Toxic Mechanism and Biological Detoxification of Fumonisins
by
 and
and
Linkai Qu
1,2,†, 
Lei Wang
2,†,
Hao Ji
2,†,
Yimeng Fang
2,
Pengyu Lei
2,
Xingxing Zhang
3,
Libo Jin
2,
Da Sun
2,* and
and
Hao Dong
1,2,* 1
College of Life Sciences, Jilin Agricultural University, Changchun 130118, China
2
Biomedical Collaborative Innovation Center of Zhejiang Province, Institute of Life Sciences, Wenzhou University, Wenzhou 325035, China
3
Department of Endocrinology and Metabolism, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou 325000, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Toxins 2022, 14(3), 182; https://doi.org/10.3390/toxins14030182
Submission received: 19 January 2022
/
Revised: 14 February 2022
/
Accepted: 26 February 2022
/
Published: 1 March 2022
Abstract
:Food safety is related to the national economy and people’s livelihood. Fumonisins are widely found in animal feed, feed raw materials, and human food. This can not only cause economic losses in animal husbandry but can also have carcinogenicity or teratogenicity and can be left in animal meat, eggs, and milk which may enter the human body and pose a serious threat to human health. Although there are many strategies to prevent fumonisins from entering the food chain, the traditional physical and chemical methods of mycotoxin removal have some disadvantages, such as an unstable effect, large nutrient loss, impact on the palatability of feed, and difficulty in mass production. As a safe, efficient, and environmentally friendly detoxification technology, biological detoxification attracts more and more attention from researchers and is gradually becoming an accepted technique. This work summarizes the toxic mechanism of fumonisins and highlights the advances of fumonisins in the detoxification of biological antioxidants, antagonistic microorganisms, and degradation mechanisms. Finally, the future challenges and focus of the biological control and degradation of fumonisins are discussed.
Keywords:
fumonisins; toxicity mechanism; toxins structure; biological detoxification; antioxidants detoxifyKey Contribution: This review summarizes the biological detoxification methods of fumonisins, explores the microorganisms that can degrade fumonisins and the possible toxicity mechanism of fumonisins as well as future research goals and directions.
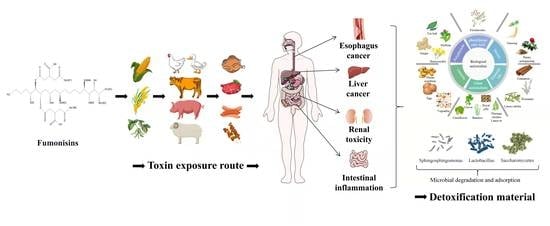
1. Introduction
Mycotoxins are toxic secondary metabolites produced by many filamentous fungi of ascomycetes [1]. Mycotoxin pollution is a persistent global problem which is inevitable and unpredictable. The production of mycotoxins is affected by the surrounding environment; even a good growth and storage environment cannot completely prevent the production of mycotoxins [2]. Fumonisins are a group of toxins that pose a significant threat to food and animal health after aflatoxins. Fumonisins have high toxicity and often appear together with aflatoxin toxicity. They cause huge economic losses to the livestock and poultry breeding industry and threaten human health [3,4]. Therefore, several studies have been exploring methods to control and alleviate fumonisin toxicity. Fumonisins easily contaminate corn, rice, and other grains, causing damage to the liver and kidneys of several animals that feed on these grains and even causing tumor problems [5,6]. In addition, fumonisin toxicity is implicated in causing human esophageal cancer and neural tube defect disease [7,8], thus fumonisins have gradually become a research hotspot after aflatoxin.
Fumonisins are a water-soluble secondary metabolite mainly produced by Fusarium verticillioides, Fusarium proliferatum, and other Fusarium species [9]. It exists on a variety of substrates, mainly on grains such as corn, and can also be found in products manufactured using grains as raw materials [5]. Fumonisins can be divided into four categories: A, B, C and P, including 28 structural analogues: FA1, FA2, FA3, PHFA3a, PHFA3b, HFA3, FAK1, FBK1, FB1, Iso-FB1, PHFB1a, PHFB1b, HFB1, FB2, FB3, FB4, FB5, FC1, N-acetyl-FC1, Iso-FC1, N-acetyl-iso-FC1, OH-FC1, N-acetyl-OH-FC1, FC3, FC4, FP1, FP2, and FP3 (Table 1). Notably, the fumonisin B family is the main and most toxic family. Fumonisin B1 (FB1) and fumonisin B2 (FB2) are the most abundant and most toxic variants that naturally contaminate maize, accounting for 70–80% and 15–25% of the total number of fumonisins [10,11].
WHO (2001) established a provisional maximum daily tolerable level of fumonisins at 2 μg/kg-BW (body weight), owing to its high levels and high toxicity [12]. The European Commission (2006 and 2007) set the maximum levels of fumonisins for unprocessed maize at 4000 μg/kg, FB at 1000 μg/kg for human corn-based foods, 800 μg/kg for corn breakfast cereals and snacks, and 200 μg/kg for corn-based baby foods [13,14]. The International Agency for Research on Cancer (IARC) classifies fumonisins into group 2B, which is a possible human carcinogen owing to their harmful effects [15]. Therefore, it is particularly significant to reduce the content and detoxify fumonisins in food.
Fumonisins are highly soluble in water and have strong thermal stability, thus they are chemically stable under various conditions. It is therefore challenging to remove them from ordinary grain processing to meet normal edible standards [16]. Physical and chemical methods cannot effectively remove fumonisins and other toxic substances from grains. Studies report that biological methods can effectively remove fumonisins in crops. Therefore, studies have widely explored the inhibition of fumonisin-producing strain growth and degradation of fumonisins through biological control and biodegradation [17,18].
In the current paper, the current prevention and control methods of fumonisin-producing strains, microorganisms that can degrade fumonisins, and biodetoxification methods were reviewed. This work will provide a reference for the biological control of fumonisin-producing strains and information on biodegradable fumonisin-producing strains.
2. Toxic Mechanism of Fumonisins
Fumonisins cause a variety of toxic effects to organisms including autophagy, apoptosis, neurotoxicity, immunotoxicity, reproductive toxicity, tissue and organ toxicity, and carcinogenicity. They can not only cause disease alone but also have a combined toxic effect with other mycotoxins such as aflatoxins. The toxicity of fumonisins is a very complex process. Previous studies report that fumonisins exert their toxicity by modulating sphingolipid metabolism and inducing oxidative stress [19,20].
2.1. Effect of Fumonisins on Sphingolipid Synthesis
Fumonisins (such as FB1) are a class of structurally similar diesters comprising different polyols and glycerol tricarboxylic acids (Figure 1). They have a similar structure to that of sphingosine (So) (Figure 2) and sphinganine (Sa) (Figure 2). Therefore, they are classified under sphingosine-like mycotoxins [21,22]. So and Sa are the main components of sphingomyelin. Sphingolipid is an important component of biofilm. Sphingolipid is involved in the regulation of several signal transduction processes such as cell proliferation, differentiation, senescence, apoptosis, and carcinogenesis. Notably, sphingolipid is the key hub of cell-to-cell recognition and interaction [23,24,25].
So and Sa are biosynthesized as condensation between palmitoyl-COA and serine as substrates under the actions of serine palmitoyltransferase, ketoreductase, dihydroceramide synthetase, dihydroceramide dehydrogenase, and other enzymes under normal physiological conditions. FB1 competes and inhibits ceramide synthetase owing to the similar structure to that of So and Sa (Figure 3). Ceramide can be produced through ab initio synthesis, or through sphingomyelin hydrolysis and sphingomyelin circulation, which is the central link of sphingomyelin metabolism. Dysregulation of ceramide synthesis affects sphingomyelin metabolism (Figure 3) [22,23], resulting in damage to the integrity of the cell membrane. Accumulation of phosphorylated products of So, Sa and So, Sa, sphingosine-1-phosphate (So-1-p) (Figure 2), and sphinganin-1-phosphate (Sa-1-p) (Figure 2) in cells can also cause cellular dysfunction [19].
The contents of Sa and So in blood and cerebrospinal fluid of pigs were significantly higher compared with those in the control group after intravenous administration of 139 nmol of FB1 or oral administration of 3425 nmol/kg-BW FB1 [21], therefore, Sa and So are biomarkers of FB1 exposure in vivo. Waes et al. reported that the content of Sa-1-p in plasma of LM/Bc mice treated with 40 μM FB1 was significantly higher relative to that of the control group [26], This dose of FB1 increases the rate of embryonic malformation and the risk of neural tube defects in pregnant LM/BC mice [27]. Moreover, Kim et al. reported that accumulation of Sa-1-p and So-1-p in serum was significant compared with that of Sa and So after intraperitoneal administration of 10 mg/kg FB1 in mice for five consecutive days [28]. In addition, the kidney is the main metabolic organ of FB1; Sa-1-p, and So-1-p produced by FB1 metabolism accumulate and last longer in the kidney relative to FB1, and So-1-p and Sa-1-p in cells can cause cellular dysfunction, implying that Sa-1-p and So-1-p can also be used as biomarkers for FB1 exposure in the body. This explains why the kidney is more vulnerable to injury compared with other organs [29,30]. Grenier et al. reported that fumonisins cause changes in the contents of Sa and So and exert a significant increase in levels of pro-inflammatory factors and Th1/Th7 in the small intestine with the increase in fumonisin concentration and feeding time [31]. Bracarense et al. reported similar findings when studying the effect of FB1 on the expression of pro-inflammatory cytokines mRNA in porcine jejunal epithelial cells. They found that the expression of IFN- γ and IL-10 in porcine jejunal epithelial cells increased significantly [32]. These findings indicate that FB1 affects sphingolipid metabolism and exerts cytotoxicity by modulating the expression of proinflammatory cytokines.
2.2. Fumonism Induces Oxidative Stress
Oxidants and antioxidants in the body are in a state of dynamic balance under physiological conditions. The body produces potentially excessive toxic aerobic free radicals, on exposure to FB1, which cannot be neutralized by antioxidants present in cells. High levels of aerobic free radicals result in lipid peroxidation, DNA oxidative damage, decreased glutathione (GSH) content, and the down-regulated expression of glutathione peroxidase (GPx) and superoxide dismutase (SOD), ultimately leading to cell tissue damage and dysfunction [33,34,35,36,37,38].
Studies report that FB1 mediates cytotoxicity partially through the induction of oxidative stress. For example, treatment of HepG2 cells with 50 μM FB1 for 0, 12, and 24 h, significantly increased the ROS content in HepG2 cells treated with FB1 compared with the level of the control group [38]. The levels of SOD-1, SOD-2, glutathione reductase (GR), and catalase (CAT) in the colon tissue of mice exposed to 2.5 mg/kg-BW FB1 for 4 consecutive days were significantly lower relative to the levels of the control group. On the contrary, expression levels of CYP450, thioredoxin, heat shock protein 70, and heat shock protein 90 were significantly higher relative to the expression levels of the control group [20]. Exposure of human SH-SY5Y neuroblastoma, rat C6 glioblastoma, and mouse GT1-7 hypothalamic cells to 0.1–100 μM FB1 for 0–144 h induced C6 and GT1-7 ROS formation in a dose-dependent manner, however, it had no significant effect on SH-SY5Y. Moreover, it downregulated GSH expression, increased malondialdehyde (MDA) production, and promoted lipid peroxidation and necrotic cell death in all cells [36].
In addition to oxidative stress, FB1 can cause damage to cell DNA. Domijan et al. observed that the apoptosis rate was significantly increased in adult male rats 48 h after administration of 5 μg/kg-BW FB1, in a dose-dependent manner. Furthermore, treatment of adult male rats with 500 μg/kg-BW FB1 induced significant DNA damage [39]. DNA damage is the basis of cell carcinogenesis. Treatment of frozen horse spermatozoa with 2.5 × 10−5 μM FB1 showed significant damage to sperm chromosomes resulting in reproductive toxicity [40]. Exposure of C6 glioma cells and p53 deleted mouse embryonic fibroblasts to FB1 significantly increased the content of MDA in C6 glioma cells, increased the apoptotic rate of C6 glioma cells, and increased the levels of 8-OH-dG and DNA fragments in C6 glioma cells in a dose-dependent manner [41]. 8-OH-dG is an important marker for DNA oxidative damage and cell carcinogenesis [42,43]. This finding indicates that FB1 leads to DNA oxidative damage. In addition, Yuan et al. reported that exposure of pig iliac endothelial cells to 50 µg/mL FB1 induced a significant increase in intracellular MDA content and a decrease in SOD, CAT, and GSH levels. Moreover, the findings showed that FB1 affects the expression of porcine vascular endothelial cells’ tight junction proteins [44]. Yu et al. conducted a subsequent study and reported that FB1 promotes cell proliferation and migration as well as induces carcinogenesis of human esophageal epithelial cells. Notably, FB1 significantly upregulates the expression of cell cycle regulatory proteins (cyclinD1 and cyclinD3) and downregulates the expression of tumor suppressor genes such as phosphatase, tensin homolog, and adenomatous polyposis, indicating that FB1 may exert its toxic or carcinogenic effects by modulating the cell cycle [45].
