Bitiscetin-3, a Novel C-Type Lectin-like Protein Cloned from the Venom Gland of the Viper Bitis arietans, Induces Platelet Agglutination and Inhibits Binding of Von Willebrand Factor to Collagen

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Construction of cDNA and a Library of cDNA-Containing Vectors
2.3. Primers
2.4. PCR and Sequencing
2.5. GenBank Accession Numbers of Bitiscetin-3α and -β
2.6. Sequence Comparisons
2.7. Recombinant Expression of cDNA
2.8. Purification of Recombinant Protein
2.9. Platelet Agglutination Assay
2.10. Western Blotting
2.11. Enzyme-Linked Immuno-Sorbent Assay (ELISA) for Analyzing the Effect of rBit-3 on Binding of GPIb to VWF
2.12. ELISA for Analyzing the Effect of rBit-3 on Binding of Bitiscetin-1 to VWF
2.13. ELISA for Evaluating the Effect of rBit-3 on VWF-to-Collagen Binding
2.14. Statistical Analysis
3. Results
3.1. cDNA Cloning of a Novel Bitiscetin
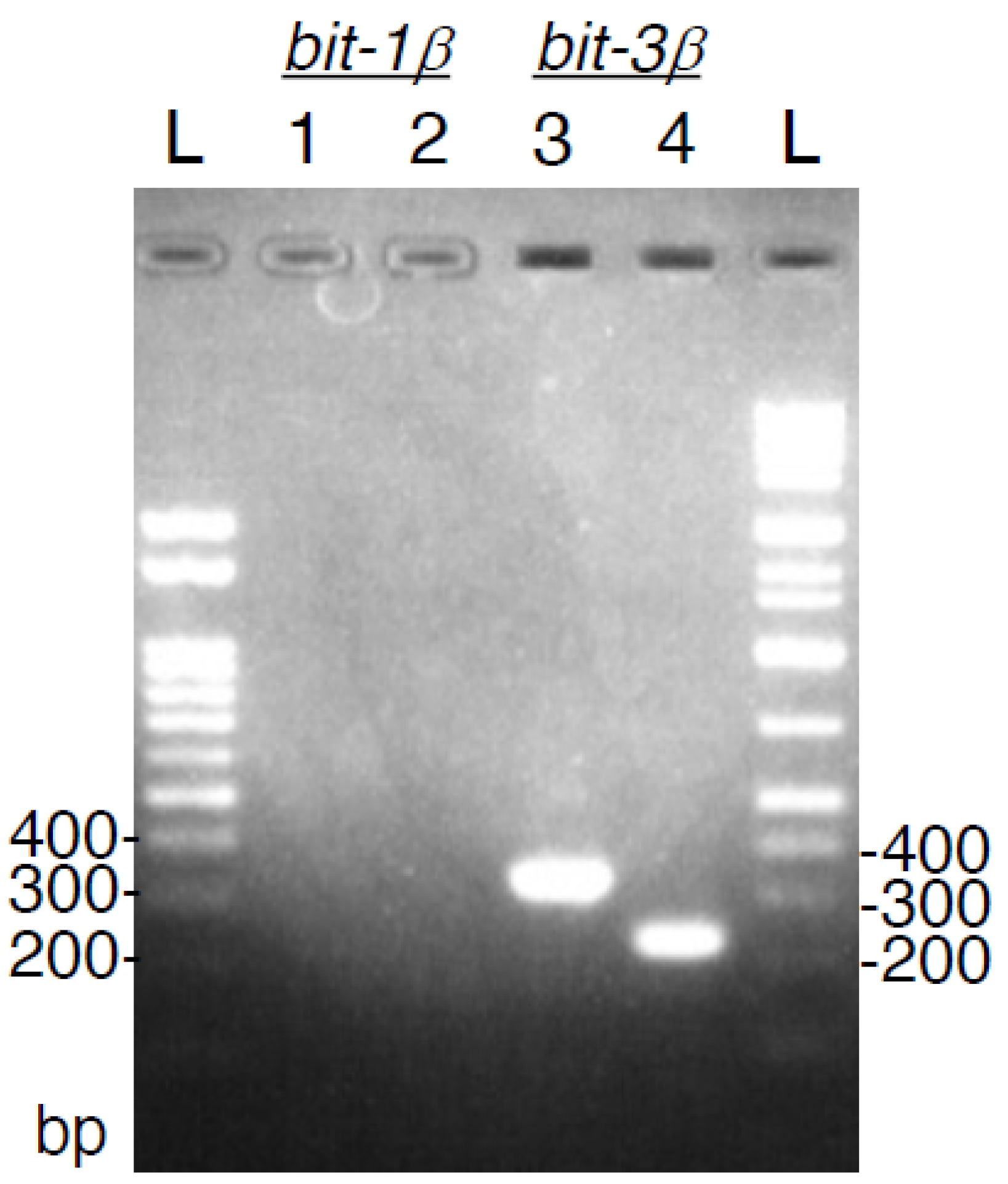
3.2. Bitiscetin-1β cDNA Could Not Be Detected
3.3. The Bitiscetin-3α and -3β Amino Acid Sequences
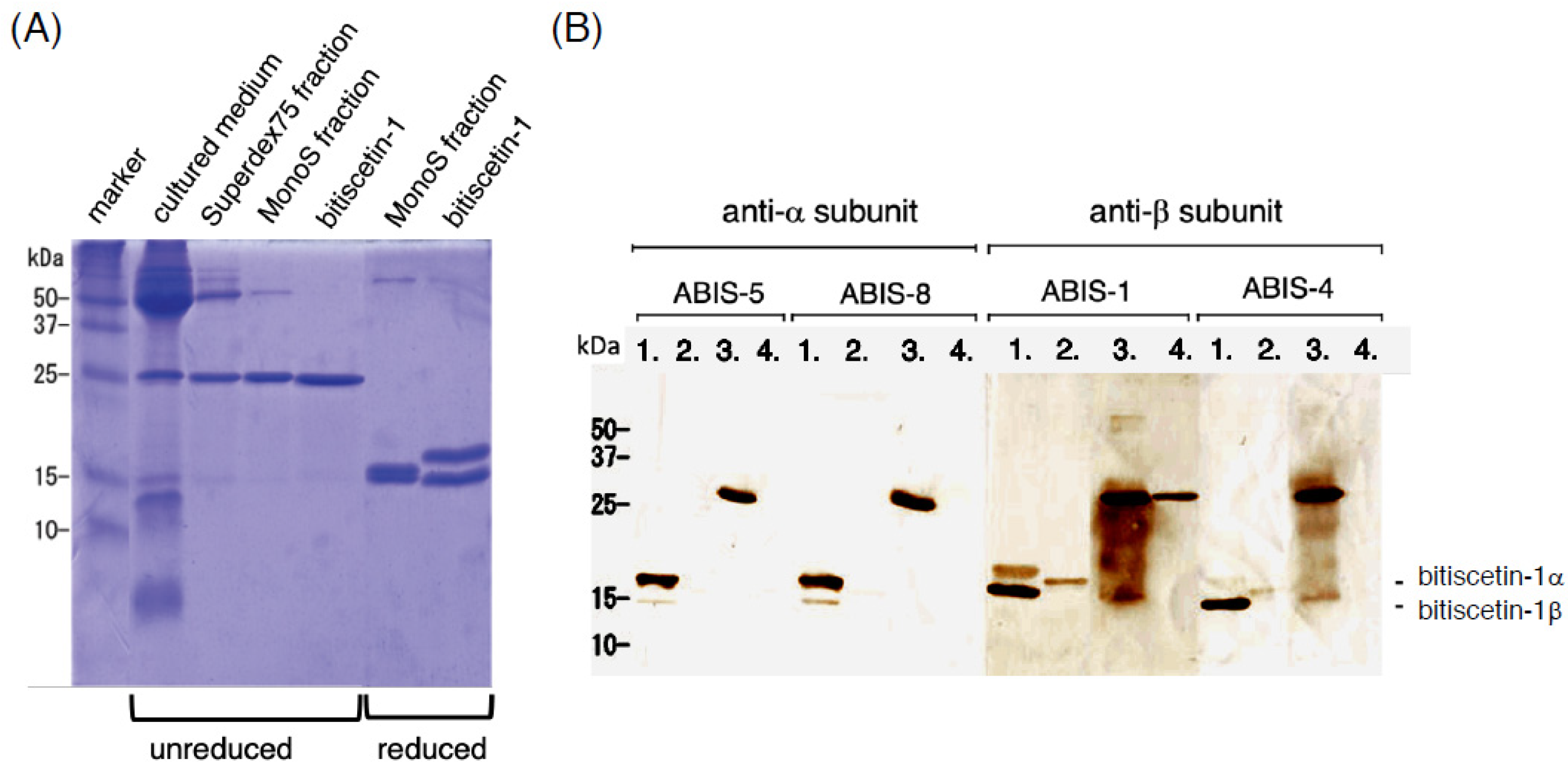
3.4. Recombinant Expression of Bitiscetin-3
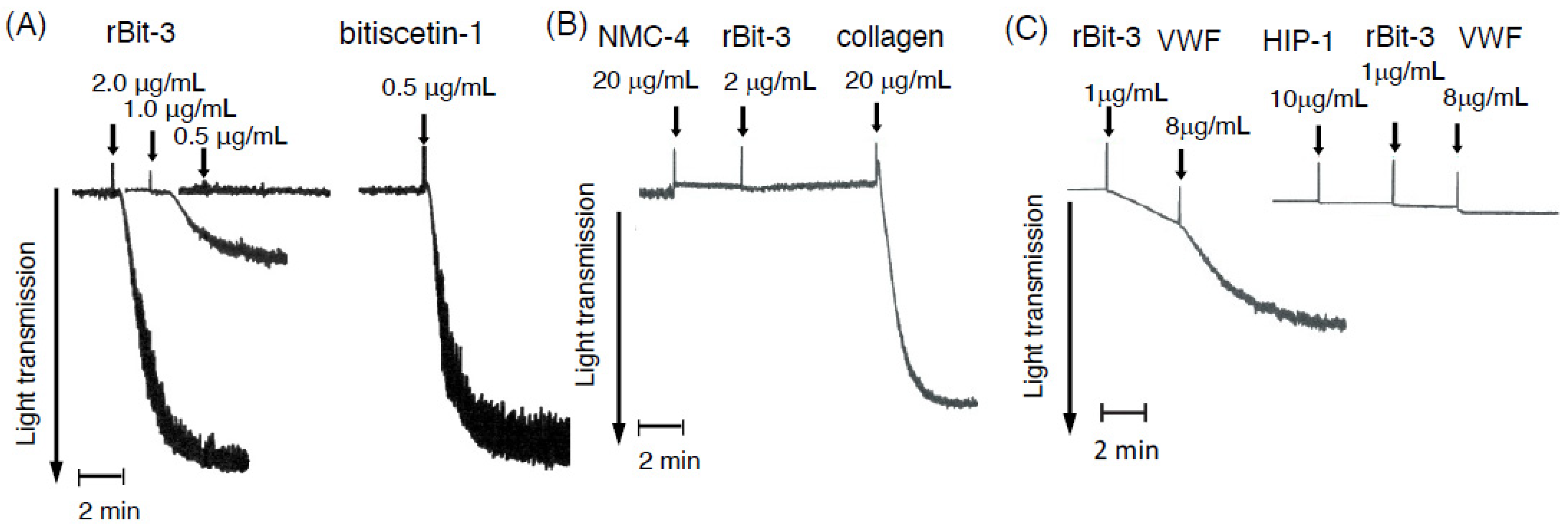
3.5. rBit-3 Induces Platelet Agglutination in Platelet-Rich Plasma, Which Can Be Blocked by Anti-VWF mAb NMC-4
3.6. rBit-3 Can Induce Agglutination of Washed Platelets, but Does So More Efficiently after the Addition of VWF
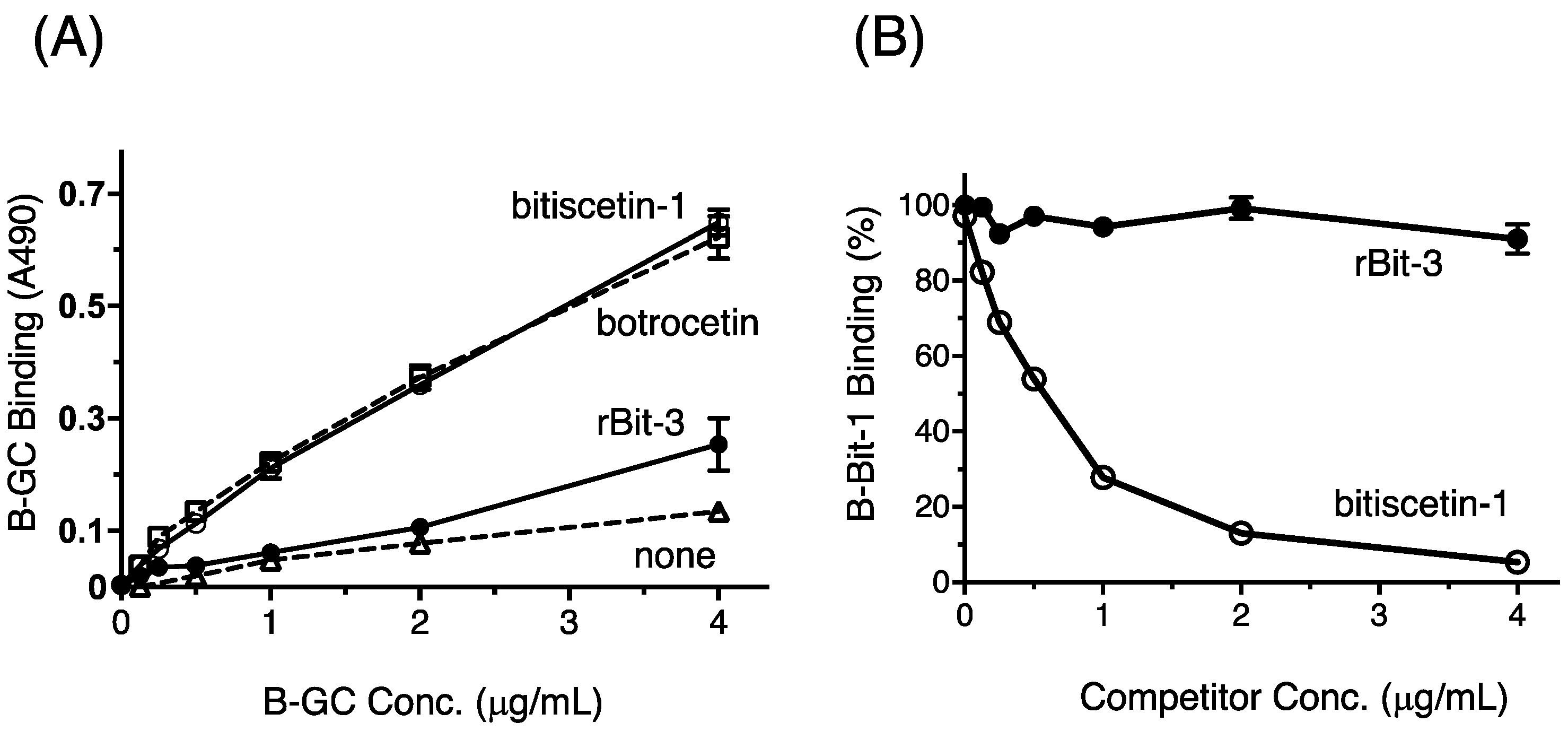
3.7. rBit-3 Enhances Binding between GPIb and VWF, but Not as Efficient as Bitiscetin-1 or Botrocetin
3.8. rBit-3 Does Not Compete with Bitiscetin-1 for Binding to VWF
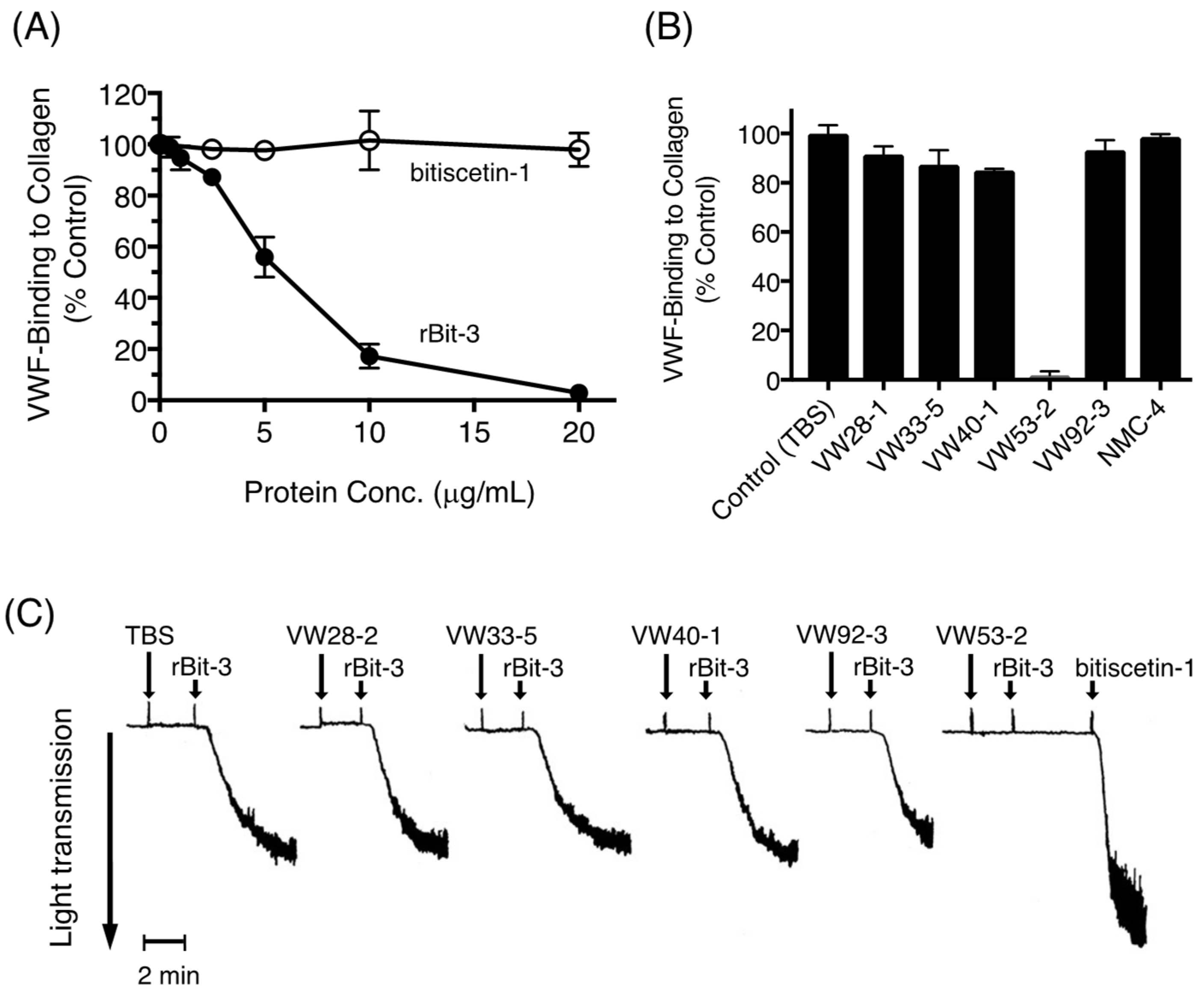
3.9. Effects of rBit-3 on the Collagen-VWF Interaction
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kini, R.M. Toxins for decoding interface selectivity in nicotinic acetylcholine receptors. Biochem. J. 2019, 476, 1515–1520. [Google Scholar] [CrossRef] [PubMed]
