
Marine Pollutant Tributyltin Affects DNA Methylation and Fitness of Banded Murex (Hexaplex trunculus) Populations
 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Area
2.2. Determination of Organotin Compounds
2.3. Phenotypic Responses
2.3.1. IMPOSEX Determination
2.3.2. Biomarker Activity Measurements
2.3.3. Morphometric Measurements
2.4. Methylation Sensitive Amplification Polymorphism (MSAP)
3. Results
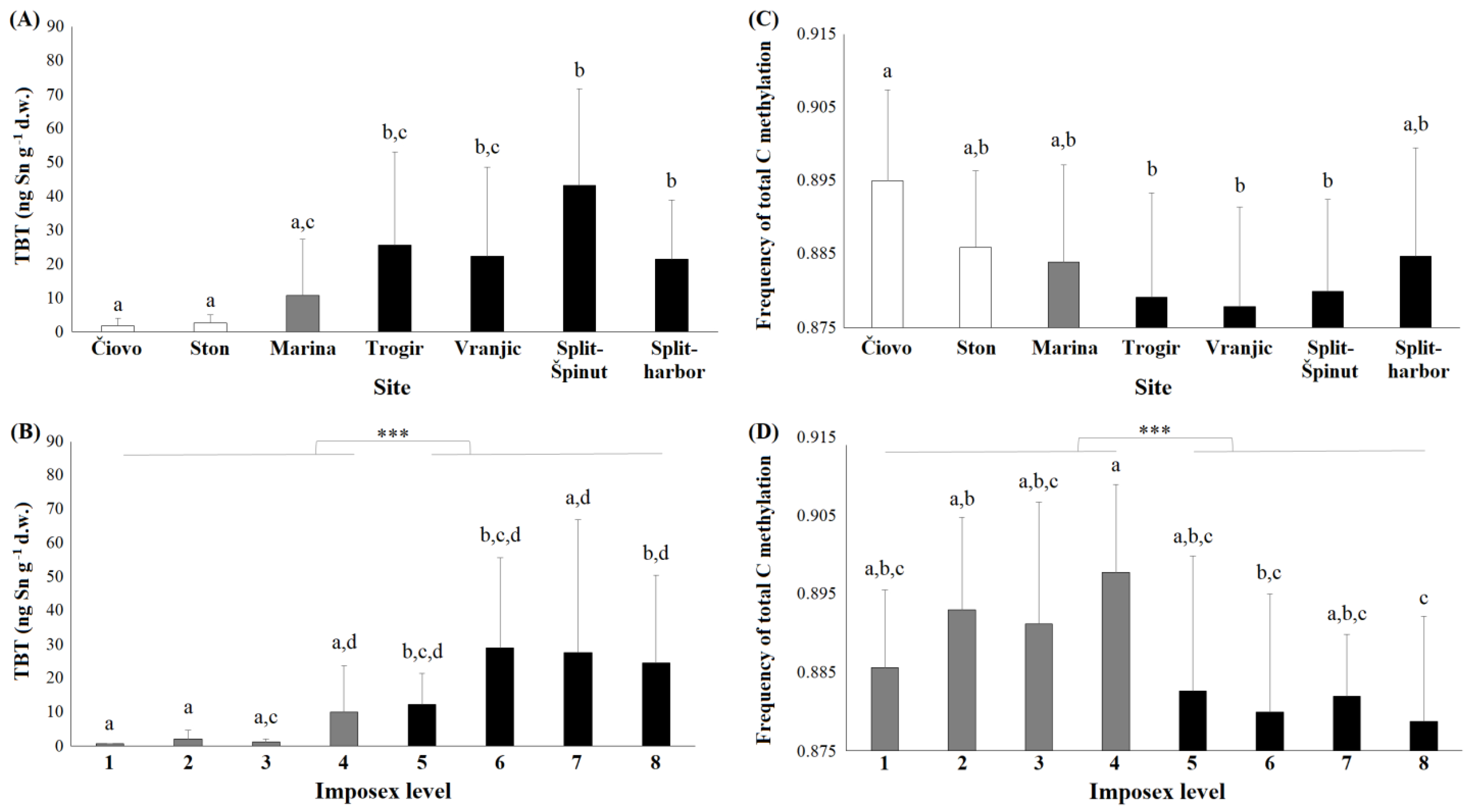
3.1. OTCs in Snail Tissue
3.2. Phenotypic Responses
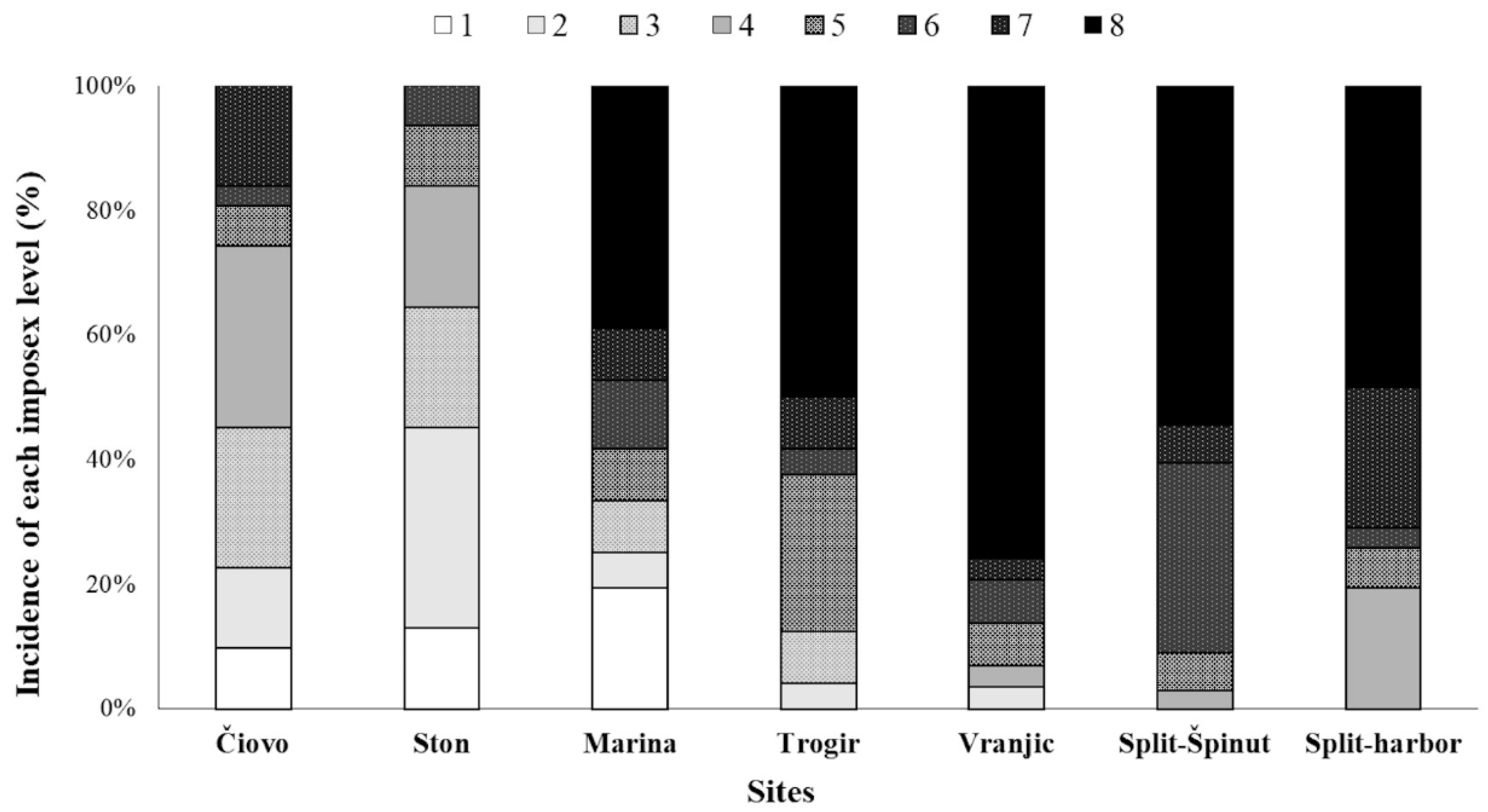
3.2.1. Imposex
3.2.2. Biomarkers
3.2.3. Morphometry
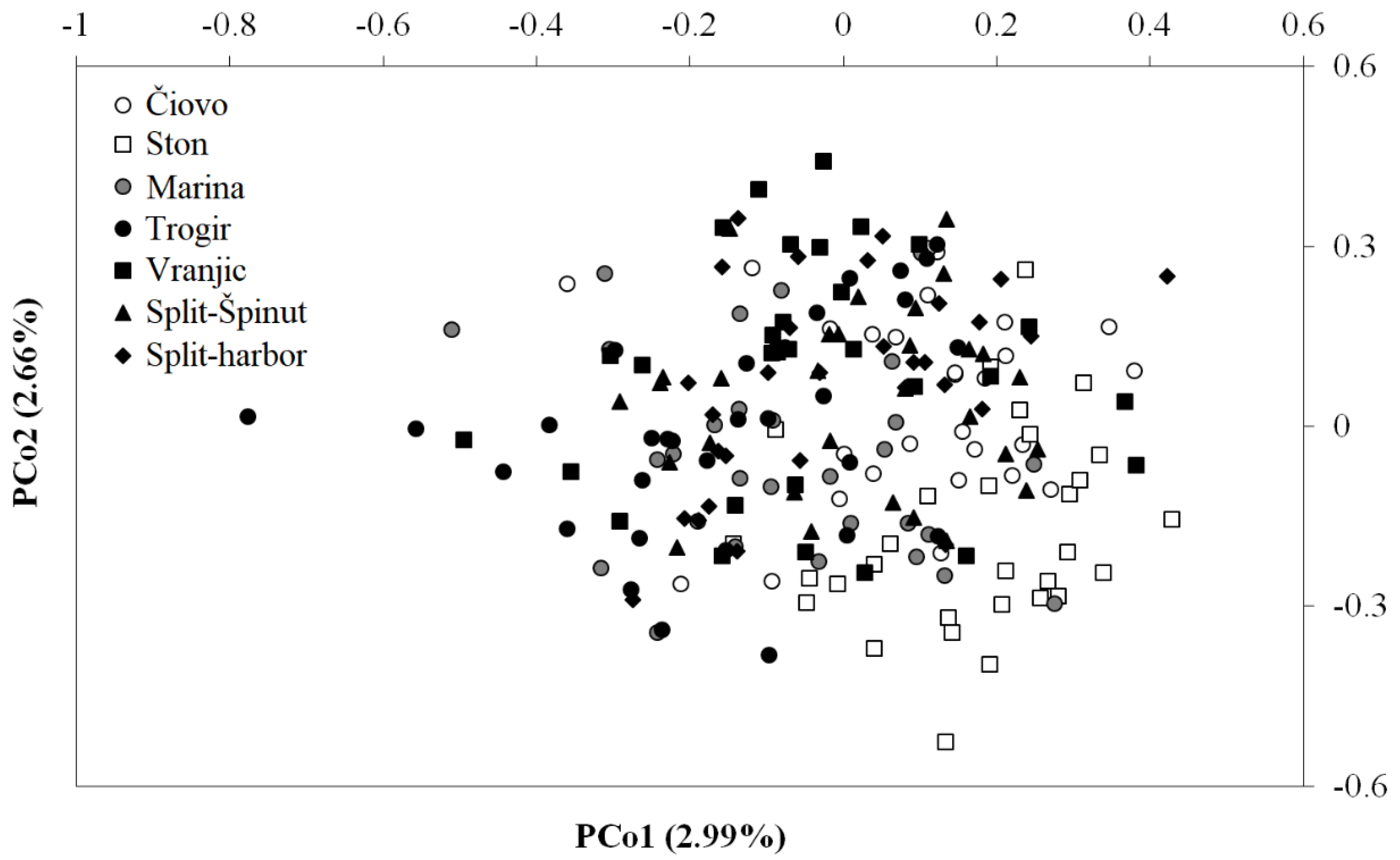
3.3. DNA Methylation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dimond, J.L.; Roberts, S.B. Germline DNA Methylation in Reef Corals: Patterns and Potential Roles in Response to Environmental Change. Mol. Ecol. 2016, 25, 1895–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, E.J.; Gluckman, P.D.; Dearden, P.K. Epigenetics, Plasticity, and Evolution: How Do We Link Epigenetic Change to Phenotype? J. Exp. Zool. Part B Mol. Dev. Evol. 2014, 322, 208–220. [Google Scholar] [CrossRef] [Green Version]
- Baldanzi, S.; Costera, E.; Marinas, D.I.; Cruces, L.; Secondary, C.A.; Author, C.; Baldanzi, S.; Watson, R.; Mcquaid, C.; Gouws, G.; et al. Evolutionary Ecology Epigenetic Variation among Natural Populations of the South African Sandhopper Talorchestia capensis. Evol. Ecol. 2017, 31, 77–91. [Google Scholar] [CrossRef]
