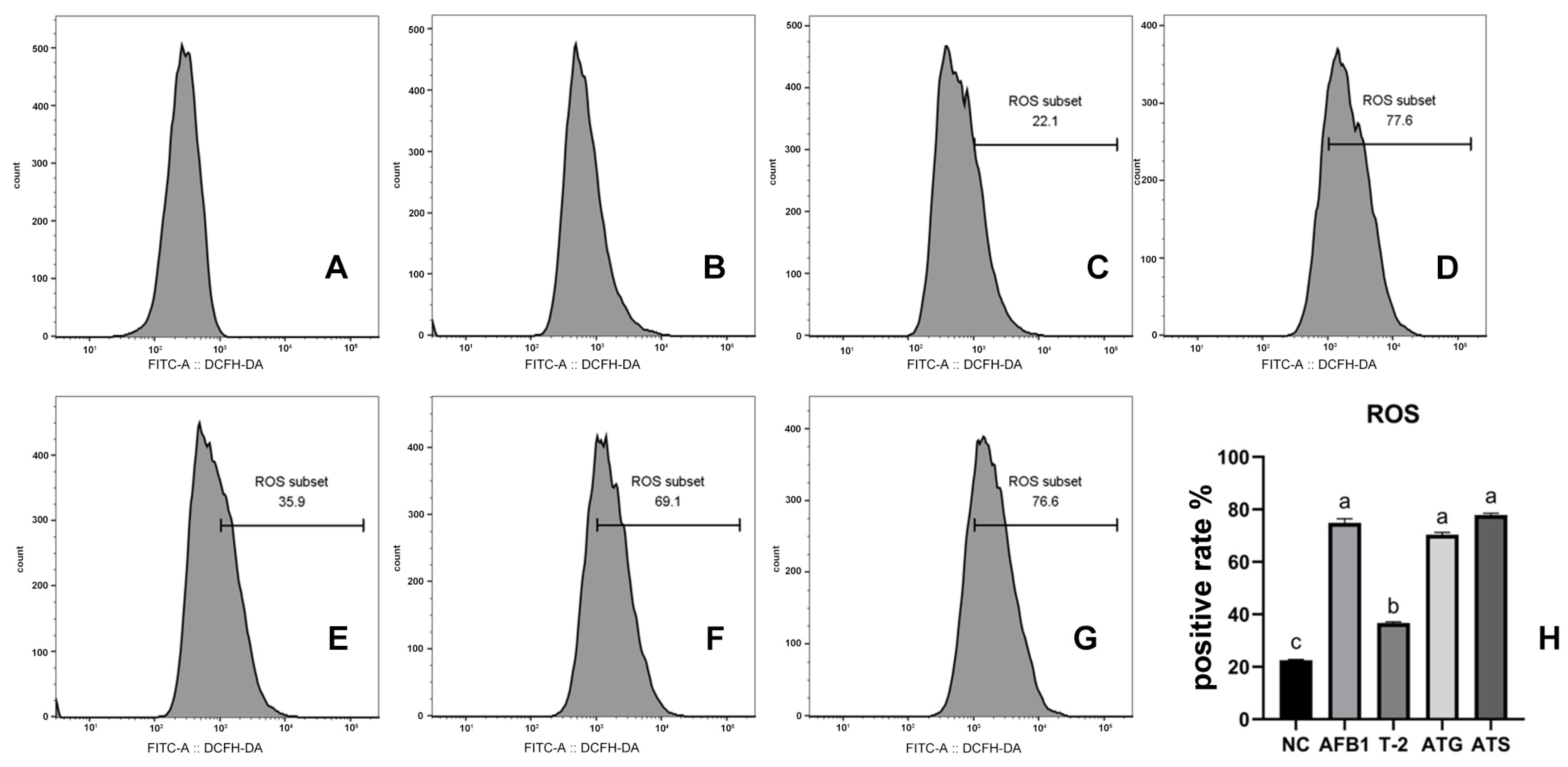
2.4. Quantification of ROS in Cells Exposed to Toxins
2.5. Assessment of Intracellular MDA, GSH, and GSH-Px Levels in Toxin-Treated Cells
2.6. Evaluation of Oxidative Stress-Related mRNA Expression in Cells
2.7. Body Weight, Weight Gain, and Food Intake
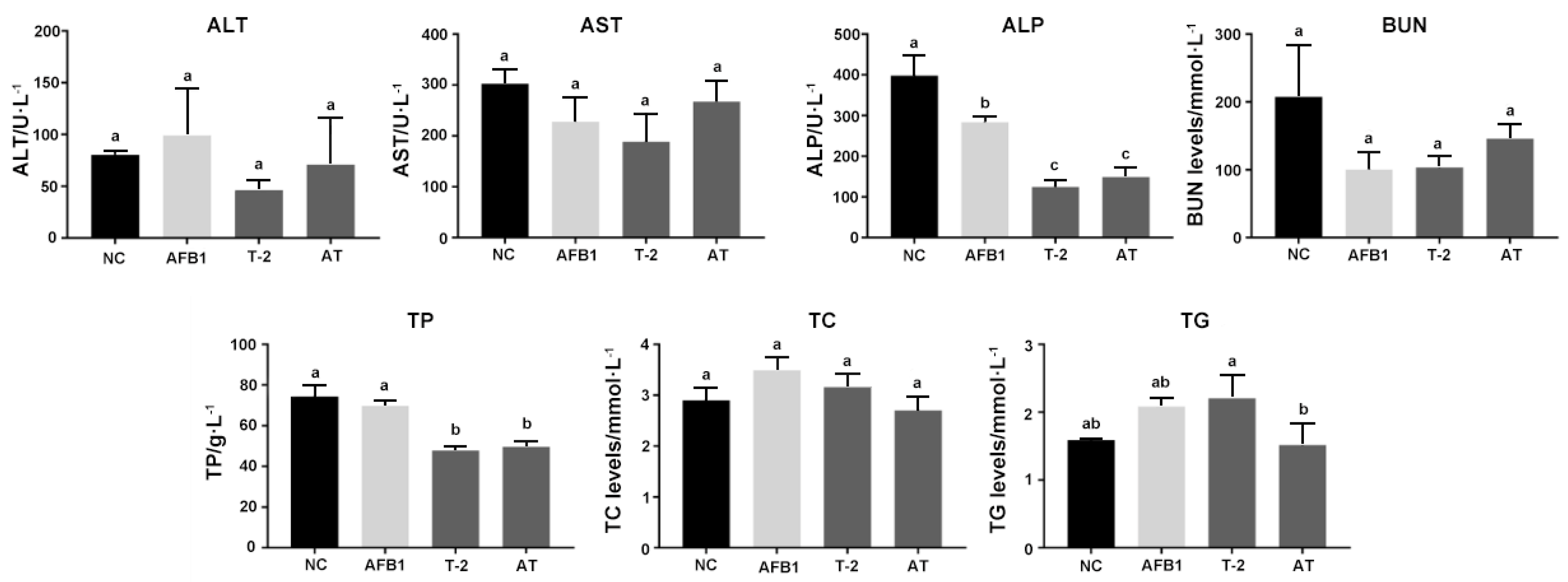
2.8. Serum Biochemistry Analysis Parameters
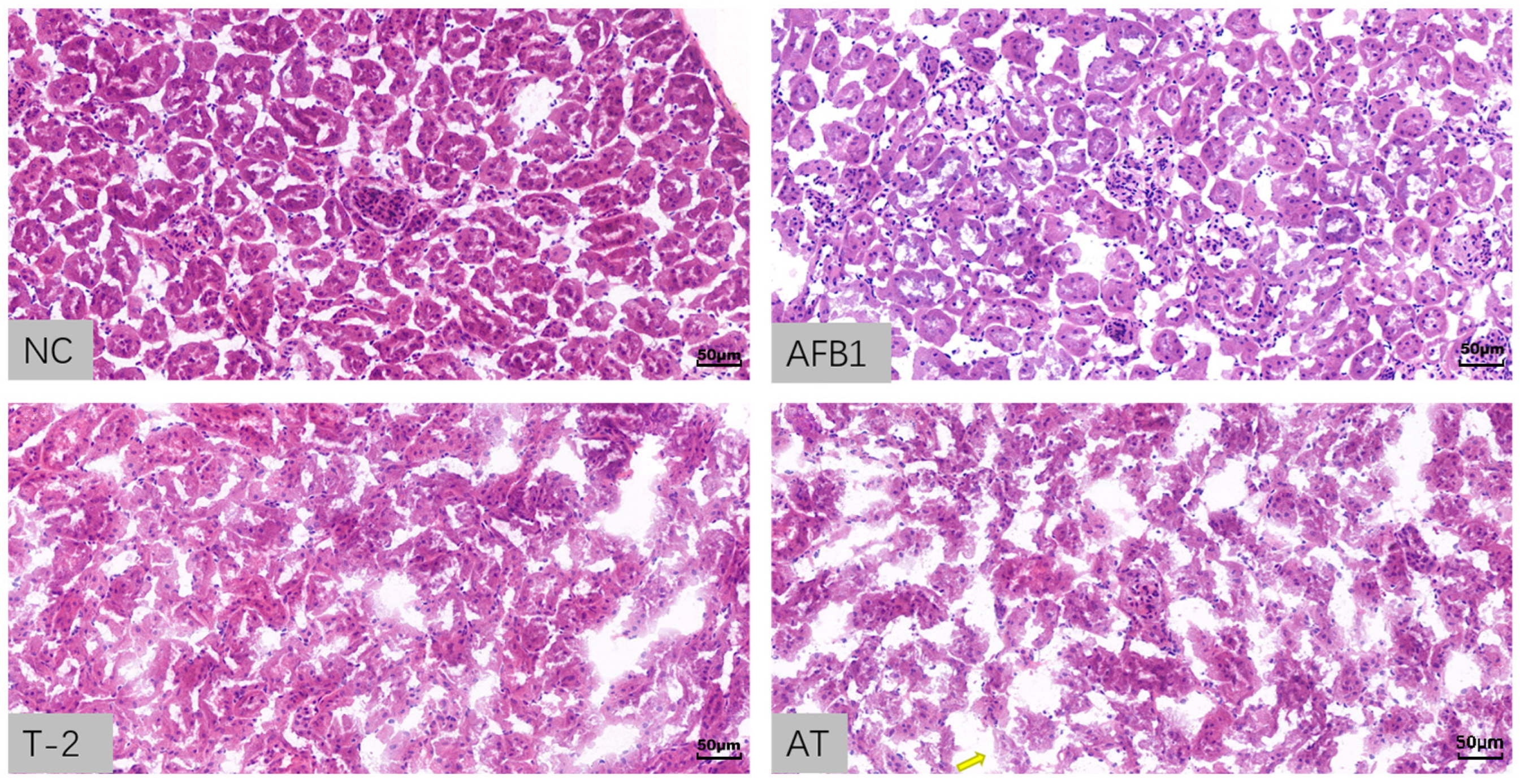
2.9. Histopathological Analysis of Kidneys
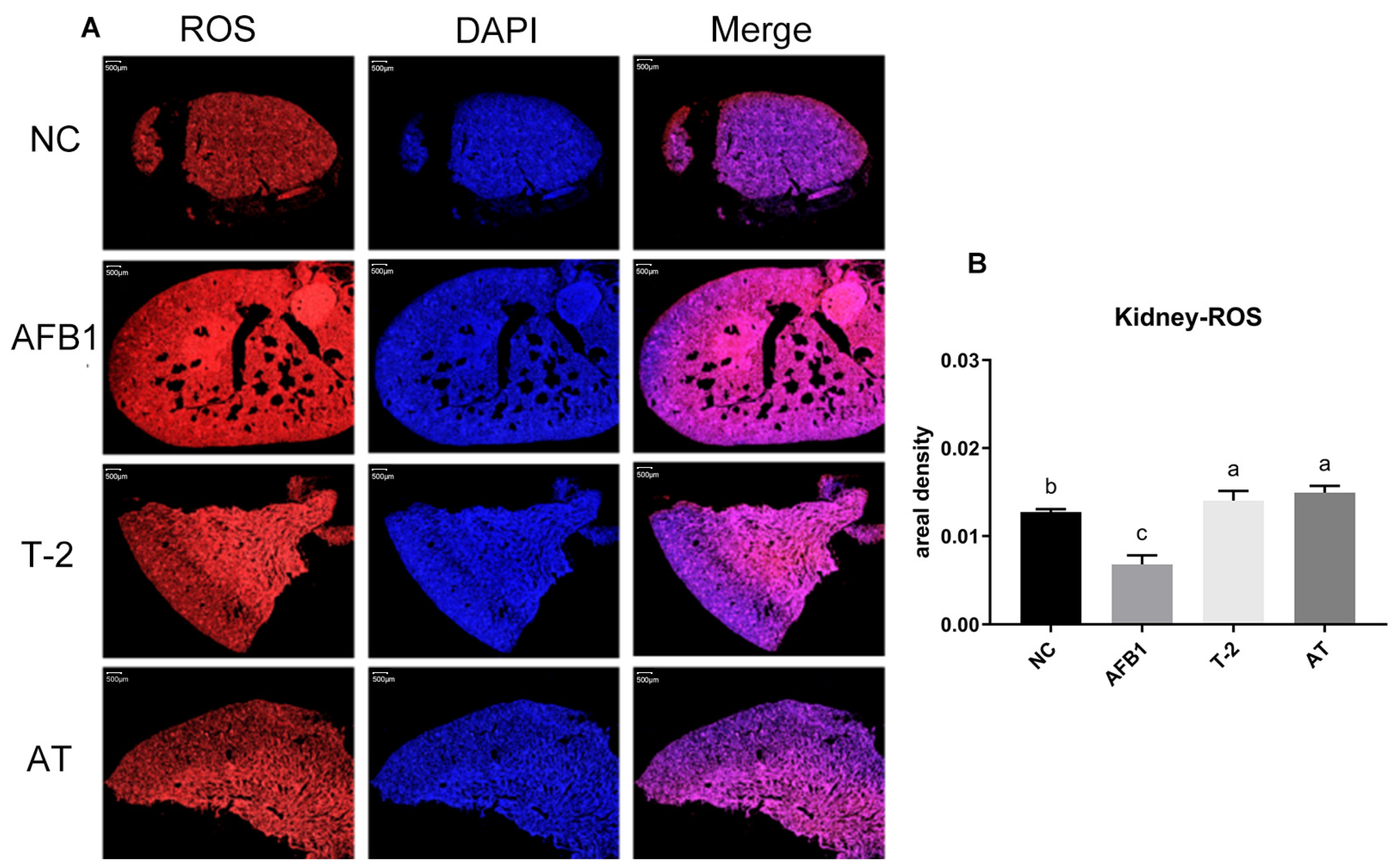
2.10. Quantification of ROS in Kidneys
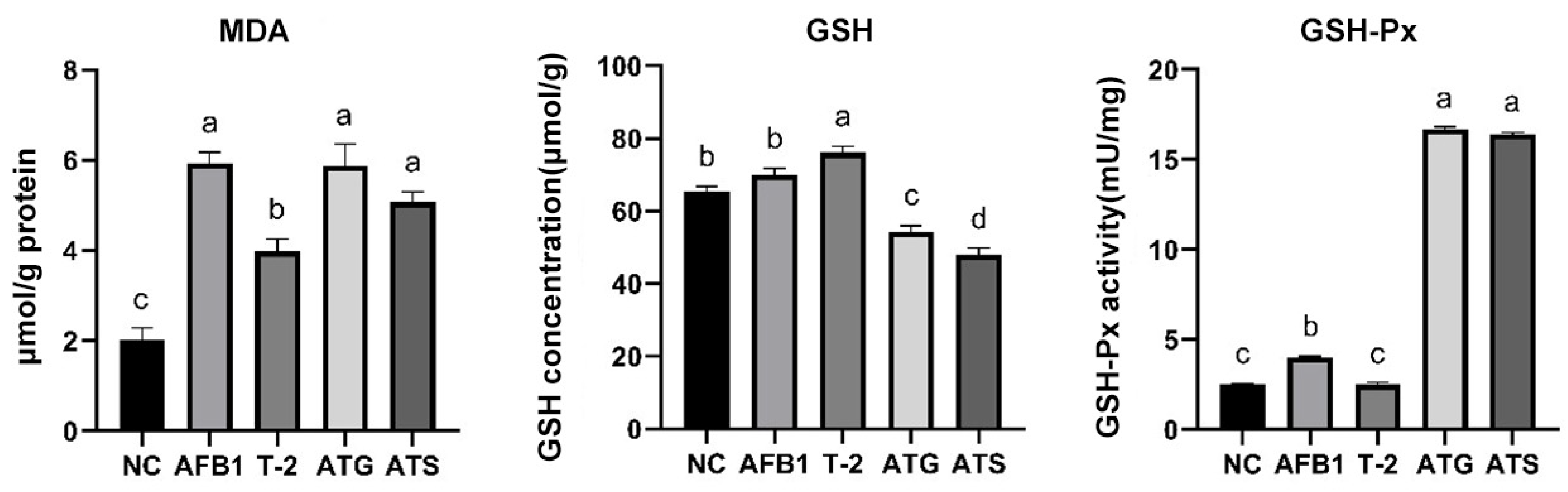
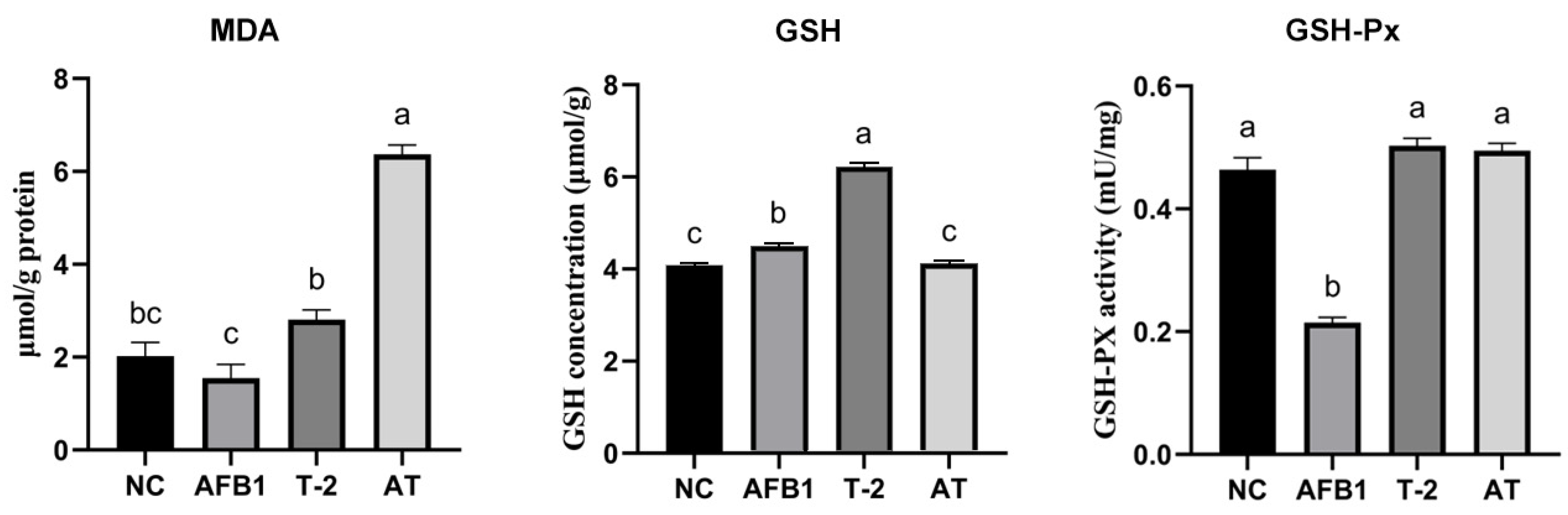
2.11. Assessment of Intracellular MDA, GSH, and GSH-Px Levels in Kidneys
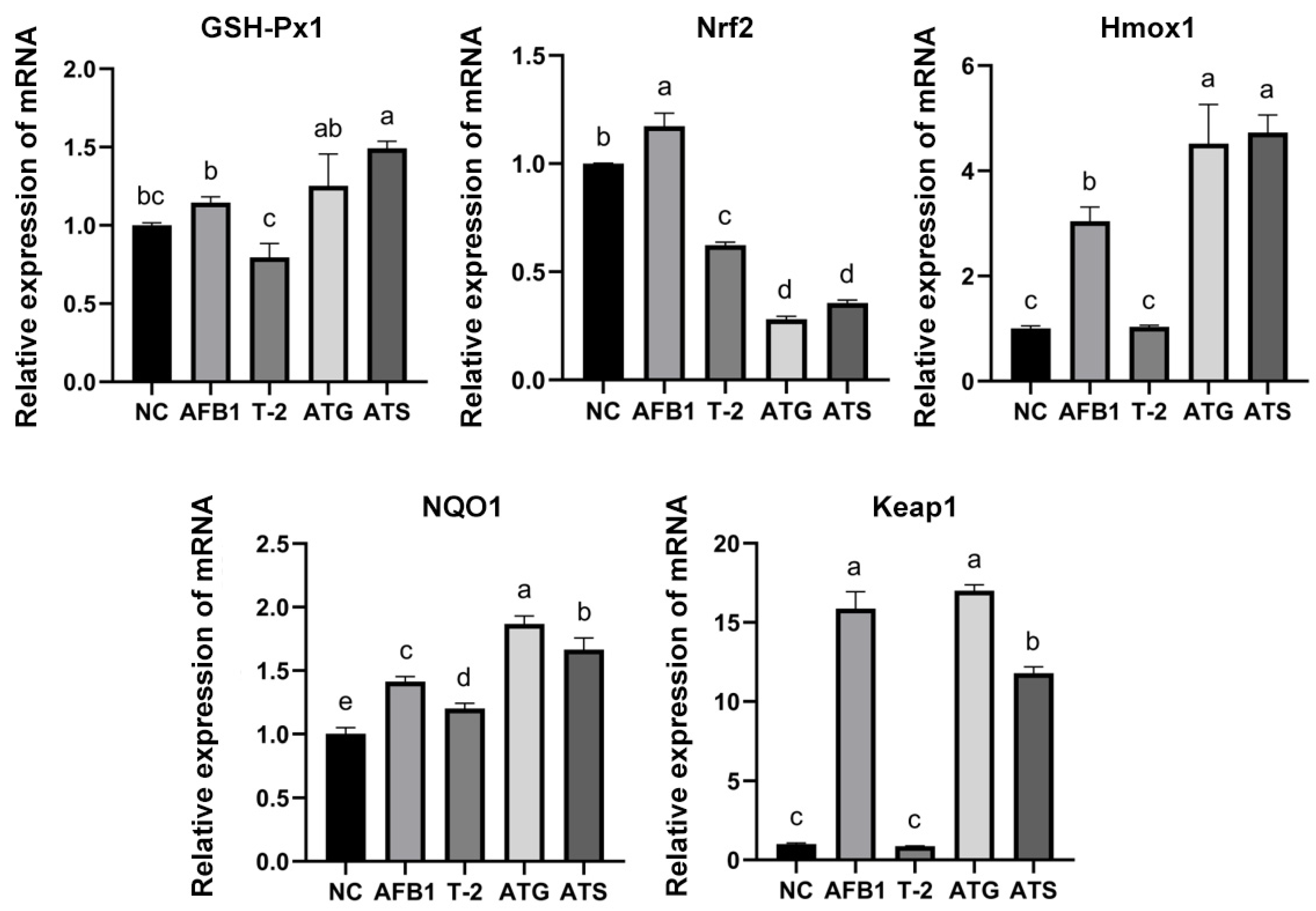
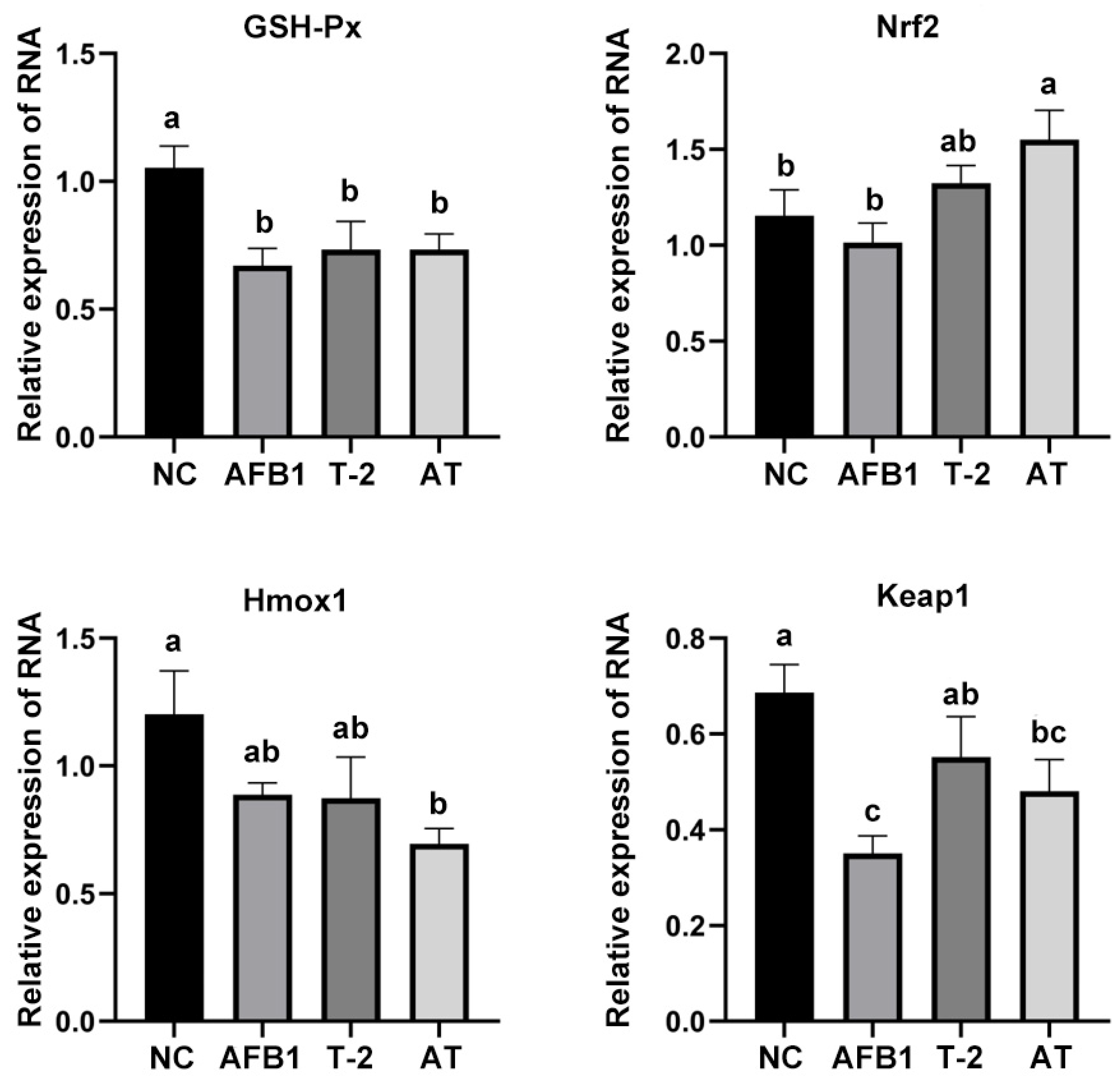
2.12. Evaluation of Oxidative Stress-Related mRNA Expression in Kidneys
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Kits
5.3. Toxicity of T-2/AFB1 In Vitro
5.3.1. Chemical Treatment
5.3.2. Chemical Treatment MTT Assay and Morphologic Evaluation
5.3.3. Measurement of ROS in Cell
5.3.4. Measurement of Intracellular MDA in Cells
5.3.5. Measurement of Intracellular GSH and GSH-Px in Cells
5.3.6. Measurement of Relative Expression Levels of Intracellular Oxidative Stress-Related mRNAs
5.4. Toxicity of T-2/AFB1 In Vivo
5.4.1. Animal Experiments
5.4.2. Serum Biochemistry Analyses
5.4.3. Observations of Kidneys Pathology
5.4.4. Measurement of ROS in Kidneys of Mice
5.4.5. Measurement of MDA, GSH, and GSH-Px in the Kidneys of Mice
5.4.6. Relative Expressions of Oxidative Stress-Related mRNAs in the Kidneys of Mice
5.5. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holmquist, G.U.; Walker, H.W.; Stahr, H.M. Influence of temperature, ph, water activity and antifungal agents on growth of aspergillus flavus and A. parasiticus. J. Food Sci. 1983, 48, 778–782. [Google Scholar] [CrossRef]
- Abbas, H.; Wilkinson, J.; Zablotowicz, R.; Accinelli, C.; Abel, C.; Bruns, H.; Weaver, M.A. Ecology of aspergillus flavus, regulation of aflatoxin production, and management strategies to reduce aflatoxin contamination of corn. Toxin Rev. 2009, 28, 142–153. [Google Scholar] [CrossRef]
- Horn, B.W.; Ramirez-Prado, J.H.; Carbone, I. The sexual state of aspergillus parasiticus. Mycologia 2009, 101, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Wu, W.; Franca, T.C.C.; Jacevic, V.; Wang, X.; Kuca, K. Immune evasion, a potential mechanism of trichothecenes: New insights into negative immune regulations. Int. J. Mol. Sci. 2018, 19, 3307. [Google Scholar] [PubMed] [Green Version]