Oxidative stress induced by FB1 partially mediates apoptosis and autophagy. Mitogen-activated protein kinase (MAPK) is an important messenger in cells. When FB1 is administered to cells, it activates protein kinase C (PKC) and regulates the c-Jun N-terminal kinase (JNK) signal pathway through MAPK [46,47]. Administration of 2.5 mg/kg-BW FB1 to mice for 4 days induced oxidative stress and the endoplasmic reticulum release of Ca2+, resulting in JNK phosphorylation, activation of the p53 apoptosis signal pathway, upregulation of the expression of pro-apoptotic factors (PUMA and Caspase3), and the induction of apoptosis [20]. Studies report that mitochondria also play a role in apoptosis [48]. Khan et al. observed that mitochondria induce the production of cytochrome C (CytoC) by p53-activated BAX. BAX is a member of the Bcl-2 family of pro-apoptotic proteins which promotes the expression of Caspase9, as well as the expression of Caspase3, ultimately inducing cell apoptosis [49]. Notably, FB1 may induce apoptosis through the Fas/FasL pathway. FB1 promotes dysfunction of the Fas receptor and activation of the caspase8 pathway inducing Caspase3 expression and resulting in cell apoptosis [50,51]. Kim et al. reported that oxidative stress mediates the JNK pathway through the effects on the endoplasmic reticulum, as well as promotes interaction between Bcl-2 and BECN-1 to release BECN1. BECN1 is a key regulator of autophagy. BECN1 modulates expression of the autophagy-associated protein LC3-II/I to induce autophagy and promotes autophagy through the endoplasmic reticulum-mediated expression of Inositol-requiring enzyme-1-α (IRE1-α), PERK-induced ATG5, ATG7, and LC3-II/I [20]. Notably, exposure to FB1 upregulates the expression of AMP-dependent protein kinase (AMPK) and downregulates expression of mammalian target of rapamycin (mTOR) by mediating endoplasmic reticulum stress, thus inducing autophagy [38]. Moreover, tumor necrosis factor-alpha (TNF-α) plays an important role in the toxicity induced by FB1. He et al. reported that the activity of TNF-α and expression level of TNF-α mRNA in heart and lung tissues of mice increased after a subcutaneous administration of 2.25 mg/kg-BW FB1 to male and female BALB/c mice for 5 days [52]. KÓCSÓ et al. reported similar findings that FB1 upregulates TNF-α mRNA expression and increases the activity of TNF-α [53]. Chen et al. reported that FB1 upregulated the expression of TNF-α mRNA in PK-15 cells and induced apoptosis in porcine kidney cells PK-15, indicating that TNF-α can be used as a biomarker for FB1 exposure in vivo [54]. Régnier et al. observed that administration of 10 mg/kg FB1 through diet upregulated the expression of NF-κB and Interleukin-8 in the liver and jejunum. Notably, NF-κB is an important target in the TNF signal pathway, implying that TNF-α may play a role in the toxicity induced by FB1 [55]. The mechanism of fumonisin toxicity is presented in Figure 4.
3. Detoxification of Fumonisins Using Biological Antioxidants
The biological detoxification mechanism of fumonisins is mainly through the antioxidant neutralization of reactive oxygen species caused by oxidative stress, thus reducing the effects of fumonisins mediated through oxidative stress. Biological antioxidants include polyphenols, sterols, phenyl propionic acids, fat-soluble and water-soluble substances, plant essential oils, and other antioxidants (Figure 5).
3.1. Polyphenols
Curcumin is a natural polyphenolic compound extracted from the rhizome of Zingiberaceae and other plants. (Figure 6) [56]. Notably, curcumin increases ceramide concentration by stimulating de novo synthesis of ceramide, activating neutral sphingomyelinase and inhibiting the activity of sphingomyelin synthase [57]. Lloyd-Evans et al. observed that curcumin reduces the intracellular accumulation of So, sphingomyelin, glycosphingolipids, and cholesterol by restoring the intracellular calcium content. These characteristics are the main features of Niemann-PicktypeC1 disease [58], as well as features for fumonisin poisoning. Feeding chicks with curcumin nanocapsules supplemented with 600 mg/kg fumonisin and 10 mg/kg curcumin, showed protecting protective effects to the liver and an antioxidant effect, as well as reducing the level of thiobarbituric acid active substance in ROS and improving the weight gain of chicks compared with the control group [59]. Moreover, curcumin reduces PK-15 death in vitro. Administration of curcumin PK-15 cells pretreated with 50 μM FB1 showed an increase in the cell survival rate from 53.7% to 77% and a decrease in the intracellular ROS content from 97.4% to 75.5% [60]. Silymarin (SIL) is also a polyphenol with a similar effect to that of curcumin (Figure 6). Sozmen et al. reported that SIL significantly reduced hepatocyte apoptosis (p < 0.0001) and upregulated the expression of Caspase-8 and TNF-α (p < 0.0001) in BALB/c mice treated with 100 mg/kg FB1 as well as 1.5 mg/kg SIL in vivo [61]. Furthermore, Ledur et al. observed that the administration of 50 μM FB1 to PK-15 cells pretreated with 2.5 μM SIL increased the cell survival rate from 53.7% to 89.2% and decreased the intracellular ROS content from 97.4% to 34.2% [60]. Moreover, Marnewick et al. reported that tea polyphenols alleviate hepatotoxicity induced by FB1. For instance, the administration in male Fischer rats of 250 mg/kg FB1 and aqueous extracts of rooibos (Aspalathus linearis), honeybush (Cyclopia intermedia), herbal, and green and black (Camellia sinensis) teas before and after fermentation showed a significant increase in the scavenging ability of mouse liver cells to free radicals. In addition, fermented herbal teas and unfermented honeybush significantly reduced liver lipid peroxidation induced by FB1. Moreover, the three tea extracts improved the activities of CAT, GPx, and GR at varying degrees [62]. Chlorogenic acid also has an inhibitory effect on fumonisin-producing strains (Figure 6). Chlorogenic acid is a common dietary polyphenol with significant bioactivity. The inhibition rate of fumonisin-producing strains after administration of chlorogenic acid was up to 70% [63].
3.2. Sterols
Hassan et al. explored the protective effect of ginseng extract (PGE) on mice exposed to FB1, as PGE contains a lot of sterols such as ginsenosides. The findings indicated that PGE reduced fragmentation of DNA in the liver and kidney after the administration of 20 mg/kg-BW of PGE and 100 μg/kg-BW FB1 to male mice at the same time. Moreover, PGE alleviated LP changes in the liver and kidney, increased GSH level, and upregulated GPx, SOD1, and CAT mRNA expression. In addition, the GPx, SOD1, and CAT mRNA expression levels of mice in the PGE group treated with 20 mg/kg-BW of FB1 were significantly higher relative to the expression levels of mice in the blank control group [64]. Additionally, Abdel-Wahhab et al. explored the effect of red ginseng on FB1 toxicity in Sprague-Dawley rats and reported consistent findings [65]. The root extract of Panax notoginseng has an inhibitory effect on the carcinogenicity of FB1. Takao et al. administered FB1 and acetone to female SENCAR mice through a skin smear to stimulate papilloma formation. The treatment group was administered with Panax notoginseng acetone extract 1 h before each administration of FB1. The findings showed that 100% of the mice in the control group developed papilloma after 12 weeks of FB1 and acetone skin smearing, whereas only about 20% and 50% of the mice in the treatment group developed papilloma after 12 and 15 weeks, respectively [66]. Moreover, daily consumption of ginseng may have a preventive or detoxifying effect on fumonisin toxicity.
3.3. Phenylpropionic Acids
Ferulic acid is a phenyl propionic acid compound derived from Ferula feruloides (Steudel) Korovin and other plants (Figure 6). Ferulic acid at 10–25 mM significantly decreased the growth rate of Fusarium oxysporum compared with the control group (p < 0.001). In addition, fumonisin production was inhibited to a certain extent [67]. Ferulic acid can be extracted from cheap agricultural by-products, therefore, the extraction of ferulic acid from low-cost agricultural by-products can be an important source in controlling the production of fumonisins in plants [68,69].
3.4. Vitamins
Vitamin E is an important antioxidant (Figure 6). Pretreatment of mice with 25 µM vitamin E (tocopherol) for 24 h before 18 µM FB1 treatment significantly reduces FB1-induced DNA damage and apoptosis [70,71]. In addition, vitamin E can be combined with selenium, CoQ10, and L-carnitine to prepare a compound with synergistic effects. In a previous study, mice were pretreated with vitamin E (30 IU/kg), selenium (1 mg/kg), CoQ10 (30 mg/kg), and L-L-carnitine (2.8 mg/kg), then intravenously administered with 1.55 mg/kg-BW FB1. The results indicated that a combination of these antioxidants alleviated DNA damage and increased the activities of aspartate aminotransferase and alanine aminotransferase by 18% and 18%, respectively, compared with the level of mice not exposed to FB1 [33]. Oginni et al. administered juvenile catfish with vitamin E and vitamin C at the same time and observed that the decrease in nutrient content in juvenile catfish induced by FB1 was improved. Notably, the crude protein content in juvenile catfish was higher compared with that of the FB1 group (p < 0.05) [72]. Furthermore, folic acid has a protective effect on cytotoxicity induced by fumonisins. Sadler et al. reported that folic acid reduced the toxic effect of FB1 on mouse embryos and improved the growth of mouse embryos after culturing embryos with a mixture of 10 mM folic acid and 2 µM FB1, indicating that folic acid improves the toxic effect of fumonisins, however, the change was not significant [73].
3.5. Essential Oil
Essential oils are unique aromatic substances extracted from plants, mainly containing alcohols, aldehydes, phenols, acetones, terpenes, and other volatile secondary metabolites synthesized by plants [74,75]. Several types of essential oils such as Litsea cubeba, cinnamon, and ginger have been reported, and most have an inhibitory effect on bacterial growth. Pante et al. conducted an in vitro experiment and reported that Litsea cubeba essential oil inhibited mycelial development of Furium verticillioides and synthesis of FB1 and FB2. The minimum inhibitory concentration of Fusarium verticillioides was 125 µg/mL and the inhibitory effect was dose dependent. The antioxidant effect of Litsea cubeba essential oil was evaluated by DPPH and ABTS methods, showing excellent antioxidant activity [76]. Bomfim et al. reported that Rosmarinus officinalis L. essential oil (REO) had a similar effect. Administration of 300 µg/mL REO caused significant morphological changes such as bacterial cell wall rupture and cell content flow out in a dose-dependent manner [77]. In addition, Zingiber officinale essential oil (GEO) inhibits the growth of fumonisin-producing bacteria and fumonisin production. Notably, administration of 2000 µg/mL GEO and 4000 µg/mL GEO significantly inhibits the production of FB1 and FB2. The inhibition rates of ergosterol biosynthesis after administration of 4000 µg/mL and 5000 µg/mL GEO were 57% and 100%, respectively [78]. Ergosterol modulates the activity of several membrane binding enzymes [79], and the reduction of ergosterol activity could result in membrane synthesis disorders, thus exhibiting a bacteriostatic effect. Castro et al. reported similar results with minimum inhibitory concentrations of Cinnamomum zeylanicum and Cymbopogon martinii essential oils to Fusarium verticillioides at 250, 250, and 500 µg/mL, respectively [80]. Plant essential oils inhibit the growth of fumonisin-producing bacteria and fumonisin production, as well as reduce or prevent toxicity caused by fumonisins. Essential oils have a strong smell and react with some drugs, thus, embedding technology is commonly used to embed essential oils. Cinnamon essential oil embedded with whey protein effectively improved the serum levels of ALT, AST, ALP, Urea, and Uric acid, and restored the normal levels in male Sprague-Dawley male rats treated with 100 mg/kg-BW FB1. Furthermore, testosterone levels in rats were restored to normal values thus reducing reproductive toxicity. Lipid peroxidation and tumor marker TNF-α in liver and kidney tissues were improved to some extent but were not restored to normal levels [74]. Studies report that the cinnamon extract glycerol monolaurate (GML) (Figure 6) has similar effects. The levels of serum triglyceride, globulin, cholesterol, liver lipid peroxidation, SOD, and serum reactive oxygen species were restored to normal or below normal levels after chicks were fed with 400 µg/kg fumonisins and GML coated with 8 mg/kg nanomaterials. However, the body weight of chicks was not improved indicating that GML does not reduce the oxidative stress caused by fumonisins to a minimum. However, it alleviates oxidative stress caused by fumonisins and enhances the activity of glutathione S-transferase which is the enzyme responsible for liver detoxification [81].
3.6. Other Antioxidants
In addition to the above-mentioned antioxidants, several other types of antioxidants have been reported in previous studies. Domijan et al. reported that sodium copper chlorophyllin (CHL) had a protective effect on FB1-induced cell and DNA damage after administration of 100 µg/mL (CHL) (Figure 6) in combination with 20 µg/mL of FB1. Oxidative stress is the main cause of DNA damage caused by FB1, thus CHL indirectly prevented FB1-induced cell death, DNA damage, and possible carcinogenesis by preventing oxidative stress [82]. Zhao et al. conducted a study whereby indole glucosinolates (IGS) (Figure 5) were infiltrated into wild-type Col-0 plants followed by a 10 µM FB1 solution into the wild-type IGS plants and compared the results with the administration of only the FB1 solution. The findings showed that IGS inhibited FB1-induced apoptosis. IGS decomposition products produced through the action of β-glucosinolase effectively reduce the accumulation of ROS, increase the activity of antioxidant enzymes, and improve ROS scavenging ability, thus reducing FB1-induced oxidative stress and apoptosis [83]. CHL and IGS are widely distributed in green leafy vegetables, thus eating more green leafy vegetables may have a preventive effect on fumonisins toxicity.
In addition to single-component antioxidants, several compound antioxidants have been reported. Hassan et al. observed that all biochemical and cytogenetic test parameters and histological images of liver tissue were significantly improved after feeding mice with an ethanol extract of Aquilegia vulgaris L. at 10 mg/kg-BW and 200 mg/kg voronisin [84]. Gbore et al. reported that the food intake of female rabbits approached the normal level after administration of Moringa leaf meal (MLM) in combination with FB1 and the effect of MLM was dose dependent. The antioxidant effect of MLM improved the adverse effects of FB1 on nutrient utilization and growth performance of female rabbits. Notably, MLM is a cheaper alternative compared with commercial antioxidants. MLM can be used as an antidote in traditional feed to reduce the harmful effects of FB1 on domestic animal production [85].
Moreover, insect products have antioxidant effects. Several honeybee products are potential sources of natural antioxidants and can counteract the effects of oxidative stress caused by various diseases [86]. Royal jelly (RJ) contains several bioactive substances and phenolic compounds, mainly comprising flavonoids and fenac, and has antioxidant activities. Liver and kidney indexes were significantly improved when male Sprague-Dawley rats were administered with a combination of 200 mg/kg fumonisins and 150 mg/kg-BW RJ compared with the levels in mice fed with FB1 alone. Liver and kidney indexes were also restored to normal levels, indicating that RJ has a protective effect on fumonisin toxicity. Notably, the protective effect was dose dependent [87].
4. Antagonistic Microorganisms and Degradation Mechanism of Fumonisins
The use of microbial control has become a research hotspot and has a wide research prospect in the field of biological control of fumonisin-producing strains and fumonisin toxin degradation. Studies report that microorganisms produce metabolites or the microorganisms themselves have potential active substances that inhibit fumonisin production. Microorganisms including lactic acid bacteria, yeasts, Klebsiella, Bacillus, sphingomonas, and other microorganisms, exert inhibitory effects on fumonisin-producing strains and fumonisin activity.