- Jablonka, E.; Raz, G. Transgenerational Epigenetic Inheritance: Prevalence, Mechanisms, and Implications for the Study of Heredity and Evolution. Q. Rev. Biol. 2009, 84, 131–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrey, A.W.; Alvarez, M.; Foust, C.M.; Kilvitis, H.J.; Lee, J.D.; Liebl, A.L.; Martin, L.B.; Richards, C.L.; Robertson, M. Ecological Epigenetics: Beyond MS-AFLP. Integr. Comp. Biol. 2013, 53, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Abidli, S.; Castro, L.F.C.; Lahbib, Y.; Reis-Henriques, M.A.; Trigui El Menif, N.; Santos, M.M. Imposex Development in Hexaplex trunculus (Gastropoda: Caenogastropoda) Involves Changes in the Transcription Levels of the Retinoid X Receptor (RXR). Chemosphere 2013, 93, 1161–1167. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.; Reis-Henriques, M.A.; Silva, R.; Santos, A.I.; Filipe, C.; Castro, L.; Santos, M.M. Tributyltin-Induced Imposex in Marine Gastropods Involves Tissue-Specific Modulation of the Retinoid X Receptor. Aquat. Toxicol. 2011, 101, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Abidli, S.; Santos, M.M.H.; Lahbib, Y.; Castro, L.F.C.; Reis-Henriques, M.A.; Trigui El Menif, N. Tributyltin (TBT) Effects on Hexaplex trunculus and Bolinus brandaris (Gastropoda: Muricidae): Imposex Induction and Sex Hormone Levels Insights. Ecol. Indic. 2012, 13, 13–21. [Google Scholar] [CrossRef]