- Marsh, N.; Williams, V. Practical applications of snake venom toxins in haemostasis. Toxicon 2005, 45, 1171–1181. [Google Scholar] [CrossRef] [PubMed]
- Clemetson, K.J.; Morita, T.; Kini, R.M. Classification and nomenclature of snake venom C-type lectins and related proteins. Toxicon 2009, 54, 83. [Google Scholar] [CrossRef] [PubMed]
- Markland, F.S.; Kettner, C.; Schiffman, S.; Shaw, E.; Bajwa, S.S.; Reddy, K.N.; Kirakossian, H.; Patkos, G.B.; Theodor, I.; Pirkle, H. Kallikrein-like activity of crotalase, a snake venom enzyme that clots fibrinogen. Proc. Natl. Acad. Sci. USA 1982, 79, 1688–1692. [Google Scholar] [CrossRef] [Green Version]
- Camargo, A.C.; Ianzer, D.; Guerreiro, J.R.; Serrano, S.M. Bradykinin-potentiating peptides: Beyond captopril. Toxicon 2012, 59, 516–523. [Google Scholar] [CrossRef]
- Péterfi, O.; Boda, F.; Szabó, Z.; Ferencz, E.; Bába, L. Hypotensive Snake Venom Components—A Mini-Review. Molecules 2019, 24, 2778. [Google Scholar] [CrossRef] [Green Version]
- Morita, T. Structures and functions of snake venom CLPs (C-type lectin-like proteins) with anticoagulant-, procoagulant-, and platelet-modulating activities. Toxicon 2005, 45, 1099–1114. [Google Scholar] [CrossRef]
- Wijeyewickrema, L.C.; Berndt, M.C.; Andrews, R.K. Snake venom probes of platelet adhesion receptors and their ligands. Toxicon 2005, 45, 1051–1061. [Google Scholar] [CrossRef]
- Matsui, T.; Hamako, J.; Titani, K. Structure and function of snake venom proteins affecting platelet plug formation. Toxins 2010, 2, 10–23. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.J.; Chung, C.H.; Huang, T.F. From Discovery of Snake Venom Disintegrins to A Safer Therapeutic Antithrombotic Agent. Toxins 2019, 11, 372. