The Effects of the Harmful Algal Bloom Species Karenia brevis on Survival of Red Porgy (Pagrus pagrus) Larvae

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
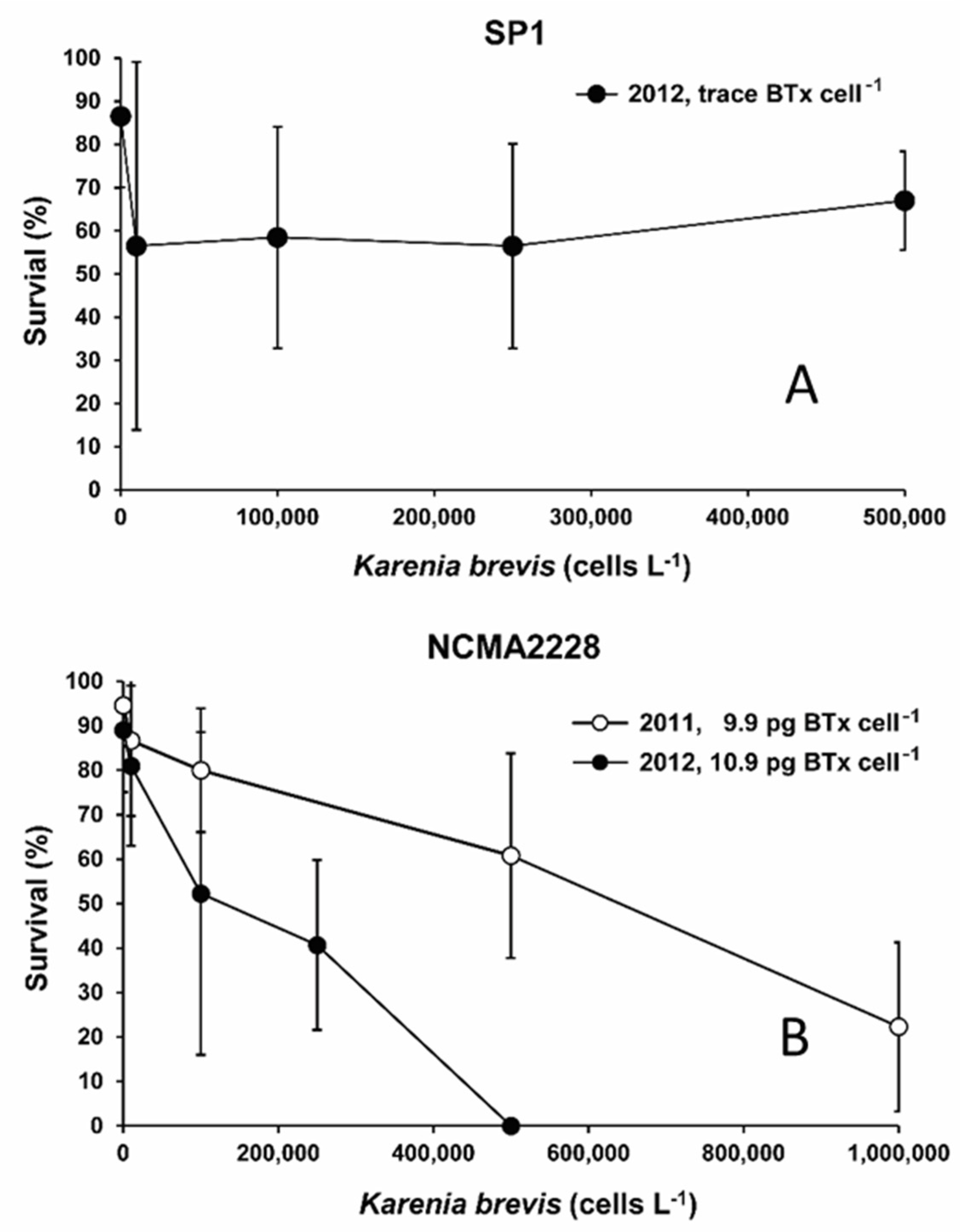
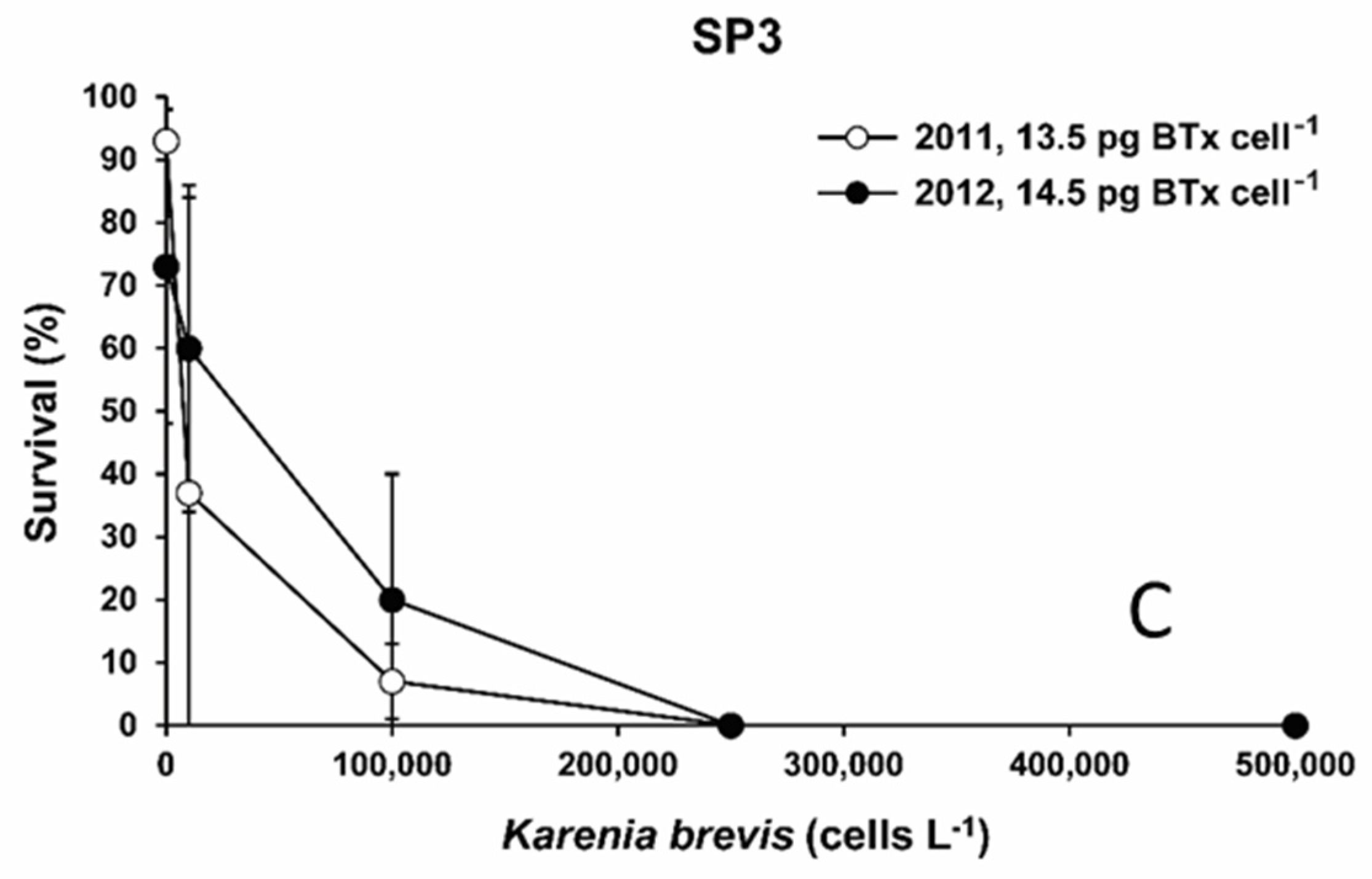
2.1. Effects of K. brevis on Larval Fish Survival
2.2. EC50 of Karenia brevis Versus Survivorship
2.3. Effect of Increased Exposure Time on Survivorship
3. Discussion
4. Materials and Methods
4.1. Larval Fish Rearing
4.2. Karenia brevis Cultures
4.3. Brevetoxin Extractions and Quantification
4.4. Exposure Experiments
4.5. Estimating the EC50 Value
4.6. Examination of How Prolonged Exposure to K. brevis Impacts Survivorship of Red Porgy Larvae
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stumpf, R.P.; Li, Y.; Kirkpatrick, B.; Litaker, R.W.; Hubbard, K.A.; Currier, R.D.; Harrison, K.K.; Tomlinson, M.C. Quantifying Karenia brevis Bloom Severity and Respiratory Irritation Impact along the Shoreline of Southwest Florida. PLoS ONE 2022, 17, e0260755. [Google Scholar] [CrossRef] [PubMed]
- Landsberg, J.H. The Effects of Harmful Algal Blooms on Aquatic Organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Poli, M.A.; Mende, T.J.; Baden, D.G. Brevetoxins, Unique Activators of Voltage-Sensitive Sodium Channels, Bind to Specific Sites in Rat Brain Synaptosomes. Mol. Pharmacol. 1986, 30, 129–135. [Google Scholar] [PubMed]
- Tester, P.A.; Shea, D.; Kibler, S.R.; Varnam, S.M.; Black, M.D.; Litaker, R.W. Relationships among Water Column Toxins, Cell Abundance and Chlorophyll Concentrations during Karenia brevis Blooms. Cont. Shelf Res. 2008, 28, 59–72. [Google Scholar] [CrossRef]
- Tester, P.A.; Turner, J.T.; Shea, D. Vectorial Transport of Toxins from the Dinoflagellate Gymnodinium breve through Copepods to Fish. J. Plankton Res. 2000, 22, 47–62. [Google Scholar] [CrossRef]