4.1. Microbial Removal of Fumonisins and Its Mechanism
Fumonisins are a chemically stable diester compound with a structure composed of different polyhydroalcohols and propanotricarboxylic acids. The skeleton is a fat chain comprising 20 carbon atoms and two identical lipid bond side chains connected to carbon 14 and 15 atoms. The degradation of fumonisins by microorganisms that use fumonisins as the only carbon source can be divided into two steps. In the case of FB1, the tricarboxylic acid groups on carbon 14 and 15 are cleaved under the action of carboxylesterase to form hydrolyzed FB1 (HFB1). The toxicity of HFB1 is significantly lower compared with that of FB1. Further, HFB1 is degraded to 2-keto-HFB1 under the action of a transaminase enzyme [88]. Further studies indicate that FB1 is first degraded into partially hydrolyzed FB1 (PHFB1a or PHFB1b) under the action of carboxylesterase. These two metabolites are formed through the cleavage of tricarboxylic acid groups on carbon 14 or 15 of FB1. The metabolites further react to form HFB1, which further reacts under the action of transaminase enzyme to form 2-keto-HFB1 as well as N-acetyl HFB1 or 2-OP1 semi-Ketal under the action of oxidative deaminase enzyme (Figure 7) [89,90,91].
The main microorganism that degrades fumonisins is sphingomonas which is widely distributed in aquatic and terrestrial environments. Sphingomonas is widely used in mycotoxin degradation. Studies report two key genes implicated in fumonisin degradation in sphingomonas namely, MTA144 which encodes transaminase and carboxylesterase. Notably, MTA144 degradation does not depend on the aerobic environment, thus it can be carried out in an anaerobic environment [89]. Moreover, the ATCC 55552 strain contains a gene that encodes a fumonisin degradation enzyme. The gene can be highly expressed in Escherichia coli and the recombinant enzyme has deamination activity in the presence of α-ketopyruvate and a pyridoxal phosphate coenzyme, thus it can deaminate HFB1. Studies should explore the genetic basis of HFB1 deamination in bacterial ATCC 55552 to fully understand the catabolism of FB1 [90].
Zhao et al. isolated a group of bacteria named SAAS79 which efficiently degraded fumonisins from mushroom residue and were mainly composed of Pseudomonas, Delftia, Sphingobacterium, Achromobacter, and other species, whereby Pseudomonas played a significant role in FB1 degradation. The findings showed that the degradation rate of 10 µg/mL FB1 was more than 90% under a pH of 5~7 and a temperature of 28~35 °C for 24 h. Moreover, the degradation rate of intracellular enzymes on approximately 10 µg/mL FB1 was about 90% after incubation for 3 h. The flora exhibited a high degradation efficiency, mild action conditions, and a wide degradation temperature range, and could degrade FB1 at a temperature ranging from 20 to 50 °C. Therefore, it has a high potential in the feed and food industry for the effective removal of FB1 [92]. Tuppia et al. explored the effect of nine strains on degrading FB1 from silage grain with high moisture content. The findings showed that Lactobacillus brevis N195 and N197 had the highest degradation effects on FB1, with degradation rates of 33% and 30% in vitro, respectively. Notably, the degradation rate of FB1 in silage grains was more than 90%, indicating that the degradation process was accompanied by adsorption, however, the specific detoxification mechanism should be explored further [93]. Benedetti et al. reported a strain that could degrade FB1 in soil. The NCB 1492 strain that degraded FB1 was isolated from the culture medium by enrichment culture. Notably, 16SrRNA analysis showed that the strain was a member of the Delftia or Comamonas family. TLC analysis showed that the NCB 1492 strain degraded FB1 24 h after culturing it in phosphate buffer saltwater (PBS) supplemented with 0.5 mg/mL FB1. FB1 spots were not detected after culturing the strain in phosphate PBS with 0.5 mg/mL FB1. No fluorescence peak was observed in HPLC analysis after incubation at 25 °C for 2 h, indicating that the NCB 1492 strain degraded fumonisins [94].
Genetically engineered bacteria have a better degradation activity of fumonisin compared with naturally occurring microorganisms. Biomin company constructed Komagataella Pastoris engineered yeast through genetic engineering technology. This strain produces fumonisin lipase named FUMzyme® (Biomin company, Wuxi, China). FUMzyme® is used in various poultry feeds. The lowest dose of 15 enzyme activity units (U)/kg and the highest dose of 300 U/kg showed the best activity. FB1 in poultry feces decreased significantly (p < 0.001) when 250 U/kg of FUMzyme® was added to the feed. The F1 in poultry feces also decreased significantly when FUMzyme® was added to feed (p < 0.05). In addition, the ratio of Sa/So in the liver and serum in poultry was lower relative to that in the control group [95]. The bacteria constructed by genetic engineering have several advantages such as a high concentration, high yield, and controllable metabolism. Moreover, the background of basic biology and molecular genetics is fully elucidated, thus biological safety is guaranteed. These findings indicate that the construction of genetically engineered bacteria has high potential in microbial removal of fumonisins in the future.
4.2. Microbial Adsorption of Fumonisins
In addition to degradation, some microorganisms can adsorb fumonisins. The adsorption effect of these microorganisms is mainly affected by temperature, pH, and bacterial structure. High-performance liquid chromatography and liquid chromatography/mass spectrometry (LC/MS) are commonly used to detect the degradation and adsorption of mycotoxins by strains. The removal of mycotoxins by strains was verified by detecting the presence of fumonisin degradation products in co-cultures. Lactic acid bacteria are widely used as probiotics owing to their large specific surface area and complex surface structure. Lactic acid bacteria are the main bacteria used for microbial adsorption and removal of fumonisins. Furthermore, they have a broad application prospect in the control of mycotoxins.
Deepthi et al. explored the effect of feeding broilers with feed containing 1 mL (109 CFU/mL) Lactobacillus plantarum MYS6 and 200 mg/kg FB1 daily for 42 days. The findings showed that Lactobacillus plantarum MYS6 restored the levels of serum parameters such as SGOT, SGPT, creatinine, cholesterol, and triglyceride to the level of the blank control group. Moreover, this strain effectively removed ROS and H2O2 from serum and liver tissue compared with the FB1 group [96]. Lactobacillus plantarum MYS6 and its extracellular products were added to the feed, and the results indicated that the content of fumonisins in the feed decreased from the original 0.870 mg/kg to 0.505 mg/kg and 0.3125 mg/kg, after co-culture of Lactobacillus plantarum MYS6 and FB1 after 2 h and 4 h, respectively, in vitro. The removal rates of FB1 were 32.9% and 61.7% at 2 h and 4 h, respectively. LC/MS analysis showed that the corresponding peaks of fumonisin degradation products were not present after 2 and 4 h of incubation, implying that the mechanism of fumonisin removal may be physical adsorption [97]. A previous study reported that Lactobacillus plantarum ZJ8 had a high binding rate to fumonisins in an acidic environment. The binding rate of the cell wall of ZJ8 strain to FB1 and FB2 was 96.8% and 100%, respectively, at pH 4 [98]. In addition, Lactobacillus plantarum B7 and Lactobacillus pentosus X8 exhibited a high scavenging effect on fumonisins, and the binding rate was dependent on the structural integrity of bacterial cell wall peptidoglycan. High peptidoglycan integrity was correlated with high binding efficiency. When peptidoglycan was removed, the removal rate of fumonisin was greatly reduced, which was speculated to be the mechanism of adsorption. The binding rates of Lactobacillus plantarum B7 and Lactobacillus pentosus X8 to FB1 and FB2 were 52.9% and 58.0%, respectively [99]. In addition to peptidoglycan integrity, acid resistance and protease resistance play an important role in mycotoxin adsorption. Ezdini et al. reported that Lactobacillus parasitum BEJ01 isolated from Tunisian butter had a strong tolerance to acid as well as to pepsin. In a previous study, male Balb/c mice were fed with 100 μg/kg FB1 and 2 × 109 CFU/mL BEJ01 for 10 days and stopped feeding for 48 h wherein the biochemical indexes of the mice were detected. The results showed that the content of MDA and the ability of binding catalase in the liver and kidney tissue were significantly lower relative to those in the FB1 group whereas the levels were similar to those of the normal feeding group [100]. Abbès et al. explored the detoxification mechanism of Lactobacillus parasitum BEJ01. Their findings indicated that Lactobacillus parasitum BEJ01 adsorbed 67.5% of FB1 in PBS in 12 h, and the adsorption rate was time dependent. The adsorption rate of FB1 reached 88.9% at 24 h and the adsorption capacity of living bacteria was significantly higher compared with that of dead bacteria. The heating and sterilization of bacteria reduced the adsorption capacity of FB1 to 25.5% [101]. Therefore, the survival of bacteria should be ensured when adsorbing toxins.
Moreover, Saccharomyces cerevisiae is used in the scavenging of fumonisins. Armando et al. used the Plackett-Burman screening design and central combinatorial design to screen and evaluate the ability of the Saccharomyces cerevisiae RC016 strain on the removal of FB1 in vitro. The findings showed that the scavenging ability of Saccharomyces cerevisiae RC016 reached 78.66% with a concentration of FB1 at 50 μg/mL and the binding ability decreased with the decrease in FB1 concentration [102]. This implies that microbial adsorption is an important mechanism in fumonisin removal.
5. Conclusions and Perspectives
Fumonisin contamination in food and feed has posed a serious threat to economic development and public health security. How to effectively control and degrade fumonisins in food and feed has become an urgent scientific problem for human health. Chemical treatments such as alkali treatment, ozone treatment, ammonia treatment, monomethylamine, and calcium hydroxide treatment may be effective. However, the residues of these chemicals in food and feed cannot be solved economically and effectively, and the application of chemical control methods is severely limited by the problems of fungal resistance, chemical residues, and environmental pollution. Meanwhile, mycotoxin adsorbents commonly used in practical production not only adsorb mycotoxin but also adsorb small molecular substances in feed, such as biotin. In addition, the adsorption capacity and adsorption effect of various adsorbents for different toxins are different, and the combination of multiple adsorbents at the same time could achieve the desired adsorption effect, though the superimposed effect needs to be demonstrated through further experiments. Biological antioxidants have the advantages of good biodegradability, low residue, and environmental friendliness and can replace chemical synthesis agents in antibacterial and mildew control. As an important source of new green mildew inhibition agents, biological antioxidants have great development potential and broad application prospects.
In all kinds of fumonisin detoxification technologies, biological detoxification has the advantages of a good detoxification effect, no residue, and small nutritional loss. Degradation of fumonisins into less toxic substances by using proteins or enzymes produced by microorganisms and the screening of microorganisms that have an antagonistic effect on virulence strains will be the focus of future research in the field of fumonisin control. It should be noted that there are still many problems to be solved in order to realize the scale application of biological detoxification technology. The degradation of fumonisins by microorganisms is affected by enzyme and microbial concentration as well as environmental parameters. Due to the complex separation and purification process of microbial enzymes and the harsh conditions of enzyme action, there are few studies on mycotoxin recombinant enzymes and their practical production and application. Therefore, screening microorganisms that can effectively remove many kinds of fumonisins and isolating and purifying the compound detoxification enzymes secreted by them are the keys to successfully controlling fumonisins. It is also an important breakthrough point and development direction in the field of fumonisin biological detoxification research to find and screen bacteria that can degrade fumonisins, to study the characteristics of the extracellular detoxification enzyme produced by bacteria, and to clone and express the detoxification enzyme gene. In the future, modern molecular biology methods and genetic engineering methods should be considered to clone and express highly active detoxifying enzyme genes so as to achieve large-scale application as an independent or combined technology.
Author Contributions
D.S., L.J. and H.D. contributed to the conception of this review. L.Q., L.W. and H.J. analyzed the literature and wrote the manuscript. Y.F., P.L. and X.Z. completed the figure drawings. D.S., L.Q., Y.F., P.L. and H.J. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (51901160), Wenzhou Science and Technology Bureau Project (S2020005, Y2020201, Y20190123).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
We declare no conflict of interest.