- Bryan, G.W.; Gibbs, P.E.; Hummerstone, L.G.; Burt, G.R. The Decline of the Gastropod Nucella lapillus Around South-West England: Evidence for the Effect of Tributyltin from Antifouling Paints. J. Mar. Biol. Assoc. U. K. 1986, 66, 611. [Google Scholar] [CrossRef]
- Furdek, M.; Vahčič, M.; Ščančar, J.; Milačič, R.; Kniewald, G.; Mikac, N. Organotin Compounds in Seawater and Mytilus galloprovincialis Mussels along the Croatian Adriatic Coast. Mar. Pollut. Bull. 2012, 64, 189–199. [Google Scholar] [CrossRef]
- Pougnet, F.; Schäfer, J.; Dutruch, L.; Garnier, C.; Tessier, E.; Dang, D.H.; Lanceleur, L.; Mullot, J.-U.; Lenoble, V.; Blanc, G. Sources and Historical Record of Tin and Butyl-Tin Species in a Mediterranean Bay (Toulon Bay, France). Environ. Sci. Pollut. Res. 2014, 21, 6640–6651. [Google Scholar] [CrossRef] [PubMed]
- Langston, W.J.; Pope, N.D.; Davey, M.; Langston, K.M.; O’ Hara, S.C.M.; Gibbs, P.E.; Pascoe, P.L. Recovery from TBT Pollution in English Channel Environments: A Problem Solved? Mar. Pollut. Bull. 2015, 95, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Furdek, M.; Mikac, N.; Bueno, M.; Tessier, E.; Cavalheiro, J.; Monperrus, M. Organotin Persistence in Contaminated Marine Sediments and Porewaters: In Situ Degradation Study Using Species-Specific Stable Isotopic Tracers. J. Hazard. Mater. 2016, 307, 263–273. [Google Scholar] [CrossRef]