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Liu, M.; Li, J.; Xu, R.; Long, C.; Li, H.; Mwangi, J.; Lu, Q.; Lai, R.; Shen, C. Snake C-Type Lectins Potentially Contribute to the Prey Immobilization in Protobothrops mucrosquamatus and Trimeresurus stejnegeri Venoms. Toxins 2020, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed Abd El-Aziz, T.; Garcia Soares, A.; Stockand, J.D. Snake Venoms in Drug Discovery: Valuable Therapeutic Tools for Life Saving. Toxins 2019, 11, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelensky, A.N.; Gready, J.E. The C-type lectin-like domain superfamily. FEBS J. 2005, 272, 6179–6217. [Google Scholar] [CrossRef] [PubMed]
- Eble, J.A. Structurally Robust and Functionally Highly Versatile-C-Type Lectin (-Related) Proteins in Snake Venoms. Toxins 2019, 11, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasoulis, T.; Isbister, G.K. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkhous, K.M.; Smith, S.V.; Read, M.S. Botrocetin and von Willebrand factor. In Hemostasis and Animal Venoms; Pirkle, H., Markland, F.S., Jr., Eds.; Marcel Dekker: New York, NY, USA, 1988; pp. 377–398. [Google Scholar]
- Usami, Y.; Fujimura, Y.; Suzuki, M.; Ozeki, Y.; Nishio, K.; Fukui, H.; Titani, K. Primary structure of two-chain botrocetin, a von Willebrand factor modulator purified from the venom of Bothrops jararaca. Proc. Natl. Acad. Sci. USA 1993, 90, 928–932. [Google Scholar] [CrossRef] [Green Version]
- Hamako, J.; Matsui, T.; Suzuki, M.; Ito, M.; Makita, K.; Fujimura, Y.; Ozeki, Y.; Titani, K. Purification and characterization of bitiscetin, a novel von Willebrand factor modulator protein from Bitis arietans snake venom. Biochem. Biophys. Res. Commun. 1996, 226, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Sadler, J.E. von Willebrand factor: Two sides of a coin. J. Thromb. Haemost. 2005, 3, 1702–1709. [Google Scholar] [CrossRef]