Abbreviations
| FB1 | fumonisins B1 |
| FB2 | fumonisins B2 |
| BW | body weight |
| So | sphingosine |
| Sa | sphinganine |
| So-1-p | sphingosine-1-phosphate |
| Sa-1-p | sphinganin-1-phosphate |
| SPT | serine palmitoyltransfe |
| 3-KeSa | 3-keto-sphinganine |
| 3-KR | 3-keto reductase |
| dhCerS | dihydroceramide synthetase |
| CerS | ceramide synthetase |
| Cer | ceramide |
| Ser | serine |
| dhDES | dihydroceramide dehydrogenase |
| C1P | ceramide-1-phosphate |
| CERK | ceramide kinase |
| C1PP | C1P phosphatase |
| CDases | ceramidase |
| SoKs | sphingosine kinase |
| So-1-pP | sphingosine-1-phosphate phosphatase |
| PE | phosphoethanolamine |
| HE | hexadecanal |
| So-1-pL | So-1-p lyase |
| SMases | sphingomyelinase |
| SMSs | sphingomyelin synthase |
| GlySM | Glycosphingolipids |
| GBA | glucerebrosidase |
| GCS | galactosyl-ceramide synthase |
| GSH | glutathione |
| GPx | glutathione peroxidase |
| SOD | superoxide dismutase |
| GR | glutathione reductase |
| CAT | catalase |
| MDA | malondialdehyde |
| MAPK | mitogen-activated protein kinase |
| PKC | protein kinase C |
| JNK | c-Jun N-terminal kinase |
| IRE1-α | Inositol-requiring enzyme-1-α |
| AMPK | AMP-dependent protein kinase |
| mTOR | mammalian target of rapamycin |
| TNF-α | tumor necrosis factor alpha |
| CytoC | cytochrome C |
| SIL | silymarin |
| PGE | protective effect of ginseng extract |
| REO | Rosmarinus officinalis L. essential oil |
| GML | glycerol monolaurate |
| CHL | chlorophyllin |
| IGS | indoleglucosinolates |
| MLM | moringa leaf meal |
| RJ | royal jelly |
| HFB1 | hydrolyzed FB1 |
| PHFB1 | partially hydrolysed FB1 |
| PBS | phosphate buffer saltwater |
References
- Alshannaq, A.; Yu, J.-H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. IJERPH 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins Affecting Animals, Foods, Humans, and Plants: Types, Occurrence, Toxicities, Action Mechanisms, Prevention, and Detoxification Strategies—A Revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Wang, S.; Hu, X.; Su, J.; Zhang, Y.; Xie, Y.; Zhang, H.; Tang, L.; Wang, J.-S. Co-Contamination of Aflatoxin B1 and Fumonisin B1 in Food and Human Dietary Exposure in Three Areas of China. Food Addit. Contam. Part A 2011, 28, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.I.; Miller, J.D. A Concise History of Mycotoxin Research. J. Agric. Food Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef] [PubMed]
- Vanara, F.; Scarpino, V.; Blandino, M. Fumonisin Distribution in Maize Dry-Milling Products and By-Products: Impact of Two Industrial Degermination Systems. Toxins 2018, 10, 357. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Song, G.; Lim, W. Effects of Mycotoxin-Contaminated Feed on Farm Animals. J. Hazard. Mater. 2020, 389, 122087. [Google Scholar] [CrossRef]
- Gelineau-van Waes, J.; Voss, K.A.; Stevens, V.L.; Speer, M.C.; Riley, R.T. Chapter 5 Maternal Fumonisin Exposure as a Risk Factor for Neural Tube Defects. In Advances in Food and Nutrition Research; Elsevier: Amsterdam, The Netherlands, 2009; Volume 56, pp. 145–181. ISBN 978-0-12-374439-5. [Google Scholar]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public Health Impacts of Foodborne Mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Tao, B.; Pang, M.; Liu, Y.; Dong, J. Natural Occurrence of Fumonisins B1 and B2 in Maize from Three Main Maize-Producing Provinces in China. Food Control 2015, 50, 838–842. [Google Scholar] [CrossRef]
- Rheeder, J.P.; Marasas, W.F.O.; Vismer, H.F. Production of Fumonisin Analogs by Fusarium Species. Appl Env. Microbiol 2002, 68, 2101–2105. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, Z.; Cheng, Y.; Gao, C.; Guo, L.; Wang, T.; Xu, J. Sphinganine-Analog Mycotoxins (SAMs): Chemical Structures, Bioactivities, and Genetic Controls. J. Fungi 2020, 6, 312. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Fumonisins. In Safety Evaluation of Certain Mycotoxins in Food; WHO Food Additives Series 47; FAO Food and Nutrition Paper 74; WHO: Geneva, Switzerland, 2001. [Google Scholar]
- European Commission Regulation No. 1881/2006, setting maximum levels for certain contaminants in foodstuffs, 19 December 2006. Off. J. Eur. Union 2006, L364, 5–24.
- European Commission Regulation No. 1126/2007, setting maximum levels for certain contaminants in foodstuffs, 28 September 2007. Off. J. Eur. Union 2007, L255, 14–17.
- International Agency for Research on Cancer. Some Traditional Herbal Medicines, Some Mycotoxins, Naphthalene and Styrene; This Publication Represents the Views and Expert Opinions of an IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, Which Met in Lyon, 12–19 February 2002; IARC monographs on the evaluation of carcinogenic risks to humans; IARC: Lyon, France, 2002; ISBN 978-92-832-1282-9. [Google Scholar]
- Bennett, J.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Hassan, Y.; Lepp, D.; Shao, S.; Zhou, T. Strategies and Methodologies for Developing Microbial Detoxification Systems to Mitigate Mycotoxins. Toxins 2017, 9, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngolong Ngea, G.L.; Yang, Q.; Castoria, R.; Zhang, X.; Routledge, M.N.; Zhang, H. Recent Trends in Detecting, Controlling, and Detoxifying of Patulin Mycotoxin Using Biotechnology Methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2447–2472. [Google Scholar] [CrossRef]
- Riley, R.T.; Merrill, A.H. Ceramide Synthase Inhibition by Fumonisins: A Perfect Storm of Perturbed Sphingolipid Metabolism, Signaling, and Disease. J. Lipid Res. 2019, 60, 1183–1189. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Singh, M.P.; Sharma, C.; Kang, S.C. Fumonisin B1 Actuates Oxidative Stress-Associated Colonic Damage via Apoptosis and Autophagy Activation in Murine Model. J. Biochem. Mol. Toxicol. 2018, 32, e22161. [Google Scholar] [CrossRef]
- Schertz, H.; Dänicke, S.; Frahm, J.; Schatzmayr, D.; Dohnal, I.; Bichl, G.; Schwartz-Zimmermann, H.; Colicchia, S.; Breves, G.; Teifke, J.; et al. Biomarker Evaluation and Toxic Effects of an Acute Oral and Systemic Fumonisin Exposure of Pigs with a Special Focus on Dietary Fumonisin Esterase Supplementation. Toxins 2018, 10, 296. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Zhang, R.-X.; Ge, S.; Zhou, T.; Liang, Y.-K. Sphingosine Kinase AtSPHK1 Functions in Fumonisin B1-Triggered Cell Death in Arabidopsis. Plant Physiol. Biochem. 2017, 119, 70–80. [Google Scholar] [CrossRef]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and Their Metabolism in Physiology and Disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191. [Google Scholar] [CrossRef]
- Ogretmen, B. Sphingolipid Metabolism in Cancer Signalling and Therapy. Nat. Rev. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Trayssac, M.; Hannun, Y.A.; Obeid, L.M. Role of Sphingolipids in Senescence: Implication in Aging and Age-Related Diseases. J. Clin. Investig. 2018, 128, 2702–2712. [Google Scholar] [CrossRef] [PubMed]
- Gelineau-van Waes, J.; Rainey, M.A.; Maddox, J.R.; Voss, K.A.; Sachs, A.J.; Gardner, N.M.; Wilberding, J.D.; Riley, R.T. Increased Sphingoid Base-1-Phosphates and Failure of Neural Tube Closure after Exposure to Fumonisin or FTY720. Birth Defects Res. Part A Clin. Mol. Teratol. 2012, 94, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Gardner, N.M.; Riley, R.T.; Showker, J.L.; Voss, K.A.; Sachs, A.J.; Maddox, J.R.; Gelineau-van Waes, J.B. Elevated Nuclear Sphingoid Base-1-Phosphates and Decreased Histone Deacetylase Activity after Fumonisin B1 Treatment in Mouse Embryonic Fibroblasts. Toxicol. Appl. Pharmacol. 2016, 298, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Yoo, H.-S.; Lee, Y.-M.; Kie, J.-H.; Jang, S.; Oh, S. Elevation of Sphinganine 1-Phosphate as a Predictive Biomarker for Fumonisin Exposure and Toxicity in Mice. J. Toxicol. Environ. Health Part A 2006, 69, 2071–2082. [Google Scholar] [CrossRef]
- Voss, K.A.; Howard, P.C.; Riley, R.T.; Sharma, R.P.; Bucci, T.J.; Lorentzen, R.J. Carcinogenicity and Mechanism of Action of Fumonisin B1: A Mycotoxin Produced by Fusarium Moniliforme (=F. Verticillioides). Cancer Detect. Prev. 2002, 26, 1–9. [Google Scholar] [CrossRef]
- Enongene, E.N. Persistence and Reversibility of the Elevation in Free Sphingoid Bases Induced by Fumonisin Inhibition of Ceramide Synthase. Toxicol. Sci. 2002, 67, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Grenier, B.; Schwartz-Zimmermann, H.; Caha, S.; Moll, W.; Schatzmayr, G.; Applegate, T. Dose-Dependent Effects on Sphingoid Bases and Cytokines in Chickens Fed Diets Prepared with Fusarium Verticillioides Culture Material Containing Fumonisins. Toxins 2015, 7, 1253–1272. [Google Scholar] [CrossRef] [Green Version]
- Bracarense, A.-P.F.L.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.-D.; Schatzmayr, G.; Oswald, I.P. Chronic Ingestion of Deoxynivalenol and Fumonisin, Alone or in Interaction, Induces Morphological and Immunological Changes in the Intestine of Piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [Green Version]
- Atroshi, F.; Rizzo, A.; Biese, I.; Veijalainen, P.; Saloniemi, H.; Sankari, S.; Andersson, K. Fumonisin B1-Introduce DNA Damage in Liver and Spleen: Effects of Pretreatment Wuth Coenzyme. Pharmacol. Res. 1999, 40, 9. [Google Scholar] [CrossRef]
- Wang, X.; Wu, Q.; Wan, D.; Liu, Q.; Chen, D.; Liu, Z.; Martínez-Larrañaga, M.R.; Martínez, M.A.; Anadón, A.; Yuan, Z. Fumonisins: Oxidative Stress-Mediated Toxicity and Metabolism in Vivo and in Vitro. Arch. Toxicol. 2016, 90, 81–101. [Google Scholar] [CrossRef] [PubMed]
- Sheik Abdul, N.; Marnewick, J.L. Fumonisin B1-induced Mitochondrial Toxicity and Hepatoprotective Potential of Rooibos: An Update. J. Appl. Toxicol. 2020, 40, 1602–1613. [Google Scholar] [CrossRef] [PubMed]
- Stockmann-Juvala, H.; Mikkola, J.; Naarala, J.; Loikkanen, J.; Elovaara, E.; Savolainen, K. Oxidative Stress Induced by Fumonisin B1 in Continuous Human and Rodent Neural Cell Cultures. Free Radic. Res. 2004, 38, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, T.; Pillay, Y.; Ghazi, T.; Nagiah, S.; Abdul, N.S.; Chuturgoon, A.A. Fumonisin B1-Induced Oxidative Stress Triggers Nrf2-Mediated Antioxidant Response in Human Hepatocellular Carcinoma (HepG2) Cells. Mycotoxin Res. 2019, 35, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Kang, S.C. Endoplasmic Reticulum Stress-Mediated Autophagy Activation Attenuates Fumonisin B1 Induced Hepatotoxicity in Vitro and in Vivo. Food Chem. Toxicol. 2017, 110, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Domijan, A.; Želježić, D.; Peraica, M.; Kovačević, G.; Gregorović, G.; Krstanac, Ž.; Horvatin, K.; Kalafatić, M. Early Toxic Effects of Fumonisin B1 in Rat Liver. Hum. Exp. Toxicol. 2008, 27, 895–900. [Google Scholar] [CrossRef]
- Minervini, F.; Lacalandra, G.M.; Filannino, A.; Garbetta, A.; Nicassio, M.; Dell’Aquila, M.E.; Visconti, A. Toxic Effects Induced by Mycotoxin Fumonisin B1 on Equine Spermatozoa: Assessment of Viability, Sperm Chromatin Structure Stability, ROS Production and Motility. Toxicol. Vitr. 2010, 24, 2072–2078. [Google Scholar] [CrossRef]
- Mobio, T.A.; Tavan, E.; Baudrimont, I.; Anane, R.; Carratú, M.-R.; Sanni, A.; Gbeassor, M.F.; Shier, T.W.; Narbonne, J.-F.; Creppy, E.E. Comparative Study of the Toxic Effects of Fumonisin B1 in Rat C6 Glioma Cells and P53-Null Mouse Embryo Fibroblasts. Toxicology 2003, 183, 65–75. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, C. 8-Hydroxy-2′-Deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef] [Green Version]
- Pilger, A.; Rüdiger, H.W. 8-Hydroxy-2′-Deoxyguanosine as a Marker of Oxidative DNA Damage Related to Occupational and Environmental Exposures. Int. Arch. Occup. Environ. Health 2006, 80, 1–15. [Google Scholar] [CrossRef]
- Yuan, Q.; Jiang, Y.; Fan, Y.; Ma, Y.; Lei, H.; Su, J. Fumonisin B1 Induces Oxidative Stress and Breaks Barrier Functions in Pig Iliac Endothelium Cells. Toxins 2019, 11, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Jia, B.; Liu, N.; Yu, D.; Zhang, S.; Wu, A. Fumonisin B1 Triggers Carcinogenesis via HDAC/PI3K/Akt Signalling Pathway in Human Esophageal Epithelial Cells. Sci. Total Environ. 2021, 787, 147405. [Google Scholar] [CrossRef] [PubMed]
- Gopee, N.V.; He, Q.; Sharma, R.P. Fumonisin B1-Induced Apoptosis Is Associated with Delayed Inhibition of Protein Kinase C, Nuclear Factor-ΚB and Tumor Necrosis Factor α in LLC-PK1 Cells. Chem. Biol. Interact. 2003, 146, 131–145. [Google Scholar] [CrossRef]
- Li, H.; Wang, M.; Kang, W.; Lin, Z.; Gan, F.; Huang, K. Non-Cytotoxic Dosage of Fumonisin B1 Aggravates Ochratoxin A-Induced Nephrocytotoxicity and Apoptosis via ROS-Dependent JNK/MAPK Signaling Pathway. Toxicology 2021, 457, 152802. [Google Scholar] [CrossRef]
- Burke, P.J. Mitochondria, Bioenergetics and Apoptosis in Cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef]
- Khan, R.; Phulukdaree, A.; Chuturgoon, A. Concentration-Dependent Effect of Fumonisin B1 on Apoptosis in Oesophageal Cancer Cells. Hum. Exp. Toxicol. 2018, 37, 762–771. [Google Scholar] [CrossRef]
- Bucciantini, M.; Nosi, D.; Forzan, M.; Russo, E.; Calamai, M.; Pieri, L.; Formigli, L.; Quercioli, F.; Soria, S.; Pavone, F.; et al. Toxic Effects of Amyloid Fibrils on Cell Membranes: The Importance of Ganglioside GM1. FASEB J. 2012, 26, 818–831. [Google Scholar] [CrossRef]
- Du, P.; Li, S.-J.; Ojcius, D.M.; Li, K.-X.; Hu, W.-L.; Lin, X.; Sun, A.-H.; Yan, J. A Novel Fas-Binding Outer Membrane Protein and Lipopolysaccharide of Leptospira Interrogans Induce Macrophage Apoptosis through the Fas/FasL-Caspase-8/-3 Pathway. Emerg. Microbes Infect. 2018, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Bhandari, N.; Sharma, R.P. Fumonisin B1 Alters Sphingolipid Metabolism and Tumor Necrosis Factor a Expression in Heart and Lung of Mice. Life Sci. 2002, 9, 2015–2023. [Google Scholar] [CrossRef]
