Insecticidal Activity of Bacillus thuringiensis Proteins against Coleopteran Pests

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. The Crystal Coleopteran-Active Proteins
2.1. Protein Structure
2.1.1. The 3-Domain Group Toxins
2.1.2. Non-3-domain Cry Toxins
2.1.3. Cyt Proteins
2.2. Insecticidal Activity
2.2.1. Host Range
2.2.2. Genetically Engineered Cry Genes
2.3. Mode of Action
2.3.1. Solubilization and Proteolytic Processing
2.3.2. Binding to the Larval Epithelium
2.3.3. Oligomerization and Pore Formation
3. The Secretable Coleopteran-Active Proteins
3.1. Protein Structure
3.2. Insecticidal Activity
3.3. Mode of Action
4. Bt Based Insecticides
5. Bt-Crops
6. Resistance and Cross-Resistance
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Raymond, B.; Johnston, P.R.; Nielsen-LeRoux, C.; Lereclus, D.; Crickmore, N. Bacillus thuringiensis: An impotent pathogen? Trends Microbiol. 2010, 18, 189–194. [Google Scholar] [CrossRef]
- Schnepf, E.; Crickmore, N.; Van Rie, J.; Lereclus, D.; Baum, J.; Feitelson, J.; Zeigler, D.R.; Dean, D.H. Bacillus thuringiensis and its pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 775–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P. Bacillus thuringiensis Toxins: An Overview of Their Biocidal Activity. Toxins 2014, 6, 3296–3325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nester, E.W.; Thomashow, L.S.; Metz, M.; Gordon, M. 100 Years of Bacillus thuringiensis: A critical scientific assessment. Am. Soc. Microbiol. 2002. [Google Scholar]
- Portela-Dussán, D.D.; Chaparro-Giralddo, A.; López-Pazos, S.A. La biotecnología de Bacillus thuringiensis en la agricultura. Nova 2013, 11, 87–96. [Google Scholar] [CrossRef]
- Krishnan, V.; Domanska, B.; Elhigazi, A.; Afolabi, F.; West, M.J.; Crickmore, N. The human cancer cell active toxin Cry41Aa from Bacillus thuringiensis acts like its insecticidal counterparts. Biochem. J. 2017, 474, 1591–1602. [Google Scholar] [CrossRef]
- Bravo, A.; Likitvivatanavong, S.; Gill, S.S.; Soberón, M. Bacillus thuringiensis: A story of a successful bioinsecticide. Insect Biochem. Mol. Biol. 2011, 41, 423–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, C. Global Status of Commercialized Biotech/GM Crops in 2017: Biotech Crop Adoption Surges as Economic Benefits Accumulate in 22 Years; ISAAA Br. No. 53; ISAAA: Ithaca, NY, USA, 2017. [Google Scholar]
- Schnepf, H.E.; Whiteley, H.R. Cloning and expression of the Bacillus thuringiensis crystal protein gene in Escherichia coli. Proc. Natl. Acad. Sci. USA 1981, 78, 2893–2897. [Google Scholar] [CrossRef] [Green Version]
- Crickmore, N.; Zeigler, D.R.; Feitelson, J.; Schnepf, E.; Van Rie, J.; Lereclus, D.; Baum, J.; Dean, D.H. Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiol. Mol. Biol. Rev. 1998, 62, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Crickmore, N.; Baum, J.A.; Bravo, A.; Lereclus, D.; Narva, K.E.; Sampson, K.; Schnepf, H.E.; Sun, M.; Zeigler, D.R. Bacillus thuringiensis Toxin Nomenclature. Available online: http://www.btnomenclature.info/ (accessed on 30 May 2020).
- Bravo, A.; Soberón, M.; Gill, S.S. Bacillus thuringiensis: Mechanisms and Use. In Insect Control: Biological and Synthetic Agents; Gilbert, L.I., Gill, S.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 175–205. [Google Scholar]
- Berry, C.; Crickmore, N. Structural classification of insecticidal proteins—Towards an in silico characterisation of novel toxins. J. Invertebr. Pathol. 2017, 142, 16–22. [Google Scholar] [CrossRef] [PubMed]
- de Maagd, R.A.; Bravo, A.; Crickmore, N. How Bacillus thuringiensis has evolved specific toxins to colonize the insect world. Trends Genet. 2001, 17, 193–199. [Google Scholar] [CrossRef]
- Pardo-López, L.; Soberón, M.; Bravo, A. Bacillus thuringiensis insecticidal three-domain Cry toxins: Mode of action, insect resistance and consequences for crop protection. FEMS Microbiol. Rev. 2013, 37, 3–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppert, B.; Ellis, R.T.; Babcock, J. Effects of Cry1F and Cry34Ab1/35Ab1 on storage pests. J. Stored Prod. Res. 2010, 46, 143–148. [Google Scholar] [CrossRef]
- Yu, Y.; Yuan, Y.; Gao, M. Construction of an environmental safe Bacillus thuringiensis engineered strain against Coleoptera. Appl. Microbiol. Biotechnol. 2016, 100, 4027–4034. [Google Scholar] [CrossRef]
- Jolivet, P. Food Habits and Food Selection of Chrysomelidae. Bionomic and Evolutionary Perspectives. In Biology of Chrysomelidae; Jolivet, P., Petitpierre, E., Hsiao, T.H., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 1–24. [Google Scholar]
- Soberón, M.; López-Díaz, J.A.; Bravo, A. Cyt toxins produced by Bacillus thuringiensis: A protein fold conserved in several pathogenic microorganisms. Peptides 2013, 41, 87–93. [Google Scholar] [CrossRef]
- Bietlot, H.P.; Vishnubhatla, I.; Carey, P.R.; Pozsgay, M.; Kaplan, H. Characterization of the cysteine residues and disulphide linkages in the protein crystal of Bacillus thuringiensis. Biochem. J. 1990, 267, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Agaisse, H.; Lereclus, D. How does Bacillus thuringiensis produce so much insecticidal crystal protein? J. Bacteriol. 1995, 177, 6027–6032. [Google Scholar] [CrossRef] [Green Version]
- Berry, C.; Ben-dov, E.; Jones, A.F.; Murphy, L.; Quail, M.A.; Holden, M.T.G.; Harris, D.; Zaritsky, A.; Parkhill, J. Complete sequence and organization of pBtoxis, the Toxin-Coding Plasmid of Bacillus thuringiensis subsp. israelensis. Appl. Environ. Microbiol. 2002, 68, 5082–5095. [Google Scholar] [CrossRef] [Green Version]
- de Maagd, R.A.; Bravo, A.; Berry, C.; Crickmore, N.; Schnepf, H.E. Structure, diversity, and evolution of protein toxins from spore-forming entomopathogenic bacteria. Annu. Rev. Genet. 2003, 37, 409–433. [Google Scholar] [CrossRef]
- Li, J.D.; Carroll, J.; Ellar, D.J. Crystal structure of insecticidal d-endotoxin from Bacillus thuringiensis at 2.5 A resolution. Nature 1991, 353, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Grochulski, P.; Masson, L.; Borisova, S.; Pusztai-Carey, M.; Schwartz, J.-L.; Brousseau, R.; Cygler, M. Bacillus thuringiensis CrylA(a) Insecticidal Toxin: Crystal structure and channel formation. J. Mol. Biol. 1995, 254, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Morse, R.J.; Yamamoto, T.; Stroud, R.M. Structure of Cry2Aa Suggests an Unexpected Receptor Binding Epitope. Structure 2001, 9, 409–417. [Google Scholar] [CrossRef]
- Galitsky, N.; Cody, V.; Wojtczak, A.; Ghosh, D.; Luft, J.R.; Pangborn, W.; English, L. Structure of the insecticidal bacterial δ-endotoxin Cry3Bb1 of Bacillus thuringiensis. Acta Crystallogr. Sect. D Biol. Crystallogr. 2001, 57, 1101–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonserm, P.; Davis, P.; Ellar, D.J.; Li, J. Crystal structure of the mosquito-larvicidal toxin Cry4Ba and its biological implications. J. Mol. Biol. 2005, 348, 363–382. [Google Scholar] [CrossRef] [PubMed]
- Boonserm, P.; Mo, M.; Angsuthanasombat, C.; Lescar, J. Structure of the functional form of the mosquito larvicidal Cry4Aa toxin from Bacillus thuringiensis at 2.8-Angstrom resolution. J. Bacteriol. 2006, 188, 3391–3401. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Ye, S.; Liu, Y.; Wei, L.; Xue, J.; Wu, H.; Song, F.; Zhang, J.; Wu, X.; Huang, D.; et al. Crystal structure of Bacillus thuringiensis Cry8Ea1: An insecticidal toxin toxic to underground pests, the larvae of Holotrichia parallela. J. Struct. Biol. 2009, 168, 259–266. [Google Scholar] [CrossRef]
- Ye, Y.; Godzik, A. FATCAT: A web server for flexible structure comparison and structure similarity searching. Nucleic Acids Res. 2004, 32, W582–W585. [Google Scholar] [CrossRef] [Green Version]
- Bravo, A.; Gill, S.S.; Soberón, M. Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon 2007, 49, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Wang, B.-C.; Yu, Z.; Sun, M. Structural Insights into Bacillus thuringiensis Cry, Cyt and Parasporin Toxins. Toxins 2014, 6, 2732–2770. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Sahara, K.; Bando, H.; Asano, S. Discovery of a novel Bacillus thuringiensis Cry8D protein and the unique toxicity of the Cry8D-class proteins against scarab beetles. J. Invertebr. Pathol. 2008, 99, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Bokori-Brown, M.; Savva, C.G.; Fernandes Da Costa, S.P.; Naylor, C.E.; Basak, A.K.; Titball, R.W. Molecular basis of toxicity of Clostridium perfringens epsilon toxin. FEBS J. 2011, 278, 4589–4601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, W.P.; Donovan, J.C.; Slaney, A.C. Bacillus thuringiensis CryET33 and CryET34 Compositions and Uses Therefor. U.S. Patent 6,063,756 A, 16 May 2000. [Google Scholar]
- Rydel, T.; Sharamitaro, J.; Brown, G.R.; Gouzov, V.; Seale, J.; Sturman, E.; Thoma, R.; Gruys, K.; Isaac, B. The crystal structure of a coleopteran insect-active binary Bt protein toxin complex at 2.5 Å resolution. In Proceedings of the Annual Meeting of the American Crystallographic Association, Los Angeles, CA, USA, 21–26 July 2001. [Google Scholar]
- Baum, J.A.; Chu, C.R.; Rupar, M.; Brown, G.R.; Donovan, W.P.; Huesing, J.E.; Ilagan, O.; Malvar, T.M.; Pleau, M.; Walters, M.; et al. Binary toxins from Bacillus thuringiensis active against the western corn rootworm, Diabrotica virgifera virgifera LeConte. Appl. Environ. Microbiol. 2004, 70, 4889–4898. [Google Scholar] [CrossRef] [Green Version]