- Sotka, E.E.; McCarty, A.; Monroe, E.A.; Oakman, N.; Van Dolah, F.M. Benthic Herbivores Are Not Deterred by Brevetoxins Produced by the Red Tide Dinoflagellate Karenia brevis. J. Chem. Ecol. 2009, 35, 851–859. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Haubois, A.-G.; Sengco, M.R.; Pierce, R.H.; Culter, J.K.; Anderson, D.M. Trophic Transfer of Brevetoxins to the Benthic Macrofaunal Community during a Bloom of the Harmful Dinoflagellate Karenia brevis in Sarasota Bay, Florida. Harmful Algae 2012, 16, 27–34. [Google Scholar] [CrossRef]
- Echevarria, M.; Naar, J.P.; Tomas, C.; Pawlik, J.R. Effects of Karenia brevis on Clearance Rates and Bioaccumulation of Brevetoxins in Benthic Suspension Feeding Invertebrates. Aquat. Toxicol. 2012, 106–107, 85–94. [Google Scholar] [CrossRef]
- Pierce, R.H.; Henry, M.S.; Blum, P.C.; Hamel, S.L.; Kirkpatrick, B.; Cheng, Y.S.; Zhou, Y.; Irvin, C.M.; Naar, J.; Weidner, A.; et al. Brevetoxin Composition in Water and Marine Aerosol along a Florida Beach: Assessing Potential Human Exposure to Marine Biotoxins. Harmful Algae 2005, 4, 965–972. [Google Scholar] [CrossRef]
- Javaruski, J.; Adhikari, P.L.; Muller, J.; Parsons, M.L. Preservation of Brevetoxins in Southwest Florida Coastal Sediments. Harmful Algae 2022, 114, 102222. [Google Scholar] [CrossRef]
- Cohen, J.H.; Tester, P.A.; Forward, R.B. Sublethal Effects of the Toxic Dinoflagellate Karenia brevis on Marine Copepod Behavior. J. Plankton Res. 2007, 29, 301–315. [Google Scholar] [CrossRef]
- Lester, K.M.; Heil, C.A.; Neely, M.B.; Spence, D.N.; Murasko, S.; Hopkins, T.L.; Sutton, T.T.; Burghart, S.E.; Bohrer, R.N.; Remsen, A.W.; et al. Zooplankton and Karenia brevis in the Gulf of Mexico. Cont. Shelf Res. 2008, 28, 99–111. [Google Scholar] [CrossRef]
- Gunter, G.; Williams, R.H.; Davis, C.C.; Smith, F.G.W. Catastrophic Mass Mortality of Marine Animals and Coincident Phytoplankton Bloom on the West Coast of Florida, November 1946 to August 1947. Ecol. Monogr. 1948, 18, 309–324. [Google Scholar] [CrossRef]