- Lenting, P.J.; Christophe, O.D.; Denis, C.V. von Willebrand factor biosynthesis, secretion, and clearance: Connecting the far ends. Blood 2015, 125, 2019–2028. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, T.; Meyer, D.; Sadler, J.E. Localization of von willebrand factor-binding sites for platelet glycoprotein Ib and botrocetin by charged-to-alanine scanning mutagenesis. J. Biol. Chem. 2000, 275, 11044–11049. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Hamako, J.; Matsushita, T.; Nakayama, T.; Fujimura, Y.; Titani, K. Binding site on human von Willebrand factor of bitiscetin, a snake venom-derived platelet aggregation inducer. Biochemistry 2002, 41, 7939–7946. [Google Scholar] [CrossRef] [PubMed]
- Read, M.S.; Shermer, R.W.; Brinkhous, K.M. Venom coagglutinin: An activator of platelet aggregation dependent on von Willebrand factor. Proc. Natl. Acad. Sci. USA 1978, 75, 4514–4518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkhous, K.M.; Reddick, R.L.; Read, M.S.; Nichols, T.C.; Bellinger, D.A.; Griggs, T.R. von Willebrand factor and animal models: Contributions to gene therapy, thrombotic thrombocytopenic purpura, and coronary artery thrombosis. Mayo Clin. Proc. 1991, 66, 733–742. [Google Scholar] [CrossRef]
- Maita, N.; Nishio, K.; Nishimoto, E.; Matsui, T.; Shikamoto, Y.; Morita, T.; Sadler, J.E.; Mizuno, H. Crystal structure of von Willebrand factor A1 domain complexed with snake venom, bitiscetin: Insight into glycoprotein Ibalpha binding mechanism induced by snake venom proteins. J. Biol. Chem. 2003, 278, 37777–37781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, K.; Doggett, T.; Laurenzi, I.J.; Liddington, R.C.; Diacovo, T.G. The snake venom protein botrocetin acts as a biological brace to promote dysfunctional platelet aggregation. Nat. Struct. Mol. Biol. 2005, 12, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Obert, B.; Houllier, A.; Meyer, D.; Girma, J.P. Conformational changes in the A3 domain of von Willebrand factor modulate the interaction of the A1 domain with platelet glycoprotein Ib. Blood 1999, 93, 1959–1968. [Google Scholar] [CrossRef] [PubMed]