- Kócsó, D.J.; Szabó-Fodor, J.; Mézes, M.; Balogh, K.; Ferenczi, S.; Szabó, A.; Bóta, B.; Kovács, M. Fumonisin B1 Exposure Increases Hsp70 Expression in the Lung and Kidney of Rats without Inducing Significant Oxidative Stress. Acta Vet. Hung. 2018, 66, 394–407. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yang, S.; Huang, S.; Yan, R.; Wang, M.; Chen, S.; Cai, J.; Long, M.; Li, P. Transcriptome Study Reveals Apoptosis of Porcine Kidney Cells Induced by Fumonisin B1 via TNF Signalling Pathway. Food Chem. Toxicol. 2020, 139, 111274. [Google Scholar] [CrossRef]
- Régnier, M.; Gourbeyre, P.; Pinton, P.; Napper, S.; Laffite, J.; Cossalter, A.; Bailly, J.; Lippi, Y.; Bertrand-Michel, J.; Bracarense, A.P.F.R.L.; et al. Identification of Signaling Pathways Targeted by the Food Contaminant FB1: Transcriptome and Kinome Analysis of Samples from Pig Liver and Intestine. Mol. Nutr. Food Res. 2017, 61, 1700433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgarbossa, A. Natural Biomolecules and Protein Aggregation: Emerging Strategies against Amyloidogenesis. IJMS 2012, 13, 17121–17137. [Google Scholar] [CrossRef] [PubMed]
- García-Seisdedos, D.; Babiy, B.; Lerma, M.; Casado, M.E.; Martínez-Botas, J.; Lasunción, M.A.; Pastor, Ó.; Busto, R. Curcumin Stimulates Exosome/Microvesicle Release in an in Vitro Model of Intracellular Lipid Accumulation by Increasing Ceramide Synthesis. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2020, 1865, 158638. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Evans, E.; Morgan, A.J.; He, X.; Smith, D.A.; Elliot-Smith, E.; Sillence, D.J.; Churchill, G.C.; Schuchman, E.H.; Galione, A.; Platt, F.M. Niemann-Pick Disease Type C1 Is a Sphingosine Storage Disease That Causes Deregulation of Lysosomal Calcium. Nat. Med. 2008, 14, 1247–1255. [Google Scholar] [CrossRef] [PubMed]
- Galli, G.M.; Griss, L.G.; Fortuoso, B.F.; Silva, A.D.; Fracasso, M.; Lopes, T.F.; Schetinger, M.R.S.; Gundel, S.; Ourique, A.F.; Carneiro, C.; et al. Feed Contaminated by Fumonisin (Fusarium Spp.) in Chicks Has a Negative Influence on Oxidative Stress and Performance, and the Inclusion of Curcumin-Loaded Nanocapsules Minimizes These Effects. Microb. Pathog. 2020, 148, 104496. [Google Scholar] [CrossRef]
- Ledur, P.C.; Santurio, J.M. Cytoprotective Effects of Curcumin and Silymarin on PK-15 Cells Exposed to Ochratoxin A, Fumonisin B1 and Deoxynivalenol. Toxicon 2020, 185, 97–103. [Google Scholar] [CrossRef]
- Sozmen, M.; Devrim, A.K.; Tunca, R.; Bayezit, M.; Dag, S.; Essiz, D. Protective Effects of Silymarin on Fumonisin B1-Induced Hepatotoxicity in Mice. J. Vet. Sci. 2014, 15, 51. [Google Scholar] [CrossRef] [Green Version]
- Marnewick, J.L.; van der Westhuizen, F.H.; Joubert, E.; Swanevelder, S.; Swart, P.; Gelderblom, W.C.A. Chemoprotective Properties of Rooibos (Aspalathus Linearis), Honeybush (Cyclopia Intermedia) Herbal and Green and Black (Camellia Sinensis) Teas against Cancer Promotion Induced by Fumonisin B1 in Rat Liver. Food Chem. Toxicol. 2009, 47, 220–229. [Google Scholar] [CrossRef]
- Atanasova-Penichon, V.; Bernillon, S.; Marchegay, G.; Lornac, A.; Pinson-Gadais, L.; Ponts, N.; Zehraoui, E.; Barreau, C.; Richard-Forget, F. Bioguided Isolation, Characterization, and Biotransformation by Fusarium Verticillioides of Maize Kernel Compounds That Inhibit Fumonisin Production. MPMI Mol. Plant Microbe Interact. 2014, 27, 1148–1158. [Google Scholar] [CrossRef] [Green Version]
- Hassan, A.M.; Abdel-Aziem, S.H.; El-Nekeety, A.A.; Abdel-Wahhab, M.A. Panax Ginseng Extract Modulates Oxidative Stress, DNA Fragmentation and up-Regulate Gene Expression in Rats Sub Chronically Treated with Aflatoxin B1 and Fumonisin B1. Cytotechnology 2015, 67, 861–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Wahhab, M.A.; Hassan, N.S.; El-Kady, A.A.; Khadrawy, Y.A.; El-Nekeety, A.A.; Mohamed, S.R.; Sharaf, H.A.; Mannaa, F.A. Red Ginseng Extract Protects against Aflatoxin B1 and Fumonisins-Induced Hepatic Pre-Cancerous Lesions in Rats. Food Chem. Toxicol. 2010, 48, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Konoshima, T.; Takasaki, M.; Tokuda, H. Anti-Carcinogenic Activity of the Roots of Panax Notoginseng. II. Biol. Pharm. Bull. 1999, 22, 1150–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrochio, L.; Cendoya, E.; Farnochi, M.C.; Massad, W.; Ramirez, M.L. Evaluation of Ability of Ferulic Acid to Control Growth and Fumonisin Production of Fusarium Verticillioides and Fusarium Proliferatum on Maize Based Media. Int. J. Food Microbiol. 2013, 167, 215–220. [Google Scholar] [CrossRef]
- Shin, H.-D.; McClendon, S.; Le, T.; Taylor, F.; Chen, R.R. A Complete Enzymatic Recovery of Ferulic Acid from Corn Residues with Extracellular Enzymes FromNeosartorya Spinosa NRRL185. Biotechnol. Bioeng. 2006, 95, 1108–1115. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E. Ferulic Acid: An Antioxidant Found Naturally in Plant Cell Walls and Feruloyl Esterases Involved in Its Release and Their Applications. Crit. Rev. Biotechnol. 2004, 24, 59–83. [Google Scholar] [CrossRef]
- Mobio, T.A.; Baudrimont, I.; Sanni, A.; Shier, T.W.; Saboureau, D.; Dano, S.D.; Ueno, Y.; Steyn, P.S.; Creppy, E.E. Prevention by Vitamin E of DNA Fragmentation and Apoptosis Induced by Fumonisin B 1 in C6 Glioma Cells. Arch. Toxicol. 2000, 74, 112–119. [Google Scholar] [CrossRef]
- Jiang, Q.; Wong, J.; Fyrst, H.; Saba, J.D.; Ames, B.N. Tocopherol or Combinations of Vitamin E Forms Induce Cell Death in Human Prostate Cancer Cells by Interrupting Sphingolipid Synthesis. Proc. Natl. Acad. Sci. USA 2004, 101, 17825–17830. [Google Scholar] [CrossRef] [Green Version]
- Oginni, O.; Gbore, F.A.; Adewole, A.M.; Eniade, A.; Adebusoye, A.J.; Abimbola, A.T.; Ajumobi, O.O. Influence of Vitamins on Flesh Yields and Proximate Compositions of Clarias Gariepinus Fed Diets Contaminated with Increasing Doses of Fumonisin B1. J. Agric. Food Res. 2020, 2, 100079. [Google Scholar] [CrossRef]
- Sadler, T.W.; Merrill, A.H.; Stevens, V.L.; Sullards, M.C.; Wang, E.; Wang, P. Prevention of Fumonisin B1-Induced Neural Tube Defects by Folic Acid. Teratology 2002, 66, 169–176. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; El-Nekeety, A.A.; Hassan, N.S.; Gibriel, A.A.Y.; Abdel-Wahhab, K.G. Encapsulation of Cinnamon Essential Oil in Whey Protein Enhances the Protective Effect against Single or Combined Sub-Chronic Toxicity of Fumonisin B1 and/or Aflatoxin B1 in Rats. Environ. Sci. Pollut. Res. 2018, 25, 29144–29161. [Google Scholar] [CrossRef] [PubMed]
- López, A.G.; Theumer, M.G.; Zygadlo, J.A.; Rubinstein, H.R. Aromatic Plants Essential Oils Activity on Fusarium Verticillioides Fumonisin B1 Production in Corn Grain. Mycopathologia 2004, 158, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Pante, G.C.; Castro, J.C.; Lini, R.S.; Romoli, J.C.Z.; de Almeida, R.T.R.; Garcia, F.P.; Nakamura, C.V.; Pilau, E.J.; de Abreu Filho, B.A.; Machinski, M. Litsea Cubeba Essential Oil: Chemical Profile, Antioxidant Activity, Cytotoxicity, Effect against Fusarium Verticillioides and Fumonisins Production. J. Environ. Sci. Health Part B 2021, 56, 387–395. [Google Scholar] [CrossRef] [PubMed]
- da Silva Bomfim, N.; Nakassugi, L.P.; Faggion Pinheiro Oliveira, J.; Kohiyama, C.Y.; Mossini, S.A.G.; Grespan, R.; Nerilo, S.B.; Mallmann, C.A.; Alves Abreu Filho, B.; Machinski, M. Antifungal Activity and Inhibition of Fumonisin Production by Rosmarinus Officinalis L. Essential Oil in Fusarium Verticillioides (Sacc.) Nirenberg. Food Chem. 2015, 166, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto-Ribeiro, M.M.G.; Grespan, R.; Kohiyama, C.Y.; Ferreira, F.D.; Mossini, S.A.G.; Silva, E.L.; de Abreu Filho, B.A.; Mikcha, J.M.G.; Machinski Junior, M. Effect of Zingiber Officinale Essential Oil on Fusarium Verticillioides and Fumonisin Production. Food Chem. 2013, 141, 3147–3152. [Google Scholar] [CrossRef] [Green Version]
- Bendaha, H.; Yu, L.; Touzani, R.; Souane, R.; Giaever, G.; Nislow, C.; Boone, C.; El Kadiri, S.; Brown, G.W.; Bellaoui, M. New Azole Antifungal Agents with Novel Modes of Action: Synthesis and Biological Studies of New Tridentate Ligands Based on Pyrazole and Triazole. Eur. J. Med. Chem. 2011, 46, 4117–4124. [Google Scholar] [CrossRef]
- Castro, J.C.; Pante, G.C.; Centenaro, B.M.; Almeida, R.T.R.D.; Pilau, E.J.; Dias Filho, B.P.; Mossini, S.A.G.; Abreu Filho, B.A.D.; Matioli, G.; Machinski Junior, M. Antifungal and Antimycotoxigenic Effects of Zingiber Officinale, Cinnamomum Zeylanicum and Cymbopogon Martinii Essential Oils against Fusarium Verticillioides. Food Addit. Contam. Part A 2020, 37, 1531–1541. [Google Scholar] [CrossRef]
- Fortuoso, B.F.; Galli, G.M.; Griss, L.G.; Armanini, E.H.; Silva, A.D.; Fracasso, M.; Mostardeiro, V.; Morsch, V.M.; Lopes, L.Q.S.; Santos, R.C.V.; et al. Effects of Glycerol Monolaurate on Growth and Physiology of Chicks Consuming Diet Containing Fumonisin. Microb. Pathog. 2020, 147, 104261. [Google Scholar] [CrossRef]
- Domijan, A.-M.; Gajski, G.; Novak Jovanović, I.; Gerić, M.; Garaj-Vrhovac, V. In Vitro Genotoxicity of Mycotoxins Ochratoxin A and Fumonisin B1 Could Be Prevented by Sodium Copper Chlorophyllin—Implication to Their Genotoxic Mechanism. Food Chem. 2015, 170, 455–462. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, J.; Liu, Y.; Miao, H.; Cai, C.; Shao, Z.; Guo, R.; Sun, B.; Jia, C.; Zhang, L.; et al. Classic Myrosinase-Dependent Degradation of Indole Glucosinolate Attenuates Fumonisin B1-Induced Programmed Cell Death in Arabidopsis. Plant J. 2015, 81, 920–933. [Google Scholar] [CrossRef]
- Hassan, A.M.; Mohamed, S.R.; El-Nekeety, A.A.; Hassan, N.S.; Abdel-Wahhab, M.A. Aquilegia Vulgaris L. Extract Counteracts Oxidative Stress and Cytotoxicity of Fumonisin in Rats. Toxicon 2010, 56, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Gbore, F.A.; Adewumi, F.H. Ameliorative Potential of Moringa Leaf Meal on Nutrient Digestibility of Rabbits Fed Fumonisin B1-Contaminated Diets. Toxicon 2021, 201, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxid. Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef]
- El-Nekeety, A.A.; El-Kholy, W.; Abbas, N.F.; Ebaid, A.; Amra, H.A.; Abdel-Wahhab, M.A. Efficacy of Royal Jelly against the Oxidative Stress of Fumonisin in Rats. Toxicon 2007, 50, 256–269. [Google Scholar] [CrossRef]
- Wu, L.Y.; Xu, Y.D.; Wang, H.M.; Sun, X.L. Research Progress on Microbial Detoxification Mechanism of Mycotoxins. Food Res. Dev. 2018, 39, 192–199. [Google Scholar] [CrossRef]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Vekiru, E.; Krska, R.; Schatzmayr, G.; Moll, W.-D.; Grabherr, R. Degradation of Fumonisin B1 by the Consecutive Action of Two Bacterial Enzymes. J. Biotechnol. 2010, 145, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Heinl, S.; Hartinger, D.; Thamhesl, M.; Schatzmayr, G.; Moll, W.-D.; Grabherr, R. An Aminotransferase from Bacterium ATCC 55552 Deaminates Hydrolyzed Fumonisin B1. Biodegradation 2011, 22, 25–30. [Google Scholar] [CrossRef]
- Blackwell, B.A.; Gilliam, J.T.; Savard, M.E.; David Miller, J.; Duvick, J.P. Oxidative Deamination of Hydrolyzed Fumonisin B1 (AP1) by Cultures OfExophiala Spinifera. Nat. Toxins 1999, 7, 31–38. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, Y.; Gong, A.; Liu, N.; Chen, S.; Zhao, X.; Li, X.; Chen, L.; Zhou, C.; Wang, J. Biodegradation of Mycotoxin Fumonisin B1 by a Novel Bacterial Consortium SAAS79. Appl. Microbiol. Biotechnol. 2019, 103, 7129–7140. [Google Scholar] [CrossRef]
- Martinez Tuppia, C.; Atanasova-Penichon, V.; Chéreau, S.; Ferrer, N.; Marchegay, G.; Savoie, J.-M.; Richard-Forget, F. Yeast and Bacteria from Ensiled High Moisture Maize Grains as Potential Mitigation Agents of Fumonisin B1: Maize Grain Silage as a Source of FB1 Degrading Microorganisms. J. Sci. Food Agric. 2017, 97, 2443–2452. [Google Scholar] [CrossRef]
- Benedetti, R.; Nazzi, F.; Locci, R.; Firrao, G. Degradation of Fumonisin B1 by a Bacterial Strain Isolated from Soil. Biodegradation 2006, 17, 31–38. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; de Lourdes Bastos, M.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Safety and Efficacy of Fumonisin Esterase (FUMzyme®) as a Technological Feed Additive for All Avian Species. EFSA J. 2016, 14, e04617. [Google Scholar] [CrossRef]
- Deepthi, B.V.; Somashekaraiah, R.; Poornachandra Rao, K.; Deepa, N.; Dharanesha, N.K.; Girish, K.S.; Sreenivasa, M.Y. Lactobacillus Plantarum MYS6 Ameliorates Fumonisin B1-Induced Hepatorenal Damage in Broilers. Front. Microbiol. 2017, 8, 2317. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, B.V.; Poornachandra Rao, K.; Chennapa, G.; Naik, M.K.; Chandrashekara, K.T.; Sreenivasa, M.Y. Antifungal Attributes of Lactobacillus Plantarum MYS6 against Fumonisin Producing Fusarium Proliferatum Associated with Poultry Feeds. PLoS ONE 2016, 11, e0155122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Qiao, Y.; Wang, X.; Pei, J.; Zheng, J.; Zhang, B. Absorption of fumonisin B1 and B2 by Lactobacillus plantarum ZJ8. Acta Microbiol. Sin. 2014, 54, 1481–1488. [Google Scholar]
- Zhao, H.; Wang, X.; Zhang, J.; Zhang, J.; Zhang, B. The Mechanism of Lactobacillus Strains for Their Ability to Remove Fumonisins B1 and B2. Food Chem. Toxicol. 2016, 97, 40–46. [Google Scholar] [CrossRef]
- Ezdini, K.; Ben Salah-Abbès, J.; Belgacem, H.; Mannai, M.; Abbès, S. Lactobacillus Paracasei Alleviates Genotoxicity, Oxidative Stress Status and Histopathological Damage Induced by Fumonisin B1 in BALB/c Mice. Toxicon 2020, 185, 46–56. [Google Scholar] [CrossRef]
- Abbès, S.; Ben Salah-Abbès, J.; Jebali, R.; Younes, R.B.; Oueslati, R. Interaction of Aflatoxin B1 and Fumonisin B1 in Mice Causes Immunotoxicity and Oxidative Stress: Possible Protective Role Using Lactic Acid Bacteria. J. Immunotoxicol. 2016, 13, 46–54. [Google Scholar] [CrossRef]
- Armando, M.R.; Galvagno, M.A.; Dogi, C.A.; Cerrutti, P.; Dalcero, A.M.; Cavaglieri, L.R. Statistical Optimization of Culture Conditions for Biomass Production of Probiotic Gut-Borne Saccharomyces Cerevisiae Strain Able to Reduce Fumonisin B1. J. Appl. Microbiol. 2013, 114, 1338–1346. [Google Scholar] [CrossRef]
Figure 1.