- van Deventer, M.; Atwood, K.; Vargo, G.A.; Flewelling, L.J.; Landsberg, J.H.; Naar, J.P.; Stanek, D. Karenia brevis Red Tides and Brevetoxin-Contaminated Fish: A High Risk Factor for Florida’s Scavenging Shorebirds? Bot. Mar. 2012, 55, 31–37. [Google Scholar] [CrossRef]
- Fire, S.E.; Flewelling, L.J.; Wang, Z.; Naar, J.; Henry, M.S.; Pierce, R.H.; Wells, R.S. Florida Red Tide and Brevetoxins: Association and Exposure in Live Resident Bottlenose Dolphins (Tursiops truncatus) in the Eastern Gulf of Mexico, USA. Mar. Mammal Sci. 2008, 24, 831–844. [Google Scholar] [CrossRef]
- Watkins, S.M.; Reich, A.; Fleming, L.E.; Hammond, R. Neurotoxic Shellfish Poisoning. Mar. Drugs 2008, 6, 431–455. [Google Scholar] [CrossRef] [Green Version]
- Flewelling, L.J.; Naar, J.P.; Abbott, J.P.; Baden, D.G.; Barros, N.B.; Bossart, G.D.; Bottein, M.-Y.D.; Hammond, D.G.; Haubold, E.M.; Heil, C.A.; et al. Red Tides and Marine Mammal Mortalities. Nature 2005, 435, 755–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazensky, R.; Silva-Sanchez, C.; Kroll, K.J.; Chow, M.; Chen, S.; Tripp, K.; Walsh, M.T.; Denslow, N.D. Investigating an Increase in Florida Manatee Mortalities Using a Proteomic Approach. Sci. Rep. 2021, 11, 4282. [Google Scholar] [CrossRef]
- Feinstein, A.; Ceurvels, A.R.; Hutton, R.F.; Snoek, E. Red Tide Outbreaks off the Florida West Coast; Florida State Board of Conservation 55-15, The Marine Laboratory, Univeristy of Miami: Coral Gables, FL, USA, 1955; p. 39. [Google Scholar]
- Landsberg, J.H.; Flewelling, L.J.; Naar, J. Karenia brevis Red Tides, Brevetoxins in the Food Web, and Impacts on Natural Resources: Decadal Advancements. Harmful Algae 2009, 8, 598–607. [Google Scholar] [CrossRef]
- Davis, C.C. Gymnodinium brevis Sp. Nov., a Cause of Discolored Water and Animal Mortality in the Gulf of Mexico. Bot. Gaz. 1948, 109, 358–360. [Google Scholar] [CrossRef]