- Obert, B.; Romijn, R.A.; Houllier, A.; Huizinga, E.G.; Girma, J.P. Characterization of bitiscetin-2, a second form of bitiscetin from the venom of Bitis arietans: Comparison of its binding site with the collagen-binding site on the von Willebrand factor A3-domain. J. Thromb. Haemost. 2006, 4, 1596–1601. [Google Scholar] [CrossRef]
- Chopek, M.W.; Girma, J.P.; Fujikawa, K.; Davie, E.W.; Titani, K. Human von Willebrand factor: A multivalent protein composed of identical subunits. Biochemistry 1986, 25, 3146–3155. [Google Scholar] [CrossRef]
- Fujimura, Y.; Usami, Y.; Titani, K.; Niinomi, K.; Nishio, K.; Takase, T.; Yoshioka, A.; Fukui, H. Studies on anti-von Willebrand factor (vWF) monoclonal antibody NMC-4, which inhibits both ristocetin- and botrocetin-induced vWF binding to platelet glycoprotein Ib. Blood 1991, 77, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Dijkstra, J.M. A method for making alignments of related protein sequences that share very little similarity; shark interleukin 2 as an example. Immunogenetics 2021, 73, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Korber, B. HIV Signature and Sequence Variation Analysis. In Computational Analysis of HIV Molecular Sequences; Rodrigo, A.G., Learn, G.H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 55–72. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, T.; Hamako, J.; Suzuki, M.; Hayashi, N.; Ito, M.; Makita, K.; Fujimura, Y.; Ozeki, Y.; Titani, K. Complete amino acid sequence of bitiscetin, a novel von Willebrand factor modulator protein, purified from snake venom of Bitis arietans. Res. Commun. Biochem. Cell Mol. Biol. 1997, 1, 271–284. [Google Scholar]