- Trenholm, H.L.; Thompson, B.K.; Foster, B.C.; Charmley, L.L.; Albassam, M.A. Effects of feeding diets containing fusarium (naturally) contaminated wheat or pure deoxynivalenol (don) in growing pigs. Can. J. Anim. Sci. 1994, 74, 361–369. [Google Scholar]
- Martínez, J.; Hernández-Rodríguez, M.; Méndez-Albores, A.; Téllez-Isaías, G.; Mera Jiménez, E.; Nicolás-Vázquez, M.I.; Miranda Ruvalcaba, R. Computational studies of aflatoxin b(1) (afb(1)): A review. Toxins 2023, 15, 135. [Google Scholar]
- Sabbioni, G. Chemical and physical properties of the major serum albumin adduct of aflatoxin b1 and their implications for the quantification in biological samples. Chem. Biol. Interact. 1990, 75, 1–15. [Google Scholar] [CrossRef]
- Grant, D.; Mendicino, M.; Levy, G.J.S. Xenotransplantation: Just around the corner? Nephrol. Dial. Transplant. 2001, 129, 243–247. [Google Scholar]
- Rawal, S.; Kim, J.E.; Coulombe, R., Jr. Aflatoxin b1 in poultry: Toxicology, metabolism and prevention. Res. Vet. Sci. 2010, 89, 325–331. [Google Scholar]
- Muhammad, I.; Wang, H.; Sun, X.; Wang, X.; Han, M.; Lu, Z.; Cheng, P.; Hussain, M.A.; Zhang, X. Dual role of dietary curcumin through attenuating afb(1)-induced oxidative stress and liver injury via modulating liver phase-i and phase-ii enzymes involved in afb(1) bioactivation and detoxification. Front. Pharmacol. 2018, 9, 554. [Google Scholar]
- Quist, C.F.; Bounous, D.I.; Kilburn, J.V.; Nettles, V.F.; Wyatt, R.D. The effect of dietary aflatoxin on wild turkey poults. J. Wildl. Dis. 2000, 36, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, E.Y.; Rhee, D.K.; Pyo, S. Inhibition of various functions in murine peritoneal macrophages by aflatoxin b1 exposure in vivo. Int. J. Immunopharmacol. 1999, 21, 47–58. [Google Scholar]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [PubMed]
- Li, Y.; Wang, Z.; Beier, R.C.; Shen, J.; De Smet, D.; De Saeger, S.; Zhang, S. T-2 toxin, a trichothecene mycotoxin: Review of toxicity, metabolism, and analytical methods. J. Agric. Food Chem. 2011, 59, 3441–3453. [Google Scholar] [PubMed]
- Schuhmacher-Wolz, U.; Heine, K.; Schneider, K. Report on toxicity data on trichothecene mycotoxins ht-2 and t-2 toxins. Eur. Food Saf. Auth. 2010, 7, 65E. [Google Scholar]