- Smith, G.B. The 1971 Red Tide and Its Impact on Certain Reef Communities in the Mid-Eastern Gulf of Mexico. Environ. Lett. 1975, 9, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Baden, D.G.; Bourdelais, A.J.; Jacocks, H.; Michelliza, S.; Naar, J. Natural and Derivative Brevetoxins: Historical Background, Multiplicity, and Effects. Environ. Health Perspect. 2005, 113, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Backer, L.C.; Baden, D.G. Overview of Aerosolized Florida Red Tide Toxins: Exposures and Effects. Environ. Health Perspect. 2005, 113, 618–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, B.; Pierce, R.; Cheng, Y.S.; Henry, M.S.; Blum, P.; Osborn, S.; Nierenberg, K.; Pederson, B.A.; Fleming, L.E.; Reich, A.; et al. Inland Transport of Aerosolized Florida Red Tide Toxins. Harmful Algae 2010, 9, 186–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirkpatrick, B.; Fleming, L.E.; Bean, J.A.; Nierenberg, K.; Backer, L.C.; Cheng, Y.S.; Pierce, R.; Reich, A.; Naar, J.; Wanner, A.; et al. Aerosolized Red Tide Toxins (Brevetoxins) and Asthma: Continued Health Effects after 1 h Beach Exposure. Harmful Algae 2011, 10, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Leverone, J.R. Comparative Effects of the Toxic Dinoflagellate, Karenia brevis, on Bivalve Molluscs from Florida. Ph.D. Thesis, University of South Florida, Tampa, FL, USA, 2007. [Google Scholar]
- Brovedani, V.; Pelin, M.; D’Orlando, E.; Poli, M. Brevetoxins: Toxicological Profile. In Marine and Freshwater Toxins; Gopalakrishnakone, P., Haddad, V., Tubaro, A., Kim, E., Kem, W.R., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 113–127. ISBN 978-94-007-6418-7. [Google Scholar]
- Warlen, S.M.; Tester, P.A.; Colby, D.R. Recruitment of Larval Fishes into a North Carolina Estuary during a Bloom of the Red Tide Dinoflagellate, Gymnodinium breve. Bull. Mar. Sci. 1998, 63, 83–95. [Google Scholar]
- Flaherty, K.E.; Landsberg, J.H. Effects of a Persistent Red Tide (Karenia brevis) Bloom on Community Structure and Species-Specific Relative Abundance of Nekton in a Gulf of Mexico Estuary. Estuaries Coasts 2011, 34, 417–439. [Google Scholar] [CrossRef]