- Ellis, R.T.; Stockhoff, B.A.; Stamp, L.; Schnepf, H.E.; Schwab, G.E.; Knuth, M.; Russell, J.; Cardineau, G.A.; Narva, K.E. Novel Bacillus thuringiensis binary insecticidal Crystal proteins active on Western Corn Rootworm, Diabrotica virgifera virgifera LeConte. Appl. Environ. Microbiol. 2002, 68, 1137–1145. [Google Scholar] [CrossRef] [Green Version]
- Kelker, M.S.; Berry, C.; Evans, S.L.; Pai, R.; McCaskill, D.G.; Wang, N.X.; Russell, J.C.; Baker, M.D.; Yang, C.; Pflugrath, J.W.; et al. Structural and Biophysical Characterization of Bacillus thuringiensis Insecticidal Proteins Cry34Ab1 and Cry35Ab1. PLoS ONE 2014, 9, e112555. [Google Scholar] [CrossRef]
- Cohen, S.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A.; Dym, O. Cyt1Aa Toxin: Crystal structure reveals implications for Its membrane-perforating function. J. Mol. Biol. 2011, 413, 804–814. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koni, P.A.; Ellar, D.J. Structure of the mosquitocidal δ-endotoxin CytB from Bacillus thuringiensis sp. kyushuensisand implications for membrane pore formation. J. Mol. Biol. 1996, 257, 129–152. [Google Scholar]
- Cohen, S.; Dym, O.; Albeck, S.; Ben-Dov, E.; Cahan, R.; Firer, M.; Zaritsky, A. High-Resolution crystal structure of activated Cyt2Ba monomer from Bacillus thuringiensis subsp. israelensis. J. Mol. Biol. 2008, 380, 820–827. [Google Scholar] [CrossRef]
- Gazit, E.; Burshtein, N.; Ellar, D.J.; Sawyer, T.; Shai, Y. Bacillus thuringiensis Cytolytic toxin associates specifically with Its synthetic Helices A and C in the membrane Bound State. Implications for the assembly of oligomeric transmembrane pores. Biochemistry 1997, 36, 15546–15554. [Google Scholar] [CrossRef]
- Promdonkoy, B.; Rungrod, A.; Promdonkoy, P.; Pathaichindachote, W.; Krittanai, C.; Panyim, S. Amino acid substitutions in αA and αC of Cyt2Aa2 alter hemolytic activity and mosquito-larvicidal specificity. J. Biotechnol. 2008, 133, 287–293. [Google Scholar] [CrossRef]
- Federici, B.A.; Park, H.-W.; Sakano, Y. Insecticidal Protein Crystals of Bacillus thuringiensis. In Inclusions in Prokaryotes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 195–236. [Google Scholar]
- Krieg, A.; Huger, A.M.; Langenbruch, G.A.; Schnetter, W. Bacillus thuringiensis var. tenebrionis: Ein neuer, gegenüber Larven von Coleopteren wirksamer Pathotyp. Zeitschrift für Angew. Entomol. 2009, 96, 500–508. [Google Scholar]
- Herrnstadt, C.; Soares, G.G.; Wilcox, E.R.; Edwards, D.L. A new strain of Bacillus thuringiensis with activity against coleopteran insects. Nat. Biotechnol. 1986, 4, 305–308. [Google Scholar]
- Barjac, H.; Frachon, E. Classification of Bacillus thuringiensis strains. Entomophaga 1990, 35, 233–240. [Google Scholar] [CrossRef]
- Rupar, M.J.; Donovan, W.P.; Groat, R.G.; Slaney, A.C.; Mattison, J.W.; Johnson, T.B.; Charles, J.F.; Dumanoir, V.C.; de Barjac, H. Two novel strains of Bacillus thuringiensis toxic to coleopterans. Appl. Environ. Microbiol. 1991, 57, 3337–3344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, B.; Höfte, H.; Annys, K.; Jansens, S.; Soetaert, P.; Peferoen, M. Novel Bacillus thuringiensis insecticidal crystal protein with a silent activity against coleopteran larvae. Appl. Environ. Microbiol. 1992, 58, 2536–2542. [Google Scholar]
- Whalon, M.E.; Miller, D.L.; Hollingworth, R.M.; Grafius, E.J.; Miller, J.R. Selection of a Colorado Potato Beetle (Coleoptera: Chrysomelidae) Strain Resistant to Bacillus thuringiensis. J. Econ. Entomol. 1993, 86, 226–233. [Google Scholar] [CrossRef]
- Bauer, L.S. Resistance: A threat to the insecticidal crystal proteins of Bacillus thuringiensis. Florida Entomol. 1995, 78, 414–443. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Keweshan, R.S.; Dunbar, M.W. Field-Evolved Resistance to Bt Maize by Western Corn Rootworm. PLoS ONE 2011, 6, e22629. [Google Scholar] [CrossRef] [Green Version]
- Ekobu, M.; Solera, M.; Kyamanywa, S.; Mwanga, R.O.M.; Odongo, B.; Ghislain, M.; Moar, W.J. Toxicity of seven Bacillus thuringiensis Cry proteins against Cylas puncticollis and Cylas brunneus (Coleoptera: Brentidae) using a novel artificial diet. J. Econ. Entomol. 2010, 103, 1493–1502. [Google Scholar] [CrossRef]
- Domínguez-Arrizabalaga, M.; Villanueva, M.; Fernandez, A.B.; Caballero, P. A strain of Bacillus thuringiensis containing a novel cry7Aa2 gene that Is toxic to Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae). Insects 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Song, P.; Wang, Q.; Nangong, Z.; Su, J.; Ge, D. Identification of Henosepilachna vigintioctomaculata (Coleoptera: Coccinellidae) midgut putative receptor for Bacillus thuringiensis insecticidal Cry7Ab3 toxin. J. Invertebr. Pathol. 2012, 109, 318–322. [Google Scholar] [CrossRef]
- Shu, C.; Yu, H.; Wang, R.; Fen, S.; Su, X.; Huang, D.; Zhang, J.; Song, F. Characterization of Two Novel cry8 genes from Bacillus thuringiensis Strain BT185. Curr. Microbiol. 2009, 58, 389–392. [Google Scholar] [CrossRef]
- Shu, C.; Yan, G.; Wang, R.; Zhang, J.; Feng, S.; Huang, D.; Song, F. Characterization of a novel cry8 gene specific to Melolonthidae pests: Holotrichia oblita and Holotrichia parallela. Appl. Microbiol. Biotechnol. 2009, 84, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Gindin, G.; Mendel, Z.; Levitin, B.; Kumar, P.; Levi, T.; Shahi, P.; Khasdan, V.; Weinthal, D.; Kuznetsova, T.; Einav, M.; et al. The basis for rootstock resilient to Capnodis species: Screening for genes encoding δ-endotoxins from Bacillus thuringiensis. Pest Manag. Sci. 2014, 70, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Hua, G.; Taylor, M.D.; Adang, M.J. A coleopteran cadherin fragment synergizes toxicity of Bacillus thuringiensis toxins Cry3Aa, Cry3Bb, and Cry8Ca against lesser mealworm, Alphitobius diaperinus (Coleoptera: Tenebrionidae). J. Invertebr. Pathol. 2014, 123, 1–5. [Google Scholar] [PubMed]
- Oliveira, G.R.; Silva, M.C.; Lucena, W.A.; Nakasu, E.Y.; Firmino, A.A.; Beneventi, M.A.; Souza, D.S.; Gomes, J.E.; de Souza, J.D.; Rigden, D.J.; et al. Improving Cry8Ka toxin activity towards the cotton boll weevil (Anthonomus grandis). BMC Biotechnol. 2011, 11, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Zhang, J.; Huang, D.; Gao, J.; Song, F. Characterization of Bacillus thuringiensis strain Bt185 toxic to the Asian Cockchafer: Holotrichia parallela. Curr. Microbiol. 2006, 53, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liu, R.; Shu, C.; Zhang, Q.; Zhao, S.; Shao, G.; Zhang, X.; Gao, J. Characterization of one novel cry8 gene from Bacillus thuringiensis strain Q52-7. World J. Microbiol. Biotechnol. 2014, 30, 3075–3080. [Google Scholar] [CrossRef] [PubMed]
- Thomson, M.; Knuth, M.; Cardineau, G. Bacillus thuringiensis Toxins with Improved Activity. U.S. Patent 5,874,288 A, 23 February 1999. [Google Scholar]
- Shrestha, G.; Reddy, G.V.P.; Jaronski, S.T. Field efficacy of Bacillus thuringiensis galleriae strain SDS-502 for the management of alfalfa weevil and its impact on Bathyplectes spp. parasitization rate. J. Invertebr. Pathol. 2018, 153, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Isaac, B.; Krieger, E.K.; Mettus, A.-M.L.; Moshiri, F.; Sivasupramanian, S. Polypeptide Compositions Toxic to Anthonomus Insects and Methods of Use. U.S. Patent 6,541,448 B2, 1 August 2002. [Google Scholar]
- Mettus, A.-M.L.; Baum, J.A. Polypeptide Compositions Toxic to Diabrotica Insects, Obteined from Bt; CryET70, and Methods of Use. Eur. Patent Application EP 1 129 198 B1, 11 May 2000. 29 September 1999. [Google Scholar]
- van Frankenhuyzen, K. Cross-order and cross-phylum activity of Bacillus thuringiensis pesticidal proteins. J. Invertebr. Pathol. 2013, 114, 76–85. [Google Scholar] [CrossRef]
- Bradley, D.; Harkey, M.A.; Kim, M.K.; Biever, K.D.; Bauer, L. The insecticidal CryIB Crystal protein of Bacillus thuringiensis ssp. thuringiensis has dual specificity to Coleopteran and Lepidopteran larvae. J. Invertebr. Pathol. 1995, 65, 162–173. [Google Scholar] [CrossRef]
- Federici, B.A.; Bauer, L.S. Cyt1Aa protein of Bacillus thuringiensis Is toxic to the Cottonwood Leaf Beetle, Chrysomela scripta, and suppresses high levels of resistance to Cry3Aa. Appl. Environ. Microbiol. 1998, 64, 4368–4371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naimov, S.; Weemen-Hendriks, M.; Dukiandjiev, S.; de Maagd, R. Bacillus thuringiensis Delta-Endotoxin Cry1 Hybrid Proteins with Increased Activity against the Colorado Potato Beetle. Appl. Environ. Microbiol. 2001, 67, 5328–5330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, É.S.; Monnerat, R.G.; Queiroz, P.R.; Dumas, V.F.; Braz, S.V.; de Souza Aguiar, R.W.; Gomes, A.C.M.M.; Sánchez, J.; Bravo, A.; Ribeiro, B.M. Midgut GPI-anchored proteins with alkaline phosphatase activity from the cotton boll weevil (Anthonomus grandis) are putative receptors for the Cry1B protein of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2010, 40, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-González, Á.; Porteous-Álvarez, A.J.; Del Val, M.; Casquero, P.A.; Escriche, B. Toxicity of five Cry proteins against the insect pest Acanthoscelides obtectus (Coleoptera: Chrisomelidae: Bruchinae). J. Invertebr. Pathol. 2020, 169. [Google Scholar] [CrossRef] [PubMed]
