The Compromised Intestinal Barrier Induced by Mycotoxins
by
Yanan Gao
1,2,3,4, 
Lu Meng
1,2,3,4,
Huimin Liu
1,2,3,4,
Jiaqi Wang
1,2,3,4,* and
Nan Zheng
1,2,3,4,* 1
Key Laboratory of Quality & Safety Control for Milk and Dairy Products of Ministry of Agriculture and Rural Affairs, Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing 100193, China
2
Laboratory of Quality and Safety Risk Assessment for Dairy Products of Ministry of Agriculture and Rural Affairs, Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing 100193, China
3
Milk and Milk Products Inspection Center of Ministry of Agriculture and Rural Affairs, Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing 100193, China
4
State Key Laboratory of Animal Nutrition, Institute of Animal Sciences, Chinese Academy of Agricultural Sciences, Beijing 100193, China
*
Authors to whom correspondence should be addressed.
Toxins 2020, 12(10), 619; https://doi.org/10.3390/toxins12100619
Submission received: 7 September 2020
/
Revised: 23 September 2020
/
Accepted: 23 September 2020
/
Published: 28 September 2020
(This article belongs to the Special Issue Effects of Single and Combined Mycotoxins)
Abstract
:Mycotoxins are fungal metabolites that occur in human foods and animal feeds, potentially threatening human and animal health. The intestine is considered as the first barrier against these external contaminants, and it consists of interconnected physical, chemical, immunological, and microbial barriers. In this context, based on in vitro, ex vivo, and in vivo models, we summarize the literature for compromised intestinal barrier issues caused by various mycotoxins, and we reviewed events related to disrupted intestinal integrity (physical barrier), thinned mucus layer (chemical barrier), imbalanced inflammatory factors (immunological barrier), and dysfunctional bacterial homeostasis (microbial barrier). We also provide important information on deoxynivalenol, a leading mycotoxin implicated in intestinal dysfunction, and other adverse intestinal effects induced by other mycotoxins, including aflatoxins and ochratoxin A. In addition, intestinal perturbations caused by mycotoxins may also contribute to the development of mycotoxicosis, including human chronic intestinal inflammatory diseases. Therefore, we provide a clear understanding of compromised intestinal barrier induced by mycotoxins, with a view to potentially develop innovative strategies to prevent and treat mycotoxicosis. In addition, because of increased combinatorial interactions between mycotoxins, we explore the interactive effects of multiple mycotoxins in this review.
Key Contribution: In this context, based on in vitro, ex vivo and in vivo model, we comprehensively summarized the compromised intestinal barrier (physical, chemical, immunological, and microbial barrier) caused by various mycotoxins.
1. Introduction
Mycotoxins are the non-enzymatic poisonous metabolites produced by fungi such as Aspergillus, Penicillium, and Fusarium genera [1,2]. In recent years, approximately 500 mycotoxins derived from these fungal species and others have been identified [3]. Of these, considerable attention has been given to several common mycotoxins, which affect both human and animal health, as well as economic growth. These include aflatoxins (AFs), ochratoxin A (OTA), deoxynivalenol (DON), fumonisins (FBs), zearalenone (ZEN), patulin (PAT), nivalenol (NIV), and citrinin (CTN) [4,5]. In terms of carcinogenicity, mycotoxins have been classified into five groups by the International Agency for Research on Cancer (IARC). AFB1 and AFM1 are categorized as Group 1, which reflects their human carcinogen status. OTA and FB1 are classified as Group 2B carcinogens, whereas, DON, ZEN, PAT, and NIV are assigned to Group 3. The toxic effects, classification, and health guidance values are summarized in Table 1.
Mycotoxins are believed to be present throughout the food chain and likely occur in raw crops and crop by-products [11,12]. If livestock consume mycotoxin-contaminated ingredients, these may become distributed in their meat [13], eggs [14], and milk [15]. Hence, a predominant source of human mycotoxin exposure is derived from the ingestion of contaminated foodstuffs. Following consumption, the upper part of the intestine absorbs these mycotoxins [16,17]. Therefore, the intestinal epithelial barrier represents the first defensive barrier towards mycotoxins, suggesting this organ is more than likely to exposed to higher mycotoxin concentrations than other tissues [18,19]. The intestine is the foremost target organ of mycotoxin toxicity, with its primary role maintaining intestinal homeostasis. It is therefore vital to understand the compromised intestinal barrier mechanism induced by mycotoxins.
2. Components of the Intestinal Barrier
The intestinal barrier is composed of interconnected physical, chemical, immunological, and microbial barriers (Figure 1). As indicated, barrier function depends on the dynamic interaction of luminal microbiota, epithelial cells, and immune cells in the lamina propria (LP). A single-cell epithelial layer covers the gut wall and is pivotal to maintaining this physical barrier [20]. The intestinal epithelium is composed of four major intestinal cell types: (i) absorptive enterocytes, which account for the main cell complement, at >80% of the epithelium; (ii) protective mucin-producing goblet cells; (iii) Paneth cells, which produce antimicrobial peptides (AMPs); (iv) hormone-secreting enteroendocrine cells [21]. Epithelial cells are connected to each other from the basolateral to apical direction through interconnected protein contacts (apical junctional complex), which consist of desmosomes, adherens junctions (AJs), and tight junctions (TJs) [22]. Molecules permeate this intestinal epithelium via transcellular and paracellular routes, the latter being regulated by TJs [23,24]. Therefore, TJs selectively regulate nutrients and stimuli flux and are considered the foremost determinants of intestinal paracellular permeability [25]. TJs are multi-protein complexes, and they consist of different transmembrane proteins, e.g., junctional adhesion molecules (JAMs), claudins, and occludin and zonula occludens (ZO) proteins [26,27].
As depicted (Figure 1), the term chemical barrier refers to the mucus layer of antimicrobial-related proteins (e.g., mucins and AMPs) secreted by intestinal epithelial cells [21,28]. This barrier prevents luminal bacteria from coming into direct contact with the intestinal epithelium [29]. Mucins are the main constituents of the mucus’ thick matrix and are produced and secreted by goblet cells [30,31]. Mucin 2 (MUC2) is the most abundant protein in the intestine and plays an important role in mucus layer integrity and function [30]. In mice lacking MUC2, luminal bacteria come into direct contact with epithelial cells, thereby inducing inflammation-related diseases such as spontaneous colitis and ulcerative colitis (UC) [32]. AMPs are primarily secreted by Paneth cells, which are only located in the small intestine, while low AMP levels are secreted by enterocytes [28].
The immunological barrier comprises immune cells in LP (e.g., dendritic cells, resident macrophages, B cells, and T cells) and the secreted immune mediators (e.g., secretory immunoglobulin A (sIgA) and cytokines) [23,33,34]. SIgA is generated by B cells in the LP, transposed across the epithelium, and secreted into the mucus layer [35]. Cytokines are the important regulators in inflammatory and immune responses, and they are secreted by immune cells and epithelial intestinal cells [18].
More than 1014 commensal bacteria, known as the microbiota, comprising hundreds of different species, colonize the mammalian intestinal tract to form the intestinal microbial barrier. This microbial ecosystem plays a key role in maintaining intestinal health, as microbial dysbiosis leads to intestinal inflammation [36,37,38]. To comprehensively understand mycotoxin-mediated damage to the intestine, we systematically analyzed dysfunction mechanisms related to these physical, chemical, immunological, and microbial barriers.
3. Experimental Models Used to Assess the Intestinal Barrier
Several in vitro, ex vivo, and in vivo approaches have been used to recapitulate animal intestinal environments to assess and characterize the intestinal barrier (Table 2).
3.1. In Vitro Intestine Model
In vitro models, representing the intestinal barrier, commonly refer to intestinal epithelial cells cultured in transwell chambers with a semipermeable membrane filter. They are typically referred to as two-dimensional (2D) cultures. To date, several intestinal epithelial cell lines from different animal species and humans have been generated. Among these, the porcine intestinal epithelial cell lines, IPEC-1 and IPEC-J2, and the well-established human Caco-2 cell line are the most frequently used intestinal models for studying barrier functions [39,44]. After seeding onto transwell chambers for 1–3 weeks, these cell lines spontaneously differentiate to form polarized monolayers, representing in vitro intestinal physical and chemical barriers [52,53,54]. In addition to these models, IPI-2I and PSI-1 cells from the adult boar ileum and adult pig small intestine, respectively, have also been used as intestinal barrier models [40,55]. However, in considering the complexity interaction between different cell types, monocultures may only partially represent the intestinal barrier. Therefore, multi-culture systems are often preferred [56]. The co-culturing of polarized IPEC-J2 cells with porcine peripheral blood mononuclear cells (PBMCs), and the co-culture of Caco-2 cells and intestinal HT29-MTX cells, has been successfully applied to evaluate intestinal barrier modulation by mycotoxins [42,43,57]. While these aforementioned cell lines are commercially available, freshly isolated intestinal primary cells from various animal species are considered as more accurate models in mimicking in vivo physiology. However, they fail to achieve long-term culture. Therefore, different cell lines are chosen based on specific research purposes.
In addition to the traditional approach of 2D intestinal epithelial cell monolayers, 3D-intestinal models (enteroids), also known as “organoids” or “mini-guts”, which are derived from individual intestinal stem cells, have been developed [41,47]. Currently, based on the enteroids technique of human and mouse models, 3D culture systems have been established for various species including pig, chicken, cow, sheep, and horse [58]. Furthermore, enteroids can be cultured in a multitude of tissues including liver, stomach, colon, etc. [59]. These systems are advantageous for the following reasons: (i) enteroids harbor most, if not all, intestinal cell types [48]; (ii) it is almost impossible to accumulate mutations; (iii) enteroids can be passaged to an almost unlimited scale [45,48,49]. Although enteroids are regarded as a new exploratory intestinal model, they have several limitations: (i) when compared with the well-established and relatively cheap 2D approaches, enteroid systems are considerably costly [41,46]; (ii) the effects of molecules on the enteroid luminal side are poorly investigated [49]. To circumvent this latter hurdle, a combinatorial 2D culture and 3D enteroid approach was developed, where the enteroids were dissociated into single-cell suspensions and then cultured in the transwell chambers [60]. When compared with conventional 2D cultures, 3D enteroids partially recapitulate the anatomy of native epithelium. If researchers require better models to simulate the intestine, and the corresponding technology is available, then 3D enteroid approaches are the best choice.
3.2. Ex Vivo Intestine Model
Ex vivo studies involve the isolation of living functional tissue or organs from an organism and culturing in an Ussing chamber [61]. This model has been applied to various species, including humans [50], rodents [62], swine [51], poultry [51], and horses [63]. A main advantage of this intestinal explant culture systems is that the model maintains the histological architecture and complex intestinal barrier integrity under in vivo conditions [64]. Similarly, researchers can isolate specific segments of the gut, from the duodenum to colon, that could answer their research questions [61]. Small bowel explant cultures have been successfully used for celiac disease studies [65], and colon explants investigations have been powerful in studying the effects of intestinal microorganisms [66,67]. However, major drawbacks exist for intestinal explants, thereby restricting their application. These include short-term preservation during culture and careful and laborious preparation [39,61]. While these limitations are technical in nature, ex vivo intestinal models are widely used to evaluate gastrointestinal toxicity induced by mycotoxins [68,69,70,71,72].
3.3. In Vivo Intestine Model
In vivo studies, based on information from whole-animal systems, can effectively emulate the toxicity mechanisms in humans. The role of live animals for education and research purposes has been recognized by the World Organisation for Animal Health (OIE) [73]. In general, several animal species, such as mice, rats, chickens, turkey, fish, pigs, sheep, and cows, have been used to evaluate the toxicology of mycotoxins. When choosing a particular animal model for toxicological research, it must be borne in mind that animal species often show different susceptibilities to mycotoxins [1,74]. In much of the contemporary research on mycotoxicology, DON has been the most frequently analyzed mycotoxin. Researchers have observed that traditional in vitro systems, based on cell models, cannot comprehensively characterize pharmacokinetics, bioavailability, and in vivo metabolism, thus necessitating the requirements for in vivo animal models [75]. However, there are the drawbacks to such experimentation (i.e., time-consuming, expensive, and individual differences cannot be ignored). Additionally, from an ethical and animal welfare perspective, the use of live animals should follow the 3R (replacement, reduction, and refinement) principle. Therefore, toxicological assessments cannot be completely dependent on animal tests. Critically, experimental models should be selected based on the scientific area, e.g., in vivo models focus on the overall changes in an organism, while in vitro model investigations are ideal if particular pathways are affected by mycotoxins.
4. Intestinal Dysfunction Induced by Mycotoxins
Figure 2 shows information on an intestinal barrier compromised by mycotoxins, including disrupted intestinal integrity (physical), thinned mucus layer (chemical), imbalanced inflammatory factors (immunological), and dysfunctional bacterial homeostasis (microbial). In the following sections we discuss the effects of common mycotoxins such as AFs, OTA, DON, ZEN, FB1, PAT, and CTN.
4.1. Effects of Mycotoxins on the Physical Barrier
Numerous studies have shown that mycotoxins disrupt the intestinal physical barrier (Table 3 and Table 4). In general, intestinal epithelial cells are self-renewed every 3–5 days [76], except for Paneth cells, which undergo renewal every 18–23 days. Thus, rapidly proliferating and regenerating intestinal epithelial cells maintain the intestinal physical barrier. Besides these cells, TJ proteins also play roles in the intestinal physical barrier. A previous study has reviewed the effects on intestinal permeability induced by mycotoxins [34]; therefore, in the present study, we will pay more attention to the latest research.
4.1.1. Effects of Mycotoxins on the In Vitro Physical Barrier
Several mycotoxins modulate the intestinal physical barrier by affecting in vitro intestinal cell proliferation. AFB1 and AFM1 significantly inhibited cell growth, causing cell cycle arrest in Caco-2 cells [77,78,79,80]. In addition, AFM1 and OTA inhibited the cell viability of the co-culture of Caco-2/HT29-MTX [43]. Equally, OTA promoted apoptosis in IPEC-J2 and Caco-2 cells by inducing mitochondrial reactive oxygen species (ROS) and arresting the cell cycle [81,82,83]. Apart from mycotoxin effects in human intestinal cells, DON inhibited intestinal epithelial cell viability and induced apoptosis in rat and pig cell models [84,85,86,87,88]. ZEN-induced cell death was confirmed in IPEC-J2, IPEC-1, and human colon carcinoma cells (HCT116 cells) [89,90,91,92]. FB1, PAT, and CTN induced cell death and apoptosis in the human colon proliferating intestinal cell line (HT-29), Caco-2 cell, and HCT116 cell [93,94,95,96,98,126,127]. These studies showed that mycotoxins caused intestinal epithelial cell death, resulting in damage to the intestinal physical barrier.
Apart from cell proliferation, TJ-mediated intestinal permeability is also modulated by mycotoxins. AFM1 and OTA exposure to both apical and basolateral surfaces increased intestinal permeability, reduced TJ proteins protein expression levels, and affected the TJ protein distribution pattern [54]. These TJ protein localization effects were confirmed by transmission electron micrographs [57]. OTA treatment resulted in intestinal barrier dysfunction, which was confirmed by increased cell permeability and microvilli disruption and TJ proteins in various cell culture systems [82,105]. These observations could be explained by ROS/Ca2+-mediated myosin light chain kinase (MLCK) activation [128]. Exposure to DON decreased transepithelial electrical resistance (TEER) values and the abundance of TJs in a concentration- and time-dependent manner [106,108,109]. It has been reported that ZEN has no effect on TEER values of IPEC-1 cell, while its metabolites (α-ZOL and β-ZOL) led to time-dependent decreased TEER values [118]. FB1 and its metabolite, hydrolyzed FB1 (HFB1), damaged the intestinal integrity in different intestinal cell systems [42]. Previous studies have shown that PAT and T-2 toxin increased differentiated Caco-2 cells permeability, via the destruction of TJs, and was accompanied by MLC2 phosphorylation [94,95,122,124]. These results indicated that TJ expression levels are associated with TEER values and potentially represent a change in intestinal permeability.
4.1.2. Effects of Mycotoxins on the Ex Vivo Physical Barrier
An increasing number of ex vivo studies have determined the effects of mycotoxins on the intestinal physical barrier. Recently, DON was reported to inhibit growth and reduce ZO-1 and claudin-1 expression levels in enteroids isolated from jejunal crypts in porcine and mice [109,110,111]. When jejunal explants from weaning piglets were exposed to DON, decreased jejunal scores and MAPK activation were recorded [72]. FB exposure to an ex vivo rat large intestine induced lipid peroxidation, which could alter cell membrane permeability [129]. Apical villi necrosis was found in pig jejunal explant exposure to PAT [123]. DON appears to be the most studied mycotoxin in ex vivo studies. Therefore, it could be speculated that an ex vivo model is also highly applicable for other mycotoxins.
4.1.3. Effects of Mycotoxins on the In Vivo Physical Barrier
AFB1 and AFM1 have the ability to evaluate caspase-3 and caspase-9 mRNA expression and reduce the Bcl-2/Bax ratio in mice intestinal tissues, suggesting these mycotoxins induce apoptotic events [100]. When broiler chicks were exposed to AFB1, researchers observed increased lactulose:rhamnose (L:R) ratio and diamine oxidase (DAO) in plasma, and reduced clauin-1 and occludin mRNA expression levels in the mid-jejunum [101,102]. DON-mediated impairment of barrier function in rats and grass carp was associated with the depressed crypt depth ratio and TJ amount in the jejunum, which was associated with Wnt/β-catenin and MLCK signaling pathways [108,110,111,113]. In addition, DON-treated weaned piglets exhibited lower disaccharidase (maltase, sucrase, and lactase) activity, suggesting detrimental effects on gut health [112]. Similarly, the effects caused by ZEN in the post-weaning gilts intestinal physical barrier were partially elucidated by ZEN-induced oxidative stress mechanisms [120]. Similar results were shown in rats challenged with ZEN; TJ proteins in the jejunum exhibited down-regulated mRNA expression [119]. The chronic ingestion of FB1 decreased villi height and occludin expression in the piglet intestine [121]. In addition, T-2 toxin treatment disrupted intestinal histology in turkeys and mice, including the duodenum, jejunum, ileum, and colon [124,125].
The adverse effects on intestinal morphology in various in vivo models have been recorded for different mycotoxins such as AFs [99,103,104], OTA [107], and DON [72,114,115,116,117]. In addition, a variety of clinical chemistry analytes such as urine, feces, and blood may also reflect defects in intestinal barrier function in vivo (e.g., fatty acid-binding protein and C-reactive protein levels in serum, and fecal hemoglobin and fecal calprotectin) [130,131]. However, these parameters have not been widely used to evaluate the effect of mycotoxin effects. Therefore, they may provide new insights in future studies.