- Riley, C.M.; Holt, S.A.; Holt, G.J.; Buskey, E.J.; Arnold, C.R. Mortality of Larval Red Drum (Sciaenops ocellatus) Associated with a Ptychodiscus brevis Red Tide. Contributions in Marine Science. Contrib. Mar. Sci. 1989, 31, 137–146. [Google Scholar]
- Kimm-Brinson, K.L.; Ramsdell, J.S. The Red Tide Toxin, Brevetoxin, Induces Embryo Toxicity and Developmental Abnormalities. Environ. Health Perspect. 2001, 109, 377–381. [Google Scholar] [CrossRef]
- DeVries, A.D. The Life History, Reproductive Ecology and Demography of the Red Porgy, Pagrus pagrus, in the Northestem Gulf of Mexico. Ph.D. Dissertation, Florida State University, Tallahassee, FL, USA, 2006. [Google Scholar]
- Red Porgy. Available online: https://en.wikipedia.org/wiki/Red_porgy (accessed on 11 April 2022).
- Hood, P.B.; Johnson, A.K. Age, Growth, Mortality, and Reproduction of Red Porgy, Pagrus pagrus, from the Eastern Gulf of Mexico. Fish. Bull. 2000, 98, 723–735. [Google Scholar]
- Tester, P.A.; Steidinger, K.A. Gymnodinium breve Red Tide Blooms: Initiation, Transport, and Consequences of Surface Circulation. Limnol. Oceanogr. 1997, 42, 1039–1051. [Google Scholar] [CrossRef] [Green Version]
- Stumpf, R.P.; Litaker, R.W.; Lanerolle, L.; Tester, P.A. Hydrodynamic Accumulation of Karenia off the West Coast of Florida. Cont. Shelf Res. 2008, 28, 189–213. [Google Scholar] [CrossRef]
- Quick, J.A.; Henderson, G.E. Evidence of Ichthyointoxicative Phenonmenon in Gymnodinium Red Tides. In Proceedings of The First International Conference on Toxic Dinoflagellate Blooms; LoCicero, V.R., Ed.; Massachusetts Science and Technology Foundation: Wakefield, MA, USA, 1977; pp. 413–422. [Google Scholar]
- Quick, J.A.; Henderson, G.E. Effects of Gymnodinium Breve Red Tide on Fishes and Birds: A Preliminary Report on Behavior, Anatomy, Hematology and Histopathology; Florida Department of Natural Resources Marine Research Laboratory: St. Petersburg. FL, USA; pp. 85–113.