- Martins, É.S.; Praça, L.B.; Dumas, V.F.; Silva-Werneck, J.O.; Sone, E.H.; Waga, I.C.; Berry, C.; Monnerat, R.G. Characterization of Bacillus thuringiensis isolates toxic to cotton boll weevil (Anthonomus grandis). Biol. Control 2007, 40, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Asano, S. Identification of cry gene from Bacillus thuringiensis by PCR and isolation of unique insecticidal bacteria. Mem. Fac. Agric. 1996, 19, 529–563. [Google Scholar]
- de Souza Aguiar, R.W.; Martins, É.S.; Ribeiro, B.M.; Monnerat, R.G. Cry10Aa Protein is highly toxic to Anthonomus grandis Boheman (Coleoptera: Curculionidae), an important insect pest in brazilian cotton crop fields. Bt Res. 2012, 3, 20–28. [Google Scholar]
- Baum, J.A.; Sukuru, U.R.; Penn, S.R.; Meyer, S.E.; Subbarao, S.; Shi, X.; Flasinski, S.; Heck, G.R.; Brown, R.S.; Clark, T.L. Cotton Plants Expressing a Hemipteran-Active Bacillus thuringiensis Crystal Protein Impact the Development and Survival of Lygus hesperus (Hemiptera: Miridae) Nymphs. J. Econ. Entomol. 2012, 105, 616–624. [Google Scholar] [CrossRef]
- Bradfisch, G.A.; Muller-Cohn, J.; Narva, K.E.; Fu, J.; Thomson, M. Bacillus thuringiensis toxins and Genes for Controlling Coleopteran Pests. U.S. Patent 6,710,027 B2, 23 March 2004. [Google Scholar]
- Schnepf, H.E.; Lee, S.; Dojillo, J.; Burmeister, P.; Fencil, K.; Morera, L.; Nygaard, L.; Narva, K.E.; Wolt, J.D. Characterization of Cry34/Cry35 Binary Insecticidal Proteins from Diverse Bacillus thuringiensis Strain Collections. Appl. Environ. Microbiol. 2005, 71, 1765–1774. [Google Scholar] [CrossRef] [Green Version]
- Moellenbeck, D.J.; Peters, M.L.; Bing, J.W.; Rouse, J.R.; Higgins, L.S.; Sims, L.; Nevshemal, T.; Marshall, L.; Ellis, R.T.; Bystrak, P.G.; et al. Insecticidal proteins from Bacillus thuringiensis protect corn from corn rootworms. Nat. Biotechnol. 2001, 19, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Rupar, M.J.; Donovan, W.P.; Tan, Y.; Slaney, A.C. Bacillus thuringiensis CryET29 Compositions Toxic to Coleopteran Insects and Ctenocephalides spp. U.S. Patent 6,093,695, 25 July 2000. [Google Scholar]
- Weathersbee, A.A.; Lapointe, S.L.; Shatters, R.G. Activity of Bacillus thuringiensis isolates against Diaprepes abbreviatus (Coleoptera: Curculionidae). Florida Entomol. 2006, 89, 441–448. [Google Scholar] [CrossRef]
- Mahmoud, S.B.; Ramos, J.E.; Shatters, R.G.; Hall, D.G.; Lapointe, S.L.; Niedz, R.P.; Rougé, P.; Cave, R.D.; Borovsky, D. Expression of Bacillus thuringiensis cytolytic toxin (Cyt2Ca1) in citrus roots to control Diaprepes abbreviatus larvae. Pesticide Biochem. Physiol. 2017, 136, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Amico, V.; Podgwaite, J.D.; Duke, S. Biological activity of Bacillus thuringiensis and associated toxins against the Asian Longhorned Beetle (Coleoptera: Cerambycidae). J. Entomol. Sci. 2004, 39, 318–324. [Google Scholar] [CrossRef]
- Chen, J.; Dai, L.-Y.; Wang, X.-P.; Tian, Y.-C.; Lu, M.-Z. The cry3Aa gene of Bacillus thuringiensis Bt886 encodes a toxin against long-horned beetles. Appl. Microbiol. Biotechnol. 2005, 67, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Peña, G.; Miranda-Rios, J.; de la Riva, G.; Pardo-López, L.; Soberón, M.; Bravo, A. A Bacillus thuringiensis S-Layer Protein Involved in Toxicity against Epilachna varivestis (Coleoptera: Coccinellidae). Appl. Environ. Microbiol. 2006, 127, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Elgizawy, K.K.; Ashry, N.M. Efficiency of Bacillus thuringiensis strains and their Cry proteins against the Red Flour Beetle, Tribolium castaneum (Herbst.) (Coleoptera: Tenebrionidae). Egypt. J. Biol. Pest Control 2019, 29, 94. [Google Scholar] [CrossRef] [Green Version]
- MacIntosh, S.C.; Stone, T.B.; Sims, S.R.; Hunst, P.L.; Greenplate, J.T.; Marrone, P.G.; Perlak, F.J.; Fischhoff, D.A.; Fuchs, R.L. Specificity and efficacy of purified Bacillus thuringiensis proteins against agronomically important insects. J. Invertebr. Pathol. 1990, 56, 258–266. [Google Scholar] [CrossRef]
- Porcar, M.; García-Robles, I.; Domínguez-Escribà, L.; Latorre, A. Effects of Bacillus thuringiensis Cry1Ab and Cry3Aa endotoxins on predatory Coleoptera tested through artificial diet-incorporation bioassays. Bull. Entomol. Res. 2010, 100, 297–302. [Google Scholar] [CrossRef]
- Chang, X.; Lu, Z.; Shen, Z.; Peng, Y.; Ye, G. Bitrophic and Tritrophic Effects of Transgenic cry1Ab/cry2Aj Maize on the Beneficial, Nontarget Harmonia axyridis (Coleoptera: Coccinellidae). Environ. Entomol. 2017, 46, 1171–1176. [Google Scholar] [CrossRef]
- Sims, S.R. Host activity spectrum of the CryIIA Bacillus thuringiensis subsp. kurstaki protein. Effects on Lepidoptera, Diptera, and non-target arthropods. Southwest. Entomol. 1997, 22, 395–404. [Google Scholar]
- Sharma, A.; Kumar, S.; Bhatnagar, R.K. Bacillus thuringiensis Protein Cry6B (BGSC ID 4D8) is Toxic to Larvae of Hypera postica. Curr. Microbiol. 2011, 62, 597–605. [Google Scholar] [CrossRef]
- Niu, L.; Tian, Z.; Liu, H.; Zhou, H.; Ma, W.; Lei, C.; Chen, L. Transgenic Bt cotton expressing Cry1Ac/Cry2Ab or Cry1Ac/EPSPS does not affect the plant bug Adelphocoris suturalis or the pollinating beetle Haptoncus luteolus. Environ. Pollut. 2018, 234, 788–793. [Google Scholar] [CrossRef] [PubMed]
- Contreras, E.; Rausell, C.; Real, M.D. Proteome response of Tribolium castaneum larvae to Bacillus thuringiensis toxin producing strains. PLoS ONE 2013, 8, e55330. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhang, S.; Luo, J.-Y.; Wang, C.-Y.; Lv, L.-M.; Wang, X.-P.; Cui, J.-J.; Lei, C.-L. Bt proteins Cry1Ah and Cry2Ab do not affect cotton aphid Aphis gossypii and ladybeetle Propylea japonica. Sci. Rep. 2016, 6, 20368. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Ellar, D.J.; Bishop, A.; Johnson, C.; Lin, S.; Hart, E.R. Characterization of a Bacillus thuringiensis δ-Endotoxin which ia toxic to insects in three orders. J. Invertebr. Pathol. 2000, 76, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.E.C.; López-Pazos, S.A.; Cerón, J. Determination of Cry toxin activity and identification of an aminopeptidase N receptor-like gene in Asymmathetes vulcanorum (Coleoptera: Curculionidae). J. Invertebr. Pathol. 2012, 111, 94–98. [Google Scholar] [CrossRef] [PubMed]
- López-Pazos, S.A.; Rojas Arias, A.C.; Ospina, S.A.; Cerón, J. Activity of Bacillus thuringiensis hybrid protein against a lepidopteran and a coleopteran pest. FEMS Microbiol. Lett. 2010, 302, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, B.S.; Park, S.H.; Choi, S.K.; Koo, B.T.; Lee, S.T.; Kim, J.I. Distribution of cryV-type insecticidal protein genes in Bacillus thuringiensis and cloning of cryV-type genes from Bacillus thuringiensis subsp. kurstaki and Bacillus thuringiensis subsp. entomocidus. Appl. Environ. Microbiol. 1995, 61, 2402–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.-K.; Shin, B.-S.; Kong, E.-M.; Rho, H.M.; Park, S.-H. Cloning of a new Bacillus thuringiensis cry1I -Type crystal protein gene. Curr. Microbiol. 2000, 41, 65–69. [Google Scholar] [CrossRef]
- Kostichka, K.; Warren, G.W.; Mullins, M.; Mullins, A.D.; Palekar, N.V.; Craig, J.A.; Koziel, M.G.; Estruch, J.J. Cloning of a cryV-type insecticidal protein gene from Bacillus thuringiensis: The cryV-encoded protein is expressed early in stationary phase. J. Bacteriol. 1996, 178, 2141–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz de Escudero, I.; Estela, A.; Porcar, M.; Martínez, C.; Oguiza, J.A.; Escriche, B.; Ferrè, J.; Caballero, P. Molecular and Insecticidal Characterization of a Cry1I Protein Toxic to Insects of the Families Noctuidae, Tortricidae, Plutellidae, and Chrysomelidae. Appl. Environ. Microbiol. 2006, 72, 4796–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossi-De-Sa, M.F.; De Magalhaes, M.Q.; Silva, M.S.; Silva, S.M.B.; Dias, S.C.; Nakasu, E.Y.T.; Brunetta, P.S.F.; Oliveira, G.R.; De Oliveira Neto, O.B.; De Oliveira, R.S.; et al. Susceptibility of Anthonomus grandis (Cotton Boll Weevil) and Spodoptera frugiperda (Fall Armyworm) to a Cry1Ia-type toxin from a Brazilian Bacillus thuringiensis strain. BMB Rep. 2007, 40, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Gleave, A.P.; Williams, R.; Hedges, R.J. Screening by polymerase chain reaction of Bacillus thuringiensis serotypes for the presence of cryV-like insecticidal protein genes and characterization of a cryV gene cloned from B. thuringiensis subsp. kurstaki. Appl. Environ. Microbiol. 1993, 59, 1683–1687. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, R.; Behle, R.; Liu, R.; Niu, L.; Song, P.; Shakoori, A.R.; Jurat-Fuentes, J.L. Activity of Bacillus thuringiensis Cry1Ie2, Cry2Ac7, Vip3Aa11 and Cry7Ab3 proteins against Anticarsia gemmatalis, Chrysodeixis includens and Ceratoma trifurcata. J. Invertebr. Pathol. 2017, 150, 70–72. [Google Scholar] [CrossRef]
- Liu, J.; Song, F.; Zhang, J.; Liu, R.; He, K.; Tan, J.; Huang, D. Identification of vip3A-type genes from Bacillus thuringiensis strains and characterization of a novel vip3A-type gene. Lett. Appl. Microbiol. 2007, 45, 432–438. [Google Scholar] [CrossRef]
- von Tersch, M.A.; Gonzalez, J.M. Bacillus thuringiensis cryET1 Toxin Gene and Protein Toxic to Lepidopteran Insects. U.S. Patent 5,356,623 A, 18 October 1994. [Google Scholar]
- Oppert, B.; Morgan, T.D.; Kramer, K.J. Efficacy of Bacillus thuringiensis Cry3Aa protoxin and protease inhibitors against coleopteran storage pests. Pest Manag. Sci. 2011, 67, 568–573. [Google Scholar] [CrossRef]
- Zhongkamg, W.; He, W.; Peng, G.; Xia, Y. Transformation and expression of specific insecticide gene Bt cry3A in resident endogenetic bacteria isolated from Apriona germari (Hope) larvae intestines. Acta Microbiol. Sin. 2008, 48, 1168–1174. [Google Scholar]