These observations show that when the intestine is affected by short-term, low-concentration mycotoxins, self-regulating cellular abilities maintain the intestinal physical barrier integrity. However, when such damage is sustained and exceeds the self-regulating capabilities, intestinal epithelial cells and TJ proteins become disrupted, leading to barrier compromise. Similarly, long-term exposure of animals to low mycotoxin doses could also adversely affect intestine health.
4.2. Effects of Mycotoxins on the Chemical Barrier
The contribution of mycotoxins to intestinal chemical barrier disruption (e.g., mucins and AMPs) has been extensively conducted across in vitro, ex vivo, and in vivo studies (Table 5). To date, hundreds of AMPs have been identified including LEAP-2A, LEAP-2B, hepcidin, and β-defensin1.
4.2.1. Effects of Mycotoxins on the In Vitro Chemical Barrier
In the co-culture of Caco-2/HT29-MTX cells, AFM1, OTA, and ZEN down-regulated the protein expression of intestinal mucins MUC2, MUC5AC, and MUC5B [43,57,135,136]. Similar results were also observed for DON where it inhibited MUC1, MUC2, and MUC3 mRNA levels in human goblet cells (HT29-16E cells) [134]. T-2 toxin exposure led to a thinned MUC2 layer in HT-29 cells and reduced MUC2 protein expression in Caco-2 cells [124]. Up-regulated porcine β-defensin 1 and β-defensin 2 mRNA expression was also observed following exposure to DON, ZEN, FB1, and NIV, whereas no significant increases in β-defensin 1 and β-defensin 2 secretion occurred in IPEC-J2 cells [137]. In addition, in co-culture studies with Caco-2/HT29-MTX cells, MUC5AC and MUC5B mRNA expression was decreased by DON and NIV exposure, while their protein levels were increased [136]. These differences in mRNA and protein levels may be partly explained by post-transcriptional or post-translational regulatory mechanisms, or protein degradation pathways. Equally, discrepancies in quantification techniques between studies may have also led to these differences, as protein quantification sensitivity is not as high as transcript measurements [144].
4.2.2. Effects of Mycotoxins on Ex Vivo and In Vivo Chemical Barriers
Few studies have investigated mycotoxin effects on the intestinal chemical barrier. One study showed that, in jejunal explants from pigs, DON time-dependently inhibited mucin mRNA expression levels [134]. In vivo exposure to a mycotoxin-contaminated diet also regulated mucin and other protein productions. Both AMPs and MUC2 mRNA expression levels were inhibited in the intestine of broiler chickens, mice, and juvenile grass carp upon DON exposure [110,111,138,139,140]. Furthermore, reduced goblet cells in the small intestine were observed in OTA and DON treatment [107,112,115,121,140,141]. In contrast, no changes in goblet cells and crude mucin production were observed in laying hens and gilts exposed to AFB1, DON, or ZEN [133,142]. Furthermore, goblet cell hyperplasia (increased goblet cell number) was observed in AF/FB1-challenged broilers and mice exposed to combined DON and ZEN [104,121,132,145]. In general, goblet cell hyperplasia led to increased mucins secretion. T-2 toxin exposure resulted in increased MUC2 mRNA levels in the jejunum of chickens [143]. These observations suggest that the continuous hyper-secretion of mucins is likely to deplete goblet cell number, ultimately disrupting the mucus layer [146].
A variety of responses (increase, no change, and/or decrease) were observed in mycotoxin-induced mucins and goblet cells. Different experimental models using different species could account for these inconsistencies. Precise mechanisms remain unclear, however, and more studies are required to resolve these issues.
4.3. Effects of Mycotoxins on the Immunological Barrier
Mycotoxin effects on the intestinal immunological barrier in in vitro and in vivo models are summarized (Table 6).
4.3.1. Effects of Mycotoxins on the In Vitro Immunological Barrier
OTA down-regulated the gene expression of cyclooxygenase-2 (COX-2) and lipoxygenase-5 (5-LOX) genes in Caco-2 cells, which have been regarded as inflammatory mediators [105]. The mRNA expressions of pro-inflammatory cytokines including interleukin-1 beta (IL-1β), IL-6, IL-8, COX-2, and tumor necrosis factor-alpha (TNF-α) were increased in the DON-exposed porcine intestinal epithelial cells, with DON-mediated inflammation partially dependent on ROS production and MAPK pathway activation [106,151,152,153,154]. Another study also reported that ZEN exerted no effects, or only marginal ones, on cytokine gene expression in IPEC-1 cells [156]. However, other studies reported an increasing tendency towards inflammatory cytokine gene expression and synthesis in ZEN-exposed IPEC-1 cells [91,118,157]. In addition, the ZEN metabolites α-ZOL and β-ZOL decreased pro- and anti-inflammatory cytokine expressions [118]. The exposure of FB1, in a concentration-dependent manner, to IPEC-1 cells depressed IL-8 mRNA and protein expression, while no changes in IL-8 secretion were recorded in HT-29 cells [93,158]. These studies show that mycotoxins, especially ZEN, exert different inflammatory cytokine expression profiles, suggesting underlying mechanisms are complex and require further in-depth study. Additionally, there is no evidence on the impact of AFs on intestinal inflammation. Therefore, more studies are required to fill this important knowledge gap.
4.3.2. Effects of Mycotoxins on the In Vivo Immunological Barrier
AFB1 and AFM1 down-regulated pro-inflammatory cytokine production in the small intestine of mice and pigs [100,147]. AFB1 also affected sIgA and polymeric immunoglobulin receptor (pIgR) expression, thereby affecting sIgA transport in epithelial cells into mucus layers. Transcriptome analysis revealed that seven processes or pathways related to the immune system were grouped in Litopenaeus vannamei as the result of AFB1 challenge [148]. OTA exposure resulted in up-regulated IL-1β and TNF-α mRNA expression in the small intestine of broiler chickens and pekin ducklings [107,150]. Similar results were also observed for DON and FB1 exposure, where cytokine production was significantly up-regulated in the intestine of piglets and juvenile grass carp [121,138,155]. In other studies, in the intestines of pregnant dams, weaned pups, and pigs treated with ZEN and FB1, pro-inflammatory cytokine mRNA expression was significantly decreased [119,158].
CD4+ cells are involved in cytokine production, and CD8+ cells play a crucial role in protecting host target cells [160,161]. Mycotoxin-contaminated diets led to decreased CD4+ and CD8+ cell percentages in chickens [162]. In addition, CD4+ cell numbers were decreased in the jejunum of broilers treated with AFB1 [149]. In addition, CTN administration altered immune cell (CD4+, CD8+, and CD19+) populations in the small intestine of mice [159]. Similarly, cell morphology effects have been observed in these models; in AF-challenged broiler chicks, the lamina propria lymphoid follicle diameter (LLFD) and lamina propria lymphoid follicle number (LLFN) in the jejunum were increased [104]. In addition, DON exposure increased the lymphocyte number in the intestine of pigs fed DON-contaminated diets [141,142], indicating mycotoxin-mediated inflammatory responses.
These data reflect the limited research on disrupted intestinal chemical barriers induced by mycotoxins in ex vivo models. While these studies have enlightened the literature on AFs, OTA, DON, and ZEN, the effects of other mycotoxins such as FB1 and PAT require greater attention.
4.4. Effects of Mycotoxins on the Microbial Barrier
Several studies have shown the impairment to the intestinal microbial barrier is caused by mycotoxins (Table 7).
4.4.1. Effects of Mycotoxins on the In Vitro Microbial Barrier
The simulator of the human intestinal microbial ecosystem (SHIME) was used to investigate OTA effects, and it showed that gut microbiota diversity was altered, with the loss of beneficial species Lactobacillus reuteri [167]. In another study, Escherichia coli (E. coli) were translocated across IPEC-J2 cell monolayers when induced by DON [152]. This finding was consistent with previous studies, demonstrating that bacterial translocations across IPEC-1 and Caco-2 cell monolayers are induced by DON, NIV, and PAT [166,169]. Importantly, bacterial translocation appears to be concentration- and species-dependent [179], with incubation times and cell line types potentially affecting the results [166].
4.4.2. Effects of Mycotoxins on the In Vivo Microbial Barrier
In broiler chicks, the dietary supplementation of AFs resulted in markedly increased ileal bacterial counts of E. coli, Klebsiella, Salmonella, and total negative bacteria [104,132,165]. In addition, AFB1 reduced intestinal bacterial flora diversity in rats, shrimp, and mice [163,164,180]. In AFB1-treated shrimp, the application of high-throughput sequencing analysis showed that Bacteroidetes relative abundance decreased, and the abundance of Proteobacteria and Firmicutes increased [180]. These results were consistent with a previous study demonstrating that OTA exposure significantly increased the relative abundance of Lactobacillus, but decreased overall gut microbiota diversity in rats [168].
Cecal species’ richness and evenness were decreased upon DON exposure in broiler chickens and mice, Bacteroidetes and Firmicutes abundance increased, while Proteobacteria decreased, suggesting an overall impaired gut microbiota community [170,171]. A fecal microbiota analysis explored DON-induced gut microbiota changes in nude mice and pigs and showed that DON treatment generated a higher abundance of Clostridiales, Lachnospiraceae, and fecal aerobic mesophilic bacteria [140,172]. In addition, DON exposure increased Bacteroides and Prevotella genera levels but decreased E. coli levels in a model of human microbiota-associated rats [174].
The biolog EcoPlate method showed that ZEN exposure significantly reduced E. coli, Clostridium perfringens, and Enterobacteriaceae levels in gilts [177]. Additionally, ZEN increased Desulfovibrio and decreased Lactobacillus in mouse colon tissue [176]. After exposure to FBs, the digestive microbiota balance was impaired in pigs [178]. In contrast, AFB1 and DON exerted no effects on gut microbiota diversity and relative abundance in broilers and rats [102,173,175].
In general, microbiota communities in the colon, cecum, and feces are representative of the intestinal bacterial flora. However, microbiota differences in different intestinal sections are still unclear. Similarity, studies on mycotoxin (i.e., ZEN and FB1) mediated alterations in the intestinal microbial barrier are limited. Therefore, more research is required in this area.
5. Contribution of a Leaky Gut to Intestinal Inflammatory Disease
Currently, links between mycotoxin exposure and some human carcinogenic and teratogenic diseases, including Reye’ syndrome, cirrhosis, hepatitis, and esophagus cancer, have been demonstrated [181]. However, potential links between mycotoxins and human chronic intestinal inflammatory diseases remain unclear. Among the chronic intestinal inflammatory diseases, inflammatory bowel diseases (IBDs) including ulcerative colitis (UC), Crohn’s disease (CD), and celiac disease (CeD) are well studied. The dysfunction of intestinal barrier and increased intestinal permeability, also known as “leaky gut”, are believed to play a prominent role in the etiology of these diseases [182,183].
Intestinal epithelial cell death and abnormal TJ expression cause increased transcellular and paracellular transport. These perturbations may cause increased access of different molecular weight xenobiotics and bacterial translocation, ultimately activating local and systemic immune responses [23,184]. Among the abnormalities of TJs, claudin switching is the most notable, which refers to the balance between different members of the claudin family [185,186]. In particular, the down-regulation of several tight claudins, such as claudins 3, 4, 5, 7, and 8, and the increased expression of leaky claudins, such as claudin 2 and 15, are often associated with an inflamed gut, including UC, CD, CeD, and irritable bowel syndrome [185,187,188,189,190]. In addition, reduced TJ expression could accelerate mucosal inflammation (ulceration and colitis), as observed in claudin 2 and claudin 7 knockout mice, and claudin 2 overexpression investigations in mice [191,192,193].
In turn, inflammatory mediators themselves could also generate negative effects on the TJs. Thus, inflammatory responses caused by the initial increases in intestinal permeability could be responsible for a stronger disruptive effect in the intestinal physical barrier [194]. Indeed, several cytokines (e.g., IL-1β, IL-6, IL-9, IL-17, IL-23, IFN-γ, and TNF-α) have been shown to alter TJ expression in in vitro and in vivo models [195,196,197,198,199,200,201,202,203].
In addition to intestinal physical and immunological barriers, chemical barrier disruption also leads to inflammatory disease. Perturbations in intestinal mucus layers, including mucus composition and production, contribute to human chronic intestinal inflammatory disease. In addition, significant modifications to gut microbiota number and composition were observed in IBD patients and were characterized by decreased anaerobic bacteria and increased Enterobacteria numbers [181]. The intestinal microflora may also affect mucus production and TJ expression, potentially leading to inflammatory responses [204,205]. As such, the intestinal barrier is an interconnected system. Therefore, disruption to one or more components may result in human intestinal disease [20,206]. Several factors are involved in the induction and persistence of the chronic intestinal inflammatory diseases: (i) abnormal TJs composition, (ii) altered production of mucus layers, (iii) changes in intestinal immunity, and (iv) modification of intestinal microflora or increased bacterial translocation (Figure 2). Given that healthy relatives of CD patients manifested increased intestinal permeability without disease [207,208], we conclude that a compromised intestinal barrier is not the sole factor causing inflammatory diseases, but intestinal dysfunction could exacerbate inflammation and enhance its severity [21,183].
6. Interactive Effects of Mycotoxins on Intestinal Barrier
Most fungi produce one or more mycotoxins. Therefore, the co-occurrence of these molecules in disease should receive more attention. A previous study demonstrated that approximately 28 mycotoxins were quantified by a liquid chromatography tandem mass spectrometry (LC-MS/MS) multi-toxin method, in samples collected from Burkina Faso and Mozambique [209]. A recent study also showed a greater co-exposure phenomenon, where the analyzed maize samples were contaminated by 5 to 41 mycotoxins in Malawi [210]. In addition, mycotoxin co-occurrence was also identified in oil, dairy milk, and infant formulas [211,212,213]. After ingestion of mycotoxin-contaminated food, these molecules may be absorbed into the blood. To corroborate this, UHPLC-MS/MS analysis identified 26 mycotoxin biomarkers, including AFB1, OTA, FB1, ZEN, and DON, in plasma and urine samples from 260 rural residents in China [214]. Equally, similar data have been observed in different populations across the globe [215,216,217,218].
The interactive effects of multi-mycotoxins are classified as synergistic, additive, and antagonistic, and they represent the effects of a components’ mixture are higher, equal, and lower than the sum of the effects induced by individual components. For the intestinal physical barrier, AFM1 cytotoxicity against Caco-2 cells was enhanced in the presence of OTA, ZEA, and α-ZOL, with additive and synergistic effects demonstrated by most combinations [219]. The additive and synergistic effects on reduced cell viability were demonstrated by combining DON and ZEN in a bi-culture Caco-2, THP-1, and HepaRG cell system [220]. The effects on increased intestinal permeability and decreased TJ expression in differentiated Caco-2 cells exposed to a combination of AFM1 and OTA were significantly stronger than the individual mycotoxins, performing additive and synergistic effect [54]. In addition, non-toxic OTA aggravated DON-induced TJ disruption in IPEC-J2 cells, suggesting a synergistic effect was at play [106].
From a chemical barrier perspective, an AFM1 and OTA combinatorial approach produced a synergistic effect by depressing mucin expression in Caco-2/HT29-MTX co-cultures [43]. In contrast, antagonistic interaction of combined DON and ZEN was observed for MUC5AC expression levels [135]. Non-additive (including synergistic and antagonistic) effects were also shown for the modulation of mucins and β-defensins expression, induced by combinations of DON, FB1, ZEN, and NIV [136,137]. Furthermore, different interactions were observed in the combination of DON and FB on intestinal immunological changes [121]. Additionally, the antagonist effect was observed in the combined DON and ZEN on gut microbiota levels [177].
To protect the gut from mycotoxin assault, several approaches have been undertaken to assess the protective effects of different materials on intestinal dysfunction. Lactoferrin [79], grape seed [147], silymarin [132], yeast cell wall [102], Lactobacillus plantarum [100], and mannanoligosaccharides [104] have alleviated AF-induced intestinal disturbances in human intestinal epithelial cells, pigs, broiler chicks, and mice. Probiotic Bacillus subtilis [82], curcumin [150], and selenium-rich yeast [107] have attenuated OTA-induced disruption of intestinal barrier in Caco-2 cells, ducks, and broiler chickens. Similarly, mixtures of Lactobacilli [156] and Bacillus cereus BC7 strain [180] efficiently renormalized ZEN-induced perturbations of intestinal inflammatory responses and microbiota. Resveratrol [152], probiotic Lactobacillus rhamnosus GG [140], methionine and its hydroxyl analogues [110], and hydrolyzed wheat gluten [111] all reduced DON-induced intestinal damage. Eugenol also protected against CTN-induced intestinal cytotoxicity in HCT116 cells [9]. More importantly, L. rhamnosus GG was reported to improve intestinal barrier functions in mice exposed to DON and ZEN combination [145].
7. Conclusions
Mycotoxin contamination of human food and animal feed results in economic loss and health detriments. In addition to the carcinogenic, mutagenic, and genotoxic effects mediated by these molecules, mycotoxin-induced impairment of the intestinal barrier has also gained considerable attention in recent years. To assess intestinal toxicity, in vitro epithelial barrier investigations frequently use human (Caco-2 and HT29 cells) and porcine epithelial cells (IPEC-1and IPEC-J2) as well as enteroids. Ex vivo models are represented by explants from mice and pigs. The predominant in vivo models use rodents (mice and rats) and pigs, followed by chickens and fish. Based on these varied models, our review revealed that the mycotoxins, i.e., AFs, OTA, ZEN, DON and FB1, decreased TJ protein expression levels, depressed mucins and AMPs secretion, activated cytokine production, and altered gut microflora composition. These elements represent the intestinal physical, chemical, immunological, and microbial barrier targeted by mycotoxins.
It is entirely plausible that mycotoxins are implicated in human chronic intestinal inflammatory disease. In this review, we showed evidence linking a compromised intestinal barrier to inflammatory disease, but further studies are required to fully confirm this hypothesis. Therefore, underlying mechanisms remain an issue. More studies are required to investigate how to control and decrease mycotoxin-contaminated foods and animal feeds, to protect human and animal health, and to reduce economic loss.