- Chaudhari, M.; Jayaraj, R.; Santhosh, S.R.; Rao, P.V.L. Oxidative damage and gene expression profile of antioxidant enzymes after t-2 toxin exposure in mice. J. Biochem. Mol. Toxicol. 2009, 23, 212–221. [Google Scholar] [CrossRef]
- Wan, Q.; Wu, G.; He, Q.; Tang, H.; Wang, Y. The toxicity of acute exposure to t-2 toxin evaluated by the metabonomics technique. Mol. Biosyst. 2015, 11, 882–891. [Google Scholar] [CrossRef]
- Wu, Q.; Huang, L.; Liu, Z.; Yao, M.; Wang, Y.; Dai, M.; Yuan, Z. A comparison of hepatic in vitro metabolism of t-2 toxin in rats, pigs, chickens, and carp. Xenobiotica 2011, 41, 863–873. [Google Scholar] [CrossRef]
- Liu, X.; Guo, P.; Liu, A.; Wu, Q.; Xue, X.; Dai, M.; Hao, H.; Qu, W.; Xie, S.; Wang, X.; et al. Nitric oxide (no)-mediated mitochondrial damage plays a critical role in t-2 toxin-induced apoptosis and growth hormone deficiency in rat anterior pituitary gh3 cells. Food Chem. Toxicol. 2017, 102, 11–23. [Google Scholar] [CrossRef]
- Liu, X.; Huang, D.; Guo, P.; Wu, Q.; Dai, M.; Cheng, G.; Hao, H.; Xie, S.; Yuan, Z.; Wang, X. Pka/creb and nf-κb pathway regulates akna transcription: A novel insight into t-2 toxin-induced inflammation and gh deficiency in gh3 cells. Toxicology 2017, 392, 81–95. [Google Scholar]
- Manafi, M.; Umakantha, B.; Mohan, K.; Swamy, H.J. Synergistic effects of two commonly contaminating mycotoxins (aflatoxin and t-2 toxin) on biochemical parameters and immune status of broiler chickens. World Appl. Sci. J. 2012, 17, 364–367. [Google Scholar]
- Calvert, T.W.; Aidoo, K.E.; Candlish, A.G.G.; Fuat, A.R.M. Comparison of in vitro cytotoxicity of fusarium mycotoxins, deoxynivalenol, t-2 toxin and zearalenone on selected human epithelial cell lines. Mycopathologia 2005, 159, 413–419. [Google Scholar] [CrossRef]
- Theumer, M.G.; López, A.G.; Masih, D.T.; Chulze, S.N.; Rubinstein, H.R.J.C.; Immunology, D.L. Immunobiological effects of fumonisin b1 in experimental subchronic mycotoxicoses in rats. Clin. Diagn. Lab. Immunol. 2002, 9, 149–155. [Google Scholar] [PubMed] [Green Version]