- Kurt, A.; Özkan, M.; Sezen, K.; Demirbaǧ, Z.; Özcengiz, G. Cry3Aa11: A new Cry3Aa δ-endotoxin from a local isolate of Bacillus thuringiensis. Biotechnol. Lett. 2005, 27, 1117–1121. [Google Scholar] [CrossRef]
- Su, Z.; Deng, L.; Yi, X.; Xiao, S.; Zhang, C. The toxicity of Cry3Aa protein in Brontispa longissima by prokaryatic expression. Genomics Appl. Biol. 2009, 28, 691–694. [Google Scholar]
- Génissel, A.; Leplé, J.C.; Millet, N.; Augustin, S.; Jouanin, L.; Pilate, G. High tolerance against Chrysomela tremulae of transgenic poplar plants expressing a synthetic cry3Aa gene from Bacillus thuringiensis ssp tenebrionis. Mol. Breed. 2003, 11, 103–110. [Google Scholar] [CrossRef]
- James, R.R.; Croft, B.A.; Strauss, S.H. Susceptibility of the Cottonwood Leaf Beetle (Coleoptera: Chrysomelidae) to Different Strains and Transgenic Toxins of Bacillus thuringiensis. Environ. Entomol. 1999, 28, 108–115. [Google Scholar] [CrossRef]
- Yan, G.; Song, F.; Shu, C.; Liu, J.; Liu, C.; Huang, D.; Feng, S.; Zhang, J. An engineered Bacillus thuringiensis strain with insecticidal activity against Scarabaeidae (Anomala corpulenta) and Chrysomelidae (Leptinotarsa decemlineata and Colaphellus bowringi). Biotechnol. Lett. 2009, 31, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jurat-Fuentes, J.L.; Oppert, B.; Fabrick, J.A.; Liu, C.; Gao, J.; Lei, Z. Increased toxicity of Bacillus thuringiensis Cry3Aa against Crioceris quatuordecimpunctata, Phaedon brassicae and Colaphellus bowringi by a Tenebrio molitor cadherin fragment. Pest Manag. Sci. 2011, 67, 1076–1081. [Google Scholar] [CrossRef]
- Park, Y.; Abdullah, M.A.F.; Taylor, M.D.; Rahman, K.; Adang, M.J. Enhancement of Bacillus thuringiensis Cry3Aa and Cry3Bb Toxicities to Coleopteran Larvae by a Toxin-Binding Fragment of an Insect Cadherin. Appl. Environ. Microbiol. 2009, 75, 3086–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Olson, M.; Lin, G.; Hey, T.; Tan, S.Y.; Narva, K.E. Bacillus thuringiensis Cry34Ab1/Cry35Ab1 interactions with Western Corn Rootworm Midgut Membrane Binding Sites. PLoS ONE 2013, 8, e53079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carroll, J.S.; Li, J.; Ellar, D.J. Proteolytic processing of a coleopteran-specific delta-endotoxin produced by Bacillus thuringiensis var. tenebrionis. Biochem. J. 1989, 261, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhang, J.; Song, F.; Wu, J.; Feng, S.; Huang, D. Engineered Bacillus thuringiensis GO33A with broad insecticidal activity against lepidopteran and coleopteran pests. Appl. Microbiol. Biotechnol. 2006, 72, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Mahadeva Swamy, H.M.; Asokan, R.; Thimmegowda, G.G.; Mahmood, R. Expression of cry3A gene and its toxicity against Asian Gray Weevil Myllocerus undecimpustulatus undatus Marshall (Coleoptera: Curculionidae). J. Basic Microbiol. 2013, 53, 664–676. [Google Scholar] [CrossRef]
- Gomez, S.; Mateus, A.C.; Hernandez, J.; Zimmermann, B.H. Recombinant Cry3Aa has insecticidal activity against the Andean potato weevil, Premnotrypes vorax. Biochem. Biophys. Res. Commun. 2000, 279, 653–656. [Google Scholar] [CrossRef]
- Johnson, T.M.; Rishi, A.S.; Nayak, P.; Sen, S.K. Cloning of a cryIIIA endotoxin gene of Bacillus thuringiensis var. tenebrionis and its transient expression inindica rice. J. Biosci. 1996, 21, 673–685. [Google Scholar]
- Adang, M.J. Enhancement of Bacillus thuringiensis Cry Toxicities to Lesser Mealworm Alphitobius diaperinus. U.S. Patent 0,201,549 A1, 18 August 2011. [Google Scholar]
- Wu, S.-J.; Dean, D.H. Functional Significance of Loops in The Receptor Binding Domain of Bacillus thuringiensis CryIIIA δ-Endotoxin. J. Mol. Biol. 1996, 255, 628–640. [Google Scholar] [CrossRef] [PubMed]
- Fabrick, J.; Oppert, C.; Lorenzen, M.D.; Morris, K.; Oppert, B.; Jurat-Fuentes, J.L. A Novel Tenebrio molitor Cadherin Is a functional receptor for Bacillus thuringiensis Cry3Aa toxin. J. Biol. Chem. 2009, 284, 18401–18410. [Google Scholar] [CrossRef] [Green Version]
- Donovan, W.P.; Rupar, M.J.; Slaney, A.C.; Malvar, T.; Gawron-Burke, M.C.; Johnson, T.B. Characterization of two genes encoding Bacillus thuringiensis insecticidal crystal proteins toxic to Coleoptera species. Appl. Environ. Microbiol. 1992, 58, 3921–3927. [Google Scholar] [CrossRef] [Green Version]
- Adang, M.J.; Adang, M.J.; Abdullah, M.A.F. Enhancement of Bacilus thuringiensis Cry Protein Toxicities to Coleopterans, and Novel Insect Cadhetin Fragments. U.S. Patent 8,486,887 B2, 16 July 2013. [Google Scholar]
- Lambert, B.; Theunis, W.; Aguda, R.; Van Audenhove, K.; Decock, C.; Jansens, S.; Seurinck, J.; Peferoen, M. Nucleotide sequence of gene crylllD encoding a novel coleopteran-active crystal protein from strain BTII09P of Bacillus thuringiensis subsp. Kurstaki Gene 1992, 110, 131–132. [Google Scholar] [CrossRef]
- Haffani, Y.Z.; Cloutier, C.; Belzile, F.J. Bacillus thuringiensis cry3Ca1 protein is toxic to the Colorado Potato Beetle, Leptinotarsa decemlineata (Say). Biotechnol. Prog. 2001, 17, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Shu, C.; Lin, Y.; Song, F.; Zhang, J. Identification, cloning and expression for novel cry7Ab gene and its insecticidal activity. J. Agric. Biotechnol. 2009, 17, 908–913. [Google Scholar]
- Wang, L.; Guo, W.; Tan, J.; Sun, W.; Liu, T.; Sun, Y. Construction of an engineering strain expressing cry7Ab7 gene cloned from Bacillus thuringiensis. Front. Agric. China 2010, 4, 328–333. [Google Scholar] [CrossRef]
- Foncerrada, L.; Sick, A.J.; Payne, J.M. Novel Coleopteran-Active Bacillus thuringiensis Isolate and a Novel Gene Encoding a Coleopteran-Active Toxin. Patent Application EP0498537 A2, 16 January 1992. pp. 1–5. [Google Scholar]
- Michaels, T.E.; Narva, K.E.; Foncerrada, L. Bacillus thuringiensis Toxins Active against Scarab Pests. U.S. Patent 5,554,534 A, 10 September 1996. [Google Scholar]
- Zhang, Y.; Zheng, G.; Tan, J.; Li, C.; Cheng, L. Cloning and characterization of a novel cry8Ab1 gene from Bacillus thuringiensis strain B-JJX with specific toxicity to scarabaeid (Coleoptera: Scarabaeidae) larvae. Microbiol. Res. 2013, 168, 512–517. [Google Scholar] [CrossRef]
- Michaels, T.E.; Foncerrada, L.; Narva, K.E. Process for Controlling Scarab Pests with Bacillus thuringiensis Isolates. International Patent Application WO 93/15206, 5 August 1993. [Google Scholar]
- Abad, A.; Duck, N.B.; Feng, X.; Flannagan, R.D.; Kahn, T.W.; Sims, L.E. Genes encoding Novel Proteins with Pesticidal Activity against Coleopterans. International Patent Application WO 02/034774 A2, 2 May 2002. [Google Scholar]
- Liu, J.; Yan, G.; Shu, C.; Zhao, C.; Liu, C.; Song, F.; Zhou, L.; Ma, J.; Zhang, J.; Huang, D. Construction of a Bacillus thuringiensis engineered strain with high toxicity and broad pesticidal spectrum against coleopteran insects. Appl. Microbiol. Biotechnol. 2010, 87, 243–249. [Google Scholar] [CrossRef]
- Jia, Y.; Zhao, C.; Wang, Q.; Shu, C.; Feng, X.; Song, F.; Zhang, J. A genetically modified broad-spectrum strain of Bacillus thuringiensis toxic against Holotrichia parallela, Anomala corpulenta and Holotrichia oblita. World J. Microbiol. Biotechnol. 2014, 30, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Sato, R.; Takeuchi, K.; Ogiwara, K.; Minami, M.; Kaji, Y.; Suzuki, N.; Hori, H.; Asano, S.; Ohba, M.; Iwahana, H. Cloning, heterologous expression, and localization of a novel crystal protein gene from Bacillus thuringiensis serovar japonensis strain Buibui toxic to scarabaeid insects. Curr. Microbiol. 1994, 28, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.-F.; Zhang, J.; Song, F.-P.; Lang, Z.-H. Microbial control and biotechnology research on Bacillus thuringiensis in China. J. Invertebr. Pathol. 2007, 95, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Asano, S.; Yamashita, C.; Iizuka, T.; Takeuchi, K.; Yamanaka, S.; Cerf, D.; Yamamoto, T. A strain of Bacillus thuringiensis subsp. galleriae containing a novel cry8 gene highly toxic to Anomala cuprea (Coleoptera: Scarabaeidae). Biol. Control 2003, 28, 191–196. [Google Scholar] [CrossRef]
- Liu, X.; Cheng, L.; Li, T.; Li, C.; Li, G. Cloning, expression and insecticidal activity of cry8Ea2 gene from Bacillus thuringiensis strain B. Acta Agric. Boreali-Sin. 2008, 23, 1–4. [Google Scholar]
- Singaravelu, B.; Crickmore, N.; Srikanth, J.; Hari, K.; Sankaranarayanan, C. Prospecting for Scarabid Specific Bacillus thuringiensis Crystal toxin cry8 gene in sugarcane ecosystem of Tamil Nadu, India. J. Sugarcane Res. 2013, 3, 141–144. [Google Scholar]
- Silva-Werneck, J.O.; Ellar, D.J. Characterization of a novel Cry9Bb δ-endotoxin from Bacillus thuringiensis. J. Invertebr. Pathol. 2008, 98, 320–328. [Google Scholar] [CrossRef]
- Brown, K.L.; Whiteley, H.R. Molecular characterization of two novel crystal protein genes from Bacillus thuringiensis subsp. thompsoni. J. Bacteriol. 1992, 174, 549–557. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Hodgman, T.C.; Krieger, L.; Schnetter, W.; Schairer, H.U. Cloning and analysis of the first cry gene from Bacillus popilliae. J. Bacteriol. 1997, 179, 4336–4341. [Google Scholar] [CrossRef] [Green Version]
- Guzov, V.M.; Malvar, T.M.; Roberts, J.K.; Sivasupramanian, S. Insect Inhibitory Bacillus thuringiensis Proteins, Fusions, and Methods of Use Therefor. U.S. Patent 7,655,838 B2, 2 February 2010. [Google Scholar]