Author Contributions
Y.G. and J.W. conceived and designed the experiments; Y.G. performed the experiments; L.M. and H.L. analyzed the data; Y.G. contributed reagents/materials/analysis tools; Y.G. and N.Z. wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by National Natural Science Foundation of China (31972190), the Independent Research Project of State Key Laboratory of Animal Nutrition (2004DA125184G1608), the Scientific Research Project for Major Achievements of Agricultural Science and Technology Innovation Program (CAAS-ZDXT2019004), and Modern Agro-Industry Technology Research System of the PR China (CARS-36).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AFs | Aflatoxins |
| AMPs | Antimicrobial peptides |
| CCK-8 | Cell counting kit-8 |
| CD | Crohn’s disease |
| CeD | Celiac disease |
| COX-2 | Cyclooxygenase-2 |
| CTN | Citrinin |
| CXCL10 | C-X-C Motif Chemokine Ligand 10 |
| DAO | Diamine oxidase |
| DON | Deoxynivalenol |
| ELISA | Enzyme-linked immunosorbent assay |
| FBs | Fumonisins |
| IBD | Inflammatory bowel diseases |
| IFN-γ | Interferon-γ |
| IL-1β | Interleukin-1 beta |
| JAMs | Junctional adhesion molecules |
| LP | Lamina propria |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MUC2 | Mucin 2 |
| NIV | Nivalenol |
| OTA | Ochratoxin A |
| PAT | Patulin |
| PMTDI | Provisional maximum tolerable daily intake |
| PTWI | Provisional tolerable weekly intake |
| RT-qPCR | Real-time quantitative PCR |
| sIgA | Secretory immunoglobulin A |
| TDI | Tolerable daily intake |
| TEER | Transepithelial electrical resistance |
| TJs | Tight junctions |
| TNF-α | Tumor necrosis factor-alpha |
| TWI | Tolerable weekly intake |
| UC | Ulcerative colitis |
| ZEN | Zearalenone |
| ZO | Zonula occludens |
References
- Cimbalo, A.; Alonso-Garrido, M.; Font, G.; Manyes, L. Toxicity of mycotoxins in vivo on vertebrate organisms: A review. Food Chem. Toxicol. 2020, 137, 111161. [Google Scholar] [CrossRef]
- Marin, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Mycotoxins: Occurrence, toxicology, and exposure assessment. Food Chem. Toxicol. 2013, 60, 218–237. [Google Scholar] [CrossRef] [PubMed]
- Broom, L. Mycotoxins and the intestine. Anim. Nutr. 2015, 1, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Adegbeye, M.J.; Reddy, P.R.K.; Chilaka, C.A.; Balogun, O.B.; Elghandour, M.; Rivas-Caceres, R.R.; Salem, A.Z.M. Mycotoxin toxicity and residue in animal products: Prevalence, consumer exposure and reduction strategies-A review. Toxicon 2020, 177, 96–108. [Google Scholar] [CrossRef]
- Yang, Y.; Li, G.; Wu, D.; Liu, J.; Li, X.; Luo, P.; Hu, N.; Wang, H.; Wu, Y. Recent advances on toxicity and determination methods of mycotoxins in foodstuffs. Trends Food Sci. Technol. 2020, 96, 233–252. [Google Scholar] [CrossRef]
- Ostry, V.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens-the IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Oswald, I.P. Effect of deoxynivalenol and other Type B trichothecenes on the intestine: A review. Toxins 2014, 6, 1615–1643. [Google Scholar] [CrossRef] [PubMed]
- Chain, E.P.o.C.i.t.F.; Knutsen, H.K.; Alexander, J.; Barregard, L.; Bignami, M.; Bruschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; et al. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar] [CrossRef]
- Saleh, I.; Goktepe, I. The characteristics, occurrence, and toxicological effects of patulin. Food Chem. Toxicol. 2019, 129, 301–311. [Google Scholar] [CrossRef]
- Pestka, J. Toxicological mechanisms and potential health effects of deoxynivalenol and nivalenol. World Mycotoxin J. 2010, 3, 323–347. [Google Scholar] [CrossRef]
- Dong, H.; Xian, Y.; Xiao, K.; Wu, Y.; Zhu, L.; He, J. Development and comparison of single-step solid phase extraction and QuEChERS clean-up for the analysis of 7 mycotoxins in fruits and vegetables during storage by UHPLC-MS/MS. Food Chem. 2019, 274, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Heshmati, A.; Zohrevand, T.; Khaneghah, A.M.; Mozaffari Nejad, A.S.; Sant’Ana, A.S. Co-occurrence of aflatoxins and ochratoxin A in dried fruits in Iran: Dietary exposure risk assessment. Food Chem. Toxicol. 2017, 106, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Pizzolato Montanha, F.; Anater, A.; Burchard, J.F.; Luciano, F.B.; Meca, G.; Manyes, L.; Pimpao, C.T. Mycotoxins in dry-cured meats: A review. Food Chem. Toxicol. 2018, 111, 494–502. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.K.; Gong, Z.Z.; Zhang, T.; Cai, W. Sodium butyrate attenuates soybean oil-based lipid emulsion-induced increase in intestinal permeability of lipopolysaccharide by modulation of P-glycoprotein in Caco-2 cells. Biochem. Biophys. Res. Commun. 2017, 482, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Sengling Cebin Coppa, C.F.; Mousavi Khaneghah, A.; Alvito, P.; Assunção, R.; Martins, C.; Eş, I.; Gonçalves, B.L.; Valganon de Neeff, D.; Sant’Ana, A.S.; Corassin, C.H.; et al. The occurrence of mycotoxins in breast milk, fruit products and cereal-based infant formula: A review. Trends Food Sci. Technol. 2019, 92, 81–93. [Google Scholar] [CrossRef]
- Hueza, I.M.; Raspantini, P.C.; Raspantini, L.E.; Latorre, A.O.; Gorniak, S.L. Zearalenone, an estrogenic mycotoxin, is an immunotoxic compound. Toxins 2014, 6, 1080–1095. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Habrowska-Gorczynska, D.E.; Piastowska-Ciesielska, A.W. Zearalenone as an endocrine disruptor in humans. Environ. Toxicol. Pharmacol. 2016, 48, 141–149. [Google Scholar] [CrossRef]
- Bouhet, S.; Oswald, I.P. The effects of mycotoxins, fungal food contaminants, on the intestinal epithelial cell-derived innate immune response. Vet. Immunol. Immunopathol. 2005, 108, 199–209. [Google Scholar] [CrossRef]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [Green Version]
- Groh, K.J.; Geueke, B.; Muncke, J. Food contact materials and gut health: Implications for toxicity assessment and relevance of high molecular weight migrants. Food Chem. Toxicol. 2017, 109, 1–18. [Google Scholar] [CrossRef]
- Natividad, J.M.; Verdu, E.F. Modulation of intestinal barrier by intestinal microbiota: Pathological and therapeutic implications. Pharmacol. Res. 2013, 69, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.E.; Araujo, R.S.; de Barros, P.A.; Soares, A.D.; Abrantes, F.A.; Generoso Sde, V.; Fernandes, S.O.; Cardoso, V.N. The role of immunomodulators on intestinal barrier homeostasis in experimental models. Clin. Nutr. 2015, 34, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, A.; Shi, Z.; He, C.; Ding, J.; Wang, X.; Ma, J.; Zhang, H. A mitochondria-mediated apoptotic pathway induced by deoxynivalenol in human colon cancer cells. Toxicol. Vitro 2012, 26, 414–420. [Google Scholar] [CrossRef]
- Drolia, R.; Bhunia, A.K. Crossing the Intestinal Barrier via Listeria Adhesion Protein and Internalin A. Trends Microbiol. 2019, 27, 408–425. [Google Scholar] [CrossRef]
- Furuse, M.; Hirase, T.; Itoh, M.; Nagafuchi, A.; Yonemura, S.; Tsukita, S. Occludin: A novel integral membrane protein localizing at tight junctions. J. Cell Biol. 1993, 123, 1777–1788. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, L.; Betanzos, A.; Avila-Flores, A. MAGUK proteins: Structure and role in the tight junction. Semin. Cell Dev. Biol. 2000, 11, 315–324. [Google Scholar] [CrossRef]
- Hooper, L.V. Do symbiotic bacteria subvert host immunity? Nat. Rev. 2009, 7, 367–374. [Google Scholar] [CrossRef]
- McGuckin, M.A.; Linden, S.K.; Sutton, P.; Florin, T.H. Mucin dynamics and enteric pathogens. Nat. Rev. Microbiol. 2011, 9, 265–278. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.E. Mucus layers in inflammatory bowel disease. Inflamm. Bowel. Dis. 2014, 20, 2124–2131. [Google Scholar] [CrossRef] [PubMed]
- Corthesy, B. Role of secretory IgA in infection and maintenance of homeostasis. Autoimmun. Rev. 2013, 12, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A.J.; Geuking, M.B.; McCoy, K.D. Immune responses that adapt the intestinal mucosa to commensal intestinal bacteria. Immunology 2005, 115, 153–162. [Google Scholar] [CrossRef]
- Keeney, K.M.; Finlay, B.B. Enteric pathogen exploitation of the microbiota-generated nutrient environment of the gut. Curr. Opin. Microbiol. 2011, 14, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Microbiol. 2018, 16, 457–470. [Google Scholar] [CrossRef]
- Yang, Y.; Jobin, C. Microbial imbalance and intestinal pathologies: Connections and contributions. Dis. Model Mech. 2014, 7, 1131–1142. [Google Scholar] [CrossRef] [Green Version]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef] [Green Version]
- Bertero, A.; Augustyniak, J.; Buzanska, L.; Caloni, F. Species-specific models in toxicology: In vitro epithelial barriers. Environ. Toxicol. Pharmacol. 2019, 70, 103203. [Google Scholar] [CrossRef]
- Clevers, H. Modeling Development and Disease with Organoids. Cell 2016, 165, 1586–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, M.J.; Han, S.E.; Hwang, K.; Mayer, E.; Reisinger, N.; Schatzmayr, D.; Park, B.C.; Han, S.H.; Yun, C.H. Hydrolyzed fumonisin B1 induces less inflammatory responses than fumonisin B1 in the co-culture model of porcine intestinal epithelial and immune cells. Toxicol. Lett. 2019, 305, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, Y.; Li, S.; Wu, C.; Wang, J.; Zheng, N. Modulation of Mucin (MUC2, MUC5AC and MUC5B) mRNA Expression and Protein Production and Secretion in Caco-2/HT29-MTX Co-Cultures Following Exposure to Individual and Combined Aflatoxin M1 and Ochratoxin A. Toxins 2019, 11, 132. [Google Scholar] [CrossRef] [Green Version]
- Nossol, C.; Barta-Boszormenyi, A.; Kahlert, S.; Zuschratter, W.; Faber-Zuschratter, H.; Reinhardt, N.; Ponsuksili, S.; Wimmers, K.; Diesing, A.K.; Rothkotter, H.J. Comparing Two Intestinal Porcine Epithelial Cell Lines (IPECs): Morphological Differentiation, Function and Metabolism. PLoS ONE 2015, 10, e0132323. [Google Scholar] [CrossRef] [Green Version]
- Olayanju, A.; Jones, L.; Greco, K.; Goldring, C.E.; Ansari, T. Application of porcine gastrointestinal organoid units as a potential in vitro tool for drug discovery and development. J. Appl. Toxicol. 2019, 39, 4–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Clevers, H. Growing self-organizing mini-guts from a single intestinal stem cell: Mechanism and applications. Science 2013, 340, 1190–1194. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.B.; Guo, S.G.; Wan, D.; Wu, X.; Yin, Y.L. Enteroids: Promising in Vitro Models for Studies of Intestinal Physiology and Nutrition in Farm Animals. J. Agric. Food Chem. 2019, 67, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Zhou, D. Organoid and Enteroid Modeling of Salmonella Infection. Front. Cell Infect. Microbiol. 2018, 8, 102. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.A.; Nighot, P.; Reyes, C.; Rawat, M.; McKee, J.; Lemon, D.; Hanson, J.; Ma, T.Y. Intestinal barrier dysfunction in human necrotizing enterocolitis. J. Pediatr. Surg. 2016, 51, 1907–1913. [Google Scholar] [CrossRef] [Green Version]
- Neirinckx, E.; Vervaet, C.; Michiels, J.; De Smet, S.; Van den Broeck, W.; Remon, J.P.; De Backer, P.; Croubels, S. Feasibility of the Ussing chamber technique for the determination of in vitro jejunal permeability of passively absorbed compounds in different animal species. J. Vet. Pharmacol. Ther. 2011, 34, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Bertero, A.; Spicer, L.J.; Caloni, F. Fusarium mycotoxins and in vitro species-specific approach with porcine intestinal and brain in vitro barriers: A review. Food Chem. Toxicol. 2018, 121, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, S.; Bao, X.; Luo, C.; Yang, H.; Wang, J.; Zhao, S.; Zheng, N. Transcriptional and Proteomic Analysis Revealed a Synergistic Effect of Aflatoxin M1 and Ochratoxin A Mycotoxins on the Intestinal Epithelial Integrity of Differentiated Human Caco-2 Cells. J. Proteome Res. 2018, 17, 3128–3142. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, S.; Wang, J.; Luo, C.; Zhao, S.; Zheng, N. Modulation of Intestinal Epithelial Permeability in Differentiated Caco-2 Cells Exposed to Aflatoxin M1 and Ochratoxin A Individually or Collectively. Toxins 2017, 10, 13. [Google Scholar] [CrossRef] [Green Version]
- Trapecar, M.; Leouffre, T.; Faure, M.; Jensen, H.E.; Granum, P.E.; Cencic, A.; Hardy, S.P. The use of a porcine intestinal cell model system for evaluating the food safety risk of Bacillus cereus probiotics and the implications for assessing enterotoxigenicity. APMIS 2011, 119, 877–884. [Google Scholar] [CrossRef]
- Adler, S.; Basketter, D.; Creton, S.; Pelkonen, O.; van Benthem, J.; Zuang, V.; Andersen, K.E.; Angers-Loustau, A.; Aptula, A.; Bal-Price, A.; et al. Alternative (non-animal) methods for cosmetics testing: Current status and future prospects-2010. Arch. Toxicol. 2011, 85, 367–485. [Google Scholar] [CrossRef]
- Wu, C.; Gao, Y.; Li, S.; Huang, X.; Bao, X.; Wang, J.; Zheng, N. Modulation of intestinal epithelial permeability and mucin mRNA (MUC2, MUC5AC, and MUC5B) expression and protein secretion in Caco-2/HT29-MTX co-cultures exposed to aflatoxin M1, ochratoxin A, and zearalenone individually or collectively. Toxicol. Lett. 2019, 309, 1–9. [Google Scholar] [CrossRef]
- Powell, R.H.; Behnke, M.S. WRN conditioned media is sufficient for in vitro propagation of intestinal organoids from large farm and small companion animals. Biol. Open 2017, 6, 698–705. [Google Scholar] [CrossRef] [Green Version]
- Merenda, A.; Fenderico, N.; Maurice, M.M. Wnt Signaling in 3D: Recent Advances in the Applications of Intestinal Organoids. Trends Cell Biol. 2020, 30, 60–73. [Google Scholar] [CrossRef] [Green Version]
- van der Hee, B.; Loonen, L.M.P.; Taverne, N.; Taverne-Thiele, J.J.; Smidt, H.; Wells, J.M. Optimized procedures for generating an enhanced, near physiological 2D culture system from porcine intestinal organoids. Stem Cell Res. 2018, 28, 165–171. [Google Scholar] [CrossRef]
- Roeselers, G.; Ponomarenko, M.; Lukovac, S.; Wortelboer, H.M. Ex vivo systems to study host-microbiota interactions in the gastrointestinal tract. Best Pract. Res. Clin. Gastroenterol. 2013, 27, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Nighot, P.K.; Blikslager, A.T. ClC-2 regulates mucosal barrier function associated with structural changes to the villus and epithelial tight junction. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G449–G456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.L.; Little, D.; Blikslager, A.T.; Papich, M.G. Mucosal permeability of water-soluble drugs in the equine jejunum: A preliminary investigation. J. Vet. Pharmacol. Ther. 2006, 29, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Randall, K.J.; Turton, J.; Foster, J.R. Explant culture of gastrointestinal tissue: A review of methods and applications. Cell Biol. Toxicol. 2011, 27, 267–284. [Google Scholar] [CrossRef]