The basic structure of fumonisins and the structural groups of fumonisin analogs (fumonisins analogs and R side chain structures are presented in Table 1). Fumonisins have a basic skeleton composed of 20 carbons and various carboxyl groups; hydroxyl groups, as well as ester bonds, are distributed on both sides of the skeleton.
Figure 1.
The basic structure of fumonisins and the structural groups of fumonisin analogs (fumonisins analogs and R side chain structures are presented in Table 1). Fumonisins have a basic skeleton composed of 20 carbons and various carboxyl groups; hydroxyl groups, as well as ester bonds, are distributed on both sides of the skeleton.

Figure 2.
Structure of Sphingosine, Sphinganine, sphingosine-1-phosphate, and sphinganin-1-phosphate. Sphingosine and Sphinganine are the key reactants in the synthesis of ceramide and participate in the composition of the membrane structure. The structures are highly similar to the structures of fumonisins, with a long chain skeleton and amino groups on the side of the skeleton.
Figure 2.
Structure of Sphingosine, Sphinganine, sphingosine-1-phosphate, and sphinganin-1-phosphate. Sphingosine and Sphinganine are the key reactants in the synthesis of ceramide and participate in the composition of the membrane structure. The structures are highly similar to the structures of fumonisins, with a long chain skeleton and amino groups on the side of the skeleton.

Figure 3.
Schematic illustration of sphingolipid metabolism. A: The synthetic pathway of ceramide. Serine (Ser) and palmitoyl coenzyme A are condensed under the action of the serine palmitoyltransferase (SPT) enzyme to form 3-keto-Sphinganine (3-KeSa). 3-KeSO is reduced under the action of 3-keto reductase (3-KR) to produce Sphinganine (Sa). Further, Sa is converted to dihydroceramide (dhCer) under the action of dihydroceramide synthetase (dhCerS)/ceramide synthetase (CerS). Finally, dhCer produces ceramide (Cer) under the action of dihydroceramide dehydrogenase (dhDES). B, C: The circulation pathway. Ceramide is phosphorylated to produce ceramide-1-phosphate (C1P) under the action of ceramide kinase (CERK). C1P can also be phosphorylated to form Cer under the action of C1P phosphatase (C1PP). In addition, ceramide produces sphingosine (So) under the action of ceramidase (CDases), which is a reversible reaction, whereby Cer can be formed under the action of CerS. Generated So is phosphorylated to form sphingosine-1-phosphate (So-1-p) under the action of sphingosine kinase (SoKs). So-1-p is converted to So under the action of sphingosine-1-phosphate phosphatase (So-1-pP). So-1-p is pyrolyzed to phosphoethanolamine (PE) and hexadecanal (HE) under the action of So-1-p lyase (So-1-pL). D, E: The hydrolysis pathway. In the hydrolysis pathway, sphingomyelin (SM) is hydrolyzed by sphingomyelinase (SMases) to form Cer and phosphocholine, whereas Cer is catalyzed by sphingomyelin synthase (SMSs) to form SM and diacylglycerol. Glycosphingolipids (GlySM) produce Cer under the action of glucerebrosidase (GBA). Further, Cer produces glycosphingolipids under the action of glucosyl-ceramide synthase or galactosyl-ceramide synthase (GCS).
Figure 3.
Schematic illustration of sphingolipid metabolism. A: The synthetic pathway of ceramide. Serine (Ser) and palmitoyl coenzyme A are condensed under the action of the serine palmitoyltransferase (SPT) enzyme to form 3-keto-Sphinganine (3-KeSa). 3-KeSO is reduced under the action of 3-keto reductase (3-KR) to produce Sphinganine (Sa). Further, Sa is converted to dihydroceramide (dhCer) under the action of dihydroceramide synthetase (dhCerS)/ceramide synthetase (CerS). Finally, dhCer produces ceramide (Cer) under the action of dihydroceramide dehydrogenase (dhDES). B, C: The circulation pathway. Ceramide is phosphorylated to produce ceramide-1-phosphate (C1P) under the action of ceramide kinase (CERK). C1P can also be phosphorylated to form Cer under the action of C1P phosphatase (C1PP). In addition, ceramide produces sphingosine (So) under the action of ceramidase (CDases), which is a reversible reaction, whereby Cer can be formed under the action of CerS. Generated So is phosphorylated to form sphingosine-1-phosphate (So-1-p) under the action of sphingosine kinase (SoKs). So-1-p is converted to So under the action of sphingosine-1-phosphate phosphatase (So-1-pP). So-1-p is pyrolyzed to phosphoethanolamine (PE) and hexadecanal (HE) under the action of So-1-p lyase (So-1-pL). D, E: The hydrolysis pathway. In the hydrolysis pathway, sphingomyelin (SM) is hydrolyzed by sphingomyelinase (SMases) to form Cer and phosphocholine, whereas Cer is catalyzed by sphingomyelin synthase (SMSs) to form SM and diacylglycerol. Glycosphingolipids (GlySM) produce Cer under the action of glucerebrosidase (GBA). Further, Cer produces glycosphingolipids under the action of glucosyl-ceramide synthase or galactosyl-ceramide synthase (GCS).

Figure 4.
Mechanism of fumonisin toxicity. Fumonisins induce oxidative stress and mediate apoptosis, autophagy, reproductive toxicity, and promote carcinogenesis. A: The apoptosis pathway. FB1 activates PKC, which activates the JNK signaling pathway through MAPK. Meanwhile, FB1 induces oxidative stress, resulting in the release of Ca2+ from the endoplasmic reticulum; increased Ca2+ release leads to phosphorylation of JNK, which further activates the P53 apoptosis signaling pathway. P53 up-regulates the expressions of pro-apoptotic factors PUMA and Caspase3 and activates BAX, which induces increased CytoC expression. It promotes the expression of Caspase9 and Caspase3 and induces apoptosis. FB1 also leads to Fas receptor dysfunction, which activates Caspase8, and Caspase8 induces increased expression of Caspase3, leading to apoptosis; FB1 induces apoptosis by activating the TNF-α signaling pathway and upregulating NF-κB, an important target of the TNF-α signaling pathway. B: The autophagy pathway. FB1 mediates the JNK pathway through oxidative stress, promotes the interaction between BAL-2 and BECN-1, and induces increased LC3-II/I expression. Meanwhile, FB1 mediates PERK through the endoplasmic reticulum, induces ATG5/7 expression, increases LC3-II/I expression, and induces apoptosis. FB1 also up-regulates the AMP-dependent AMPK expression through endoplasmic reticulum stress and increased AMPK expression down-regulates the mTOR induced autophagy. C: Reproductive toxicity and carcinogenesis. Oxidative stress induces DNA damage which in turn induces reproductive toxicity and carcinogenesis.
Figure 4.
Mechanism of fumonisin toxicity. Fumonisins induce oxidative stress and mediate apoptosis, autophagy, reproductive toxicity, and promote carcinogenesis. A: The apoptosis pathway. FB1 activates PKC, which activates the JNK signaling pathway through MAPK. Meanwhile, FB1 induces oxidative stress, resulting in the release of Ca2+ from the endoplasmic reticulum; increased Ca2+ release leads to phosphorylation of JNK, which further activates the P53 apoptosis signaling pathway. P53 up-regulates the expressions of pro-apoptotic factors PUMA and Caspase3 and activates BAX, which induces increased CytoC expression. It promotes the expression of Caspase9 and Caspase3 and induces apoptosis. FB1 also leads to Fas receptor dysfunction, which activates Caspase8, and Caspase8 induces increased expression of Caspase3, leading to apoptosis; FB1 induces apoptosis by activating the TNF-α signaling pathway and upregulating NF-κB, an important target of the TNF-α signaling pathway. B: The autophagy pathway. FB1 mediates the JNK pathway through oxidative stress, promotes the interaction between BAL-2 and BECN-1, and induces increased LC3-II/I expression. Meanwhile, FB1 mediates PERK through the endoplasmic reticulum, induces ATG5/7 expression, increases LC3-II/I expression, and induces apoptosis. FB1 also up-regulates the AMP-dependent AMPK expression through endoplasmic reticulum stress and increased AMPK expression down-regulates the mTOR induced autophagy. C: Reproductive toxicity and carcinogenesis. Oxidative stress induces DNA damage which in turn induces reproductive toxicity and carcinogenesis.

Figure 5.
Summary diagram of biological antioxidants. Antioxidants are widely found in vegetables and other green plants, indicating that eating more green leafy vegetables may have a protective effect on fumonisin-induced toxicity.
Figure 5.
Summary diagram of biological antioxidants. Antioxidants are widely found in vegetables and other green plants, indicating that eating more green leafy vegetables may have a protective effect on fumonisin-induced toxicity.

Figure 6.
Active components of antioxidants. Hydroxyl groups are widely found in antioxidants and have a strong reducing ability; thus, they can neutralize excessive reactive oxygen species produced under oxidative stress and achieve an antioxidant effect.
Figure 6.
Active components of antioxidants. Hydroxyl groups are widely found in antioxidants and have a strong reducing ability; thus, they can neutralize excessive reactive oxygen species produced under oxidative stress and achieve an antioxidant effect.

Figure 7.
A schematic representation of the degradation process of FB1. Degradation of FB1 occurs through a three-step reaction. The structure of FB1 is gradually disintegrated after the completion of the reaction and is converted from toxic to low-toxic or non-toxic compounds.
Figure 7.
A schematic representation of the degradation process of FB1. Degradation of FB1 occurs through a three-step reaction. The structure of FB1 is gradually disintegrated after the completion of the reaction and is converted from toxic to low-toxic or non-toxic compounds.