- Herman, R.A.; Scherer, P.N.; Young, D.L.; Mihaliak, C.A.; Woodsworth, A.T.; Stockhoff, B.A.; Narva, K.E. Binary Insecticidal Crystal Protein from Bacillus thuringiensis, Strain PS149B1: Effects of Individual Protein Components and Mixtures in Laboratory Bioassays. J. Econ. Entomol. 2002, 95, 635–639. [Google Scholar] [CrossRef]
- Rupar, M.J.; Donovan, W.P.; Chu, C.R.; Pease, E.; Tan, Y.; Slaney, A.C.; Malvar, T.M.; Baum, J. Coleopteran Toxin Polypeptide Compositions and Insect-Resistant Transgenic Plants. U.S. Patent 090,094,714 A1, 9 April 2009. [Google Scholar]
- Yokoyama, T.; Tanaka, M.; Hasegawa, M. Novel cry gene from Paenibacillus lentimorbus strain Semadara inhibits ingestion and promotes insecticidal activity in Anomala cuprea larvae. J. Invertebr. Pathol. 2004, 85, 25–32. [Google Scholar] [CrossRef] [PubMed]
- de Maagd, R.A.; Kwa, M.S.; van der Klei, H.; Yamamoto, T.; Schipper, B.; Vlak, J.M.; Stiekema, W.J.; Bosch, D. Domain III substitution in Bacillus thuringiensis delta-endotoxin CryIA(b) results in superior toxicity for Spodoptera exigua and altered membrane protein recognition. Appl. Environ. Microbiol. 1996, 62, 1537–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadenkov, A.A.; Kadyrov, R.M.; Uzbekova, S.V.; Kuz’min, E.V.; Osterman, A.L.; Chestukhina, G.G.; Shemiakin, M.F. Creation of a hybrid protein gene based on Bacillus thuringiensis delta endotoxins CryIIIA and CrIA(a) and expression of its derivatives in Escherichia coli. Mol. Biol. 1993, 27, 952–959. [Google Scholar]
- Walters, F.S.; DeFontes, C.M.; Hart, H.; Warren, G.W.; Chen, J.S. Lepidopteran-Active Variable-Region Sequence Imparts Coleopteran Activity in eCry3.1Ab, an Engineered Bacillus thuringiensis Hybrid Insecticidal Protein. Appl. Environ. Microbiol. 2010, 76, 3082–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- English, L.; Brussock, M.; Malvar, T.M.; Bryson, J.; Kulesza, C.; Walters, F.S.; Slatin, S.; von Tersch, M.A.; Romano, C. Nucleic Acid Segments Encoding Modified Bacillus thuringiensis Coleopteran-Toxic Crystal Proteins. U.S. Patent 6,060,594, 9 May 2000. [Google Scholar]
- Walters, F.S.; Stacy, C.M.; Lee, M.K.; Palekar, N.; Chen, J.S. An engineered Chymotrypsin/Cathepsin G site in Domain I renders Bacillus thuringiensis Cry3A active against Western Corn Rootworm larvae. Appl. Environ. Microbiol. 2008, 74, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Jouzani, G.S.; Valijanian, E.; Sharafi, R. Bacillus thuringiensis: A successful insecticide with new environmental features and tidings. Appl. Microbiol. Biotechnol. 2017, 101, 2691–2711. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Crickmore, N. Specificity determinants for Cry insecticidal proteins: Insights from their mode of action. J. Invertebr. Pathol. 2017, 142, 5–10. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Li, J.; Liu, Y.; Song, F.; Zhang, J. The role of DNA binding with the Cry8Ea1 toxin of Bacillus thuringiensis. FEMS Microbiol. Lett. 2011, 317, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Xia, L.; Sun, Y.; Ding, X.; Fu, Z.; Mo, X.; Zhang, H.; Yuan, Z. Identification of cry-Type Genes on 20-kb DNA Associated with Cry1 Crystal Proteins from Bacillus thuringiensis. Curr. Microbiol. 2005, 51, 53–58. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, X.; Sun, Y.; Li, W.; Xia, L.; Ding, X.; Yin, J.; Hu, S.; Yu, Z.; Tang, Y. Construction of gene library of 20 kb DNAs from parasporal Crystal in Bacillus thuringiensis strain 4.0718: Phylogenetic analysis and molecular docking. Curr. Microbiol. 2012, 64, 106–111. [Google Scholar] [CrossRef]
- Ai, B.; Li, J.; Feng, D.; Li, F.; Guo, S. The Elimination of DNA from the Cry Toxin-DNA Complex Is a Necessary Step in the Mode of Action of the Cry8 Toxin. PLoS ONE 2013, 8, e81335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koller, C.N.; Bauer, L.S.; Hollingworth, R.M. Characterization of the pH-mediated solubility of Bacillus thuringiensis var. san diego native δ-endotoxin crystals. Biochem. Biophys. Res. Commun. 1992, 184, 692–699. [Google Scholar] [CrossRef]
- Chougule, N.P.; Doyle, E.; Fitches, E.; Gatehouse, J.A. Biochemical characterization of midgut digestive proteases from Mamestra brassicae (cabbage moth; Lepidoptera: Noctuidae) and effect of soybean Kunitz inhibitor (SKTI) in feeding assays. J. Insect Physiol. 2008, 54, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Michaud, D.; Bernier-Vadnais, N.; Overney, S.; Yelle, S. Constitutive expression of digestive cysteine proteinase forms during development of the colorado potato beetle, Leptinotarsa decemlineata Say (Coleoptera: Chrysomelidae). Insect Biochem. Mol. Biol. 1995, 25, 1041–1048. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Sahara, K.; Bando, H.; Asano, S. Intramolecular proteolytic nicking and binding of Bacillus thuringiensis Cry8Da toxin in BBMVs of Japanese beetle. J. Invertebr. Pathol. 2010, 105, 243–247. [Google Scholar] [CrossRef]
- Rausell, C.; García-Robles, I.; Sánchez, J.; Muñoz-Garay, C.; Martínez-Ramírez, A.C.; Real, M.D.; Bravo, A. Role of toxin activation on binding and pore formation activity of the Bacillus thuringiensis Cry3 toxins in membranes of Leptinotarsa decemlineata (Say). Biochim. Biophys. Acta Biomembr. 2004, 1660, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Martínez, P.; Vera-Velasco, N.M.; Martínez-Solís, M.; Ghislain, M.; Ferré, J.; Escriche, B. Shared binding sites for the Bacillus thuringiensis proteins Cry3Bb, Cry3Ca, and Cry7Aa in the African Sweet Potato Pest Cylas puncticollis (Brentidae). Appl. Environ. Microbiol. 2014, 80, 7545–7550. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Campuzano, C.; Real, M.D.; Martínez-Ramírez, A.C.; Bravo, A.; Rausell, C. An ADAM metalloprotease is a Cry3Aa Bacillus thuringiensis toxin receptor. Biochem. Biophys. Res. Commun. 2007, 362, 437–442. [Google Scholar] [CrossRef]
- Zúñiga-Navarrete, F.; Gómez, I.; Peña, G.; Bravo, A.; Soberón, M. A Tenebrio molitor GPI-anchored alkaline phosphatase is involved in binding of Bacillus thuringiensis Cry3Aa to brush border membrane vesicles. Peptides 2013, 41, 81–86. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Waterfield, N.; Ffrench-Constant, R.H. Expressed sequence tags from Diabrotica virgifera virgifera midgut identify a coleopteran cadherin and a diversity of cathepsins. Insect Mol. Biol. 2005, 14, 137–143. [Google Scholar] [CrossRef]
- Sayed, A.; Nekl, E.R.; Siqueira, H.A.A.; Wang, H.-C.; Ffrench-Constant, R.H.; Bagley, M.; Siegfried, B.D. A novel cadherin-like gene from western corn rootworm, Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae), larval midgut tissue. Insect Mol. Biol. 2007, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras, E.; Schoppmeier, M.; Real, M.D.; Rausell, C. Sodium Solute Symporter and Cadherin proteins Act as Bacillus thuringiensis Cry3Ba toxin functional receptors in Tribolium castaneum. J. Biol. Chem. 2013, 288, 18013–18021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, C.; Tan, S.; Yin, J.; Soberón, M.; Bravo, A.; Liu, C.; Geng, L.; Song, F.; Li, K.; Zhang, J. Assembling of Holotrichia parallela (dark black chafer) midgut tissue transcriptome and identification of midgut proteins that bind to Cry8Ea toxin from Bacillus thuringiensis. Appl. Microbiol. Biotechnol. 2015, 99, 7209–7218. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Huang, Y.; Shu, C.; Soberón, M.; Bravo, A.; Liu, C.; Song, F.; Lai, J.; Zhang, J. Holotrichia oblita Midgut Proteins That Bind to Bacillus thuringiensis Cry8-Like Toxin and Assembly of the H. oblita Midgut Tissue Transcriptome. Appl. Environ. Microbiol. 2017, 83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Bando, H.; Asano, S. Identification of a Bacillus thuringiensis Cry8Da toxin-binding glucosidase from the adult Japanese beetle, Popillia japonica. J. Invertebr. Pathol. 2013, 113, 123–128. [Google Scholar] [CrossRef]
- Pérez, C.; Muñoz-Garay, C.; Portugal, L.C.; Sánchez, J.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis ssp. israelensis Cyt1Aa enhances activity of Cry11Aa toxin by facilitating the formation of a pre-pore oligomeric structure. Cell. Microbiol. 2007, 9, 2931–2937. [Google Scholar]
- Khorramnejad, A.; Domínguez-Arrizabalaga, M.; Caballero, P.; Escriche, B.; Bel, Y. Study of the Bacillus thuringiensis Cry1Ia protein oligomerization promoted by midgut Brush Border Membrane Vesicles of lepidopteran and coleopteran insects, or cultured insect cells. Toxins 2020, 12, 133. [Google Scholar] [CrossRef] [Green Version]
- Warren, G.W.; Koziel, M.G.; Mullins, M.A.; Kostichka, K.; Estruch, J.J. Novel Pesticidal Proteins and Strains. Patent Coop. Treaty WO 96/10083 A1, 4 April 1996. [Google Scholar]
- Donovan, W.P.; Engleman, J.T.; Donovan, J.C.; Baum, J.A.; Bunkers, G.J.; Chi, D.J.; Clinton, W.P.; English, L.; Heck, G.R.; Ilagan, O.M.; et al. Discovery and characterization of Sip1A: A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae. Appl. Microbiol. Biotechnol. 2006, 72, 713–719. [Google Scholar] [CrossRef]
- Sattar, S.; Maiti, M.K. Molecular Characterization of a Novel Vegetative Insecticidal Protein from Bacillus thuringiensis Effective Against Sap-Sucking Insect Pest. J. Microbiol. Biotechnol. 2011, 21, 937–946. [Google Scholar] [CrossRef] [Green Version]