- Tortora, R.; Russo, I.; De Palma, G.D.; Luciani, A.; Rispo, A.; Zingone, F.; Iovino, P.; Capone, P.; Ciacci, C. In vitro gliadin challenge: Diagnostic accuracy and utility for the difficult diagnosis of celiac disease. Am. J. Gastroenterol. 2012, 107, 111–117. [Google Scholar] [CrossRef]
- Chow, J.; Lee, S.M.; Shen, Y.; Khosravi, A.; Mazmanian, S.K. Host-bacterial symbiosis in health and disease. Adv. Immunol. 2010, 107, 243–274. [Google Scholar] [CrossRef] [Green Version]
- Jarry, A.; Cremet, L.; Caroff, N.; Bou-Hanna, C.; Mussini, J.M.; Reynaud, A.; Servin, A.L.; Mosnier, J.F.; Lievin-Le Moal, V.; Laboisse, C.L. Subversion of human intestinal mucosa innate immunity by a Crohn’s disease-associated E. coli. Mucosal. Immunol. 2015, 8, 572–581. [Google Scholar] [CrossRef]
- Garcia, G.R.; Dogi, C.A.; Poloni, V.L.; Fochesato, A.S.; De Moreno de Leblanc, A.; Cossalter, A.M.; Payros, D.; Oswald, I.P.; Cavaglieri, L.R. Beneficial effects of Saccharomyces cerevisiae RC016 in weaned piglets: In vivo and ex vivo analysis. Benef. Microbes 2019, 10, 33–42. [Google Scholar] [CrossRef]
- Gerez, J.; Buck, L.; Marutani, V.H.; Calliari, C.M.; Bracarense, A.P. Low Levels of Chito-Oligosaccharides Are Not Effective in Reducing Deoxynivalenol Toxicity in Swine Jejunal Explants. Toxins 2018, 10, 276. [Google Scholar] [CrossRef] [Green Version]
- Kolf-Clauw, M.; Castellote, J.; Joly, B.; Bourges-Abella, N.; Raymond-Letron, I.; Pinton, P.; Oswald, I.P. Development of a pig jejunal explant culture for studying the gastrointestinal toxicity of the mycotoxin deoxynivalenol: Histopathological analysis. Toxicol. Vitro 2009, 23, 1580–1584. [Google Scholar] [CrossRef]
- Kolf-Clauw, M.; Sassahara, M.; Lucioli, J.; Rubira-Gerez, J.; Alassane-Kpembi, I.; Lyazhri, F.; Borin, C.; Oswald, I.P. The emerging mycotoxin, enniatin B1, down-modulates the gastrointestinal toxicity of T-2 toxin in vitro on intestinal epithelial cells and ex vivo on intestinal explants. Arch. Toxicol. 2013, 87, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Lucioli, J.; Pinton, P.; Callu, P.; Laffitte, J.; Grosjean, F.; Kolf-Clauw, M.; Oswald, I.P.; Bracarense, A.P. The food contaminant deoxynivalenol activates the mitogen activated protein kinases in the intestine: Interest of ex vivo models as an alternative to in vivo experiments. Toxicon 2013, 66, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellafiora, L.; Filipello, V.; Dall’Asta, C.; Finazzi, G.; Galaverna, G.; Losio, M.N. A Structural Study on the Listeria Monocytogenes Internalin A-Human E-cadherin Interaction: A Molecular Tool to Investigate the Effects of Missense Mutations. Toxins 2020, 12, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schelstraete, W.; Devreese, M.; Croubels, S. Comparative toxicokinetics of Fusarium mycotoxins in pigs and humans. Food Chem. Toxicol. 2020, 137, 111140. [Google Scholar] [CrossRef]
- Cheli, F.; Giromini, C.; Baldi, A. Mycotoxin mechanisms of action and health impact: ‘in vitro’ or ‘in vivo’ tests, that is the question. World Mycotoxin J. 2015, 8, 573–589. [Google Scholar] [CrossRef]
- van der Flier, L.G.; Clevers, H. Stem cells, self-renewal, and differentiation in the intestinal epithelium. Annu. Rev. Physiol. 2009, 71, 241–260. [Google Scholar] [CrossRef]
- Nones, J.; Solhaug, A.; Eriksen, G.S.; Macuvele, D.L.P.; Poli, A.; Soares, C.; Trentin, A.G.; Riella, H.G.; Nones, J. Bentonite modified with zinc enhances aflatoxin B1 adsorption and increase survival of fibroblasts (3T3) and epithelial colorectal adenocarcinoma cells (Caco-2). J. Hazard. Mater. 2017, 337, 80–89. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, N.; Liu, J.; Li, F.D.; Li, S.L.; Wang, J.Q. Aflatoxin B1 and aflatoxin M1 induced cytotoxicity and DNA damage in differentiated and undifferentiated Caco-2 cells. Food Chem. Toxicol. 2015, 83, 54–60. [Google Scholar] [CrossRef]
- Zheng, N.; Zhang, H.; Li, S.; Wang, J.; Liu, J.; Ren, H.; Gao, Y. Lactoferrin inhibits aflatoxin B1- and aflatoxin M1-induced cytotoxicity and DNA damage in Caco-2, HEK, Hep-G2, and SK-N-SH cells. Toxicon 2018, 150, 77–85. [Google Scholar] [CrossRef]
- Bao, X.Y.; Li, S.L.; Gao, Y.N.; Wang, J.Q.; Zheng, N. Transcriptome analysis revealed that aflatoxin M1 could cause cell cycle arrest in differentiated Caco-2 cells. Toxicol. Vitro 2019, 59, 35–43. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Y.; Zhai, N.; Chen, X.; Gan, F.; Li, H.; Huang, K. Ochratoxin A-Induced Apoptosis of IPEC-J2 Cells through ROS-Mediated Mitochondrial Permeability Transition Pore Opening Pathway. J. Agric. Food Chem. 2017, 65, 10630–10637. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Liu, J.; Liang, Z. Probiotic Bacillus subtilis CW14 reduces disruption of the epithelial barrier and toxicity of ochratoxin A to Caco-2cells. Food Chem. Toxicol. 2019, 126, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gao, Y.; Yan, Q.; Bao, X.; Zhao, S.; Wang, J.; Zheng, N. Transcriptome Analysis of Ochratoxin A-Induced Apoptosis in Differentiated Caco-2 Cells. Toxins 2019, 12, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano-Sancho, G.; Gonzalez-Arias, C.A.; Ramos, A.J.; Sanchis, V.; Fernandez-Cruz, M.L. Cytotoxicity of the mycotoxins deoxynivalenol and ochratoxin A on Caco-2 cell line in presence of resveratrol. Toxicol. Vitro 2015, 29, 1639–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, G.; Fontanella, B.; Severino, L.; Quaroni, A.; Autore, G.; Marzocco, S. Nivalenol and deoxynivalenol affect rat intestinal epithelial cells: A concentration related study. PLoS ONE 2012, 7, e52051. [Google Scholar] [CrossRef]
- Diesing, A.K.; Nossol, C.; Danicke, S.; Walk, N.; Post, A.; Kahlert, S.; Rothkotter, H.J.; Kluess, J. Vulnerability of polarised intestinal porcine epithelial cells to mycotoxin deoxynivalenol depends on the route of application. PLoS ONE 2011, 6, e17472. [Google Scholar] [CrossRef] [Green Version]
- Diesing, A.K.; Nossol, C.; Panther, P.; Walk, N.; Post, A.; Kluess, J.; Kreutzmann, P.; Danicke, S.; Rothkotter, H.J.; Kahlert, S. Mycotoxin deoxynivalenol (DON) mediates biphasic cellular response in intestinal porcine epithelial cell lines IPEC-1 and IPEC-J2. Toxicol. Lett. 2011, 200, 8–18. [Google Scholar] [CrossRef]
- Broekaert, N.; Devreese, M.; Demeyere, K.; Berthiller, F.; Michlmayr, H.; Varga, E.; Adam, G.; Meyer, E.; Croubels, S. Comparative in vitro cytotoxicity of modified deoxynivalenol on porcine intestinal epithelial cells. Food Chem. Toxicol. 2016, 95, 103–109. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Fang, H.; Zhao, Y.; Jin, Y.; Shen, J.; Zhou, C.; Zhou, Y.; Fu, Y.; Wang, J.; et al. Transcriptional profiling of zearalenone-induced inhibition of IPEC-J2 cell proliferation. Toxicon 2019, 172, 8–14. [Google Scholar] [CrossRef]
- Shen, T.; Miao, Y.; Ding, C.; Fan, W.; Liu, S.; Lv, Y.; Gao, X.; De Boevre, M.; Yan, L.; Okoth, S.; et al. Activation of the p38/MAPK pathway regulates autophagy in response to the CYPOR-dependent oxidative stress induced by zearalenone in porcine intestinal epithelial cells. Food Chem. Toxicol. 2019, 131, 110527. [Google Scholar] [CrossRef]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Beridan Neagoe, I.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Abassi, H.; Ayed-Boussema, I.; Shirley, S.; Abid, S.; Bacha, H.; Micheau, O. The mycotoxin zearalenone enhances cell proliferation, colony formation and promotes cell migration in the human colon carcinoma cell line HCT116. Toxicol. Lett. 2016, 254, 1–7. [Google Scholar] [CrossRef]
- Minervini, F.; Garbetta, A.; D’Antuono, I.; Cardinali, A.; Martino, N.A.; Debellis, L.; Visconti, A. Toxic mechanisms induced by fumonisin b1 mycotoxin on human intestinal cell line. Arch. Environ. Contam. Toxicol. 2014, 67, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Assuncao, R.; Alvito, P.; Kleiveland, C.R.; Lea, T.E. Characterization of in vitro effects of patulin on intestinal epithelial and immune cells. Toxicol. Lett. 2016, 250–251, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Assuncao, R.; Pinhao, M.; Loureiro, S.; Alvito, P.; Silva, M.J. A multi-endpoint approach to the combined toxic effects of patulin and ochratoxin a in human intestinal cells. Toxicol. Lett. 2019, 313, 120–129. [Google Scholar] [CrossRef]
- Boussabbeh, M.; Ben Salem, I.; Prola, A.; Guilbert, A.; Bacha, H.; Abid-Essefi, S.; Lemaire, C. Patulin induces apoptosis through ROS-mediated endoplasmic reticulum stress pathway. Toxicol. Sci. 2015, 144, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.; Soung, N.K.; Thimmegowda, N.R.; Jeong, S.J.; Jang, J.H.; Moon, D.O.; Chung, J.K.; Lee, K.S.; Kwon, Y.T.; Erikson, R.L.; et al. Patulin induces colorectal cancer cells apoptosis through EGR-1 dependent ATF3 up-regulation. Cell Signal 2012, 24, 943–950. [Google Scholar] [CrossRef] [Green Version]
- Salah, A.; Bouaziz, C.; Amara, I.; Abid-Essefi, S.; Bacha, H. Eugenol protects against citrinin-induced cytotoxicity and oxidative damages in cultured human colorectal HCT116 cells. Environ. Sci. Pollut. Res. Int. 2019, 26, 31374–31383. [Google Scholar] [CrossRef]
- Akinrinmade, F.J.; Akinrinde, A.S.; Amid, A. Changes in serum cytokine levels, hepatic and intestinal morphology in aflatoxin B1-induced injury: Modulatory roles of melatonin and flavonoid-rich fractions from Chromolena odorata. Mycotoxin Res. 2016, 32, 53–60. [Google Scholar] [CrossRef]
- Jebali, R.; Ben Salah-Abbes, J.; Abbes, S.; Hassan, A.M.; Abdel-Aziem, S.H.; El-Nekeety, A.A.; Oueslati, R.; Abdel-Wahhab, M.A. Lactobacillus plantarum alleviate aflatoxins (B1 and M1) induced disturbances in the intestinal genes expression and DNA fragmentation in mice. Toxicon 2018, 146, 13–23. [Google Scholar] [CrossRef]
- Chen, X.; Naehrer, K.; Applegate, T.J. Interactive effects of dietary protein concentration and aflatoxin B1 on performance, nutrient digestibility, and gut health in broiler chicks. Poult. Sci. 2016, 95, 1312–1325. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, J.Q.; Jia, S.C.; Chen, Y.K.; Wang, J.P. Effect of yeast cell wall on the growth performance and gut health of broilers challenged with aflatoxin B1 and necrotic enteritis. Poult. Sci. 2018, 97, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.D.; He, J.; Ao, X.; Chen, D.W. Effects of maize naturally contaminated with aflatoxin B1 on growth performance, intestinal morphology, and digestive physiology in ducks. Poult. Sci. 2017, 96, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Jahanian, E.; Mahdavi, A.H.; Asgary, S.; Jahanian, R. Effect of dietary supplementation of mannanoligosaccharides on growth performance, ileal microbial counts, and jejunal morphology in broiler chicks exposed to aflatoxins. Livest. Sci. 2016, 190, 123–130. [Google Scholar] [CrossRef]
- Gonzalez-Arias, C.A.; Crespo-Sempere, A.; Marin, S.; Sanchis, V.; Ramos, A.J. Modulation of the xenobiotic transformation system and inflammatory response by ochratoxin A exposure using a co-culture system of Caco-2 and HepG2 cells. Food Chem. Toxicol. 2015, 86, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Ying, C.; Hong, W.; Nianhui, Z.; Chunlei, W.; Kehe, H.; Cuiling, P. Nontoxic concentrations of OTA aggravate DON-induced intestinal barrier dysfunction in IPEC-J2 cells via activation of NF-kappaB signaling pathway. Toxicol. Lett. 2019, 311, 114–124. [Google Scholar] [CrossRef]
- Tong, C.; Li, P.; Yu, L.-H.; Li, L.; Li, K.; Chen, Y.; Yang, S.-H.; Long, M. Selenium-rich yeast attenuates ochratoxin A-induced small intestinal injury in broiler chickens by activating the Nrf2 pathway and inhibiting NF-KB activation. J. Funct. Foods 2020, 66. [Google Scholar] [CrossRef]
- Luo, S.; Terciolo, C.; Bracarense, A.; Payros, D.; Pinton, P.; Oswald, I.P. In vitro and in vivo effects of a mycotoxin, deoxynivalenol, and a trace metal, cadmium, alone or in a mixture on the intestinal barrier. Environ. Int. 2019, 132, 105082. [Google Scholar] [CrossRef]
- Li, X.G.; Zhu, M.; Chen, M.X.; Fan, H.B.; Fu, H.L.; Zhou, J.Y.; Zhai, Z.Y.; Gao, C.Q.; Yan, H.C.; Wang, X.Q. Acute exposure to deoxynivalenol inhibits porcine enteroid activity via suppression of the Wnt/beta-catenin pathway. Toxicol. Lett. 2019, 305, 19–31. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Wang, Z.; Zhang, S.W.; Lin, H.L.; Gao, C.Q.; Zhao, J.C.; Yang, C.; Wang, X.Q. Methionine and Its Hydroxyl Analogues Improve Stem Cell Activity To Eliminate Deoxynivalenol-Induced Intestinal Injury by Reactivating Wnt/beta-Catenin Signaling. J. Agric. Food Chem. 2019, 67, 11464–11473. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Zhang, S.W.; Lin, H.L.; Gao, C.Q.; Yan, H.C.; Wang, X.Q. Hydrolyzed wheat gluten alleviates deoxynivalenol-induced intestinal injury by promoting intestinal stem cell proliferation and differentiation via upregulation of Wnt/beta-catenin signaling in mice. Food Chem. Toxicol. 2019, 131, 110579. [Google Scholar] [CrossRef] [PubMed]
- Martinez, G.; Dieguez, S.N.; Fernandez Paggi, M.B.; Riccio, M.B.; Perez Gaudio, D.S.; Rodriguez, E.; Amanto, F.A.; Tapia, M.O.; Soraci, A.L. Effect of fosfomycin, Cynara scolymus extract, deoxynivalenol and their combinations on intestinal health of weaned piglets. Anim. Nutr. 2019, 5, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wu, P.; Jiang, W.D.; Liu, Y.; Zeng, Y.Y.; Jiang, J.; Kuang, S.Y.; Tang, L.; Zhang, Y.A.; Zhou, X.Q.; et al. Deoxynivalenol decreased the growth performance and impaired intestinal physical barrier in juvenile grass carp (Ctenopharyngodon idella). Fish. Shellfish Immunol. 2018, 80, 376–391. [Google Scholar] [CrossRef]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Hess, M.; Bohm, J. Single and combined effects of deoxynivalenol mycotoxin and a microbial feed additive on lymphocyte DNA damage and oxidative stress in broiler chickens. PLoS ONE 2014, 9, e88028. [Google Scholar] [CrossRef] [PubMed]
- Gerez, J.R.; Pinton, P.; Callu, P.; Grosjean, F.; Oswald, I.P.; Bracarense, A.P. Deoxynivalenol alone or in combination with nivalenol and zearalenone induce systemic histological changes in pigs. Exp. Toxicol. Pathol. 2015, 67, 89–98. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Su, Y.; Shen, D.; Dai, P.; Li, C. Short-term ingestion of deoxynivalenol in naturally contaminated feed alters piglet performance and gut hormone secretion. Anim. Sci. J. 2018, 89, 1134–1143. [Google Scholar] [CrossRef]
- Yunus, A.W.; Blajet-Kosicka, A.; Kosicki, R.; Khan, M.Z.; Rehman, H.; Bohm, J. Deoxynivalenol as a contaminant of broiler feed: Intestinal development, absorptive functionality, and metabolism of the mycotoxin. Poult. Sci. 2012, 91, 852–861. [Google Scholar] [CrossRef]