- Reddy, K.; Salleh, B.; Saad, B.; Abbas, H.; Abel, C.; Shier, W. An overview of mycotoxin contamination in foods and its implications for human health. Toxin Rev. 2010, 29, 3–26. [Google Scholar] [CrossRef]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed—Focus on europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelaziem, S.H.; Hassan, A.M.; Abdelwahhab, M.A. Dietary supplementation with whey protein and ginseng extract counteracts oxidative stress and DNA damage in rats fed an aflatoxin-contaminated diet. Mutat. Res. 2011, 723, 65–71. [Google Scholar] [CrossRef] [PubMed]
- McKean, C.; Tang, L.; Billam, M.; Tang, M.; Theodorakis, C.W.; Kendall, R.J.; Wang, J.S. Comparative acute and combinative toxicity of aflatoxin b1 and t-2 toxin in animals and immortalized human cell lines. J. Appl. Toxicol. 2006, 26, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhao, Z.; Guo, W.; Ling, A.; Wang, J.; Wang, X.; Yang, J. Potential role of individual and combined effects of t-2 toxin, ht-2 toxin and neosolaniol on the apoptosis of porcine leydig cells. Toxins 2022, 14, 145. [Google Scholar] [CrossRef]
- Hou, L.; Gan, F.; Zhou, X.; Zhou, Y.; Qian, G.; Liu, Z.; Huang, K. Immunotoxicity of ochratoxin a and aflatoxin b1 in combination is associated with the nuclear factor kappa b signaling pathway in 3d4/21 cells. Chemosphere 2018, 199, 718–727. [Google Scholar] [CrossRef]
- Wu, C.; Gao, Y.; Li, S.; Huang, X.; Bao, X.; Wang, J.; Zheng, N. Modulation of intestinal epithelial permeability and mucin mrna (muc2, muc5ac, and muc5b) expression and protein secretion in caco-2/ht29-mtx co-cultures exposed to aflatoxin m1, ochratoxin a, and zearalenone individually or collectively. Toxicol. Lett. 2019, 309, 1–9. [Google Scholar] [CrossRef]
- Golli-Bennour, E.E.; Kouidhi, B.; Bouslimi, A.; Abid-Essefi, S.; Hassen, W.; Bacha, H. Cytotoxicity and genotoxicity induced by aflatoxin b1, ochratoxin a, and their combination in cultured vero cells. J. Biochem. Mol. Toxicol. 2010, 24, 42–50. [Google Scholar] [CrossRef]
- Klarić, M.S.; Pepeljnjak, S.; Domijan, A.M.; Petrik, J. Lipid peroxidation and glutathione levels in porcine kidney pk15 cells after individual and combined treatment with fumonisin b(1), beauvericin and ochratoxin a. Basic Clin. Pharmacol. Toxicol. 2007, 100, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Klarić, M.S.; Rumora, L.; Ljubanović, D.; Pepeljnjak, S. Cytotoxicity and apoptosis induced by fumonisin b(1), beauvericin and ochratoxin a in porcine kidney pk15 cells: Effects of individual and combined treatment. Arch. Toxicol. 2008, 82, 247–255. [Google Scholar] [PubMed]