- Estruch, J.J.; Warren, G.W.; Mullins, M.A.; Nye, G.J.; Craig, J.A.; Koziel, M.G. Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proc. Natl. Acad. Sci. USA 1996, 93, 5389–5394. [Google Scholar] [CrossRef] [Green Version]
- Sha, J.; Zhang, J.; Chi, B.; Liu, R.; Li, H.; Gao, J. sip1Ab gene from a native Bacillus thuringiensis strain QZL38 and its insecticidal activity against Colaphellus bowringi Baly. Biocontrol Sci. Technol. 2018, 28, 459–467. [Google Scholar] [CrossRef]
- Warren, G.W. Vegetative Insecticidal Proteins. In Advances in Insect Control; Taylor & Francis: Abingdon, UK, 1997; pp. 109–122. [Google Scholar]
- Han, S.; Craig, J.A.; Putnam, C.D.; Carozzi, N.B.; Tainer, J.A. Evolution and mechanism from structures of an ADP-ribosylating toxin and NAD complex. Nat. Struct. Biol. 1999, 6, 932–936. [Google Scholar]
- Shi, Y.; Ma, W.; Yuan, M.; Sun, F.; Pang, Y. Cloning of vip1/vip2 genes and expression of Vip1Ca/Vip2Ac proteins in Bacillus thuringiensis. World J. Microbiol. Biotechnol. 2007, 23, 501–507. [Google Scholar] [CrossRef]
- Barth, H.; Aktories, K.; Popoff, M.R.; Stiles, B.G. Binary bacterial toxins: Biochemistry, biology, and applications of common Clostridium and Bacillus proteins. Microbiol. Mol. Biol. Rev. 2004, 68, 373–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakroun, M.; Banyuls, N.; Bel, Y.; Escriche, B.; Ferré, J. Bacterial Vegetative Insecticidal Proteins (Vip) from Entomopathogenic Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 329–350. [Google Scholar] [CrossRef] [Green Version]
- Feitelson, J.S.; Schnepf, H.E.; Narva, K.E.; Stockhoff, B.A.; Schmeits, J.; Loewer, D.; Dullum, C.J.; Muller-Cohn, J.; Stamp, L. Pesticidal Toxins and Nucleotide Sequences Which Encode These Toxins. U.S. Patent 6,204,435 B1, 20 March 2001. [Google Scholar]
- Boets, A.; Arnaut, G.; Van Rie, J.; Damme, N. Toxins. U.S. Patent 7,919,609 B2, 5 April 2011. [Google Scholar]
- Bi, Y.; Zhang, Y.; Shu, C.; Crickmore, N.; Wang, Q.; Du, L.; Song, F.; Zhang, J. Genomic sequencing identifies novel Bacillus thuringiensis Vip1/Vip2 binary and Cry8 toxins that have high toxicity to Scarabaeoidea larvae. Appl. Microbiol. Biotechnol. 2015, 99, 753–760. [Google Scholar] [CrossRef]
- Leuber, M.; Orlik, F.; Schiffler, B.; Sickmann, A.; Benz, R. Vegetative Insecticidal Protein (Vip1Ac) of Bacillus thuringiensis HD201: Evidence for oligomer and channel formation. Biochemistry 2006, 45, 283–288. [Google Scholar] [CrossRef]
- Geng, J.; Jiang, J.; Shu, C.; Wang, Z.; Song, F.; Geng, L.; Duan, J.; Zhang, J. Bacillus thuringiensis Vip1 functions as a receptor of Vip2 toxin for binary insecticidal activity against Holotrichia parallela. Toxins 2019, 11, 440. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Xu, W.; Yuan, M.; Tang, M.; Chen, J.; Pang, Y. Expression of vip1/vip2 genes in Escherichia coli and Bacillus thuringiensis and the analysis of their signal peptides. J. Appl. Microbiol. 2004, 97, 757–765. [Google Scholar] [CrossRef]
- Yu, X.; Liu, T.; Liang, X.; Tang, C.; Zhu, J.; Wang, S.; Li, S.; Deng, Q.; Wang, L.; Zheng, A.; et al. Rapid detection of vip1-type genes from Bacillus cereus and characterization of a novel vip binary toxin gene. FEMS Microbiol. Lett. 2011, 325, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Baranek, J.; Kaznowski, A.; Konecka, E.; Naimov, S. Activity of vegetative insecticidal proteins Vip3Aa58 and Vip3Aa59 of Bacillus thuringiensis against lepidopteran pests. J. Invertebr. Pathol. 2015, 130, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Wraight, S.P.; Lacey, L.A.; Kabaluk, J.T.; Gottel, M.S. Potential for Microbial Biological Control of Coleopteran and Hemipteran Pests of Potato—Agriculture and Agri-Food Canada (AAFC). Fruit Veg. Cereal Sci. Biotechnol. 2009, 3, 25–38. [Google Scholar]
- Beveridge, N.; Elek, J.A. Insect and host-tree species influence the effectiveness of a Bacillus thuringiensis ssp. tenebrionis-based insecticide for controlling chrysomelid leaf beetles. Aust. J. Entomol. 2001, 40, 386–390. [Google Scholar] [CrossRef]
- Coyle, D.R.; McMillin, J.D.; Krause, S.C.; Hart, E.R. Laboratory and field evaluations of two Bacillus thuringiensis formulations, Novodor and Raven, for control of Cottonwood Leaf Beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2000, 93, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Way, M.O.; Wallace, R.G.; Harper, H.B.; Landry, C.L. Control of Rice Water Weevil with Novodor 3, 1999. Arthropod Manag. Tests 2000, 25, 133. [Google Scholar] [CrossRef]
- Baum, J.A.; Johnson, T.B. Bacillus thuringiensis: Natural and recombinant bioinsecticide products. In Methods in Biotechnology, Vol5: Biopesticides: Use and Deliver; Humana Press Inc.: Totowa, NJ, USA, 1999; pp. 189–209. [Google Scholar]
- Sanchis, V. From microbial sprays to insect-resistant transgenic plants: History of the biospesticide Bacillus thuringiensis. A review. Agron. Sustain. Dev. 2011, 31, 217–231. [Google Scholar] [CrossRef]
- Gawron-Burke, C.; Baum, J.A. Genetic Manipulation of Bacillus thuringiensis Insecticidal Crystal Protein Genes in Bacteria. In Genetic Engineering; Springer US: Boston, MA, USA, 1991; Volume 13, pp. 237–263. [Google Scholar]
- Baum, J.A.; Kakefuda, M.; Gawron-burke, C. Engineering Bacillus thuringiensis Bioinsecticides with an Indigenous Site-Specific Recombination System. Appl. Environ. Microbiol. 1996, 62, 4367–4373. [Google Scholar] [CrossRef] [Green Version]
- Perlak, F.; Stone, T.; Muskopf, Y.; Petersen, L.; Parker, G.; McPherson, S.; Wyman, J.; Love, S.; Reed, G.; Biever, D. Genetically improved potatoes: Protection from damage by Colorado potato beetles. Plant Mol. Biol. 1993, 22, 313–321. [Google Scholar] [CrossRef]
- Duncan, D.R.; Hammond, D.; Zalewski, J.; Cudnohufsky, J.; Kaniewski, W.; Thornton, M.; Bookout, J.T.; Lavrik, P.; Rogan, G.J.; Feldman-Riebe, J. Field performance of “Transgenic” potato, with resistance to Colorado Potato Beetle and Viruses. HortScience 2002, 37, 556E–557. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.E.; Kaniewski, W.K.; Lawson, E.C. Reduced Field Spread of Potato Leafroll Virus in Potatoes Transformed with the Potato Leafroll Virus Coat Protein Gene. Plant Dis. 1997, 81, 1447–1453. [Google Scholar] [CrossRef]
- Thornton, M. The Rise and Fall of NewLeaf Potatoes. NABC Rep. 2004, 15, 235–243. [Google Scholar]
- Vaughn, T.; Cavato, T.; Brar, G.; Coombe, T.; DeGooyer, T.; Ford, S.; Groth, M.; Howe, A.; Johnson, S.; Kolacz, K.; et al. A Method of controlling corn rootworm feeding using a Bacillus thuringiensis protein expressed in transgenic maize. Crop Sci. 2005, 45, 931–938. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Shrestha, R.B.; Jakka, S.R.K.; Dunbar, M.W.; Clifton, E.H.; Paolino, A.R.; Ingber, D.A.; French, B.W.; Masloski, K.E.; Dounda, J.W.; et al. Evidence of resistance to Cry34/35Ab1 Corn by Western corn rootworm (Coleoptera: Chrysomelidae): Root Injury in the Field and Larval Survival in Plant-Based Bioassays. J. Econ. Entomol. 2016, 109, 1872–1880. [Google Scholar] [CrossRef]
- Zukoff, S.N.; Ostlie, K.R.; Potter, B.; Meihls, L.N.; Zukoff, A.L.; French, L.; Ellersieck, M.R.; French, B.W.; Hibbard, B.E. Multiple assays indicate varying levels of cross resistance in Cry3Bb1-selected field populations of the western corn rootworm to mCry3A, eCry3.1Ab, and Cry34/35Ab1. J. Econ. Entomol. 2016, 109, 1387–1398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wangila, D.S.; Gassmann, A.J.; Petzold-Maxwell, J.L.; French, B.W.; Meinke, L.J. Susceptibility of Nebraska Western Corn Rootworm (Coleoptera: Chrysomelidae) populations to Bt corn events. J. Econ. Entomol. 2015, 108, 742–751. [Google Scholar] [CrossRef] [Green Version]
- Jakka, S.R.K.; Shrestha, R.B.; Gassmann, A.J. Broad-spectrum resistance to Bacillus thuringiensis toxins by western corn rootworm (Diabrotica virgifera virgifera). Sci. Rep. 2016, 6, 27860. [Google Scholar] [CrossRef]
- Shrestha, R.B.; Dunbar, M.W.; French, B.W.; Gassmann, A.J. Effects of field history on resistance to Bt maize by western corn rootworm, Diabrotica virgifera virgifera LeConte. PLoS ONE 2018, 13, e0200156. [Google Scholar] [CrossRef] [Green Version]
- Ferré, J.; Van Rie, J.; Macintosh, S.C. Insecticidal Genetically Modified Crops and Insect Resistance Management (IRM). In Integration of Insect-Resistant Genetically Modified Cropts within IPM Programs; Springer Science + Business Media: Dordrecht, The Netherlands, 2008. [Google Scholar]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing yramided transgenic Bt crops for sustainable pest management. Nat. Biotechnol. 2015, 33, 61–168. [Google Scholar] [CrossRef] [PubMed]
- Onstad, D.W.; Meinke, L.J. Modeling evolution of Diabrotica virgifera virgifera to transgenic corn with two insecticidal traits. J. Econ. Entomol. 2010, 103, 849–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]





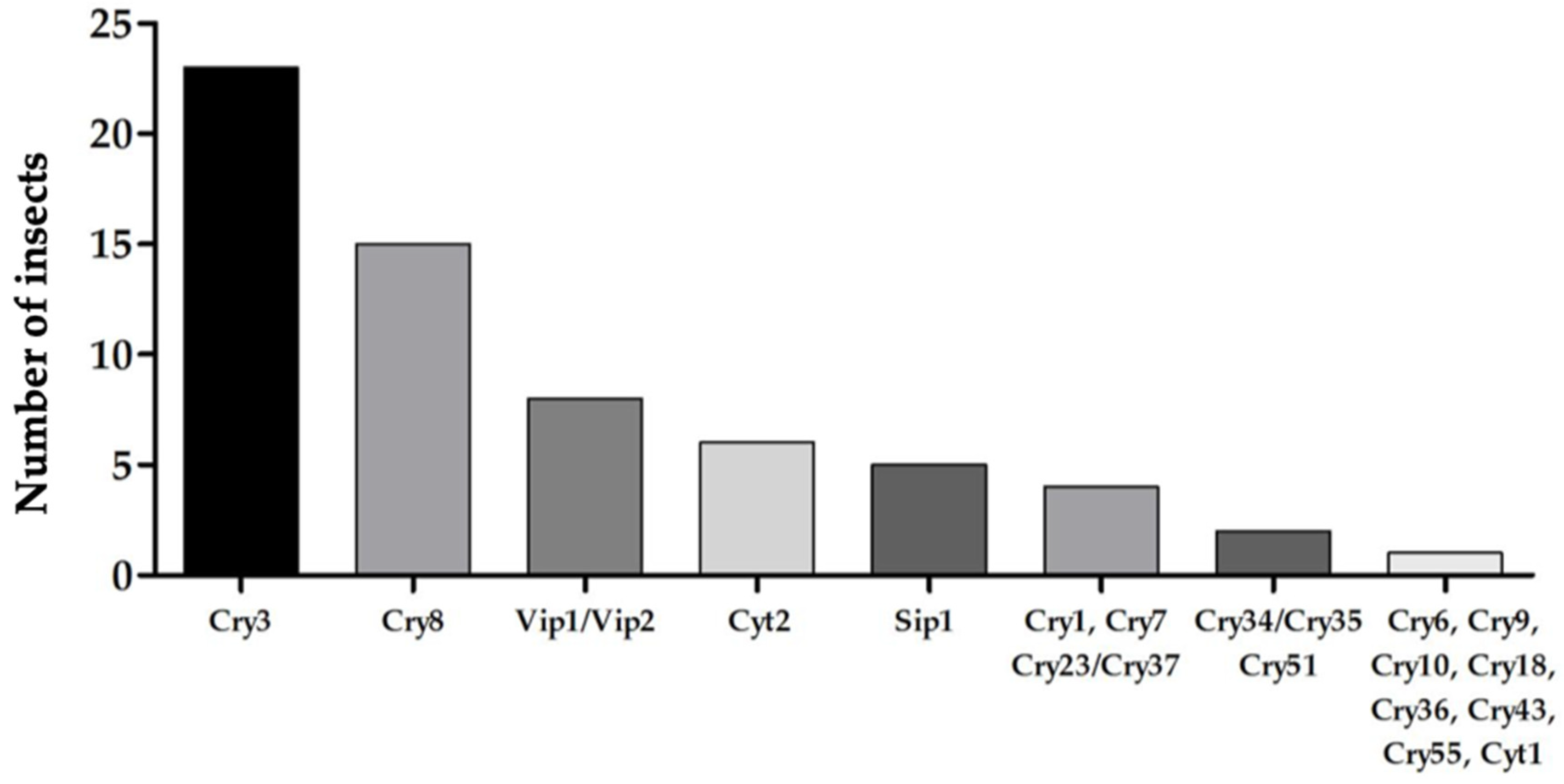{kind=link}
{kind=link}
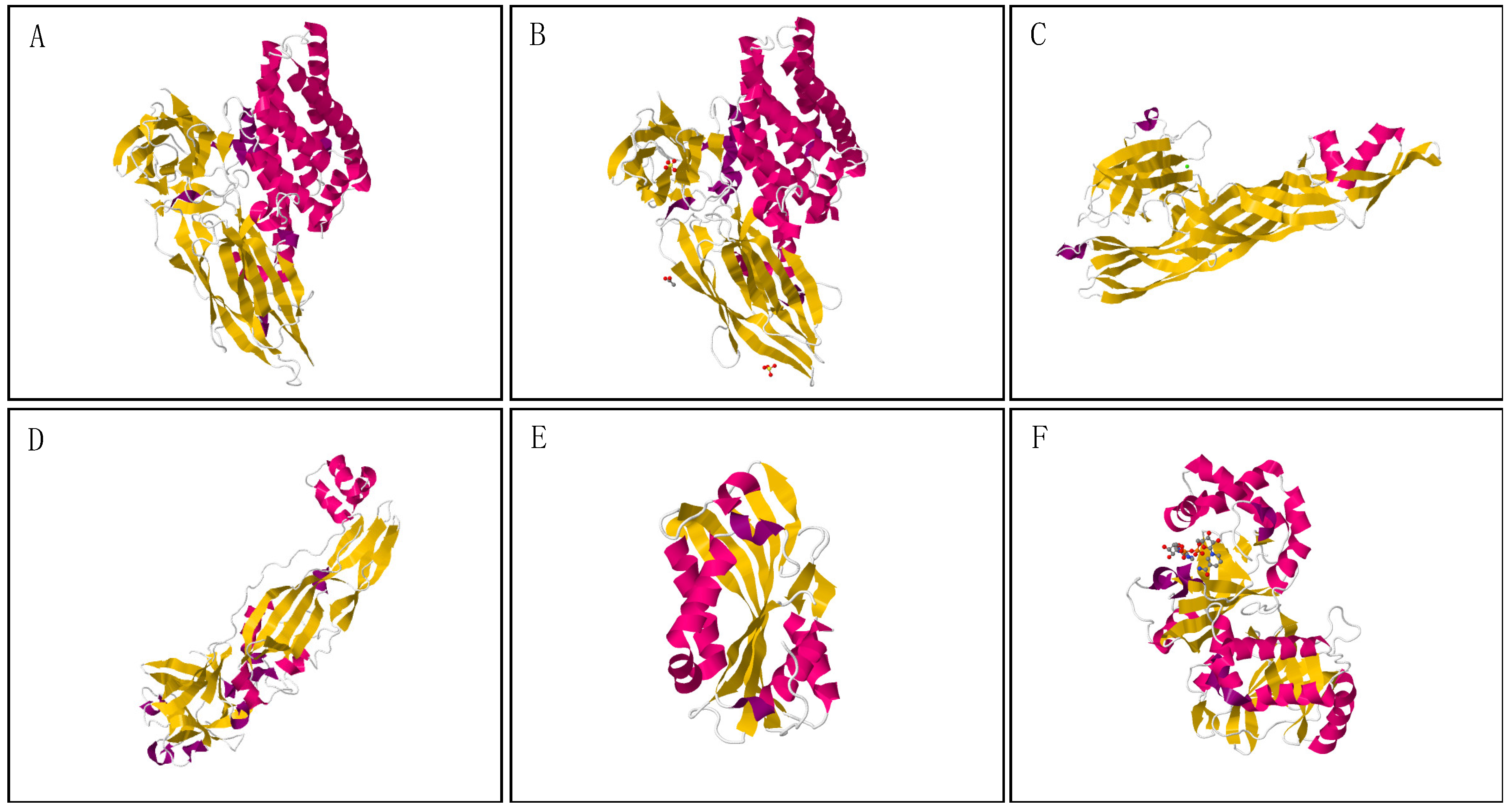{kind=link}
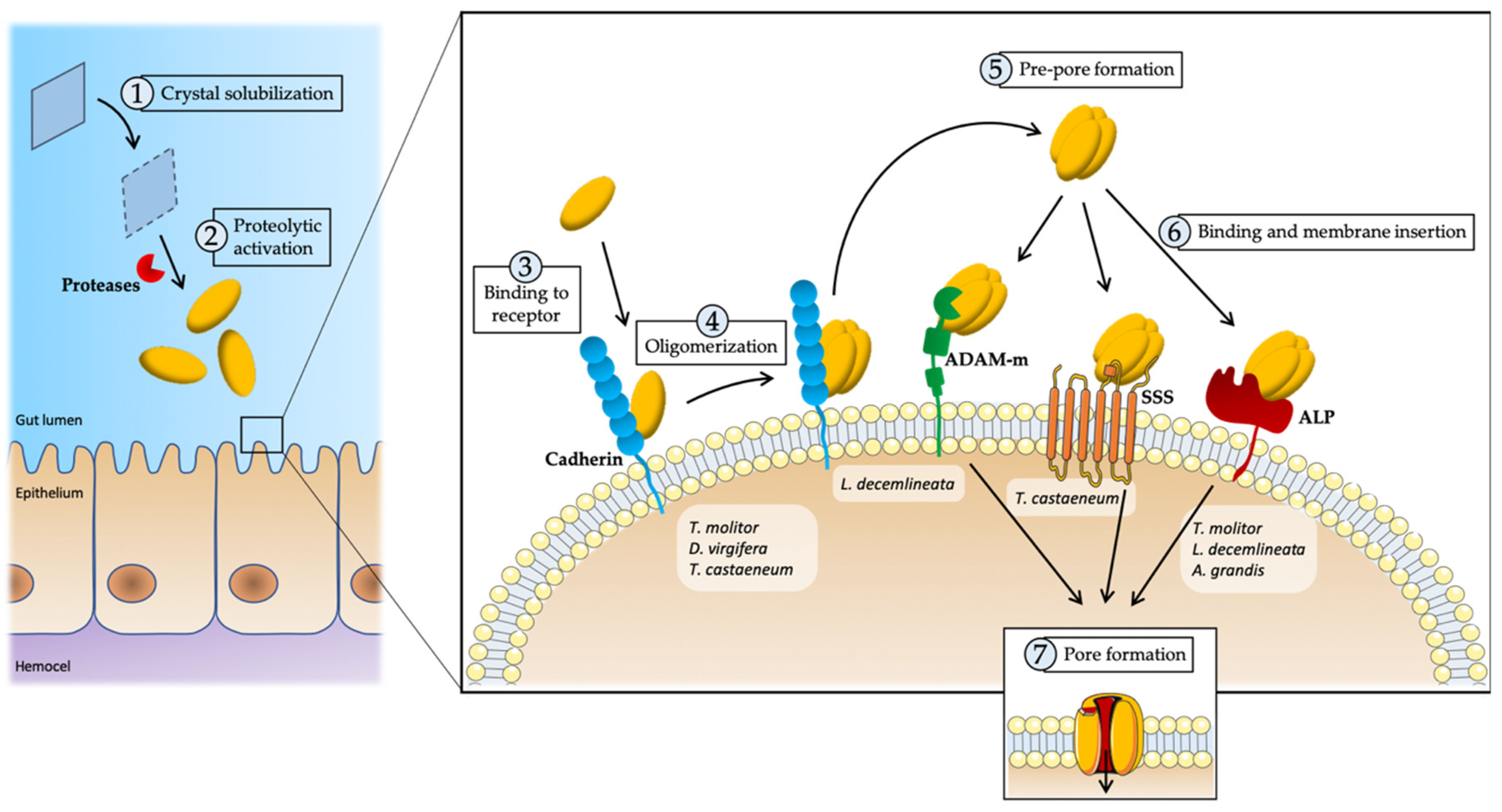{kind=link}
| Crystal Type Toxin | Target Insect | Activity (a) | LC50 (b) | Reference | |
|---|---|---|---|---|---|
| Scientific Name | Family | ||||
| Cry1Aa | Anoplophora glabripennis | Cerambycidae | N | [86] | |
| Apriona germari | Cerambycidae | N | [87] | ||
| Epilachna varivestis | Coccinellidae | A | [88] | ||
| Tribolium castaneum | Tenebrionidae | LA | [89] | ||
| Cry1Ab | Diabrotica undecimpuntata | Chrysomelidae | N | [90] | |
| Leptinotarsa decemlineata | Chrysomelidae | N | [90] | ||