- Marin, D.E.; Motiu, M.; Taranu, I. Food contaminant zearalenone and its metabolites affect cytokine synthesis and intestinal epithelial integrity of porcine cells. Toxins (Basel) 2015, 7, 1979–1988. [Google Scholar] [CrossRef]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Jiang, S.; Huang, L.; Wang, Y.; Yang, W.; Yang, Z.; Ge, J. Effects of zearalenone-induced oxidative stress and Keap1-Nrf2 signaling pathway-related gene expression in the ileum and mesenteric lymph nodes of post-weaning gilts. Toxicology 2020, 429, 152337. [Google Scholar] [CrossRef]
- Bracarense, A.P.; Lucioli, J.; Grenier, B.; Drociunas Pacheco, G.; Moll, W.D.; Schatzmayr, G.; Oswald, I.P. Chronic ingestion of deoxynivalenol and fumonisin, alone or in interaction, induces morphological and immunological changes in the intestine of piglets. Br. J. Nutr. 2012, 107, 1776–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawauchiya, T.; Takumi, R.; Kudo, Y.; Takamori, A.; Sasagawa, T.; Takahashi, K.; Kikuchi, H. Correlation between the destruction of tight junction by patulin treatment and increase of phosphorylation of ZO-1 in Caco-2 human colon cancer cells. Toxicol. Lett. 2011, 205, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Maidana, L.; Gerez, J.R.; El Khoury, R.; Pinho, F.; Puel, O.; Oswald, I.P.; Bracarense, A. Effects of patulin and ascladiol on porcine intestinal mucosa: An ex vivo approach. Food Chem. Toxicol. 2016, 98, 189–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, R.; Sun, Y.; Ye, W.; Zheng, T.; Wen, J.; Deng, Y. T-2 toxin inhibits the production of mucin via activating the IRE1/XBP1 pathway. Toxicology 2019, 424, 152230. [Google Scholar] [CrossRef] [PubMed]
- Sklan, D.; Shelly, M.; Makovsky, B.; Geyra, A.; Klipper, E.; Friedman, A. The effect of chronic feeding of diacetoxyscirpenol and T-2 toxin on performance, health, small intestinal physiology and antibody production in turkey poults. Br. Poult. Sci. 2003, 44, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Angius, F.; Spolitu, S.; Uda, S.; Deligia, S.; Frau, A.; Banni, S.; Collu, M.; Accossu, S.; Madeddu, C.; Serpe, R.; et al. High-density lipoprotein contribute to G0-G1/S transition in Swiss NIH/3T3 fibroblasts. Sci. Rep. 2015, 5, 17812. [Google Scholar] [CrossRef] [Green Version]
- Katsuyama, A.; Konno, T.; Shimoyama, S.; Kikuchi, H. The mycotoxin patulin decreases expression of density-enhanced phosphatase-1 by down-regulating PPARgamma in human colon cancer cells. Tohoku J. Exp. Med. 2014, 233, 265–274. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhai, N.; Chen, Y.; Fu, C.; Huang, K. OTA induces intestinal epithelial barrier dysfunction and tight junction disruption in IPEC-J2 cells through ROS/Ca(2+)-mediated MLCK activation. Environ. Pollut. 2018, 242, 106–112. [Google Scholar] [CrossRef]
- Garbetta, A.; Debellis, L.; De Girolamo, A.; Schena, R.; Visconti, A.; Minervini, F. Dose-dependent lipid peroxidation induction on ex vivo intestine tracts exposed to chyme samples from fumonisins contaminated corn samples. Toxicol. Vitro 2015, 29, 1140–1145. [Google Scholar] [CrossRef]
- Al-Saffar, A.K.; Meijer, C.H.; Gannavarapu, V.R.; Hall, G.; Li, Y.; Diaz Tartera, H.O.; Lordal, M.; Ljung, T.; Hellstrom, P.M.; Webb, D.L. Parallel Changes in Harvey-Bradshaw Index, TNFalpha, and Intestinal Fatty Acid Binding Protein in Response to Infliximab in Crohn’s Disease. Gastroenterol. Res. Pract. 2017, 2017, 1745918. [Google Scholar] [CrossRef] [Green Version]
- Webb, D.L. Tests of intestinal mucosal hyperpermeability: Many diseases, many biomarkers and a bright future. Best Pract. Res. Clin. Gastroenterol. 2019, 40–41, 101636. [Google Scholar] [CrossRef] [PubMed]
- Jahanian, E.; Mahdavi, A.H.; Asgary, S.; Jahanian, R. Effects of dietary inclusion of silymarin on performance, intestinal morphology and ileal bacterial count in aflatoxin-challenged broiler chicks. J. Anim. Physiol. Anim. Nutr. (Berl) 2017, 101, e43–e54. [Google Scholar] [CrossRef] [PubMed]
- Applegate, T.J.; Schatzmayr, G.; Prickel, K.; Troche, C.; Jiang, Z. Effect of aflatoxin culture on intestinal function and nutrient loss in laying hens. Poult. Sci. 2009, 88, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Graziani, F.; Pujol, A.; Nicoletti, C.; Paris, O.; Ernouf, P.; Di Pasquale, E.; Perrier, J.; Oswald, I.P.; Maresca, M. Deoxynivalenol inhibits the expression by goblet cells of intestinal mucins through a PKR and MAP kinase dependent repression of the resistin-like molecule beta. Mol. Nutr. Food Res. 2015, 59, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.K.; Wan, L.Y.M.; Turner, P.C.; El-Nezami, H. Individual and combined effects of deoxynivalenol and zearalenone on modulation of mucin expression on intestinal epithelial cells. Toxicol. Lett. 2016, 259, S135–S136. [Google Scholar] [CrossRef]
- Wan, L.Y.; Allen, K.J.; Turner, P.C.; El-Nezami, H. Modulation of mucin mRNA (MUC5AC and MUC5B) expression and protein production and secretion in Caco-2/HT29-MTX co-cultures following exposure to individual and combined Fusarium mycotoxins. Toxicol. Sci. 2014, 139, 83–98. [Google Scholar] [CrossRef] [Green Version]
- Wan, M.L.; Woo, C.S.; Allen, K.J.; Turner, P.C.; El-Nezami, H. Modulation of porcine beta-defensins 1 and 2 upon individual and combined Fusarium toxin exposure in a swine jejunal epithelial cell line. Appl. Environ. Microbiol. 2013, 79, 2225–2232. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Feng, L.; Jiang, W.D.; Wu, P.; Liu, Y.; Zeng, Y.Y.; Jiang, J.; Kuang, S.Y.; Tang, L.; Zhou, X.Q. Deoxynivalenol decreased intestinal immune function related to NF-kappaB and TOR signalling in juvenile grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2019, 84, 470–484. [Google Scholar] [CrossRef]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Janssens, G.P.; De Baere, S.; Mountzouris, K.C.; Su, S.; Wong, E.A.; De Meulenaer, B.; et al. Mycotoxins Deoxynivalenol and Fumonisins Alter the Extrinsic Component of Intestinal Barrier in Broiler Chickens. J. Agric. Food Chem. 2015, 63, 10846–10855. [Google Scholar] [CrossRef]
- Lin, R.; Sun, Y.; Mu, P.; Zheng, T.; Mu, H.; Deng, F.; Deng, Y.; Wen, J. Lactobacillus rhamnosus GG supplementation modulates the gut microbiota to promote butyrate production, protecting against deoxynivalenol exposure in nude mice. Biochem. Pharmacol. 2020, 175, 113868. [Google Scholar] [CrossRef]
- Wu, L.; Liao, P.; He, L.; Ren, W.; Yin, J.; Duan, J.; Li, T. Growth performance, serum biochemical profile, jejunal morphology, and the expression of nutrients transporter genes in deoxynivalenol (DON)- challenged growing pigs. BMC Vet. Res. 2015, 11, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewczuk, B.; Przybylska-Gornowicz, B.; Gajecka, M.; Targonska, K.; Ziolkowska, N.; Prusik, M.; Gajecki, M. Histological structure of duodenum in gilts receiving low doses of zearalenone and deoxynivalenol in feed. Exp. Toxicol. Pathol. 2016, 68, 157–166. [Google Scholar] [CrossRef]
- Levkut, M.; Revajová, V.; Herich, R.; Levkutová, M.; Ševčíková, Z.; Levkut, M. Effect of Glucan and Low Doses of T-2 Toxin on Mucin, IgA Expression and IgA+ Lymphocytes in the Intestine of Chickens. J. Comp. Pathol. 2015, 152, 51–52. [Google Scholar] [CrossRef]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, M.L.Y.; Turner, P.C.; Allen, K.J.; El-Nezami, H. Lactobacillus rhamnosus GG modulates intestinal mucosal barrier and inflammation in mice following combined dietary exposure to deoxynivalenol and zearalenone. J. Funct. Foods 2016, 22, 34–43. [Google Scholar] [CrossRef]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef]
- Taranu, I.; Marin, D.E.; Palade, M.; Pistol, G.C.; Chedea, V.S.; Gras, M.A.; Rotar, C. Assessment of the efficacy of a grape seed waste in counteracting the changes induced by aflatoxin B1 contaminated diet on performance, plasma, liver and intestinal tissues of pigs after weaning. Toxicon 2019, 162, 24–31. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, B.; Liu, M.; Jiang, K.; Wang, M.; Wang, L. Comparative transcriptome analysis reveals the different roles between hepatopancreas and intestine of Litopenaeus vannamei in immune response to aflatoxin B1 (AFB1) challenge. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2019, 222, 1–10. [Google Scholar] [CrossRef]
- Kraieski, A.L.; Hayashi, R.M.; Sanches, A.; Almeida, G.C.; Santin, E. Effect of aflatoxin experimental ingestion and Eimeira vaccine challenges on intestinal histopathology and immune cellular dynamic of broilers: Applying an Intestinal Health Index. Poult. Sci. 2017, 96, 1078–1087. [Google Scholar] [CrossRef]
- Ruan, D.; Wang, W.C.; Lin, C.X.; Fouad, A.M.; Chen, W.; Xia, W.G.; Wang, S.; Luo, X.; Zhang, W.H.; Yan, S.J.; et al. Effects of curcumin on performance, antioxidation, intestinal barrier and mitochondrial function in ducks fed corn contaminated with ochratoxin A. Animal 2019, 13, 42–52. [Google Scholar] [CrossRef]
- Kang, R.; Li, R.; Dai, P.; Li, Z.; Li, Y.; Li, C. Deoxynivalenol induced apoptosis and inflammation of IPEC-J2 cells by promoting ROS production. Environ. Pollut. 2019, 251, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Ling, K.H.; Wan, M.L.; El-Nezami, H.; Wang, M. Protective Capacity of Resveratrol, a Natural Polyphenolic Compound, against Deoxynivalenol-Induced Intestinal Barrier Dysfunction and Bacterial Translocation. Chem. Res. Toxicol. 2016, 29, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.; Yang, H.; Lee, S.H. Modulation of early growth response gene 1 and interleukin-8 expression by ribotoxin deoxynivalenol (vomitoxin) via ERK1/2 in human epithelial intestine 407 cells. Biochem. Biophys. Res. Commun. 2007, 362, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kim, J.; Kim, D.; Moon, Y. Mycotoxin detoxifiers attenuate deoxynivalenol-induced pro-inflammatory barrier insult in porcine enterocytes as an in vitro evaluation model of feed mycotoxin reduction. Toxicol. Vitro 2017, 38, 108–116. [Google Scholar] [CrossRef]
- Lessard, M.; Savard, C.; Deschene, K.; Lauzon, K.; Pinilla, V.A.; Gagnon, C.A.; Lapointe, J.; Guay, F.; Chorfi, Y. Impact of deoxynivalenol (DON) contaminated feed on intestinal integrity and immune response in swine. Food Chem. Toxicol. 2015, 80, 7–16. [Google Scholar] [CrossRef]
- Taranu, I.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Pelinescu, D. Induction of pro-inflammatory gene expression by Escherichia coli and mycotoxin zearalenone contamination and protection by a Lactobacillus mixture in porcine IPEC-1 cells. Toxicon 2015, 97, 53–63. [Google Scholar] [CrossRef]
- Fan, W.; Lv, Y.; Ren, S.; Shao, M.; Shen, T.; Huang, K.; Zhou, J.; Yan, L.; Song, S. Zearalenone (ZEA)-induced intestinal inflammation is mediated by the NLRP3 inflammasome. Chemosphere 2018, 190, 272–279. [Google Scholar] [CrossRef]
- Bouhet, S.; Le Dorze, E.; Peres, S.; Fairbrother, J.M.; Oswald, I.P. Mycotoxin fumonisin B1 selectively down-regulates the basal IL-8 expression in pig intestine: In vivo and in vitro studies. Food Chem. Toxicol. 2006, 44, 1768–1773. [Google Scholar] [CrossRef]
- Islam, M.R.; Roh, Y.S.; Cho, A.; Kim, J.; Kim, J.H.; Eo, S.K.; Lim, C.W.; Kim, B. Immune modulatory effects of the foodborne contaminant citrinin in mice. Food Chem. Toxicol. 2012, 50, 3537–3547. [Google Scholar] [CrossRef]
- Prado-Garcia, H.; Romero-Garcia, S.; Aguilar-Cazares, D.; Meneses-Flores, M.; Lopez-Gonzalez, J.S. Tumor-induced CD8+ T-cell dysfunction in lung cancer patients. Clin. Dev. Immunol. 2012, 2012, 741741. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.H.; Lillehoj, H.S.; Lillehoj, E.P. Intestinal immune responses to coccidiosis. Dev. Comp. Immunol. 2000, 24, 303–324. [Google Scholar] [CrossRef]
- Girgis, G.N.; Barta, J.R.; Girish, C.K.; Karrow, N.A.; Boermans, H.J.; Smith, T.K. Effects of feed-borne Fusarium mycotoxins and an organic mycotoxin adsorbent on immune cell dynamics in the jejunum of chickens infected with Eimeria maxima. Vet. Immunol. Immunopathol. 2010, 138, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tang, L.; Glenn, T.C.; Wang, J.S. Aflatoxin B1 Induced Compositional Changes in Gut Microbial Communities of Male F344 Rats. Toxicol. Sci. 2016, 150, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Liu, L.; Chen, J.; Xiao, A. Response of Intestinal Bacterial Flora to the Long-term Feeding of Aflatoxin B1 (AFB1) in Mice. Toxins 2017, 9, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galarza-Seeber, R.; Latorre, J.D.; Bielke, L.R.; Kuttappan, V.A.; Wolfenden, A.D.; Hernandez-Velasco, X.; Merino-Guzman, R.; Vicente, J.L.; Donoghue, A.; Cross, D.; et al. Leaky Gut and Mycotoxins: Aflatoxin B1 Does Not Increase Gut Permeability in Broiler Chickens. Front. Vet. Sci. 2016, 3, 10. [Google Scholar] [CrossRef]
- Maresca, M.; Yahi, N.; Younes-Sakr, L.; Boyron, M.; Caporiccio, B.; Fantini, J. Both direct and indirect effects account for the pro-inflammatory activity of enteropathogenic mycotoxins on the human intestinal epithelium: Stimulation of interleukin-8 secretion, potentiation of interleukin-1beta effect and increase in the transepithelial passage of commensal bacteria. Toxicol. Appl. Pharmacol. 2008, 228, 84–92. [Google Scholar] [CrossRef]
- Ouethrani, M.; Van de Wiele, T.; Verbeke, E.; Bruneau, A.; Carvalho, M.; Rabot, S.; Camel, V. Metabolic fate of ochratoxin A as a coffee contaminant in a dynamic simulator of the human colon. Food Chem. 2013, 141, 3291–3300. [Google Scholar] [CrossRef]
- Guo, M.; Huang, K.; Chen, S.; Qi, X.; He, X.; Cheng, W.H.; Luo, Y.; Xia, K.; Xu, W. Combination of metagenomics and culture-based methods to study the interaction between ochratoxin a and gut microbiota. Toxicol. Sci. 2014, 141, 314–323. [Google Scholar] [CrossRef]
- Pinton, P.; Nougayrede, J.P.; Del Rio, J.C.; Moreno, C.; Marin, D.E.; Ferrier, L.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. The food contaminant deoxynivalenol, decreases intestinal barrier permeability and reduces claudin expression. Toxicol. Appl. Pharmacol. 2009, 237, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.J.; Zhang, R.Q.; Zhai, Q.Y.; Liu, J.C.; Li, N.; Liu, W.X.; Li, L.; Shen, W. Metagenomic analysis of gut microbiota alteration in a mouse model exposed to mycotoxin deoxynivalenol. Toxicol. Appl. Pharmacol. 2019, 372, 47–56. [Google Scholar] [CrossRef]
- Lucke, A.; Bohm, J.; Zebeli, Q.; Metzler-Zebeli, B.U. Dietary Deoxynivalenol Contamination and Oral Lipopolysaccharide Challenge Alters the Cecal Microbiota of Broiler Chickens. Front. Microbiol. 2018, 9, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wache, Y.J.; Valat, C.; Postollec, G.; Bougeard, S.; Burel, C.; Oswald, I.P.; Fravalo, P. Impact of deoxynivalenol on the intestinal microflora of pigs. Int. J. Mol. Sci. 2009, 10, 1–17. [Google Scholar] [CrossRef]