- Babich, H.; Borenfreund, E. Cytotoxicity of t-2 toxin and its metabolites determined with the neutral red cell viability assay. Appl. Environ. Microbiol. 1991, 57, 2101–2103. [Google Scholar] [CrossRef] [PubMed]
- Cid-Castro, C.; Hernández-Espinosa, D.R.; Morán, J. Ros as regulators of mitochondrial dynamics in neurons. Cell. Mol. Neurobiol. 2018, 38, 995–1007. [Google Scholar] [CrossRef]
- Dai, C.; Ciccotosto, G.D.; Cappai, R.; Tang, S.; Li, D.; Xie, S.; Xiao, X.; Velkov, T. Curcumin attenuates colistin-induced neurotoxicity in n2a cells via anti-inflammatory activity, suppression of oxidative stress, and apoptosis. Mol. Neurobiol. 2018, 55, 421–434. [Google Scholar] [CrossRef]
- Nakajima, K.; Masubuchi, Y.; Ito, Y.; Inohana, M.; Takino, M.; Saegusa, Y.; Yoshida, T.; Sugita-Konishi, Y.; Shibutani, M. Developmental exposure of citreoviridin transiently affects hippocampal neurogenesis targeting multiple regulatory functions in mice. Food Chem. Toxicol. 2018, 120, 590–602. [Google Scholar] [CrossRef]
- Pearson, J.N.; Patel, M. The role of oxidative stress in organophosphate and nerve agent toxicity. Ann. N. Y. Acad. Sci. 2016, 1378, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poersch, A.B.; Trombetta, F.; Souto, N.S.; de Oliveira Lima, C.; Braga, A.C.; Dobrachinski, F.; Ribeiro, L.R.; Soares, F.A.; Fighera, M.R.; Royes, L.F.; et al. Fumonisin b1 facilitates seizures induced by pentylenetetrazol in mice. Neurotoxicol. Teratol. 2015, 51, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, W.; Fan, M.; Meng, T.; Chen, X.; Jiang, Y.; Zhu, D.; Hu, W.; Gong, J.; Feng, S.; et al. Deoxynivalenol induces apoptosis in pc12 cells via the mitochondrial pathway. Environ. Toxicol. Pharmacol. 2016, 43, 193–202. [Google Scholar] [CrossRef]
- Wang, X.; Fan, M.; Chu, X.; Zhang, Y.; Rahman, S.U.; Jiang, Y.; Chen, X.; Zhu, D.; Feng, S.; Li, Y.; et al. Deoxynivalenol induces toxicity and apoptosis in piglet hippocampal nerve cells via the mapk signaling pathway. Toxicon 2018, 155, 1–8. [Google Scholar] [PubMed]
- Wang, W.J.; Xu, Z.L.; Yu, C.; Xu, X.H. Effects of aflatoxin b1 on mitochondrial respiration, ros generation and apoptosis in broiler cardiomyocytes. Anim. Sci. J. 2017, 88, 1561–1568. [Google Scholar] [PubMed]