- Ito, M.; Hamako, J.; Sakurai, Y.; Matsumoto, M.; Fujimura, Y.; Suzuki, M.; Hashimoto, K.; Titani, K.; Matsui, T. Complete amino acid sequence of kaouthiagin, a novel cobra venom metalloproteinase with two disintegrin-like sequences. Biochemistry 2001, 40, 4503–4511. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Hori, A.; Hamako, J.; Matsushita, F.; Ozeki, Y.; Sakurai, Y.; Hayakawa, M.; Matsumoto, M.; Fujimura, Y. Mutant botrocetin-2 inhibits von Willebrand factor-induced platelet agglutination. J. Thromb. Haemost. 2017, 15, 538–548. [Google Scholar] [CrossRef]
- Harrison, R.A.; Oliver, J.; Hasson, S.S.; Bharati, K.; Theakston, R.D. Novel sequences encoding venom C-type lectins are conserved in phylogenetically and geographically distinct Echis and Bitis viper species. Gene 2003, 315, 95–102. [Google Scholar] [CrossRef]
- Whiteley, G.; Logan, R.A.; Leung, K.Y.; Newberry, F.J.; Rowley, P.D.; Dunbar, J.P.; Wagstaff, S.C.; Casewell, N.R.; Harrison, R.A. Stabilising the Integrity of Snake Venom mRNA Stored under Tropical Field Conditions Expands Research Horizons. PLoS Negl. Trop. Dis. 2016, 10, e0004615. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Khedkar, S.; Bork, P. SMART: Recent updates, new developments and status in 2020. Nucleic Acids Res. 2021, 49, D458–D460. [Google Scholar] [CrossRef]
- Hopp, T.P.; Woods, K.R. Prediction of protein antigenic determinants from amino acid sequences. Proc. Natl. Acad. Sci. USA 1981, 78, 3824–3828. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Delaney, M.K.; O’Brien, K.A.; Du, X. Signaling during platelet adhesion and activation. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 2341–2349. [Google Scholar] [CrossRef] [Green Version]
- Matsui, T.; Kunishima, S.; Hamako, J.; Katayama, M.; Kamiya, T.; Naoe, T.; Ozeki, Y.; Fujimura, Y.; Titani, K. Interaction of von Willebrand factor with the extracellular matrix and glycocalicin under static conditions. J. Biochem. 