- Harrison, T.D.; Gilmour, G.; McNeill, M.T.; Armour, N.; McIlroy, L. Survey of Imposex in Nucella lapillus as an Indicator of Tributyltin Pollution in Northern Irish Coastal Waters, 2004 to 2017. Mar. Pollut. Bull. 2020, 159, 111474. [Google Scholar] [CrossRef] [PubMed]
- Formalewicz, M.M.; Rampazzo, F.; Noventa, S.; Gion, C.; Petranich, E.; Crosera, M.; Covelli, S.; Faganeli, J.; Berto, D. Organotin Compounds in Touristic Marinas of the Northern Adriatic Sea: Occurrence, Speciation and Potential Recycling at the Sediment-Water Interface. Environ. Sci. Pollut. Res. 2019, 26, 31142–31157. [Google Scholar] [CrossRef] [PubMed]
- Nicolaus, E.E.M.; Barry, J. Imposex in the Dogwhelk (Nucella lapillus): 22-Year Monitoring around England and Wales. Environ. Monit. Assess. 2015, 187, 736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz, J.M.; Carro, B.; Albaina, N.; Couceiro, L.; Míguez, A.; Quintela, M.; Barreiro, R. Bi-Species Imposex Monitoring in Galicia (NW Spain) Shows Contrasting Achievement of the OSPAR Ecological Quality Objective for TBT. Mar. Pollut. Bull. 2017, 114, 715–723. [Google Scholar] [CrossRef]