- Zhang, J.; Zheng, N.; Liu, J.; Li, F.D.; Li, S.L.; Wang, J.Q. Aflatoxin b1 and aflatoxin m1 induced cytotoxicity and DNA damage in differentiated and undifferentiated caco-2 cells. Food Chem. Toxicol. 2015, 83, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Sosa, V.; Moliné, T.; Somoza, R.; Paciucci, R.; Kondoh, H.; Leonart, M.E. Oxidative stress and cancer: An overview. Ageing Res. Rev. 2013, 12, 376–390. [Google Scholar]
- Dkhil, M.A.; Al-Khalifa, M.S.; Al-Quraishy, S.; Zrieq, R.; Abdel Moneim, A.E. Indigofera oblongifolia mitigates lead-acetate-induced kidney damage and apoptosis in a rat model. Drug Des. Dev. Ther. 2016, 10, 1847–1856. [Google Scholar]
- Gugliandolo, E.; Peritore, A.F.; D’Amico, R.; Licata, P.; Crupi, R. Evaluation of neuroprotective effects of quercetin against aflatoxin b1-intoxicated mice. Animals 2020, 10, 898. [Google Scholar] [CrossRef]
- Kalantari, H.; Moosavi, M. Review on t-2 toxin. Jundishapur J. Nat. Pharm. Prod. 2010, 5, 26–38. [Google Scholar]
- He, C.H.; Fan, Y.H.; Wang, Y.; Huang, C.Y.; Wang, X.C.; Zhang, H.B. The individual and combined effects of deoxynivalenol and aflatoxin b1 on primary hepatocytes of cyprinus carpio. Int. J. Mol. Sci. 2010, 11, 3760–3768. [Google Scholar] [CrossRef] [Green Version]
- Swamy, H.V.; Smith, T.K.; MacDonald, E.J. Effects of feeding blends of grains naturally contaminated with fusarium mycotoxins on brain regional neurochemistry of starter pigs and broiler chickens. J. Anim. Sci. 2004, 82, 2131–2139. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xing, L.; Zhang, M.; Wang, J.; Zheng, N. The toxic effects of aflatoxin b1 and aflatoxin m1 on kidney through regulating l-proline and downstream apoptosis. BioMed Res. Int. 2018, 2018, 9074861. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, S.; Yang, H.; Wang, Y.; Wang, J.; Zheng, N. L-proline alleviates kidney injury caused by afb1 and afm1 through regulating excessive apoptosis of kidney cells. Toxins 2019, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Dong, R.; Yang, Y.; Xie, H.; Huang, Y.; Chen, X.; Wang, D.; Zhang, Z. Protective effect of organic selenium on oxidative damage and inflammatory reaction of rabbit kidney induced by t-2 toxin. Biol. Trace Elem. Res. 2021, 199, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, D.; Priya, P.; Chattopadhyay, A. Sodium fluoride affects zebrafish behaviour and alters mrna expressions of biomarker genes in the brain: Role of nrf2/keap1. Environ. Toxicol. Pharmacol. 2015, 40, 352–359. [Google Scholar]