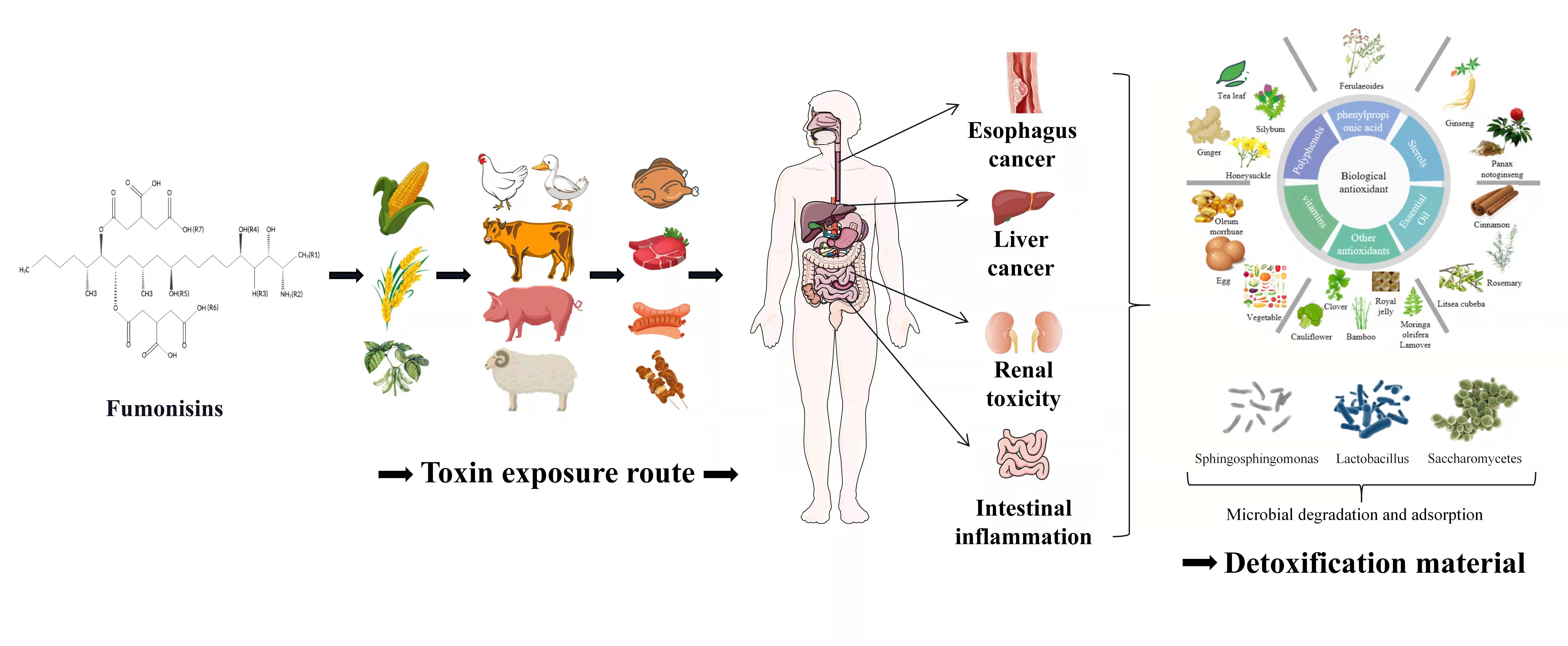{kind=link}
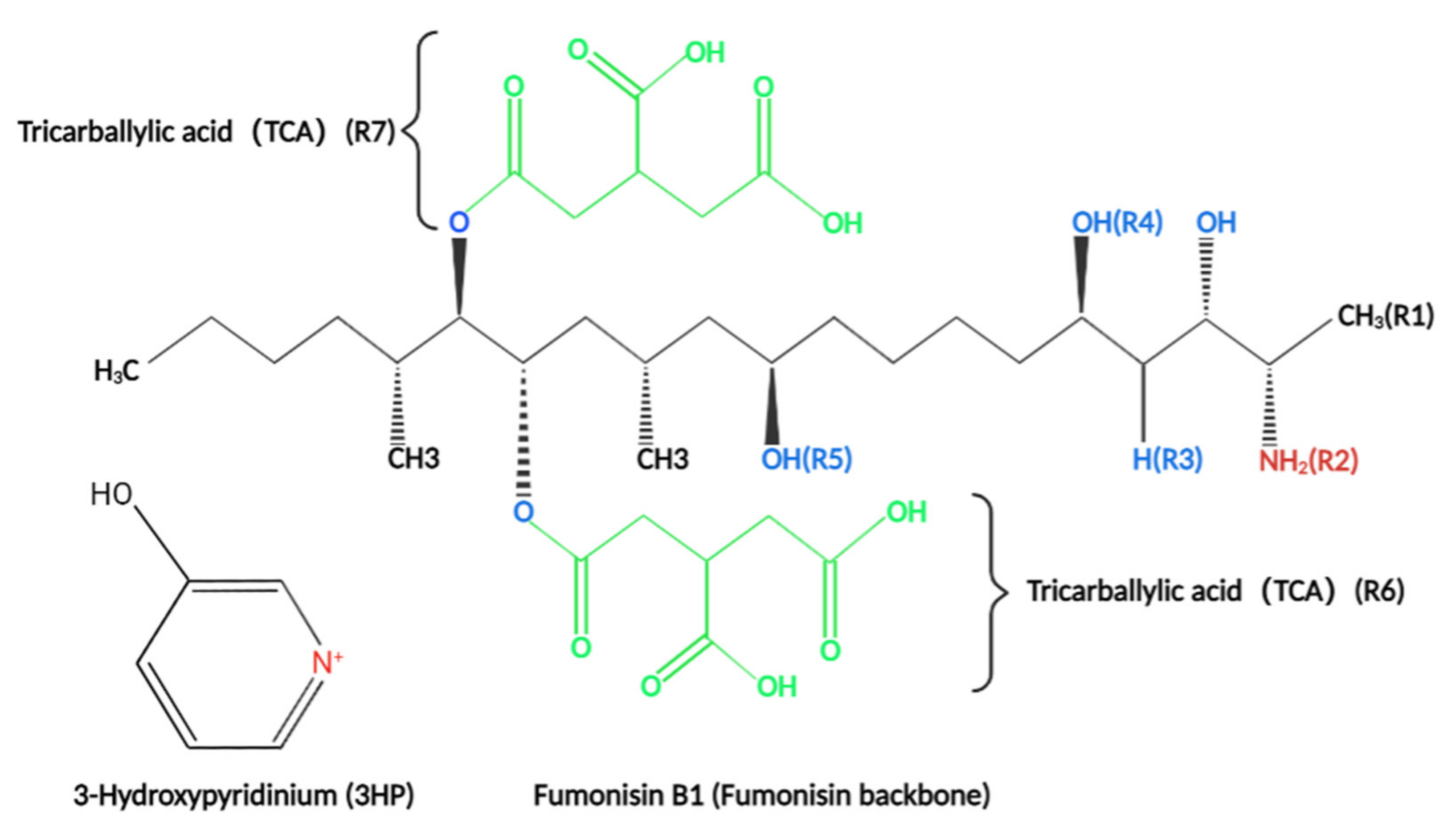{kind=link}
{kind=link}
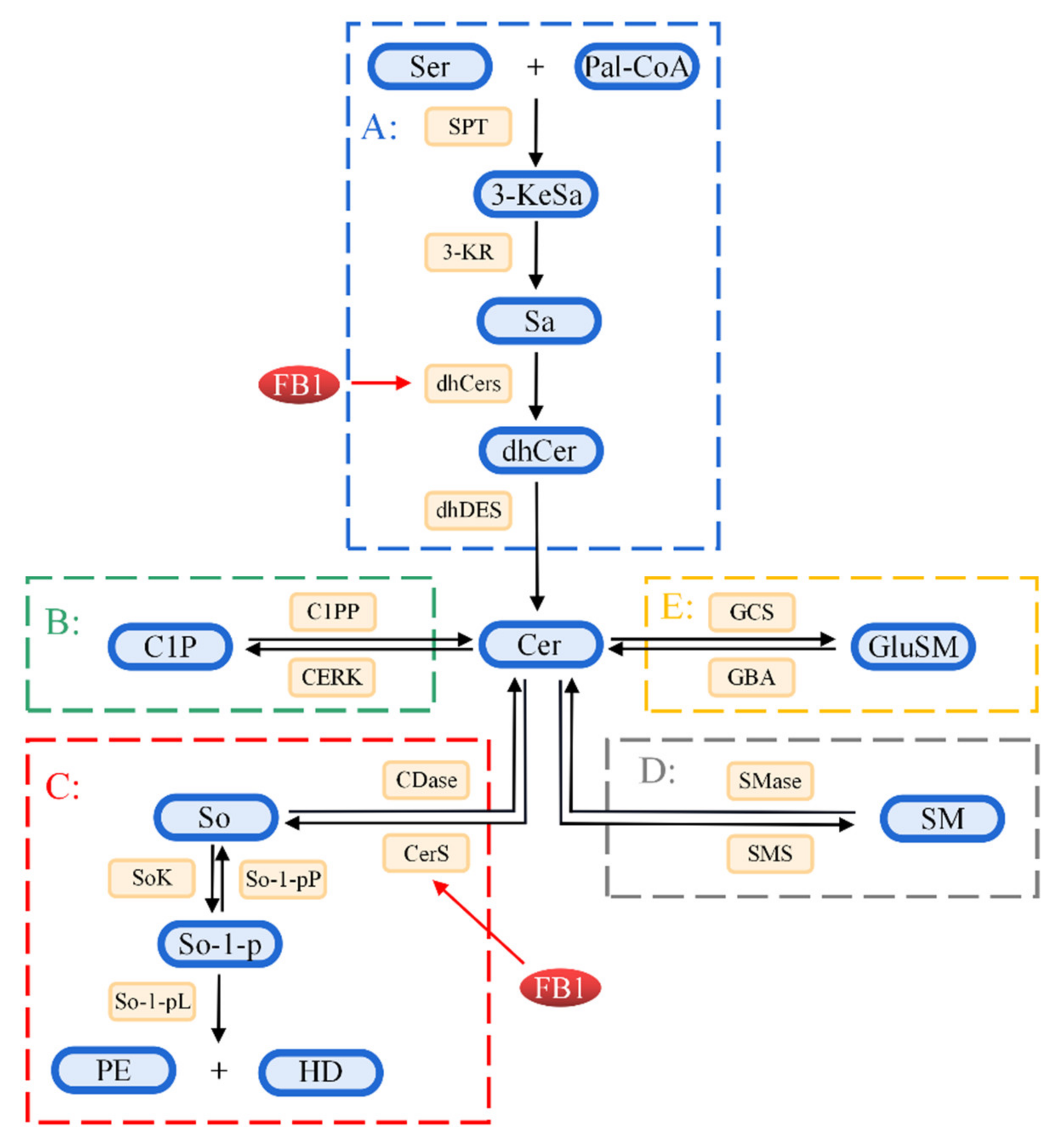{kind=link}
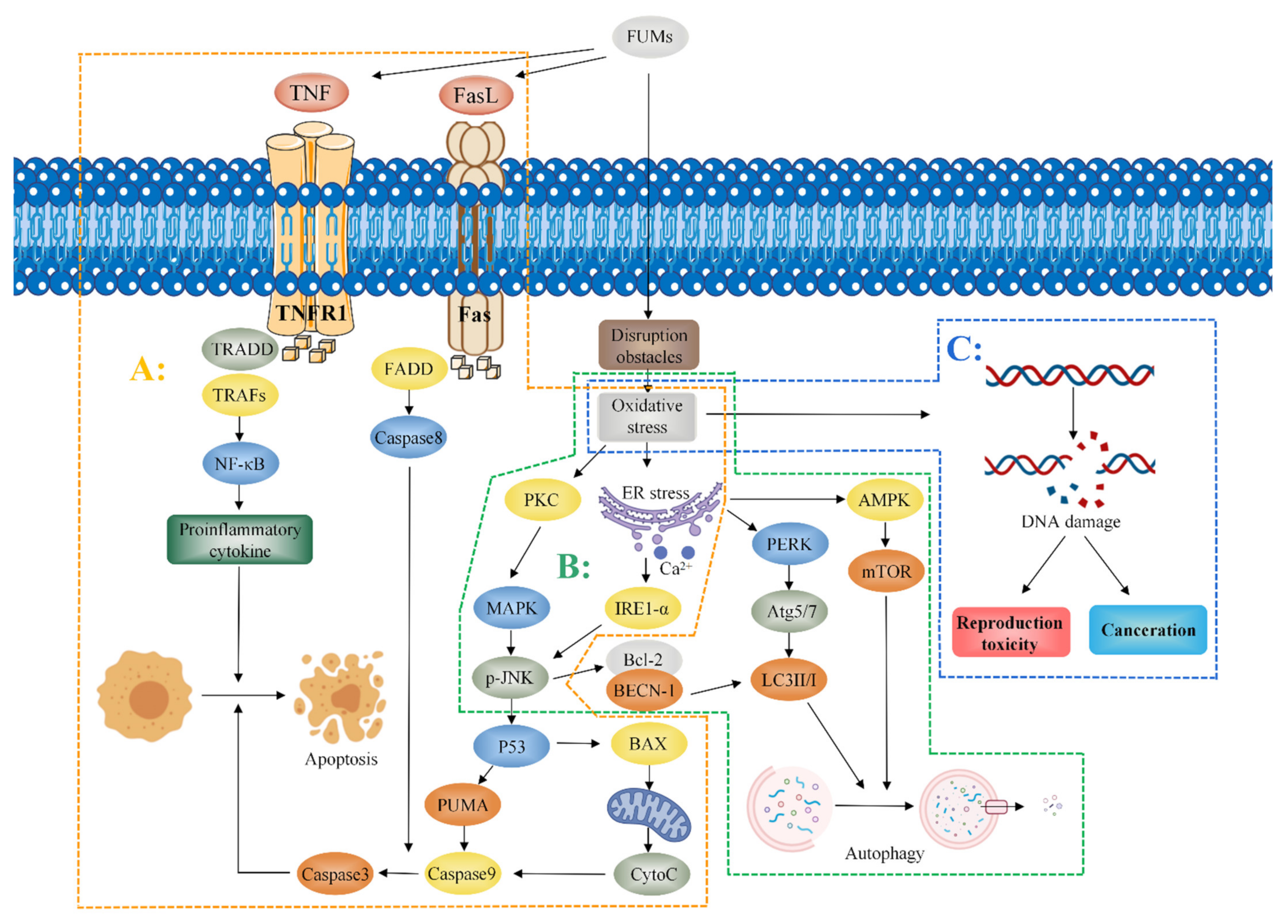{kind=link}
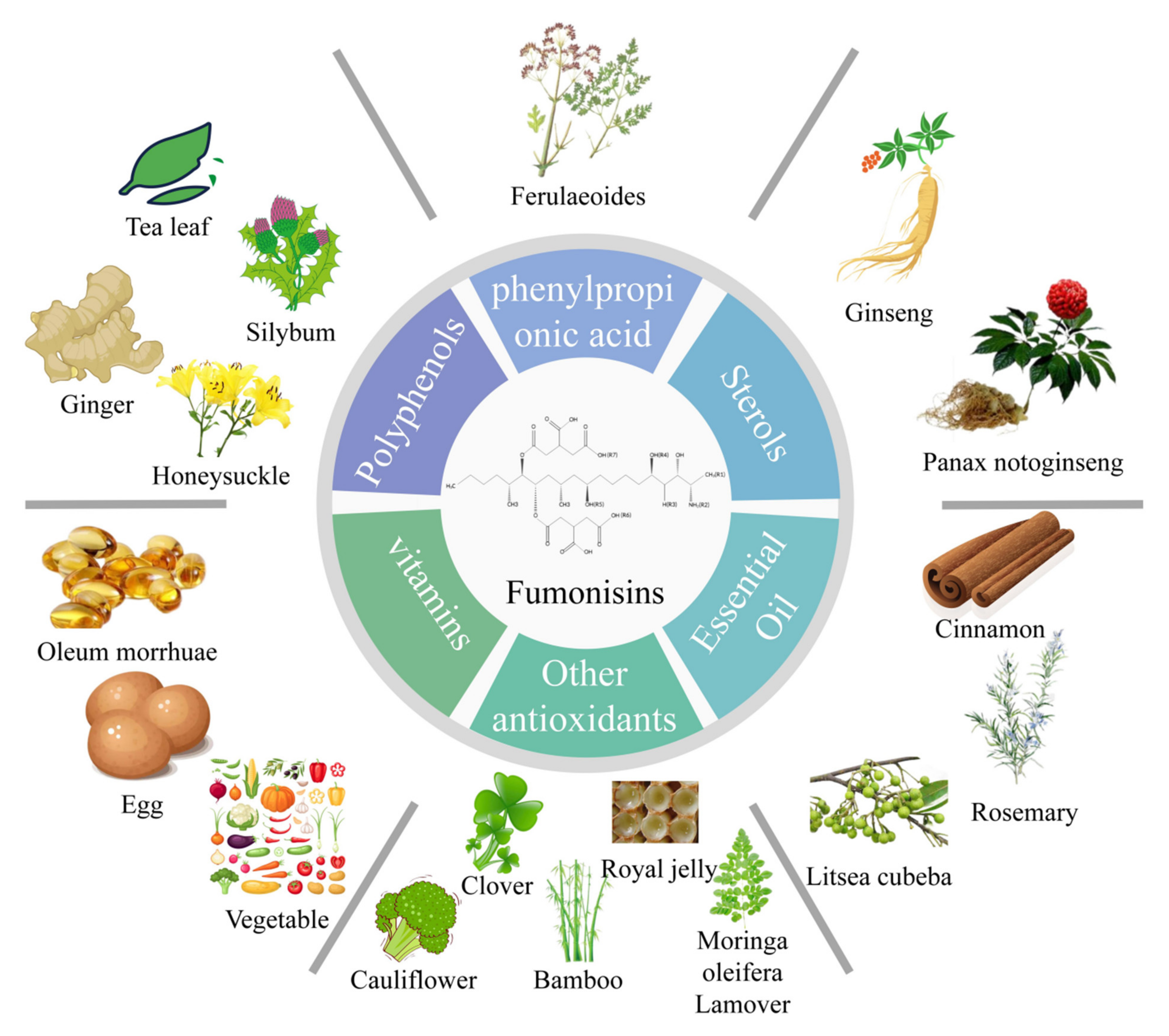{kind=link}
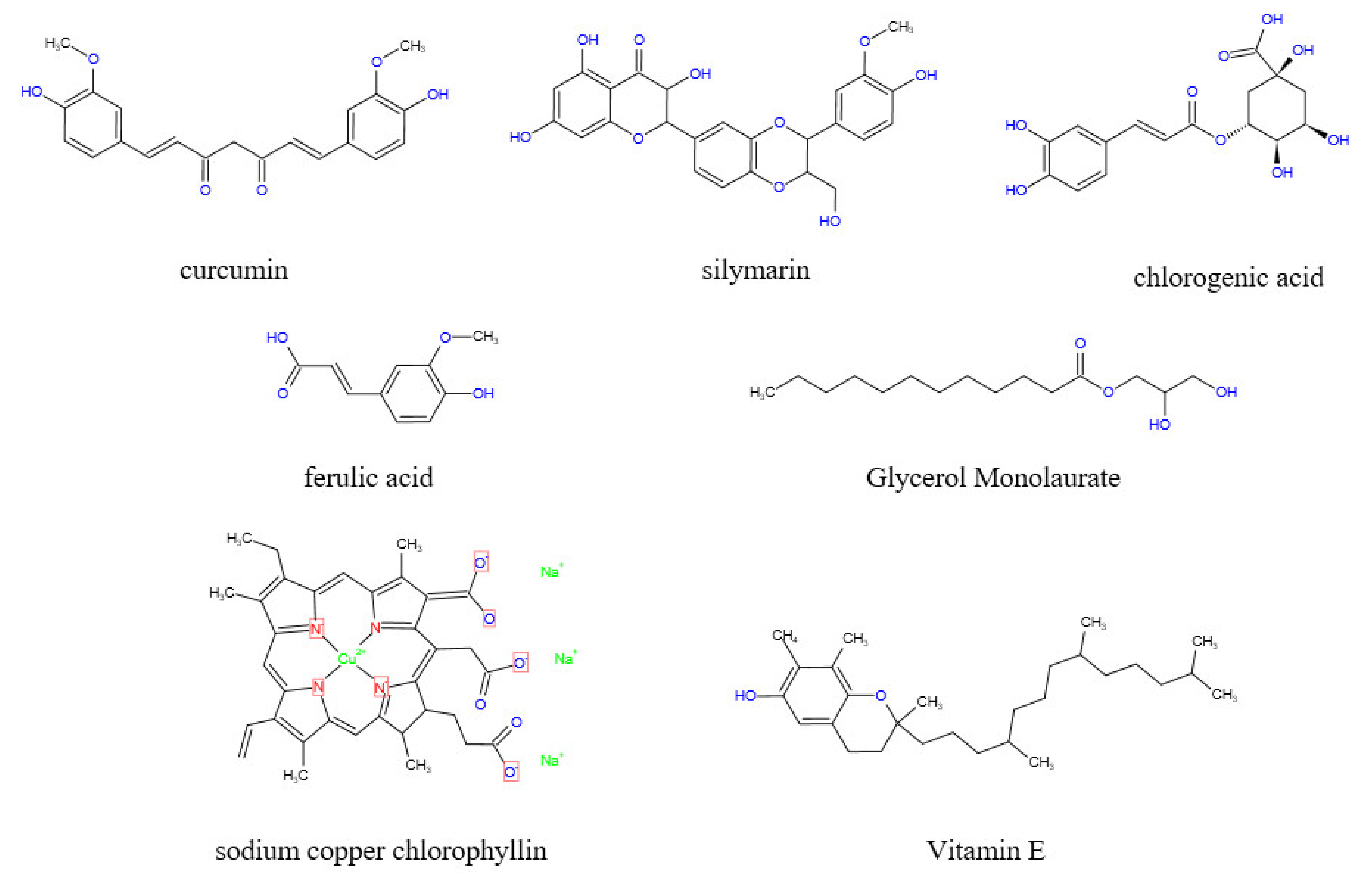{kind=link}
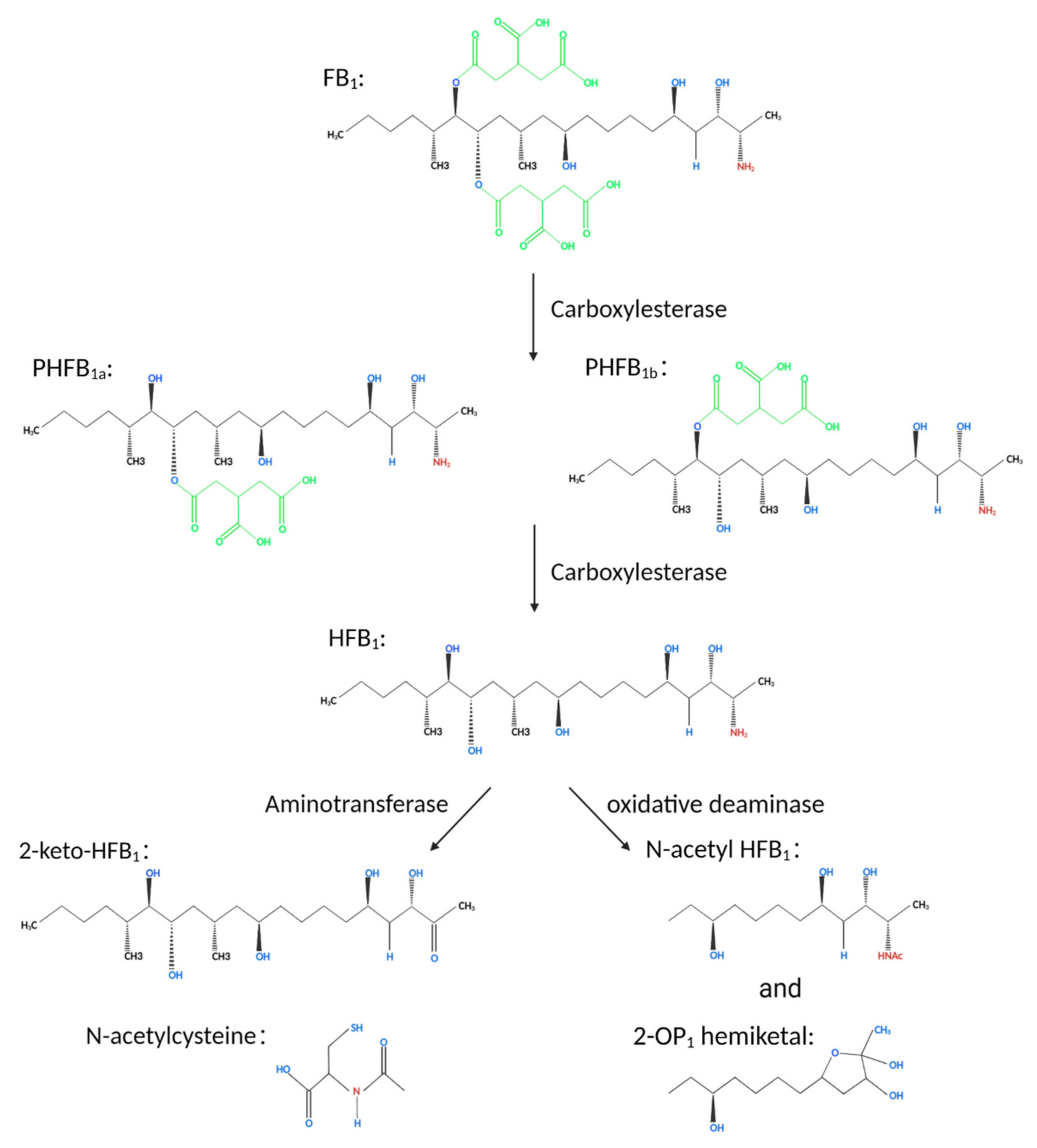{kind=link}
Table 1.
Fumonisins and their structural analogs [10].
Table 1.
Fumonisins and their structural analogs [10].
| Fumonisins | R1 | R2 | R3 | R4 | R5 | R6 | R7 |
|---|---|---|---|---|---|---|---|
| FB1 | CH3 | NH2 | H | OH | OH | TCA | TCA |
| Iso-FB1 | CH3 | NH2 | OH | H | OH | TCA | TCA |
| PHFB1a | CH3 | NH2 | H | OH | OH | OH | TCA |
| PHFB1b | CH3 | NH2 | H | OH | OH | TCA | OH |
| HFB1 | CH3 | NH2 | H | OH | OH | OH | OH |
| FB2 | CH3 | NH2 | H | OH | H | TCA | TCA |
| FB3 | CH3 | NH2 | H | H | OH | TCA | TCA |
| FB4 | CH3 | NH2 | H | H | H | TCA | TCA |
| FBK1 | CH3 | NH2 | H | H | OH | TCA | =O |
| FA1 | CH3 | NHCOCH3 | H | OH | OH | TCA | TCA |
| FA2 | CH3 | NHCOCH3 | H | OH | H | TCA | TCA |
| FA3 | CH3 | NHCOCH3 | H | H | OH | TCA | TCA |
| PHFA3a | CH3 | NHCOCH3 | H | H | OH | OH | TCA |
| PHFA3b | CH3 | NHCOCH3 | H | H | OH | TCA | OH |
| HFA3 | CH3 | NHCOCH3 | H | H | OH | OH | OH |
| FAK1 | CH3 | NHCOCH3 | H | OH | OH | TCA | =O |