1997, 121, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Romijn, R.A.; Westein, E.; Bouma, B.; Schiphorst, M.E.; Sixma, J.J.; Lenting, P.J.; Huizinga, E.G. Mapping the collagen-binding site in the von Willebrand factor-A3 domain. J. Biol. Chem. 2003, 278, 15035–15039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daltry, J.C.; Wuster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Davies, E.L.; Arbuckle, K. Coevolution of Snake Venom Toxic Activities and Diet: Evidence that Ecological Generalism Favours Toxicological Diversity. Toxins 2019, 11, 711. [Google Scholar] [CrossRef] [Green Version]
- Huizinga, E.G.; Martijn van der Plas, R.; Kroon, J.; Sixma, J.J.; Gros, P. Crystal structure of the A3 domain of human von Willebrand factor: Implications for collagen binding. Structure 1997, 5, 1147–1156. [Google Scholar] [CrossRef] [Green Version]
- Brondijk, T.H.; Bihan, D.; Farndale, R.W.; Huizinga, E.G. Implications for collagen I chain registry from the structure of the collagen von Willebrand factor A3 domain complex. Proc. Natl. Acad. Sci. USA 2012, 109, 5253–5258. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Lin, J.; Cruz, M.A.; Dong, J.F.; Zhu, C. Force-induced cleavage of single VWFA1A2A3 tridomains by ADAMTS-13. Blood 2010, 115, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Crawley, J.T.; de Groot, R.; Xiang, Y.; Luken, B.M.; Lane, D.A. Unraveling the scissile bond: How ADAMTS13 recognizes and cleaves von Willebrand factor. Blood 2011, 118, 3212–3221. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, Y.; Fujimura, Y.; Kokubo, T.; Imamura, K.; Kawasaki, T.; Handa, M.; Suzuki, M.; Matsui, T.; Titani, K.; Yoshioka, A. The cDNA cloning and molecular characterization of a snake venom platelet glycoprotein Ib-binding protein, mamushigin, from Agkistrodon halys blomhoffii venom. Thromb. Haemost. 1998, 79, 1199–1207. [Google Scholar] [CrossRef]