- Mackei, M.; Orbán, K.; Molnár, A.; Pál, L.; Dublecz, K.; Husvéth, F.; Neogrády, Z.; Mátis, G. Cellular effects of t-2 toxin on primary hepatic cell culture models of chickens. Toxins 2020, 12, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domijan, A.M.; Marjanović Čermak, A.M.; Vulić, A.; Tartaro Bujak, I.; Pavičić, I.; Pleadin, J.; Markov, K.; Mihaljević, B. Cytotoxicity of gamma irradiated aflatoxin b(1) and ochratoxin a. J. Environ. Sci. Health B 2019, 54, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agyin, J.K.; Santhamma, B.; Nair, H.B.; Roy, S.S.; Tekmal, R.R. Bu-32: A novel proteasome inhibitor for breast cancer. Breast Cancer Res. 2009, 11, R74. [Google Scholar]
- Sun, C.; Wang, L.; Xianyu, B.; Li, T.; Gao, S.; Xu, H. Selenoxide elimination manipulate the oxidative stress to improve the antitumor efficacy. Biomaterials 2019, 225, 119514. [Google Scholar] [CrossRef]
- Zhong, J.; Xie, J.; Xiao, J.; Li, D.; Xu, B.; Wang, X.; Wen, H.; Zhou, Z.; Cheng, Y.; Xu, J.; et al. Inhibition of pde4 by fcpr16 induces ampk-dependent autophagy and confers neuroprotection in sh-sy5y cells and neurons exposed to mpp(+)-induced oxidative insult. Free Radic. Biol. Med. 2019, 135, 87–101. [Google Scholar] [CrossRef]
- Li, D.Y.; Xue, M.Y.; Geng, Z.R.; Chen, P.Y. The suppressive effects of bursopentine (bp5) on oxidative stress and nf-ĸb activation in lipopolysaccharide-activated murine peritoneal macrophages. Cell. Physiol. Biochem. 2012, 29, 9–20. [Google Scholar] [CrossRef]
- Chiu, K.Y.; Loke, S.L.; Ho, F.C. Immunohistochemical demonstration of c-erbb-2 oncoprotein in gastric adenocarcinoma: Comparison of cryostat and paraffin wax sections and effect of fixation. J. Clin. Pathol. 1994, 47, 117–121. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell | AFB1 IC50 (nM) | AFB1 95% CL (nM) | T-2 IC50 (nM) | T-2 95% CL (nM) |
|---|---|---|---|---|
| PK15 | 30,060 | 22,480–43,740 | 1.179 | 0.5892–2.369 |
| Concentration (IC50) | AFB1 (nM) | T-2 (nM) | Mix (nM) | Cell Viability (%) Mean ± SD |
|---|---|---|---|---|
| 0 (NC) | 0 | 0 | 0 | 100 ± 6 |
| 0.125 | 3750 | 0.15 | 3750.15 | 83 ± 3 |
| 0.25 | 7500 | 0.30 | 7500.30 | 83 ± 3 |
| 0.375 | 11,250 | 0.45 | 11,250.45 | 70 ± 1 |
| 0.50 | 15,000 | 0.60 | 15,000.60 | 65 ± 4 |
| 0.625 | 18,750 | 0.75 | 18,750.75 | 59 ± 2 |
| 0.75 | 22,500 | 0.90 | 22,500.90 | 53 ± 2 |
| 0.875 | 26,250 | 1.05 | 26,251.05 | 40 ± 1 |