- Leverone, J.R.; Blake, N.J.; Pierce, R.H.; Shumway, S.E. Effects of the Dinoflagellate Karenia brevis on Larval Development in Three Species of Bivalve Mollusc from Florida. Toxicon 2006, 48, 75–84. [Google Scholar] [CrossRef]
- Rolton, A.; Vignier, J.; Soudant, P.; Shumway, S.E.; Bricelj, V.M.; Volety, A.K. Effects of the Red Tide Dinoflagellate, Karenia brevis, on Early Development of the Eastern Oyster Crassostrea virginica and Northern Quahog Mercenaria mercenaria. Aquat. Toxicol. 2014, 155, 199–206. [Google Scholar] [CrossRef]
- Shen, H.; Song, X.; Zhang, Y.; Zhang, P.; Li, J.; Song, W.; Yu, Z. Profiling of Brevetoxin Metabolites Produced by Karenia brevis 165 Based on Liquid Chromatography-Mass Spectrometry. Toxins 2021, 13, 354. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Sunda, W.G.; Litaker, R.W.; Shea, D.; Tester, P.A. Nitrogen Limitation Increases Brevetoxins in Karenia brevis (Dinophyceae): Implications for Bloom Toxicity: N-Limitation Increases Brevetoxins. J. Phycol. 2012, 48, 844–858. [Google Scholar] [CrossRef]
- Hardison, D.R.; Sunda, W.G.; Shea, D.; Litaker, R.W. Increased Toxicity of Karenia brevis during Phosphate Limited Growth: Ecological and Evolutionary Implications. PLoS ONE 2013, 8, e58545. [Google Scholar] [CrossRef] [Green Version]
- Dupont, J.; Hallock, P.; Jaap, W. Ecological Impacts of the 2005 Red Tide on Artificial Reef Epibenthic Macroinvertebrate and Fish Communities in the Eastern Gulf of Mexico. Mar. Ecol. Prog. Ser. 2010, 415, 189–200. [Google Scholar] [CrossRef]
- Gannon, D.; Berens McCabe, E.; Camilleri, S.; Gannon, J.; Brueggen, M.; Barleycorn, A.; Palubok, V.; Kirkpatrick, G.; Wells, R. Effects of Karenia brevis Harmful Algal Blooms on Nearshore Fish Communities in Southwest Florida. Mar. Ecol. Prog. Ser. 2009, 378, 171–186. [Google Scholar] [CrossRef]
- Walters, S.; Lowerre-Barbieri, S.; Bickford, J.; Tustison, J.; Landsberg, J. Effects of Karenia brevis Red Tide on the Spatial Distribution of Spawning Aggregations of Sand Seatrout Cynoscion arenarius in Tampa Bay, Florida. Mar. Ecol. Prog. Ser. 2013, 479, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Rolton, A.; Vignier, J.; Volety, A.; Shumway, S.; Bricelj, V.M.; Soudant, P. Impacts of Exposure to the Toxic Dinoflagellate Karenia brevis on Reproduction of the Northern Quahog, Mercenaria mercenaria. Aquat. Toxicol. 2018, 202, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Rolton, A.; Soudant, P.; Vignier, J.; Pierce, R.; Henry, M.; Shumway, S.E.; Bricelj, V.M.; Volety, A.K. Susceptibility of Gametes and Embryos of the Eastern Oyster, Crassostrea virginica, to Karenia brevis and Its Toxins. Toxicon 2015, 99, 6–15. [Google Scholar] [CrossRef]
- McFarland, K.; Jean, F.; Thebault, J.; Volety, A.K. Potential Impacts of Blooms of the Toxic Dinoflagellate Karenia brevis on the Growth, Survival and Juvenile Recruitment of the Non-Native Green Mussel Perna viridis in Southeastern United States. Toxicon 2016, 109, 94–102. [Google Scholar] [CrossRef]