- Miro-Abella, E.; Torrell, H.; Herrero, P.; Canela, N.; Arola, L.; Borrull, F.; Ras, R.; Fontanals, N. Monitoring and evaluation of the interaction between deoxynivalenol and gut microbiota in Wistar rats by mass spectrometry-based metabolomics and next-generation sequencing. Food Chem. Toxicol. 2018, 121, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, M.J.; Perrin-Guyomard, A.; Houee, P.; Rolland, J.G.; Laurentie, M. Evaluation of an oral subchronic exposure of deoxynivalenol on the composition of human gut microbiota in a model of human microbiota-associated rats. PLoS ONE 2013, 8, e80578. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Dobrindt, U.; Martin, P.; Secher, T.; Bracarense, A.P.; Boury, M.; Laffitte, J.; Pinton, P.; Oswald, E.; Oswald, I.P. The Food Contaminant Deoxynivalenol Exacerbates the Genotoxicity of Gut Microbiota. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, J.; Wang, Y.; Wang, K.; Wei, H.; Shen, L. Isolation and characterization of the Bacillus cereus BC7 strain, which is capable of zearalenone removal and intestinal flora modulation in mice. Toxicon 2018, 155, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Slizewska, K.; Nowak, A.; Zielonka, L.; Zakowska, Z.; Gajecka, M.; Gajecki, M. The effect of experimental fusarium mycotoxicosis on microbiota diversity in porcine ascending colon contents. Toxins 2014, 6, 2064–2081. [Google Scholar] [CrossRef] [Green Version]
- Burel, C.; Tanguy, M.; Guerre, P.; Boilletot, E.; Cariolet, R.; Queguiner, M.; Postollec, G.; Pinton, P.; Salvat, G.; Oswald, I.P.; et al. Effect of low dose of fumonisins on pig health: Immune status, intestinal microbiota and sensitivity to Salmonella. Toxins 2013, 5, 841–864. [Google Scholar] [CrossRef] [Green Version]
- Pinto, F.E.L., Jr.; Brandt, C.T.; Medeiros, A.C.; Oliveira, A.J.F.; Jeronimo, S.M.; Brito, H.M.F. Bacterial translocation in rats nonfunctioning diverted distal colon. Acta Cir. Bras. 2007, 22, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, B.; Liu, M.; Jiang, K.; Wang, M.; Wang, L. Aflatoxin B1 (AFB1) induced dysregulation of intestinal microbiota and damage of antioxidant system in pacific white shrimp (Litopenaeus vannamei). Aquaculture 2018, 495, 940–947. [Google Scholar] [CrossRef]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.S.; Pratt-Phillips, S.; Gonzalez, L.M. Alterations in Intestinal Permeability: The Role of the “Leaky Gut” in Health and Disease. J. Equine Vet. Sci. 2017, 52, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Choi, W.; Yeruva, S.; Turner, J.R. Contributions of intestinal epithelial barriers to health and disease. Exp. Cell Res. 2017, 358, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Dheer, R.; Santaolalla, R.; Davies, J.M.; Lang, J.K.; Phillips, M.C.; Pastorini, C.; Vazquez-Pertejo, M.T.; Abreu, M.T. Intestinal Epithelial Toll-Like Receptor 4 Signaling Affects Epithelial Function and Colonic Microbiota and Promotes a Risk for Transmissible Colitis. Infect. Immun. 2016, 84, 798–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barmeyer, C.; Schulzke, J.D.; Fromm, M. Claudin-related intestinal diseases. Semin. Cell Dev. Biol. 2015, 42, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Capaldo, C.T.; Nusrat, A. Claudin switching: Physiological plasticity of the Tight Junction. Semin. Cell Dev. Biol. 2015, 42, 22–29. [Google Scholar] [CrossRef]
- Goswami, P.; Das, P.; Verma, A.K.; Prakash, S.; Das, T.K.; Nag, T.C.; Ahuja, V.; Gupta, S.D.; Makharia, G.K. Are alterations of tight junctions at molecular and ultrastructural level different in duodenal biopsies of patients with celiac disease and Crohn’s disease? Virchows Arch. 2014, 465, 521–530. [Google Scholar] [CrossRef]
- Luissint, A.C.; Parkos, C.A.; Nusrat, A. Inflammation and the Intestinal Barrier: Leukocyte-Epithelial Cell Interactions, Cell Junction Remodeling, and Mucosal Repair. Gastroenterology 2016, 151, 616–632. [Google Scholar] [CrossRef] [Green Version]
- Oshima, T.; Miwa, H.; Joh, T. Changes in the expression of claudins in active ulcerative colitis. J. Gastroenterol. Hepatol. 2008, 23 (Suppl. 2), S146–S150. [Google Scholar] [CrossRef]
- Zeissig, S.; Burgel, N.; Gunzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef]
- Ahmad, R.; Chaturvedi, R.; Olivares-Villagomez, D.; Habib, T.; Asim, M.; Shivesh, P.; Polk, D.B.; Wilson, K.T.; Washington, M.K.; Van Kaer, L.; et al. Targeted colonic claudin-2 expression renders resistance to epithelial injury, induces immune suppression, and protects from colitis. Mucosal. Immunol. 2014, 7, 1340–1353. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Lu, Z.; Foreman, O.; Tatum, R.; Lu, Q.; Renegar, R.; Cao, J.; Chen, Y.H. Inflammation and disruption of the mucosal architecture in claudin-7-deficient mice. Gastroenterology 2012, 142, 305–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, M.; Yoshida, M.; Nishiumi, S.; Furuse, M.; Azuma, T. Claudin-2 regulates colorectal inflammation via myosin light chain kinase-dependent signaling. Dig. Dis. Sci. 2013, 58, 1546–1559. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A.; Shea-Donohue, T. Mechanisms of disease: The role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 416–422. [Google Scholar] [CrossRef]
- Capaldo, C.T.; Nusrat, A. Cytokine regulation of tight junctions. Biochim. Biophys. Acta 2009, 1788, 864–871. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, K.; McKenzie, A.N.; Neurath, M.F.; Weigmann, B. IL-9 regulates intestinal barrier function in experimental T cell-mediated colitis. Tissue Barriers 2015, 3, e983777. [Google Scholar] [CrossRef] [Green Version]
- Haines, R.J.; Beard, R.S., Jr.; Chen, L.; Eitnier, R.A.; Wu, M.H. Interleukin-1beta Mediates beta-Catenin-Driven Downregulation of Claudin-3 and Barrier Dysfunction in Caco2 Cells. Dig. Dis. Sci. 2016, 61, 2252–2261. [Google Scholar] [CrossRef] [Green Version]
- Haines, R.J.; Beard, R.S., Jr.; Eitner, R.A.; Chen, L.; Wu, M.H. TNFalpha/IFNgamma Mediated Intestinal Epithelial Barrier Dysfunction Is Attenuated by MicroRNA-93 Downregulation of PTK6 in Mouse Colonic Epithelial Cells. PLoS ONE 2016, 11, e0154351. [Google Scholar] [CrossRef] [Green Version]
- Mankertz, J.; Amasheh, M.; Krug, S.M.; Fromm, A.; Amasheh, S.; Hillenbrand, B.; Tavalali, S.; Fromm, M.; Schulzke, J.D. TNFalpha up-regulates claudin-2 expression in epithelial HT-29/B6 cells via phosphatidylinositol-3-kinase signaling. Cell Tissue Res. 2009, 336, 67–77. [Google Scholar] [CrossRef]
- Naydenov, N.G.; Baranwal, S.; Khan, S.; Feygin, A.; Gupta, P.; Ivanov, A.I. Novel mechanism of cytokine-induced disruption of epithelial barriers: Janus kinase and protein kinase D-dependent downregulation of junction protein expression. Tissue Barriers 2013, 1, e25231. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Yoshinaga, N.; Tanabe, S. Interleukin-6 (IL-6) regulates claudin-2 expression and tight junction permeability in intestinal epithelium. J. Biol. Chem. 2011, 286, 31263–31271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Chao, K.; Ng, S.C.; Bai, A.H.; Yu, Q.; Yu, J.; Li, M.; Cui, Y.; Chen, M.; Hu, J.F.; et al. Pro-inflammatory miR-223 mediates the cross-talk between the IL23 pathway and the intestinal barrier in inflammatory bowel disease. Genome Biol. 2016, 17, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, C.R.; Raleigh, D.R.; Su, L.; Shen, L.; Sullivan, E.A.; Wang, Y.; Turner, J.R. Epithelial myosin light chain kinase activation induces mucosal interleukin-13 expression to alter tight junction ion selectivity. J. Biol. Chem. 2010, 285, 12037–12046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spasova, D.S.; Surh, C.D. Blowing on embers: Commensal microbiota and our immune system. Front. Immunol. 2014, 5, 318. [Google Scholar] [CrossRef] [PubMed]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Kurashima, Y.; Goto, Y.; Kiyono, H. Mucosal innate immune cells regulate both gut homeostasis and intestinal inflammation. Eur. J. Immunol. 2013, 43, 3108–3115. [Google Scholar] [CrossRef]
- Katz, K.D.; Hollander, D.; Vadheim, C.M.; McElree, C.; Delahunty, T.; Dadufalza, V.D.; Krugliak, P.; Rotter, J.I. Intestinal permeability in patients with Crohn’s disease and their healthy relatives. Gastroenterology 1989, 97, 927–931. [Google Scholar] [CrossRef]
- Peeters, M.; Geypens, B.; Claus, D.; Nevens, H.; Ghoos, Y.; Verbeke, G.; Baert, F.; Vermeire, S.; Vlietinck, R.; Rutgeerts, P. Clustering of increased small intestinal permeability in families with Crohn’s disease. Gastroenterology 1997, 113, 802–807. [Google Scholar] [CrossRef]
- Warth, B.; Parich, A.; Atehnkeng, J.; Bandyopadhyay, R.; Schuhmacher, R.; Sulyok, M.; Krska, R. Quantitation of mycotoxins in food and feed from Burkina Faso and Mozambique using a modern LC-MS/MS multitoxin method. J. Agric. Food Chem. 2012, 60, 9352–9363. [Google Scholar] [CrossRef]
- Matumba, L.; Sulyok, M.; Monjerezi, M.; Biswick, T.; Krska, R. Fungal metabolites diversity in maize and associated human dietary exposures relate to micro-climatic patterns in Malawi. World Mycotoxin J. 2015, 8, 269–282. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, B.; He, X.; Cheng, W.H.; Xu, W.; Luo, Y.; Liang, R.; Luo, H.; Huang, K. Analysis of individual and combined effects of ochratoxin A and zearalenone on HepG2 and KK-1 cells with mathematical models. Toxins 2014, 6, 1177–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signorini, M.L.; Gaggiotti, M.; Molineri, A.; Chiericatti, C.A.; Zapata de Basilico, M.L.; Basilico, J.C.; Pisani, M. Exposure assessment of mycotoxins in cow’s milk in Argentina. Food Chem. Toxicol. 2012, 50, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Zheng, N.; Zheng, B.Q.; Wen, F.; Cheng, J.B.; Han, R.W.; Xu, X.M.; Li, S.L.; Wang, J.Q. Simultaneous determination of aflatoxin M1, ochratoxin A, zearalenone and alpha-zearalenol in milk by UHPLC-MS/MS. Food Chem. 2014, 146, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Xu, J.; Jiang, K.; Liu, X.; Meng, J.; Di Mavungu, J.D.; Guo, W.; Zhang, Z.; Jing, J.; Li, H.; et al. Determination of multiple mycotoxins in paired plasma and urine samples to assess human exposure in Nanjing, China. Environ. Pollut. 2019, 248, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Abia, W.A.; Warth, B.; Sulyok, M.; Krska, R.; Tchana, A.; Njobeh, P.B.; Turner, P.C.; Kouanfack, C.; Eyongetah, M.; Dutton, M.; et al. Bio-monitoring of mycotoxin exposure in Cameroon using a urinary multi-biomarker approach. Food Chem. Toxicol. 2013, 62, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Sabuncuoglu, S.; Erkekoglu, P.; Aydin, S.; Sahin, G.; Kocer-Gumusel, B. The effects of season and gender on the serum aflatoxins and ochratoxin A levels of healthy adult subjects from the Central Anatolia Region, Turkey. Eur. J. Nutr. 2015, 54, 629–638. [Google Scholar] [CrossRef]
- Shirima, C.P.; Kimanya, M.E.; Routledge, M.N.; Srey, C.; Kinabo, J.L.; Humpf, H.U.; Wild, C.P.; Tu, Y.K.; Gong, Y.Y. A prospective study of growth and biomarkers of exposure to aflatoxin and fumonisin during early childhood in Tanzania. Environ. Health Perspect. 2015, 123, 173–178. [Google Scholar] [CrossRef] [Green Version]
- Solfrizzo, M.; Gambacorta, L.; Visconti, A. Assessment of multi-mycotoxin exposure in southern Italy by urinary multi-biomarker determination. Toxins 2014, 6, 523–538. [Google Scholar] [CrossRef]
- Gao, Y.N.; Wang, J.Q.; Li, S.L.; Zhang, Y.D.; Zheng, N. Aflatoxin M1 cytotoxicity against human intestinal Caco-2 cells is enhanced in the presence of other mycotoxins. Food Chem. Toxicol. 2016, 96, 79–89. [Google Scholar] [CrossRef]
- Smith, M.C.; Gheux, A.; Coton, M.; Madec, S.; Hymery, N.; Coton, E. In vitro co-culture models to evaluate acute cytotoxicity of individual and combined mycotoxin exposures on Caco-2, THP-1 and HepaRG human cell lines. Chem. Biol. Interact. 2018, 281, 51–59. [Google Scholar] [CrossRef]
Figure 1.
Normal intestinal homeostasis. The intestinal barrier is equipped with four levels to protect the intestine from external stimuli. This includes a physical barrier (a single layer of semi-permeable epithelial cells), chemical barrier (a mucus layer consisting of mucins and antimicrobial peptides, secreted by goblet cells and Paneth cells, respectively), immunological barrier (immune cells in the lamina propria and secreted immune mediators such as cytokines and secretory immunoglobulin A (sIgA)), and microbial barrier (commensal bacteria in the intestinal lumen). Adjacent epithelial cells are connected by tight junctions, which are composed of transmembrane proteins, junctional adhesion molecules (JAMs), claudins, and occludin that are linked to the actin cytoskeleton through zonula occludens (ZO) proteins.
Figure 1.
Normal intestinal homeostasis. The intestinal barrier is equipped with four levels to protect the intestine from external stimuli. This includes a physical barrier (a single layer of semi-permeable epithelial cells), chemical barrier (a mucus layer consisting of mucins and antimicrobial peptides, secreted by goblet cells and Paneth cells, respectively), immunological barrier (immune cells in the lamina propria and secreted immune mediators such as cytokines and secretory immunoglobulin A (sIgA)), and microbial barrier (commensal bacteria in the intestinal lumen). Adjacent epithelial cells are connected by tight junctions, which are composed of transmembrane proteins, junctional adhesion molecules (JAMs), claudins, and occludin that are linked to the actin cytoskeleton through zonula occludens (ZO) proteins.

Figure 2.
Summary of the negative effects induced by mycotoxins on intestinal barrier. Relevant aspects include (i) increased permeability (paracellular and transcellular transport), which is induced by disrupted epithelial cells and tight junctions, and (ii) the thinned mucus layer. The compromised intestinal barrier results in the penetration of xenobiotics of different molecular weights and bacterial translocation, ultimately contributing to an imbalance of inflammatory responses and the activation of local and systemic immunity, causing the occurrence of inflammatory-related diseases.
Figure 2.
Summary of the negative effects induced by mycotoxins on intestinal barrier. Relevant aspects include (i) increased permeability (paracellular and transcellular transport), which is induced by disrupted epithelial cells and tight junctions, and (ii) the thinned mucus layer. The compromised intestinal barrier results in the penetration of xenobiotics of different molecular weights and bacterial translocation, ultimately contributing to an imbalance of inflammatory responses and the activation of local and systemic immunity, causing the occurrence of inflammatory-related diseases.


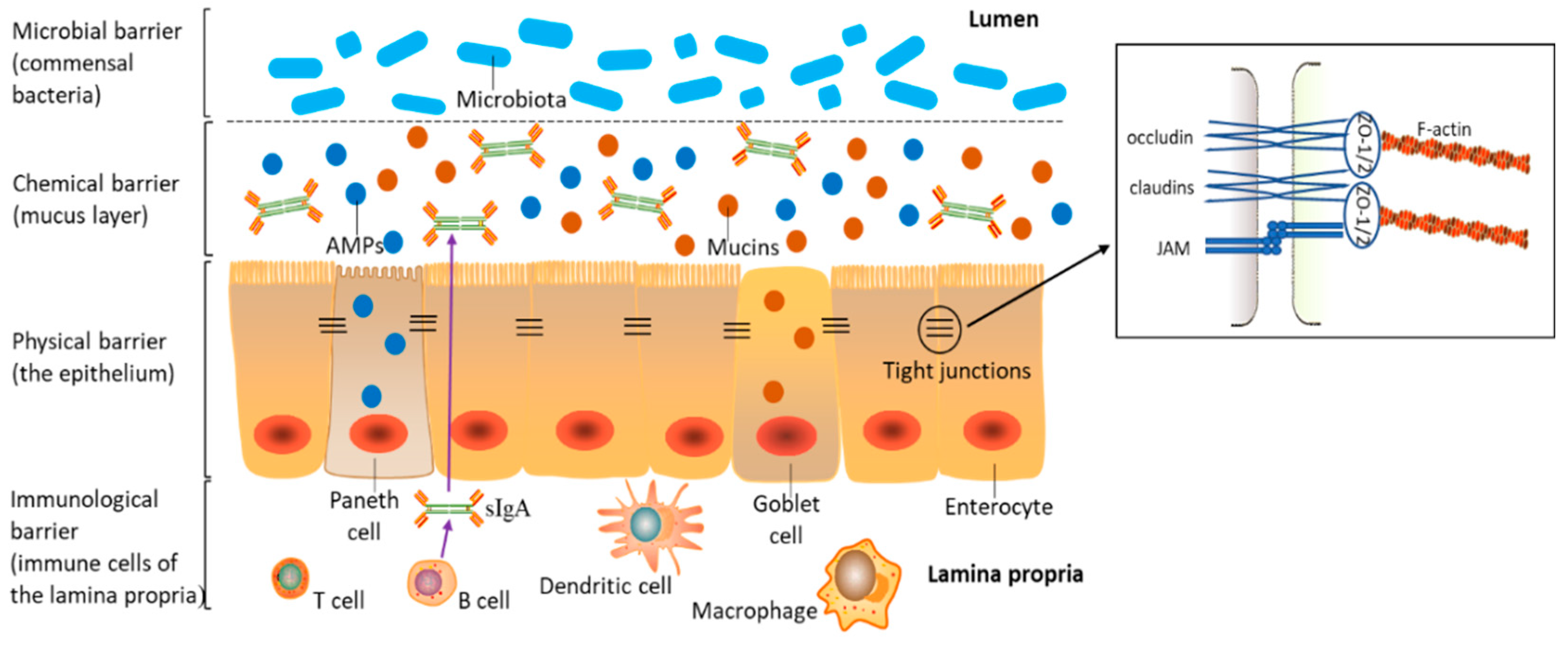{kind=link}
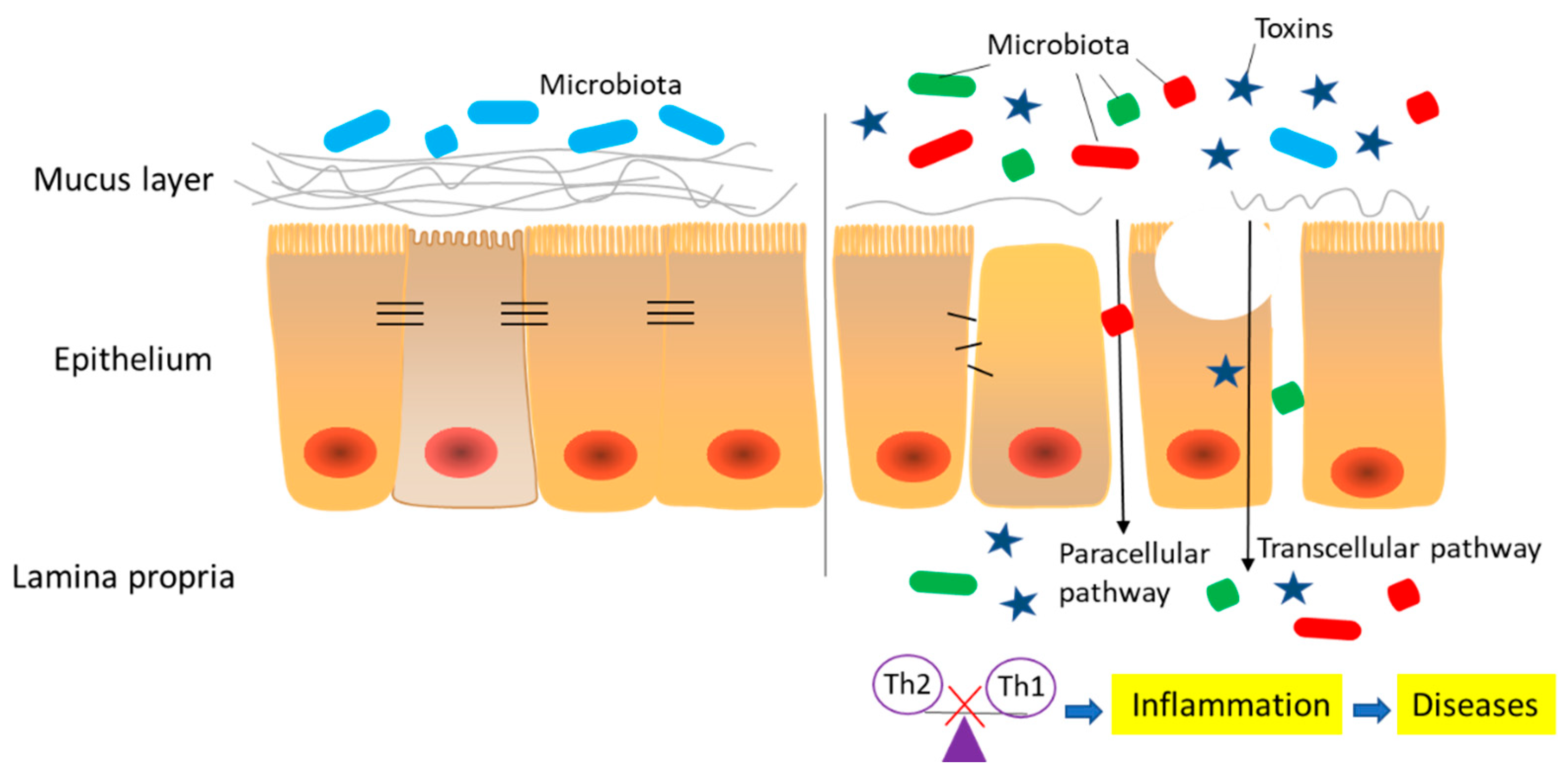{kind=link}
Table 1.