| FC1 | H | NH2 | H | OH | OH | TCA | TCA |
| N-acetyl-FC1 | H | NHCOCH3 | H | OH | OH | TCA | TCA |
| Iso-FC1 | H | NH2 | OH | H | OH | TCA | TCA |
| N-acetyl-iso-FC1 | H | NHCOCH3 | OH | H | OH | TCA | TCA |
| OH-FC1 | H | NH2 | OH | OH | OH | TCA | TCA |
| N-acetyl-OH-FC1 | H | NHCOCH3 | OH | OH | OH | TCA | TCA |
| FC3 | H | NH2 | H | H | OH | TCA | TCA |
| FC4 | H | NH2 | H | H | H | TCA | TCA |
| FP1 | CH3 | 3HP | H | OH | OH | TCA | TCA |
| FP2 | CH3 | 3HP | H | OH | H | TCA | TCA |
| FP3 | CH3 | 3HP | H | H | OH | TCA | TCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qu, L.; Wang, L.; Ji, H.; Fang, Y.; Lei, P.; Zhang, X.; Jin, L.; Sun, D.; Dong, H. Toxic Mechanism and Biological Detoxification of Fumonisins. Toxins 2022, 14, 182. https://doi.org/10.3390/toxins14030182
AMA Style
Qu L, Wang L, Ji H, Fang Y, Lei P, Zhang X, Jin L, Sun D, Dong H. Toxic Mechanism and Biological Detoxification of Fumonisins. Toxins. 2022; 14(3):182. https://doi.org/10.3390/toxins14030182
Chicago/Turabian StyleQu, Linkai, Lei Wang, Hao Ji, Yimeng Fang, Pengyu Lei, Xingxing Zhang, Libo Jin, Da Sun, and Hao Dong. 2022. "Toxic Mechanism and Biological Detoxification of Fumonisins" Toxins 14, no. 3: 182. https://doi.org/10.3390/toxins14030182
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.