- Simon, J.L.; Dauer, D.M. A Quantitative Evaluation of Red-Tide Induced Mass Mortalities of Benthic Invertebrates in Tampa Bay, Florida. Environ. Lett. 1972, 3, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.B. Relationship of Eastern Gulf of Mexico Reef-Fish Communities to the Species Equilibrium Theory of Insular Biogeography. J. Biogeogr. 1979, 6, 49. [Google Scholar] [CrossRef]
- Lawrence, J.M.; Cobb, J.; Swigart, J.P. Red-Tide–Associated Mass Mortality in 2005 of the Sand Dollars Encope aberrans, Encope michelini, and Mellita tenuis (Echinodermata: Echinoidea) on the Central Florida Gulf Coast Shelf. Gulf Mex. Sci. 2006, 24, 83–87. [Google Scholar] [CrossRef]
- Lerma, L. The Effects of a Red Tide, Karenia brevis Episode on the Benthic Macroinvertebrate Communities of South Padre Island, Texas. Master’s Thesis, The University Texas at Brownsville, Brownsville, TX, USA, 2013. [Google Scholar]
- Brown, A.R.; Lilley, M.; Shutler, J.; Lowe, C.; Artioli, Y.; Torres, R.; Berdalet, E.; Tyler, C.R. Assessing Risks and Mitigating Impacts of Harmful Algal Blooms on Mariculture and Marine Fisheries. Rev. Aquac. 2019, 12, 1663–1688. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.; Ueno, M.; Liang, Y.; Kim, D.; Oda, T. Generation of Reactive Oxygen Species (ROS) by Harmful Algal Bloom (HAB)-Forming Phytoplankton and Their Potential Impact on Surrounding Living Organisms. Antioxidants 2022, 11, 206. [Google Scholar] [CrossRef]
- Gao, Y.; Erdner, D.L. Dynamics of Cell Death across Growth Stages and the Diel Cycle in the Dinoflagellate Karenia brevis. J. Eukaryot. Microbiol. 2022, 69, e12874. [Google Scholar] [CrossRef]
- Diaz, J.M.; Plummer, S.; Tomas, C.; Alves-de-Souza, C. Production of Extracellular Superoxide and Hydrogen Peroxide by Five Marine Species of Harmful Bloom-Forming Algae. J. Plankton Res. 2018, 40, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Rolton, A.; Rhodes, L.; Hutson, K.S.; Biessy, L.; Bui, T.; MacKenzie, L.; Symonds, J.E.; Smith, K.F. Effects of Harmful Algal Blooms on Fish and Shellfish Species: A Case Study of New Zealand in a Changing Environment. Toxins 2022, 14, 341. [Google Scholar] [CrossRef] [PubMed]
- Dorantes-Aranda, J.J.; Seger, A.; Mardones, J.I.; Nichols, P.D.; Hallegraeff, G.M. Progress in Understanding Algal Bloom-Mediated Fish Kills: The Role of Superoxide Radicals, Phycotoxins and Fatty Acids. PLoS ONE 2015, 10, e0133549. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; McNabb, P.; Rhodes, L.; Holland, P.; Webb, S.; Adamson, J.; Immers, A.; Gooneratne, R.; Holland, J. The Toxic Effects of Three Dinoflagellate Species from the Genus Karenia on Invertebrate Larvae and Finfish. N. Z. J. Mar. Freshw. Res. 2012, 46, 149–165. [Google Scholar] [CrossRef]
- Perrault, J.R.; Schmid, J.R.; Walsh, C.J.; Yordy, J.E.; Tucker, A.D. Brevetoxin Exposure, Superoxide Dismutase Activity and Plasma Protein Electrophoretic Profiles in Wild-Caught Kemp’s Ridley Sea Turtles (Lepidochelys kempii) in Southwest Florida. Harmful Algae 2014, 37, 194–202. [Google Scholar] [CrossRef]
- Ross, C.; Ritson-Williams, R.; Pierce, R.; Bullington, J.B.; Henry, M.; Paul, V.J. Effects of the Florida Red Tide Dinoflagellate, Karenia brevis, on Oxidative Stress and Metamorphosis of Larvae of the Coral Porites astreoides. Harmful Algae 2010, 9, 173–179. [Google Scholar] [CrossRef]