The overview of mycotoxins.
| Mycotoxin | Toxic Effects | IARC Classification | Health Guidance Value | References |
|---|---|---|---|---|
| AFB1, AFM1 | carcinogenic | Group 1 | As low as reasonably achievable (ALARA principle) | [5,6] |
| OTA | nephrotoxic, teratogenic, immunotoxic, neurotoxic | Group 2B | TWI = 120 ng/kg bw/w PTWI = 100 ng/kg bw/w | [5,6] |
| FB1 | hepatotoxicity, nephrotoxicity | Group 2B | PMTDI (FB1+FB2 +FB 3) = 2 μg/kg bw/d TDI (FB1+FB2 +FB3) = 2 μg/kg bw/d | [1,5,6] |
| DON | immunotoxic | Group 3 | TDI = 1 μg/kg bw/d | [5,6,7,8] |
| ZEN | reproductive toxicity | Group 3 | PMTDI = 0.5 μg/kg bw/d TDI = 0.25 μg/kg bw/d | [5,6] |
| PAT | hepatotoxicity | Group 3 | PMTDI = 0.4 μg/kg bw/d | [1,5,9] |
| NIV | immunotoxicity, hematotoxicity, myelotoxicity | Group 3 | TDI = 1.2 μg/kg bw/d | [2,5,10] |
| T-2, HT-2 | toxic on the skin and mucous membranes | Group 3 | PMTDI (T-2+HT-2) = 0.06 μg/kg bw/d TDI (T-2+HT-2) = 0.1 μg/kg bw/d | [5] |
Group 1, carcinogenic to humans; Group 2A, probably carcinogenic to humans; Group 2B, possibly carcinogenic to humans; Group 3, not classifiable as to its carcinogenicity to humans.
Table 2.
The characteristics of in vitro, ex vivo, and in vivo intestine models.
| Models | Types | Advantages | Limitations | References |
|---|---|---|---|---|
| In vitro-2D intestinal model | Caco-2 cells, IPEC-1 cells, IPEC-J2 cells, IPI-2I cells and PSI-1 cells, co-culture of different cell lines) | well-established and relatively cheap | only containing a single cell type without villus and crypt domain | [39,40,41,42,43,44] |
| In vitro-3D intestinal model | enteroids, also known as organoids or mini-guts | partially recapitulate the anatomy of native epithelium, have the ability to passage at an almost unlimited scale | the effects of substances on the luminal side are poorly investigated, considerable cost, do not contain the immune and stromal cells | [41,45,46,47,48,49] |
| Ex vivo | applied in humans, rodents, swine, poultry and horse | a more accurate model to mimic the physiology in vivo | fail to achieve long-term culture, careful and laborious preparation | [45,50,51] |
| In vivo | commonly used models include mouse, rat, chicken, turkey, fish, pig, sheep and bovine | provide the information based on the whole animals, thus they could corroborate the toxicity in humans effectively | the use of live animals should follow 3R (replacement, reduction and refinement) principle | [1] |
Table 3.
Modulation of intestinal epithelial cells (physical barrier) induced by mycotoxins.
| Model | Dose/Administration Route | Exposure Time | Technique | Damage | References |
|---|---|---|---|---|---|
| Aflatoxin | |||||
| Caco-2 cells | AFB1: 1–50 μM | 24 h | Alamar blue assay | Decrease cell viability | [77] |
| Caco-2 cells | AFB1: 0.01–1 μg/mL AFM1: 0.01–1 μg/mL | 24, 48, 72 h | MTT assay | AFB1: decrease cell viability AFM1: decrease cell viability | [78] |
| Caco-2 cells | AFB1: 4 μg/mL AFM1: 4 μg/mL | 24 h | MTT assay | AFB1: decrease cell viability AFM1: decrease cell viability | [79] |
| Caco-2 cells | AFM1: 0.0005–4 μg/mL | 48 h | RNA-Seq, CCK-8 assay, Flow cytometry analysis | No effect on cell viability Induce cell cycle arrest | [80] |
| Caco-2/HT29-MTX cells | AFM1: 0.05, 4 μg/mL | 48 h | CCK-8 assay | Decrease cell viability | [43] |
| Ochratoxin A | |||||
| Caco-2/HT29-MTX cells | OTA: 0.05, 4 μg/mL | 48 h | CCK-8 assay | Decrease cell viability | [43] |
| IPEC-J2 Cells | OTA: 0.5–32 μM | 6, 12, 24 h | MTT assay | Decrease cell viability | [81] |
| Caco-2 cells | OTA: 0.1–30 μM | 24 h | RNA-Seq, CCK-8 assay, Flow cytometry analysis | Decrease cell viability Arrest cell cycle in G2/M phase Induce apoptosis | [82] |
| Caco-2 cells | OTA: 0.0005–4 μg/mL | 48 h | RNA-Seq, Flow cytometry analysis | Induce cell apoptosis | [83] |
| Caco-2 cells | OTA: 0.5–160 μM | 48 h | MTS assay | Decrease cell viability | [84] |
| Deoxynivalenol | |||||
| Caco-2 cells | DON: 0.25–30 μM | 48 h | MTS assay | Decrease cell viability | [84] |
| HT-29 cells | DON: 125–2000 ng/mL | 24 h | Western blot analysis | Induce cell apoptosis | [24] |
| IEC-6 cells | DON: 0.5–80 μM | 24 h | Propidium iodide staining | Induce cell apoptosis | [85] |
| IPEC-J2 Cells | DON: 200, 2000 ng/mL | 24, 48, 72 h | BrdU incorporation assay, Western blot analysis | Decrease cell viability Induce cell apoptosis | [86] |
| IPEC-1 cells, IPEC-J2 Cells | DON: 100–4000 ng/mL | 24, 48, 72 h | MTT assay | Decrease cell viability | [87] |
| IPEC-J2 Cells | DON: 1–20 μg/mL | 72 h | Flow cytometry analysis | Decrease cell viability Induce cell apoptosis | [88] |
| Zearalenone | |||||
| IPEC-J2 Cells | ZEN: 40 μM | 24 h | CCK-8 assay, Flow cytometry analysis, RNA-Seq | Decrease cell viability Arrest cell cycle in the G2/M phase | [89] |
| IPEC-J2 cells | ZEN: 6, 8 μg/mL | 12–48 h | MTT assay, PCR | Decrease cell viability Induce cell apoptosis | [90] |
| IPEC-1 cells | ZEN: 0.1–100 μM | 24 h | XTT assay, Microarray assay | Decrease cell viability | [91] |
| HCT116 cells | ZEN: 0–320 μM | 48 h | Methylene blue staining assay | Increase cell viability at very low concentrations, decrease cell viability at high concentrations | [92] |
| Fumonisin B1 | |||||
| HT-29 cells | FB1: 1.1–69 μM | 72 h | MTT assay | Decrease cell viability | [93] |
| Patulin | |||||
| Caco-2 cells | PAT: 1–150 μM | 24 h | MTT assay | Decrease cell viability | [94] |
| Caco-2 cells | PAT: 0.7–18 μM | 24 h | MTT assay | Decrease cell viability | [95] |
| HCT116 cells | PAT: 5–25 μM | 24 h | FDA assay, Western blot analysis | Decrease cell viability Induce cell apoptosis | [96] |
| HCT116 cells | PAT: 1.25–20 μM | 1–4 days, 24 h | MTT assay, Western blot analysis | Decrease cell viability Induce apoptotic cells death | [97] |
| Citrinin | |||||
| HCT116 cells | CTN: 150 μM | 24 h | MTT assay | Decrease cell viability | [98] |
Table 4.
Modulation of tight junctions (physical barrier) induced by mycotoxins.
| Model | Dose/Administration Route | Exposure Time | Technique | Damage | References |
|---|---|---|---|---|---|
| Aflatoxin | |||||
| Caco-2 cells | AFM1: 0.12, 12 μM | 48 h | Western blot analysis, Immunofluorescent staining | Decrease in TEER value Increase in permeability of LY and 4 and 40 kDa FITC-dextran Decrease the protein expression of ZO-1, occludin, claudin-4, and claudin-3 Affect the distribution pattern of ZO-1, occludin, claudin-4, and claudin-3 | [54] |
| Caco-2/HT29-MTX cells | AFM1: 12 μM | 48 h | Transmission electron micrographs | Affect the distribution pattern of ZO-1, occludin, claudin-4, and claudin-3 disrupt TJs structure | [57] |
| Rat (Wistar, n = 35) | AFB1: 2.5 mg/kg Intraperitoneal administration | 7 days | Histopathological analysis | Villi degeneration of duodenum and ileum | [99] |
| Mice (Balb/c, n = 60) | AFB1: 100 μg/kg b.w. AFM1: 100 μg/kg b.w. Oral administration | 14 days | RT-PCR, Western blot analysis | Induce small intestine apoptosis | [100] |
| Broiler chicks (Ross 708, n = 288) | AFB1: 1.5 mg/kg Contaminated feed | 20 days | Serum biochemistry, RT-PCR | Increase in serum lactulose/rhamnose ratio Increase in transcript level of claudin-1 in jejunum | [101] |
| Broiler (Cobb, n = 576) | AFB1: 40 μg/kg Contaminated feed | 21 days | Serum biochemistry, RT-PCR | Increase in serum diamine oxidase concentration Decrease in mRNA level of occludin and claudin-1 in jejunum | [102] |
| Duck (Cherry Valley, n = 640) | AFB1: 195.4 μg/kg Contaminated feed | 35 days | Intestinal morphology analysis | Increase in crypt depth, villus width of duodenum Increase in villus height, villus width of jejunum | [103] |
| Broiler chicks (Ross 308, n = 336) | AFs 0.5 and 2 mg/kg feed Contaminated feed | 28, 42 days | Intestinal morphology analysis | Decrease in villi height to crypt depth ratio | [104] |
| Ochratoxin A | |||||
| Caco-2 cells | OTA: 0.12, 12 μM | 48 h | Western blot analysis, Immunofluorescent staining | Decrease in TEER value Increase in permeability of LY and 4 and 40 kDa FITC-dextran Decrease in protein expression of ZO-1, occluding, claudin-4, and claudin-3 Affect the distribution pattern of ZO-1, occluding, claudin-4, and claudin-3 | [54] |
| Caco-2 cells | OTA: 5–45 μM | 3, 12, 24 h | TEER measurement | Decrease in TEER value | [105] |
| Caco-2 cells | OTA: 15 μM | 5 h | Transmission electron microscope, Immunofluorescent staining | Reduce the microvilli on cell surface Alter the localization and distribution of claudin-1 and ZO-1 | [82] |
| IPEC-J2 cells | OTA: 4–128 μM | 6, 12, 24 h | Measurement of epithelial monolayer paracellular permeability | Decrease in TEER value Increase in permeability of 4 kDa FITC-dextran | [106] |
| Broiler chickens (n = 80) | OTA: 50 μg/kg b.w. Oral administration | 21 days | Intestinal morphology analysis | Decrease in villi height to crypt depth in duodenum, jejunum, and ileum | [107] |
| Deoxynivalenol | |||||
| Caco-2 cells | DON: 1–30 μM | 24, 40, 48 h | Western blot analysis, Immunofluorescent staining | Decrease in TEER value Increase in permeability of 4 kDa FITC-dextran Decrease in protein expression of claudin-7, occludin and E-cadherin Alter the localization and distribution of occludin and E-cadherin | [108] |
| IPEC-J2 cells | DON: 250, 500 ng/mL | 0.5–120 h | TEER measurement | Decrease in TEER value | [109] |
| IPEC-J2 cells | DON: 0.5–16 μM | 6, 12, 24 h | Western blot analysis, Immunofluorescent staining | Decrease in TEER value Increase in permeability of 4 kDa FITC-dextran Decrease in protein expression of claudin-3, and claudin-4 Alter the localization and distribution of claudin-3, and claudin-4 | [106] |
| Mouse enteroids, Mice (C57BL/6, n = 72) | DON: 250 ng/mL, DON: 2 mg/kg b.w. Oral administration | 72 h, 1–12 days | Immunofluorescent staining, Western blot analysis | Alter the localization and distribution of claudin-1 Decrease in protein expression of claudin-1, and ZO-1 in jejunum | [110] |
| Piglets jejunal explants | DON: 5, 10 μM | 4 h | histological analysis | Induce histological lesions on the intestine | [72] |
| Mice (C57BL/6, n = 72) | DON: 2 mg/kg b.w. Oral administration | 14 days | Serum biochemistry, Intestinal morphology analysis | Increase in serum diamine oxidase activity Decrease in villus/crypt ratio | [111] |
| Rat (Wistar, n = 32) | DON: 8.2 mg/kg feed Contaminated feed | 28 days | Histological and morphometric assessment, Immunohistochemical assessment | Decrease in crypt depth in jejunum, Decrease in the expression of occludin and E-cadherin in jejunum | [108] |
| Pig (piglet, n = 24) | DON: 50 μg/kg b.w. Contaminated feed | 15 days | Disaccharidases activity | Decrease in maltase, sucrase and lactase activity of the small intestine (duodenum, proximal and medium jejunum and ileum) | [112] |
| Fish (juvenile grass carp, n = 1440) | DON: 27–1515 μg/kg diet Contaminated feed | 60 days | RT-PCR | Decrease in the mRNA levels of ZO-1, ZO-2b, occludin, claudin-c, -f, -7a, -7b, -11 in fish intestine, Increase in the mRNA levels of claudin-12, -15a in fish intestine | [113] |
| Broiler chickens (n = 40) | DON: 10 mg/kg feed Contaminated feed | 35 days | Thiobarbituric acid reactive substance estimation | Increase in thiobarbituric acid reactive substance level, an indicator of oxidative stress, in jejunum | [114] |
| Pig (piglet, n = 20) | DON: 2 mg/kg feed Contaminated feed | 28 days | Histological assessment | Increase in the lesional score in intestine | [115] |
| Pig (piglet, n = 48) | DON: 1000–3000 μg/kg feed Contaminated feed | 21 days | Histological assessment | Decrease in villi height/crypt depth ratio in jejunum | [116] |
| Pig (piglet, n = 12) | DON: 2.3 mg/kg feed Contaminated feed | 20 days | Histological analysis | Decrease in the histological score in the jejunum | [72] |
| Broiler (Ross, n = 75) | DON: 1.7, 12.2 mg/kg feed Contaminated feed | 35 days | Histological analysis | Decrease in relative density (weight: length) of the small intestine | [117] |
| Zearalenone | |||||
| IPEC-1 cells | ZEN: 25, 50 μM α-ZOL: 25, 50 μM β-ZOL: 25, 50 μM | 1–10 days | TEER measurement | ZEN: no effect in TEER value α-ZOL: decrease in TEER value β-ZOL: decrease in TEER value | [118] |
| Rat (Sprague-Dawley, n = 96) | ZEN: 1.3–146.0 mg/kg Contaminated feed | 7 days | RT-PCR | Decrease in the mRNA expression of claudin-4 and occludin in jejunum | [119] |
| Pig (gilt, n = 40) | ZEN: 0.5–1.5 mg/kg Contaminated feed | 10 days | RT-PCR, Western blot analysis | Increase in the expression of oxidative stress related proteins | [120] |
| Fumonisin B1 | |||||
| IPEC-J2 cells | FB1: 50, 100 μM HFB1: 50, 100 μM | 1-9 days | TEER measurement | FB1: decrease in TEER value HFB1: decrease in TEER value | [42] |
| Pig (piglet, n = 24) | FB1: 6 mg/kg Contaminated feed | 35 days | Western blot analysis | Decrease in the protein expression of occludin in ileum | [121] |
| Patulin | |||||
| Caco-2 cells | PAT: 3–50 μM | 24 h | Western blot analysis | Decrease in TEER value Decrease in protein expression of ZO-1 | [94] |
| Caco-2 cells | PAT: 5–100 μM | 24 h | TEER measurement | Decrease in TEER value | [95] |
| Caco-2 cells | PAT: 50 μM | 3–72 h | Western blot analysis, Immunofluorescent staining | Decrease in TEER value Alter the localization and distribution of claudin-4, occludin and ZO-1 Decrease in protein expression of ZO-1 | [122] |
| Pigs jejunal explants | PAT: 10–100 μM | 4 h | Histological and morphometric analysis | Induce apical villi necrosis and alter lateral intercellular disruption | [123] |
| T-2 toxin | |||||
| Caco-2 cells | T-2: 50–100 ng/mL | 24 h | Western blot analysis | Decrease in TEER value Decrease in the expression of occludin | [124] |
| Turkey poults (n = 24) | T-2: 241–982 ppb Contaminated feed | 32 days | Histological analysis | Decrease in villi height in the jejunum | [125] |
| Mice (BALB/c, n = 30) | T-2: 0.5, 2.0 mg/kg b.w. Oral administration | 28 days | Histological analysis | Decrease in villi height in the ileum | [124] |
Table 5.