| Phyllotreta armoraciae | Chrysomelidae | N | [90] | ||
| Adalia bipunctata | Coccinellidae | N | [91] | ||
| Atheta coriaria | Coccinellidae | N | [91] | ||
| Cryptolaemus montrouzieri | Coccinellidae | N | [91] | ||
| Harmonia axyridis | Coccinellidae | N | [92] | ||
| Anthonomus grandis | Curculionidae | N | [90] | ||
| Hypera postica | Curculionidae | N | [90] | ||
| Popillia japonica | Scarabaeidae | N | [90] | ||
| Cry1Ac | Diabrotica undecimpuntata | Chrysomelidae | N | [90] | |
| Leptinotarsa decemlineata | Chrysomelidae | N | [90] | ||
| Phyllotreta armoraciae | Chrysomelidae | N | [90] | ||
| Hippodamia convergens | Coccinellidae | N | [93] | ||
| Anthonomus grandis | Curculionidae | N | [90] | ||
| Hypera postica | Curculionidae | N | [90,94] | ||
| Haptoncus luteolus | Nitidulidae | N | [95] | ||
| Tribolium castaneum | Tenebrionidae | N | [96] | ||
| Popillia japonica | Scarabaeidae | N | [90] | ||
| Cry1Ah | Propylea japónica | Coccinellidae | N | [97] | |
| Cry1Aj | Harmonia axyridis | Coccinellidae | N | [92] | |
| Cry1Ba | Anoplophora glabripennis | Cerambycidae | N | [86] | |
| Acanthoscelides obtectus | Chrysomelidae | A | [75] | ||
| Chrysomela scripta F | Chrysomelidae | A | 1.8 // 5.9 | [71,72] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | 1050 // 142 | [71,73] | |
| Phaedon cochleariae | Chrysomelidae | N | [98] | ||
| Anthonomus grandis | Curculionidae | A | 305.32 | [74] | |
| Asymmathetes vulcanorum | Curculionidae | N | [99] | ||
| Hypothenemus hampei | Curculionidae | A | [100] | ||
| Tribolium castaneum | Tenebrionidae | N | [89] | ||
| Cry1Ca | Tribolium castaneum | Tenebrionidae | N | [89] | |
| Cry1Da | Tribolium castaneum | Tenebrionidae | N | [89] | |
| Cry1Ea | Tribolium castaneum | Tenebrionidae | N | [89] | |
| Cry1Fa | Cryptolestes pusillus | Laemophloeidae | N | [17] | |
| Tribolium castaneum | Tenebrionidae | N | [17] | ||
| Cry1Fb | Tribolium castaneum | Tenebrionidae | N | [89] | |
| Cry1Ia | Acanthoscelides obtectus | Chrysomelidae | A | [75] | |
| Agelastica coerulea | Chrysomelidae | N | [101,102] | ||
| Diabrotica undecimpuntata | Chrysomelidae | N | [103] | ||
| Leptinotarsa decemlineata | Chrysomelidae | A | 33.7 // 10 | [73,104] | |
| Phaedom brassicae | Chrysomelidae | N | [101] | ||
| Anthonomus grandis | Curculionidae | A | 21.5 // 230 | [76,105] | |
| Asymmathetes vulcanorum | Curculionidae | N | [99] | ||
| Tenebrio molitor | Tenebrionidae | N | [106] | ||
| Tribolium castaneum | Tenebrionidae | N | [89] | ||
| Cry1Ib | Phaedom brassicae | Chrysomelidae | N | [101] | |
| Agelastica coerulea | Chrysomelidae | N | [101] | ||
| Cry1Id | Agelastica coerulea | Chrysomelidae | N | [102] | |
| Cry1Ie | Ceratoma trifurcata | Chrysomelidae | N | [107] | |
| Pyrrhalta aenescens | Chrysomelidae | N | [108] | ||
| Cry1Jb | Diabrotica undecimpuntata | Chrysomelidae | N | [109] | |
| Leptinotarsa decemlineata | Chrysomelidae | N | [109] | ||
| Cry2Aa | Diabrotica undecimpuntata | Chrysomelidae | N | [93] | |
| Diabrotica virgifera | Chrysomelidae | N | [93] | ||
| Leptinotarsa decemlineata | Chrysomelidae | N | [93] | ||
| Hippodamia convergens | Coccinellidae | N | [93] | ||
| Anthonomus grandis | Curculionidae | N | [93] | ||
| Cry2Ab | Propylea japonica | Coccinellidae | N | [97] | |
| Haptoncus luteolus | Nitidulidae | N | [95] | ||
| Cry3Aa | Rhyzophertha dominica | Bostrichidae | A | 1.17 μg/mg | [110] |
| Cylas brunneus | Brentidae | A | 1.88 μg/g | [56] | |
| Cylas puncticollis | Brentidae | A | 1.99 μg/g | [56] | |
| Apriona germari | Cerambycidae | A | [94,111] | ||
| Acanthoscelides obtectus | Chrysomelidae | A | [75] | ||
| Agelastica alni | Chrysomelidae | A | [112] | ||
| Brontispa longissimi | Chrysomelidae | A | 0.475 mg/mL | [113] | |
| Chrysomela tremulae | Chrysomelidae | A | [114] | ||
| Chrysomela scripta F | Chrysomelidae | A | [115] | ||
| Chrysomela scripta F | Chrysomelidae | A | 2.22 // 1.8 | [71,72] | |
| Colaphellus bowringi | Chrysomelidae | A | 2.68 // 1.33 | [116,117] | |
| Crioceris quaturdicerumpunctata | Chrysomelidae | A | 3.82 | [117] | |
| Diabrotica undecimpuntata | Chrysomelidae | N | [90,118] | ||
| Diabrotica virgifera | Chrysomelidae | N | [118,119] | ||
| Leptinotarsa decemlineata | Chrysomelidae | A | 1.84 // 3.56 | [73,118] | |
| Phaedom brassicae | Chrysomelidae | A | 1.11 | [117] | |
| Phaedon cochleariae | Chrysomelidae | A | [120] | ||
| Phyllotreta armoraciae | Chrysomelidae | N | [90] | ||
| Plagiodera versicolora | Chrysomelidae | A | 1.13 // 3.09 | [18] | |
| Pyrrhalta aenescens | Chrysomelidae | A | 0.22 mg/ml | [121] | |
| Pyrrhalta luteola | Chrysomelidae | A | 0.12 μg/cm2 | [49] | |
| Adalia bipunctata | Coccinellidae | N | [91] | ||
| Atheta coriaria | Coccinellidae | N | [91] | ||
| Cryptolaemus montrouzieri | Coccinellidae | N | [91] | ||
| Epilachna varivestis | Coccinellidae | A | [88] | ||
| Anthonomus grandis | Curculionidae | N | [90] | ||
| Asymmathetes vulcanorum | Curculionidae | N | [99] | ||
| Hypera postica | Curculionidae | N | [90] | ||
| Hypothenemus hampei | Curculionidae | A | [100] | ||
| Myllocerus undecimpustulatus | Curculionidae | A | 152 ng/cm2 | [122] | |
| Premnotrypes vorax | Curculionidae | LA | [123] | ||
| Sitophilus oryzae | Curculionidae | A | [124] | ||
| Amphimallon solstitiale | Scarabaeidae | A | [112] | ||
| Anomala corpulenta | Scarabaeidae | N | [116] | ||
| Melontha melontha | Scarabaeidae | A | [112] | ||
| Popillia japonica | Scarabaeidae | N | [90] | ||
| Alphitobius diaperinus | Tenebrionidae | A | 9.58 // 8 μg/cm2 | [62,125] | |
| Tribolium castaneum | Tenebrionidae | N | [96,110] | ||
| Tribolium castaneum | Tenebrionidae | A | 0.46 g/10 g | [89] | |
| Tenebrio molitor | Tenebrionidae | A | 11.4 μg/larve | [126] | |
| Tenebrio molitor | Tenebrionidae | A | [110,127] | ||
| Cry3Ba | Cylas brunneus | Brentidae | A | 1.304 μg/g | [56] |
| Cylas puncticollis | Brentidae | A | 1.273 μg/g | [56] | |
| Chrysomela scripta F | Chrysomelidae | A | [115] | ||
| Diabrotica undecimpuntata | Chrysomelidae | A | 107 ng/mm2 | [128] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | 1.35 ng/mm2 | [128] | |
| Epilachna varivestis | Coccinellidae | N | [88] | ||
| Popillia japonica | Scarabaeidae | A | 1 | [37] | |
| Tribolium castaneum | Tenebrionidae | A | 1.60 g/10 g | [89] | |
| Tribolium castaneum | Tenebrionidae | A | 13.55 mg/mL | [37,96] | |
| Cry3Bb | Cylas brunneus | Brentidae | A | 1.83 μg/g | [56] |
| Cylas puncticollis | Brentidae | A | 1.82 μg/g | [56] | |
| Anoplophora glabripennis | Cerambycidae | N | [86] | ||
| Diabrotica undecimpuntata | Chrysomelidae | A | 9.49 // 1.18 | [118,129] | |
| Diabrotica virgifera | Chrysomelidae | A | 2.10 // 5.18 | [118,129] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | 6.86 // 6.54 | [118,129] | |
| Alphitobius diaperinus | Tenebrionidae | A | 26.52 // 50 μg/cm2 | [62,125] | |
| Cry3Ca | Cylas brunneus | Brentidae | A | 0.69 μg/g | [56] |
| Cylas puncticollis | Brentidae | A | 0.57 μg/g | [56] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | 0.7 // 320.13 | [130,131] | |
| Tribolium castaneum | Tenebrionidae | N | [96] | ||
| Cry6Aa | Diabrotica virgifera | Chrysomelidae | A | 77 µg/cm2 | [66,119] |
| Hypera brunneipennis | Curculionidae | A | [66] | ||
| Hypera postica | Curculionidae | A | [66] | ||
| Cry6Ba | Hypera postica | Curculionidae | A | 280 ng/μl | [94] |
| Cry7Aa | Cylas brunneus | Brentidae | A | 0.44 μg/g | [56] |
| Cylas puncticollis | Brentidae | A | 0.34 μg/g | [56] | |
| Anoplophora glabripennis | Cerambycidae | N | [86] | ||
| Diabrotica undecimpuntata | Chrysomelidae | N | [52] | ||
| Leptinotarsa decemlineata | Chrysomelidae | A | 13.1 // 18.8 | [52,57] | |
| Anthonomus grandis | Curculionidae | N | [52] | ||
| Cry7Ab | Acanthoscelides obtectus | Chrysomelidae | A | [75] | |
| Ceratoma trifurcata | Chrysomelidae | N | [107] | ||
| Colaphellus bowringi | Chrysomelidae | A | 293.79 | [132] | |
| Pyrrhalta aenescens | Chrysomelidae | N | [58] | ||
| Henosepilachna vigintioctomaculata | Coccinellidae | A | 209 | [58,133] | |
| Anomala corpulenta | Scarabaeidae | N | [58] | ||
| Tribolium castaneum | Tenebrionidae | LA | [89] | ||
| Cry8Aa | Leptinotarsa decemlineata | Chrysomelidae | A | [134] | |
| Cotinis spp | Scarabaeidae | A | [135] | ||
| Tribolium castaneum | Tenebrionidae | LA | [89] | ||
| Cry8Ab | Holotrichia oblita | Scarabaeidae | A | 5.72 μg/g | [136] |
| Holotrichia parallela | Scarabaeidae | A | 2.00 μg/g | [136] | |
| Tenebrio molitor | Tenebrionidae | N | [136] | ||
| Cry8Ba | Diabrotica virgifera | Chrysomelidae | A | [119] | |
| Cotinis spp | Scarabaeidae | A | [137] | ||
| Cyclocephala borealis | Scarabaeidae | A | [135] | ||
| Cyclocephala pasadenae | Scarabaeidae | A | [135] | ||
| Popillia japonica | Scarabaeidae | A | [135] | ||
| Cry8Bb | Diabrotica undecimpuntata | Chrysomelidae | A | [138] | |
| Diabrotica virgifera | Chrysomelidae | A | [138] | ||
| Leptinotarsa decemlineata | Chrysomelidae | A | [138] | ||
| Cry8Ca | Anoplophora glabripennis | Cerambycidae | N | [86] | |
| Colaphellus bowringi | Chrysomelidae | N | [116] | ||
| Leptinotarsa decemlineata | Chrysomelidae | N | [116] | ||
| Epilachna varivestis | Coccinellidae | A | [88] | ||
| Anomala corpulenta | Scarabaeidae | A | 1.75 × 10 × 108 CFU/g | [116] | |
| Anomala corpulenta | Scarabaeidae | A | 1.6 × 10 × 108 CFU/g | [139,140] | |