- Yoshida, E.; Fujimura, Y.; Miura, S.; Sugimoto, M.; Fukui, H.; Narita, N.; Usami, Y.; Suzuki, M.; Titani, K. Alboaggregin-B and botrocetin, two snake venom proteins with highly homologous amino acid sequences but totally distinct functions on von Willebrand factor binding to platelets. Biochem. Biophys. Res. Commun. 1993, 191, 1386–1392. [Google Scholar] [CrossRef]
- Kawasaki, T.; Fujimura, Y.; Usami, Y.; Suzuki, M.; Miura, S.; Sakurai, Y.; Makita, K.; Taniuchi, Y.; Hirano, K.; Titani, K. Complete amino acid sequence and identification of the platelet glycoprotein Ib-binding site of jararaca GPIb-BP, a snake venom protein isolated from Bothrops jararaca. J. Biol. Chem. 1996, 271, 10635–10639. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nashimoto, Y.; Matsushita, F.; Dijkstra, J.M.; Nakamura, Y.; Akiyama, H.; Hamako, J.; Morita, T.; Araki, S.; Matsui, T. Bitiscetin-3, a Novel C-Type Lectin-like Protein Cloned from the Venom Gland of the Viper Bitis arietans, Induces Platelet Agglutination and Inhibits Binding of Von Willebrand Factor to Collagen. Toxins 2022, 14, 236. https://doi.org/10.3390/toxins14040236
Nashimoto Y, Matsushita F, Dijkstra JM, Nakamura Y, Akiyama H, Hamako J, Morita T, Araki S, Matsui T. Bitiscetin-3, a Novel C-Type Lectin-like Protein Cloned from the Venom Gland of the Viper Bitis arietans, Induces Platelet Agglutination and Inhibits Binding of Von Willebrand Factor to Collagen. Toxins. 2022; 14(4):236. https://doi.org/10.3390/toxins14040236
Chicago/Turabian StyleNashimoto, Youhei, Fumio Matsushita, Johannes M. Dijkstra, Yuta Nakamura, Hidehiko Akiyama, Jiharu Hamako, Takashi Morita, Satohiko Araki, and Taei Matsui. 2022. "Bitiscetin-3, a Novel C-Type Lectin-like Protein Cloned from the Venom Gland of the Viper Bitis arietans, Induces Platelet Agglutination and Inhibits Binding of Von Willebrand Factor to Collagen" Toxins 14, no. 4: 236. https://doi.org/10.3390/toxins14040236