| 1 | 30,000 | 1.20 | 30,001.20 | 42 ± 2 |
| Treatment | Initial Weight (g) Mean ± SD | Final Weight (g) Mean ± SD | ADFI (g) Mean ± SD |
|---|---|---|---|
| NC | 25.51 ± 0.46 | 37.53 ± 0.89 a | 5.77 ± 0.35 a |
| AFB1 | 26.06 ± 0.37 | 36.26 ± 1.20 a | 5.64 ± 0.55 a |
| T-2 | 25.77 ± 0.35 | 20.50 ± 1.58 c | 4.81 ± 0.81 a |
| AT | 25.82 ± 0.32 | 26.47 ± 2.64 b | 3.20 ± 0.20 b |
| Gene | Upstream Primers (5′ → 3′) | Downstream Primers (5′ → 3′) |
|---|---|---|
| GAPDH | AGGGCATCCTGGGCTACACT | TCCACCACCCTGTTGCTGTA |
| GSH-Px1 | CGTGCAACCAGTTTGGACAT | AGCATGAAGTTGGGCTCGAA |
| Nrf2 | CACTAAACCCAATCCAACCCC | TTGTGAGATGAGCCTCCAAGC |
| Hmox1 | CGCTCCCGAATGAACACTCT | GCGAGGGTCTCTGGTCCTTA |
| NQO1 | GATCATACTGGCCCACTCCG | GAGCAGTCTCGGCAGGATAC |
| Keap1 | CGGAGGCTATGATGGTCACA | ATTCCATCCCTAGCGTGCAG |
| Reagent | Dosage |
|---|---|
| cDNA | 1 μL |
| SYBR Green Mix | 5 μL |
| Upstream primers | 0.2 μL |
| Downstream primers | 0.2 μL |
| RNase free H2O | 3.6 μL |
| Gene | Upstream Primers (5′ → 3′) | Downstream Primers (5′ → 3′) |
|---|---|---|
| GAPDH | ATGACTCCACTCACGGCAAA | CGGCCTCACCCCATTTGATG |
| GSH-Px1 | TGCAATCAGTTCGGACACCA | AAGGTAAAGAGCGGGTGAGC |
| Nrf2 | ATCAGGCCCAGTCCCTCAAT | CAGCCAGCTGCTTGTTTTCG |
| Hmox1 | CAGAAGAGGCTAAGACCGCC | CTCTGACGAAGTGACGCCAT |
| NQO1 | GGTAGCGGCTCCATGTACTC | CCAGACGGTTTCCAGACGTT |
| Keap1 | GATGGGCAGGACCAGTTGAA | CCGGGTCATAGCATTCCACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, S.; Wu, Y.; Gao, S.; Zhou, M.; Liu, Z.; Xiong, Q.; Jiang, L.; Yuan, G.; Li, L.; Yang, L. Deciphering the Hazardous Effects of AFB1 and T-2 Toxins: Unveiling Toxicity and Oxidative Stress Mechanisms in PK15 Cells and Mouse Kidneys. Toxins 2023, 15, 503. https://doi.org/10.3390/toxins15080503
Xiao S, Wu Y, Gao S, Zhou M, Liu Z, Xiong Q, Jiang L, Yuan G, Li L, Yang L. Deciphering the Hazardous Effects of AFB1 and T-2 Toxins: Unveiling Toxicity and Oxidative Stress Mechanisms in PK15 Cells and Mouse Kidneys. Toxins. 2023; 15(8):503. https://doi.org/10.3390/toxins15080503
Chicago/Turabian StyleXiao, Shuai, Yingxin Wu, Suisui Gao, Mingxia Zhou, Zhiwei Liu, Qianbo Xiong, Lihuang Jiang, Guoxiang Yuan, Linfeng Li, and Lingchen Yang. 2023. "Deciphering the Hazardous Effects of AFB1 and T-2 Toxins: Unveiling Toxicity and Oxidative Stress Mechanisms in PK15 Cells and Mouse Kidneys" Toxins 15, no. 8: 503. https://doi.org/10.3390/toxins15080503