| Anomala cuprea | Scarabaeidae | A | [141] | ||
| Anomala exoleta | Scarabaeidae | A | [142] | ||
| Holotrichia parallela | Scarabaeidae | A | 9.24 × 10 × 108 CFU/g | [140] | |
| Popillia japonica | Scarabaeidae | A | 12.3 μg/g | [35] | |
| Alphitobius diaperinus | Tenebrionidae | A | 7.71 // 10 μg/cm2 | [62,125] | |
| Tribolium castaneum | Tenebrionidae | N | [89] | ||
| Cry8Da | Anomala cuprea | Scarabaeidae | A | [143] | |
| Anomala orientalis | Scarabaeidae | A | [143] | ||
| Popillia japonica | Scarabaeidae | A | 17.0 μg/g | [35,143] | |
| Cry8Db | Popillia japonica | Scarabaeidae | A | 19.6 μg/g | [35] |
| Cry8Ea | Plagiodera versicolora | Chrysomelidae | A | [144] | |
| Anomala corpulenta | Scarabaeidae | A | [140] | ||
| Holotrichia parallela | Scarabaeidae | A | 0.9 × 10 × 108 CFU/mL | [59,64,144] | |
| Popillia japonica | Scarabaeidae | A | [144] | ||
| Tenebrio molitor | Tenebrionidae | N | [64] | ||
| Tribolium castaneum | Tenebrionidae | N | [64,89] | ||
| Cry8Fa | Anomala corpulenta | Scarabaeidae | N | [59] | |
| Holotrichia oblita | Scarabaeidae | N | [59] | ||
| Holotrichia parallela | Scarabaeidae | N | [59] | ||
| Tribolium castaneum | Tenebrionidae | N | [89] | ||
| Cry8Ga | Holotrichia oblita | Scarabaeidae | N | [60] | |
| Holotrichia parallela | Scarabaeidae | N | [60] | ||
| Cry8Ka | Anthonomus grandis | Curculionidae | A | 2.83–8.93 | [63] |
| Cry8Na | Anomala corpulenta | Scarabaeidae | N | [65] | |
| Holotrichia oblita | Scarabaeidae | N | [65] | ||
| Holotrichia parallela | Scarabaeidae | A | 3.18 × 10 × 1010 CFU/g | [65] | |
| Cry8Sa | Holotrichia serrata (F.) | Scarabaeidae | A | [145] | |
| Cry9Bb | Diabrotica undecimpuntata | Chrysomelidae | N | [146] | |
| Diabrotica virgifera | Chrysomelidae | N | [146] | ||
| Leptinotarsa decemlineata | Chrysomelidae | N | [146] | ||
| Anthonomus grandis | Curculionidae | N | [146] | ||
| Cry9Da | Anomala cuprea | Scarabaeidae | A | [77] | |
| Tribolium castaneum | Tenebrionidae | N | [89] | ||
| Cry10Aa | Anthonomus grandis | Curculionidae | A | 7.12 | [78] |
| Cry14Aa | Tribolium castaneum | Tenebrionidae | LA | [89] | |
| Cry15Aa | Leptinotarsa decemlineata | Chrysomelidae | N | [147] | |
| Cry18Aa1 | Melontha melontha | Scarabaeidae | A | [148] | |
| Cry22Aa | Anthonomus grandis | Curculionidae | A | 0.75 μg/well | [68] |
| Tribolium castaneum | Tenebrionidae | A | 1.25 g/10 g | [89] | |
| Cry22Ab | Cylas brunneus | Brentidae | A | 1.01 μg/g | [56] |
| Cylas puncticollis | Brentidae | A | 0.78 μg/g | [56] | |
| Diabrotica virgifera | Chrysomelidae | A | 39.4 μg/cm2 | [69] | |
| Diabrotica undecimpuntata | Chrysomelidae | N | [69] | ||
| Leptinotarsa decemlineata | Chrysomelidae | N | [69] | ||
| Anthonomus grandis | Curculionidae | A | 3.12 μg/well | [68] | |
| Cry22Ba | Diabrotica virgifera | Chrysomelidae | N | [68] | |
| Anthonomus grandis | Curculionidae | A | [68] | ||
| Cry23Aa/37Aa | Cylas brunneus | Brentidae | A | 0.46 μg/g | [56] |
| Cylas puncticollis | Brentidae | A | 0.42 μg/g | [56] | |
| Acanthoscelides obtectus | Chrysomelidae | A | [75] | ||
| Anthonomus grandis | Curculionidae | A | [149] | ||
| Popillia japonica | Scarabaeidae | A | [37] | ||
| Tribolium castaneum | Tenebrionidae | A | 6.30 μg SC/μL | [37,61] | |
| Cry34Aa | Diabrotica virgifera | Chrysomelidae | N | [40] | |
| Cry34Ab | Diabrotica undecimpuntata | Chrysomelidae | LA | [150] | |
| Diabrotica virgifera | Chrysomelidae | N | [40,82] | ||
| Cry34Ac | Diabrotica virgifera | Chrysomelidae | N | [40] | |
| Cry34Aa/35Aa | Diabrotica undecimpuntata | Chrysomelidae | A | 34.1 μg/well | [151] |
| Diabrotica virgifera | Chrysomelidae | A | 34 μg/cm2 | [81,151] | |
| Cry34Ab/35Ab | Rhyzophertha dominica | Bostrichidae | N | [17] | |
| Diabrotica undecimpuntata | Chrysomelidae | A | [150] | ||
| Diabrotica virgifera | Chrysomelidae | A | 3 μg/cm2 | [40,81] | |
| Oryzaephilus surinamensis | Cucujidae | LA | [17] | ||
| Sitophilus oryzae | Curculionidae | LA | [17] | ||
| Trogoderma variabile | Dermestidae | N | [17] | ||
| Tenebrio molitor | Tenebrionidae | LA | [17] | ||
| Tribolium castaneum | Tenebrionidae | LA | [17] | ||
| Tribolium castaneum | Tenebrionidae | N | [96] | ||
| Cry34Ac/35Ac | Diabrotica virgifera | Chrysomelidae | A | 7 μg/cm2 | [40,81] |
| Cry34Ba/35Ba | Diabrotica virgifera | Chrysomelidae | A | [39] | |
| Cry35Aa | Diabrotica virgifera | Chrysomelidae | N | [40] | |
| Cry35Ab | Diabrotica virgifera | Chrysomelidae | N | [40,82] | |
| Cry35Ac | Diabrotica virgifera | Chrysomelidae | N | [40] | |
| Cry36A | Diabrotica virgifera | Chrysomelidae | A | 147.3 μg/well | [151] |
| Cry37Aa | Tribolium castaneum | Tenebrionidae | A | 1.25 g/10 g | [89] |
| Cry38Aa | Diabrotica virgifera | Chrysomelidae | N | [39] | |
| Cry43Aa | Anomala cuprea | Scarabaeidae | A | [152] | |
| Cry43Ba | Anomala cuprea | Scarabaeidae | N | [152] | |
| Cry51Aa | Diabrotica undecimpuntata | Chrysomelidae | N | [79] | |
| Diabrotica virgifera | Chrysomelidae | N | [79] | ||
| Leptinotarsa decemlineata | Chrysomelidae | A | [79] | ||
| Tribolium castaneum | Tenebrionidae | A | 1.45 g/10 g | [89] | |
| Cry55Aa | Phyllotreta cruciferae | Chrysomelidae | A | [80] | |
| Tribolium castaneum | Tenebrionidae | N | [89] | ||
| Cyt1Aa | Chrysomela scripta F | Chrysomelidae | A | 132.6 | [72] |
| Cyt2Ca | Diabrotica undecimpuntata | Chrysomelidae | A | 25 μg/well | [83] |
| Diabrotica virgifera | Chrysomelidae | A | 10.8 μg/well | [83] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | [83] | ||
| Diapepes abbreviatus | Curculionidae | A | 50.7 | [84,85] | |
| Popillia japonica | Scarabaeidae | A | [83] | ||
| Tribolium castaneum | Tenebrionidae | A | [83] | ||
| Crystal Type Toxin | Target Insect | Activity (a) | LC50 (b) | Reference | |
|---|---|---|---|---|---|
| Scientific Name | Family | ||||
| Sip1Aa | Diabrotica undecimpuntata | Chrysomelidae | A | [181] | |
| Diabrotica virgifera | Chrysomelidae | A | [181] | ||
| Colaphellus bowringi | Chrysomelidae | A | 1.07 | [184] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | 24 | [181] | |
| Sip1Ab | Colaphellus bowringi | Chrysomelidae | A | 1.05 | [184] |
| Hloltrichia diomphalia | Scarabaeidae | N | [184] | ||
| Vip1Aa | Diabrotica virgifera | Chrysomelidae | N | [185] | |
| Vip1Ac | Holotrichia oblita | Scarabaeidae | N | [196] | |
| Tenebrio molitor | Tenebrionidae | N | [195] | ||
| Vip1ad | Anomala corpulenta | Scarabaeidae | N | [192] | |
| Holotrichia oblita | Scarabaeidae | N | [192] | ||
| Holotrichia parallela | Scarabaeidae | N | [192] | ||
| Vip1Da | Diabrotica virgifera | Chrysomelidae | N | [191] | |
| Vip2Aa | Diabrotica virgifera | Chrysomelidae | N | [185] | |
| Vip2Ac | Tenebrio molitor | Tenebrionidae | N | [195] | |
| Vip2Ad | Diabrotica virgifera | Chrysomelidae | N | [191] | |
| Vip2Ae | Holotrichia oblita | Scarabaeidae | N | [196] | |
| Tenebrio molitor | Tenebrionidae | N | [196] | ||
| Vip2Ag | Anomala corpulenta | Scarabaeidae | N | [192] | |
| Holotrichia oblita | Scarabaeidae | N | [192] | ||
| Holotrichia parallela | Scarabaeidae | N | [192] | ||
| Vip1Aa+Vip2Aa | Diabrotica longicornis B. | Chrysomelidae | A | [185] | |
| Diabrotica undecimpuntata | Chrysomelidae | A | [185] | ||
| Diabrotica virgifera | Chrysomelidae | A | [185] | ||
| Leptinotarsa decemlineata | Chrysomelidae | N | [185] | ||
| Tenebrio molitor | Tenebrionidae | N | [185] | ||
| Vip1Aa+Vip2Ab | Diabrotica virgifera | Chrysomelidae | A | [185] | |
| Vip1Ab+Vip2Aa | Diabrotica virgifera | Chrysomelidae | N | [185] | |
| Vip1Ab+Vip2Ab | Diabrotica virgifera | Chrysomelidae | N | [185] | |
| Vip1Ac+Vip2Ac | Tenebrio molitor | Tenebrionidae | N | [195] | |
| Vip1Ac+Vip2Ae | Holotrichia oblita | Scarabaeidae | N | [196] | |
| Tenebrio molitor | Tenebrionidae | N | [196] | ||
| Vip1Ad+Vip2Ag | Anomala corpulenta | Scarabaeidae | A | 220 ng/g soil | [192] |
| Holotrichia oblita | Scarabaeidae | A | 120 ng/g soil | [192] | |
| Holotrichia parallela | Scarabaeidae | A | 80 // 2.33 ng/g soil | [195,197] | |
| Vip1Ca+Vip2Aa | Tenebrio molitor | Tenebrionidae | N | [187] | |
| Vip1Da+Vip2Ad | Diabrotica longicornis B. | Chrysomelidae | A | 213 | [191] |
| Diabrotica undecimpuntata | Chrysomelidae | A | 4.91 | [191] | |
| Diabrotica virgifera | Chrysomelidae | A | 437 | [191] | |
| Leptinotarsa decemlineata | Chrysomelidae | A | 37 | [191] | |
| Anthonomus grandis | Curculionidae | A | 207 | [191] | |
| Vip1Ba+Vip2Ba | Diabrotica virgifera | Chrysomelidae | A | [190] | |
| Vip1Bb+Vip2Bb | Diabrotica virgifera | Chrysomelidae | A | [190] | |
| Vip3Aa | Tenebrio molitor | Tenebrionidae | N | [197] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domínguez-Arrizabalaga, M.; Villanueva, M.; Escriche, B.; Ancín-Azpilicueta, C.; Caballero, P. Insecticidal Activity of Bacillus thuringiensis Proteins against Coleopteran Pests. Toxins 2020, 12, 430. https://doi.org/10.3390/toxins12070430
Domínguez-Arrizabalaga M, Villanueva M, Escriche B, Ancín-Azpilicueta C, Caballero P. Insecticidal Activity of Bacillus thuringiensis Proteins against Coleopteran Pests. Toxins. 2020; 12(7):430. https://doi.org/10.3390/toxins12070430
Chicago/Turabian StyleDomínguez-Arrizabalaga, Mikel, Maite Villanueva, Baltasar Escriche, Carmen Ancín-Azpilicueta, and Primitivo Caballero. 2020. "Insecticidal Activity of Bacillus thuringiensis Proteins against Coleopteran Pests" Toxins 12, no. 7: 430. https://doi.org/10.3390/toxins12070430