- Walsh, C.J.; Butawan, M.; Yordy, J.; Ball, R.; Flewelling, L.; de Wit, M.; Bonde, R.K. Sublethal Red Tide Toxin Exposure in Free-Ranging Manatees (Trichechus manatus) Affects the Immune System through Reduced Lymphocyte Proliferation Responses, Inflammation, and Oxidative Stress. Aquat. Toxicol. 2015, 161, 73–84. [Google Scholar] [CrossRef]
- Murray, T.F. Chapter 3—Neurotoxic: Ciguatoxin and Brevetoxin—From Excitotoxicity to Neurotherapeutics. In Advances in Neurotoxicology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 6, pp. 89–104. ISBN 978-0-12-822014-6. [Google Scholar]
- Hort, V.; Abadie, E.; Arnich, N.; Dechraoui Bottein, M.-Y.; Amzil, Z. Chemodiversity of Brevetoxins and Other Potentially Toxic Metabolites Produced by Karenia Spp. and Their Metabolic Products in Marine Organisms. Mar. Drugs 2021, 19, 656. [Google Scholar] [CrossRef]
- Ostrowski, A.D. Effects of Salinity and Temperature on the Growth, Survival, Whole Body Osmolality, Stress Resistance, and MRNA Expression of Na+/k+—ATPase in Red Porgy Larvae, Pagrus pagrus. Master’s Thesis, University of North Carolina Wilmington, Wilmington, NC, USA, 2009. [Google Scholar]
- Errera, R.M.; Bourdelais, A.; Drennan, M.A.; Dodd, E.B.; Henrichs, D.W.; Campbell, L. Variation in Brevetoxin and Brevenal Content among Clonal Cultures of Karenia brevis May Influence Bloom Toxicity. Toxicon 2010, 55, 195–203. [Google Scholar] [CrossRef]
- Waggett, R.J.; Hardison, D.R.; Tester, P.A. Toxicity and Nutritional Inadequacy of Karenia brevis: Synergistic Mechanisms Disrupt Top-down Grazer Control. Mar. Ecol. Prog. Ser. 2012, 444, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Sunda, W.G.; Hardison, D.R. Ammonium Uptake and Growth Limitation in Marine Phytoplankton. Limnol. Oceanogr. 2007, 52, 2496–2506. [Google Scholar] [CrossRef]
- Cheng, Y.S.; Zhou, Y.; Irvin, C.M.; Pierce, R.H.; Naar, J.; Backer, L.C.; Fleming, L.E.; Kirkpatrick, B.; Baden, D.G. Characterization of Marine Aerosol for Assessment of Human Exposure to Brevetoxins. Environ. Health Perspect. 2005, 113, 638–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, W.G.; Mead, R.N.; Brand, L.E.; Shea, D. Determination of Brevetoxin in Recent Marine Sediments. Chemosphere 2008, 73, 1373–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Litaker, R.W.; Bogdanoff, A.K.; Hardison, D.R.; Holland, W.C.; Ostrowski, A.; Morris, J.A. The Effects of the Harmful Algal Bloom Species Karenia brevis on Survival of Red Porgy (Pagrus pagrus) Larvae. Toxins 2022, 14, 439. https://doi.org/10.3390/toxins14070439
Litaker RW, Bogdanoff AK, Hardison DR, Holland WC, Ostrowski A, Morris JA. The Effects of the Harmful Algal Bloom Species Karenia brevis on Survival of Red Porgy (Pagrus pagrus) Larvae. Toxins. 2022; 14(7):439. https://doi.org/10.3390/toxins14070439
Chicago/Turabian StyleLitaker, Richard Wayne, Alex K. Bogdanoff, Donnie Ransom Hardison, William C. Holland, Andrew Ostrowski, and James A. Morris. 2022. "The Effects of the Harmful Algal Bloom Species Karenia brevis on Survival of Red Porgy (Pagrus pagrus) Larvae" Toxins 14, no. 7: 439. https://doi.org/10.3390/toxins14070439