Modulation of the intestinal chemical barrier induced by mycotoxins.
| Model | Dose/Administration Route | Exposure Time | Technique | Damage | References |
|---|---|---|---|---|---|
| Aflatoxin | |||||
| Cao-2/HT29-MTX cells | AFM1: 0.05, 4 μg/mL | 48 h | RT-PCR, ELISA | Change the mRNA and protein expression level of MUC2, MUC5AC and MUC 5B in different proportions of co-cultured cells | [43] |
| Cao-2/HT29-MTX cells | AFM1: 12 μM | 48 h | RT-PCR, ELISA | No effect on the mRNA and protein expression level of MUC2, MUC5AC and MUC 5B | [57] |
| Broiler chicks (Ross 308, n = 336) | AFs (AFB1+AFB2+AFG1+AFG2) 0.5 and 2 mg/kg feed Contaminated feed | 28, 42 days | Histological analysis | Increase in the goblet cell counts at 28 and 42 d | [104] |
| Broiler chicks (Ross 308, n = 336) | AFs (AFB1+AFB2+AFG1+AFG2) 0.5 and 2 ppm feed Contaminated feed | 28, 42 days | Histological analysis | Increase in the goblet cell counts at 28 and 42 d | [132] |
| Hens (Hyline W36, n = 64) | AFB1: 0.5–2.0 mg/kg | 14 days | Histological analysis | No changes in goblet cell number and crude mucin production | [133] |
| Ochratoxin A | |||||
| Cao-2/HT29-MTX cells | OTA: 0.05, 4 μg/mL | 48 h | RT-PCR, ELISA | Modulate the mRNA level of MUC2, MUC5AC and MUC 5B, Increase in the protein expression of MUC2 and MUC5B at low concentration, while decrease at high concentration | [43] |
| Broiler chickens (n = 80) | OTA: 50 μg/kg b.w. Oral administration | 21 days | Histological analysis | Decrease in goblet cells number in the small intestine | [107] |
| Deoxynivalenol | |||||
| HT29-16E cells | DON: 0.1–100 μM | 3–48 h | RT-PCR | Decrease in the transcript level of MUC1, MUC2 and MUC3 | [134] |
| Cao-2 cells | DON: 2 μM | 5 min–24 h | RT-PCR | Increase in the transcript level of MUC5AC | [135] |
| Cao-2/HT29-MTX cells | DON: 2 μM | 48 h | RT-PCR, ELISA | Decrease in the transcript level of MUC5AC and MUC5B in the 90:10 ratio | [136] |
| IPEC-J2 cells | DON: 2 μM | 48 h | RT-PCR, ELISA | Increase in the transcript level of β-defensin1 and β-defensin2, while no effect on protein expression | [137] |
| porcine intestinal explants | DON: 10 μM | 8, 12 h | RT-PCR | Decrease in the transcript level of MUC1, MUC2 and MUC3 | [134] |
| Fish (juvenile grass carp, n = 1440) | DON: 318–1515 μg/kg diet Contaminated feed | 60 days | RT-PCR | Decrease in the mRNA expression of MUC2 and AMPs (β-defensin1, hepcidin, LEAP-2A and LEAP-2B) in proximal, middle and distal intestine | [138] |
| Broiler Chickens (Ross 308, n = 112) | DON: 4.6 mg/kg feed Contaminated feed | 15 days | RT-PCR | Decrease in the mRNA expression of MUC2 in duodenum | [139] |
| Mice (BALB/c, n = 42) | DON: 3.0 mg/kg Gavage | 15 days | RT-PCR | Decrease in the mRNA expression of MUC2 | [140] |
| Mice (C57BL/6, n = 72) | DON: 2 mg/kg b.w. Oral administration | 1–12 days | Immunohistochemistry staining | Decrease in the MUC2+ cells and LYZ+ cells number in jejunum | [110] |
| Mice (C57BL/6, n = 72) | DON: 2 mg/kg b.w. Oral administration | 14 days | Immunohistochemistry staining | Decrease in the MUC2+ cells and LYZ+ cells number in jejunum | [111] |
| Pig (piglet, n = 24) | DON: 3.0 mg/kg Contaminated feed | 35 days | Histological analysis | Decrease in the goblet cells number in jejunum and ileum | [121] |
| Pig (piglet, n = 20) | DON: 1.5 mg/kg Contaminated feed | 28 days | Histological analysis | Decrease in the number of goblet cells in jejunum and ileum | [115] |
| Mice (BALB/c, n = 42) | DON: 3.0 mg/kg Gavage | 15 days | Histological analysis | Decrease in the goblet cells number | [140] |
| Pig (piglet, n = 24) | DON: 50 μg/kg b.w. Contaminated feed | 15 days | Histological analysis | Decrease in the goblet cells number in villi, but no effects at crypts level | [112] |
| Pig (growing pigs, n = 24) | DON: 3, 6, 12 mg/kg feed Contaminated feed | 21 days | Histological analysis | Decrease in the goblet cells number of jejunum of pigs fed with diets 6 mg/kg DON-contaminated | [141] |
| Pig (gilt, n = 72) | DON: 12 μg/kg b.w. Contaminated feed | 7–42 days | Histological analysis | No effect on the goblet cells number of duodenum | [142] |
| Zearalenone | |||||
| Cao-2 cells | ZEN: 40 μM | 5 min–24 h | RT-PCR | Decrease in the mRNA expression of MUC5AC | [135] |
| Cao-2/HT29-MTX cells | ZEN: 40 μM | 48 h | RT-PCR, ELISA | Decrease in the transcript level of MUC5AC | [136] |
| IPEC-J2 cells | ZEN: 40 μM | 48 h | RT-PCR, ELISA | Increase in the transcript level of β-defensin1 and β-defensin2, while no effect on protein expression | [137] |
| Pig (gilt, n = 72) | ZEN: 40 μg/kg b.w. Contaminated feed | 7–42 days | Histological analysis | No effect on the goblet cells number of duodenum | [142] |
| Fumonisin B1 | |||||
| IPEC-J2 cells | FB1: 40 μM | 48 h | RT-PCR, ELISA | Increase in the mRNA expression of β-defensin1 and β-defensin2, while no effect on protein expression | [137] |
| Pig (piglet, n = 24) | DON: 6.0 mg/kg Contaminated feed | 35 days | Histological analysis | Inductive effect on goblet cell in jejunum | [121] |
| T-2 toxin | |||||
| HT-29 cells Cao-2 cells | T-2: 50–100 ng/mL | 24 h | Immunofluorescence staining, Western blot analysis | Decrease in the mucus layer in Caco-2 cells and HT-29 cells Decrease the protein expression of MUC2 | [124] |
| Chickens (n = 20) | T-2: 145 μg/kg diet Contaminated feed | 14 days | RT-PCR | Increase MUC2 mRNA expression level in jejunum | [143] |
| Nivalenol | |||||
| Cao-2/HT29-MTX cells | NIV: 2 μM | 48 h | RT-PCR, ELISA | Decrease in the mRNA expression of MUC5AC and MUC5B in the 90:10 ratio | [136] |
| IPEC-J2 cells | NIV: 2 μM | 48 h | RT-PCR, ELISA | Increase in the transcript level of β-defensin1 and β-defensin2, while no effect on protein expression | [137] |
Table 6.
Modulation of the intestinal immunological barrier induced by mycotoxins.
| Model | Dose/Administration Route | Exposure Time | Technique | Damage | References |
|---|---|---|---|---|---|
| Aflatoxin | |||||
| Mice (Balb/c, n = 60) | AFB1: 100 μg/kg b.w. AFM1: 100 μg/kg b.w. Oral administration | 14 days | Western blot analysis | AFB1: Decrease the protein level of TNF-α AFM1: Decrease the protein level of TNF-α | [100] |
| Pig (piglet, n = 24) | AFB1: 320 ppb Contaminated feed | 30 days | ELISA | Decrease in the protein expression of IL-1β, IL-6, IFN-γ, TNF-α and in duodenum | [147] |
| Broiler (Cobb, n = 576) | AFB1: 40 μg/kg Contaminated feed | 21 days | RT-PCR | Decrease in the transcript level of sIgA | [102] |
| Broiler chicks (Ross 308, n = 336) | AFs (AFB1+AFB2+AFG1+AFG2) 0.5 and 2 mg/kg feed Contaminated feed | 28, 42 days | Histological analysis | Increase in the number and diameter of lamina propria lymphoid follicles in jejunum | [104] |
| Shrimp (Litopenaeus vannamei, n = 100) | AFB1: 5 ppm Contaminated feed | 30 days | RNA-Seq | Identify 7 process or pathways related to immune system | [148] |
| Broiler chickens (Cobb 500, n = 240) | AFB1: 400 ppb Contaminated feed | 21 days | Immunohistochemistry | Decrease in the CD4+ cells number in jejunum | [149] |
| Ochratoxin A | |||||
| Caco-2 cells | OTA: 5–45 μM | 3, 12, 24 h | RT-PCR | Decrease in the mRNA expression of COX-2 and 5-LOX | [105] |
| Broiler chickens (n = 80) | OTA: 50 μg/kg b.w. Oral administration | 21 days | RT-PCR | Increase in the transcript level of TNF-α and IL-1β in small intestine | [107] |
| Duck (White Pekin ducklings, n = 540) | OTA: 2 mg/kg Contaminated feed | 21 days | ELISA | Increase in the protein level of TNF-α and IL-1β in jejunum | [150] |
| Deoxynivalenol | |||||
| IPEC-J2 cells | DON: 4 μM | 24 h | RT-PCR | Increase in the mRNA expression of TNF-α and IL-8 | [106] |
| IPEC-J2 cells | DON: 0.5–2.0 μg/mL | 4, 8, 12 h | RT-PCR | Increase in the mRNA expression IL-1β, IL-6, COX-2, and TNF-α | [151] |
| IPEC-J2 cells | DON: 4 μM | 12 h | Luminex multiplex assay | Increase in IL-6 and IL-8 protein level | [152] |
| Intestine 407 cells | DON: 25–1000 ng/mL | 12 h | Luciferase assay, RT-PCR | Increase in IL-8 secretion and mRNA expression | [153] |
| IPEC-1 cells | DON: 1000 ng/mL | 1 h | RT-PCR | Increase in IL-8 and MCP-1 mRNA expression | [154] |
| Pig (gilt, n = 72) | DON: 12 μg/kg b.w. Contaminated feed | 7-42 days | Histological analysis | Increase in the lymphocytes number in intestine | [142] |
| Pig (growing pigs, n = 24) | DON: 3, 6, 12 mg/kg feed Contaminated feed | 21 days | Histological analysis | Increase in the lymphocytes number in intestine | [141] |
| Fish (juvenile grass carp, n = 1440) | DON: 318–1515 μg/kg diet Contaminated feed | 60 days | RT-PCR | Increase in the transcript level of pro-inflammatory cytokines (IL-1β, IL-6, IL-8, IL-12p35, IL-12p40, IL-15, IL-17D, TNF-α and IFN-γ) in intestine Decrease in the mRNA expression of anti-inflammatory cytokines (IL-10, IL-11, IL-4/13A, IL-4/13B and TGF-β1) in intestine | [138] |
| Pig (piglet, n = 24) | DON: 3.0 mg/kg Contaminated feed | 35 days | RT-PCR | Increase the mRNA expression of IL-1β, IL-2, IL-6, IL-12p40, and MIP-1β in jejunum Increase the mRNA expression of IL-1β, IL-6, and TNF-α in ileum | [121] |
| Pig (piglet, n = 16) | DON: 3.5 mg/kg Contaminated feed | 42 days | RT-PCR | Increase the mRNA expression of IL-4 and CXCL10 in jejunum Increase the transcript level of IFN-γ, IL-6, IL-8 and CXCL10 in ileum | [155] |
| Zearalenone | |||||
| IPEC-1 cells | ZEN: 25 μM | 1 h | RT-PCR | No effect on the transcript level IL-1β, IL-6, IL-8, IL-17, and TNF-α Decrease in transcript level of IL-4 and IFN-γ | [156] |
| IPEC-1 cells | ZEN: 10–100 μM α-ZOL: 10–100 μM β-ZOL: 10–100 μM | 24 h | ELISA | ZEN: a tendency to increase the secretion of IL-8 and IL-10 α-ZOL: decrease the secretion of IL-8 and IL-10 β-ZOL: decrease the secretion of IL-8 and IL-10 | [118] |
| IPEC-1 cells | ZEN: 10 μM | 24 h | RT-PCR | A tendency to increase the transcript level of IL-10, IL-18, CCL20 and MCP-1 | [91] |
| IPEC-J2 cells | ZEN: 6, 8 μg/mL | 24 h | RT-PCR | Increase in the transcript level of pro-IL-1β and pro-IL-18 | [157] |
| Rat (Sprague-Dawley, n = 96) | ZEN: 1.3–146.0 mg/kg Contaminated feed | 7 days | RT-PCR | Decrease in the mRNA expression of TNF-α and IL-1β in pregnant dams | [119] |
| Fumonisin B1 | |||||
| HT-29 cells | FB1: 1.1–69.0 μM | 48 h | Immunoenzymatic bioassay | No changes on IL-8 secretion | [93] |
| IPEC-1 cells | FB1: 2.6–100 μM | 4 days | RT-PCR, ELISA | Decrease in the mRNA and protein expression of IL-8 | [158] |
| Pig (piglet, n = 24) | FB1: 6 mg/kg Contaminated feed | 35 days | RT-PCR | Increase the transcript level of IL-10 and IFN-γ in jejunum Increase the mRNA expression of TNF-α and IL-1β in ileum | [121] |
| Pig (n = 17) | FB1: 0.5 mg/kg b.w./d Gavage | 7 days | RT-PCR | Decrease in the mRNA expression of IL-8 in the ileum | [158] |
| Citrinin | |||||
| Mice (BALB/c, n = 20) | CTN: 1, 5, and 10 mg/kg b.w. Gavage | 14 days | Staining of immune cells for flow cytometric analysis | Increase CD8+ cells in intra-epithelial, Decrease CD19+ cells in intra-epithelial | [159] |
Table 7.
Modulation of the intestinal microbial barrier induced by mycotoxins.
| Model | Dose/Administration Route | Exposure Time | Technique | Damage | References |
|---|---|---|---|---|---|
| Aflatoxin | |||||
| Broiler chicks (Ross 308, n = 336) | AFs (AFB1+AFB2+AFG1+AFG2) 0.5 and 2 mg/kg feed Contaminated feed | 28, 42 days | Bacteriological examinations | Increase in the total negative bacteria, including Escherichia coli, Salmonella, Klebsiella in ileum | [104] |
| Broiler chicks (Ross 308, n = 336) | AFs (AFB1+AFB2+AFG1+AFG2) 0.5 and 2 ppm feed Contaminated feed | 28, 42 days | Bacterial examinations | Increase in the total negative bacteria, including Escherichia coli, Salmonella, Klebsiella in ileum | [132] |
| Broiler (Cobb, n = 576) | AFB1: 40 μg/kg Contaminated feed | 21 days | Bacterial examinations | No effect in the ileal bacteria populations, including Lactobacilli, Bifidobacteria, C. perfringens, Escherichia coli | [102] |
| Shrimp (Litopenaeus vannamei, n = 100) | AFB1: 5 ppm Contaminated feed | 30 days | 16S rRNA gene sequencing | Decrease in the types of intestinal microbiota Increase in the relative abundance of Proteobacteria and Firmicutes Decrease in the relative abundance of Bacteroidetes | [128] |
| Rat (Fischer 344, n = 20) | AFB1: 5, 25, 75 μg/kg b.w. Gavage | 28 days | 16S rRNA gene sequencing | Decrease in the fecal microbial diversity but increase evenness of community composition Decrease in the lactic acid bacteria number | [163] |
| Mice (Kunming, n = 24) | AFB1: 2.5, 4, 10 mg/L Gavage | 60 days | 16S rRNA gene sequencing | Decrease in the intestinal microbial diversity | [164] |
| Broiler Chickens (Cobb-Vantress, n = 480) | AFB1: 1, 1.5, 2 ppm Contaminated feed | 21 days | Bacti flat bottom plate assay | Increase in the number of total gram-negative bacteria, total aerobic bacteria number, and total lactic acid bacteria in cecum | [165] |
| Ochratoxin A | |||||
| Caco-2 cells | OTA: 1–100 μM | 12 h | Bacterial translocation assay | Increase in Escherichia coli translocation across monolayer | [166] |
| human intestinal microbial ecosystem | OTA: 2.5 μM | 14 days | PCR-TTGE and PCR-DGGE | Alter gut microbiota diversity and cause the loss of beneficial species Lactobacillus reuteri | [167] |
| Rat (F344, n = 18) | OTA: 70, 210 μg/kg b.w. Gavage | 28 days | 16S rRNA sequencing, shotgun sequencing | Decrease in the diversity of the gut microbiota Increase in the relative abundance of Lactobacillus | [168] |
| Deoxynivalenol | |||||
| IPEC-J2 cells | DON: 4 μM | 12 h | Bacterial translocation assay | Increase in Escherichia coli translocation across monolayer | [152] |
| IPEC-1 cells | DON: 5–50 μM | 48 h | Bacterial translocation assay | Increase in Escherichia coli translocation across monolayer | [169] |
| Caco-2 cells | DON: 1-100 μM | 12 h | Bacterial translocation assay | Increase in Escherichia coli translocation across monolayer | [166] |
| Mice (BALB/c, n = 42) | DON: 3.0 mg/kg Gavage | 15 days | 16S rRNA gene sequencing | Increase in the abundance of Clostridiales, Lachnospiraceae and Blautia | [140] |
| Mice (CD-1, n = 36) | DON: 1.0, 5.0 mg/kg Gavage | 14 days | shotgun sequencing | Increase in the abundance of Firmicutes and Bacteroidetes in cecum | [170] |
| Broiler chickens (ROSS 308, n = 80) | DON: 2.5, 5 and 10 mg/kg diet Contaminated feed | 35 days | 16S rRNA gene sequencing | Increase in the abundance of Firmicutes in cecum Decrease in the abundance of Proteobacteria in cecum | [171] |
| Pig (piglet, n = 24) | DON: 2.5 mg/kg Contaminated feed | 28 days | Capillary electrophoresis single-stranded conformation polymorphism | Increase in fecal aerobic mesophilic bacteria number | [172] |
| Rat (Wistar, n = 18) | DON: 60, 120 μg/kg b.w. Gavage | 40 days | 16S rRNA gene sequencing | Increase in the relative abundance of Coprococcus genus | [173] |
| Rat (Sprague-Dawley, n = 20) | DON: 100 μg/kg b.w. Gavage | 28 days | RT-PCR | Increase in the concentration of Bacteroides and Prevotella genera Decrease in the expression Escherichia coli | [174] |
| Rat (Wistar, N = 80) | DON: 2, 10 mg/kg Contaminated feed | 28 days | 16S rRNA gene sequencing | No effect on the composition of the gut microbiota | [175] |
| Zearalenone | |||||
| Mice (BALB/c, n = 40) | ZEN: 10 mg/kg b.w. Gavage | 14 days | 16S rRNA gene sequencing | Decrease in the abundance of Firmicutes Decrease in the abundance of Bacteroidetes | [176] |
| Pig (gilt, n = 75) | ZEN: 40 μg/kg b.w. Oral administration | 42 days | EcoPlate tests | Decrease in mesophilic aerobic bacteria number Decrease in the level of Enterobacteriaceae family | [177] |
| Fumonisin B1 | |||||
| Pig (piglet, n = 24) | FBs (FB1+FB2): 11.8 ppm Contaminated feed | 63 days | Capillary single-stranded conformation polymorphism analysis | Alter the digestive microbiota balance | [178] |
| Patulin | |||||
| Caco-2 cells | PAT: 1–100 μM | 12 h | Bacterial translocation assay | Increase in Escherichia coli translocation across monolayer | [166] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, Y.; Meng, L.; Liu, H.; Wang, J.; Zheng, N. The Compromised Intestinal Barrier Induced by Mycotoxins. Toxins 2020, 12, 619. https://doi.org/10.3390/toxins12100619
AMA Style
Gao Y, Meng L, Liu H, Wang J, Zheng N. The Compromised Intestinal Barrier Induced by Mycotoxins. Toxins. 2020; 12(10):619. https://doi.org/10.3390/toxins12100619
Chicago/Turabian StyleGao, Yanan, Lu Meng, Huimin Liu, Jiaqi Wang, and Nan Zheng. 2020. "The Compromised Intestinal Barrier Induced by Mycotoxins" Toxins 12, no. 10: 619. https://doi.org/10.3390/toxins12100619
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.