- Sousa, A.C.A.; Pastorinho, M.R.; Takahashi, S.; Tanabe, S. History on Organotin Compounds, from Snails to Humans. Environ. Chem. Lett. 2014, 12, 117–137. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Zhang, J.; Chen, Y.; Zuo, Z. DNA Hypomethylation Induced by Tributyltin, Triphenyltin, and a Mixture of These in Sebastiscus marmoratus Liver. Aquat. Toxicol. 2009, 95, 93–98. [Google Scholar] [CrossRef]
- Kirchner, S.; Kieu, T.; Chow, C.; Casey, S.; Blumberg, B. Prenatal Exposure to the Environmental Obesogen Tributyltin Predisposes Multipotent Stem Cells to Become Adipocytes. Mol. Endocrinol. 2010, 24, 526–539. [Google Scholar] [CrossRef] [Green Version]
- Bastos Sales, L.; Kamstra, J.H.; Cenijn, P.H.; van Rijt, L.S.; Hamers, T.; Legler, J. Effects of Endocrine Disrupting Chemicals on in Vitro Global DNA Methylation and Adipocyte Differentiation. Toxicol. Vitr. 2013, 27, 1634–1643. [Google Scholar] [CrossRef] [PubMed]
- Cocci, P.; Mosconi, G.; Palermo, F.A. Effects of Tributyltin on Retinoid X Receptor Gene Expression and Global DNA Methylation during Intracapsular Development of the Gastropod Tritia mutabilis (Linnaeus, 1758). Environ. Toxicol. Pharmacol. 2021, 88, 103753. [Google Scholar] [CrossRef]
- Chamorro-Garcia, R.; Diaz-Castillo, C.; Shoucri, B.M.; Käch, H.; Leavitt, R.; Shioda, T.; Blumberg, B. Ancestral Perinatal Obesogen Exposure Results in a Transgenerational Thrifty Phenotype in Mice. Nat. Commun. 2017, 8, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grün, F.; Blumberg, B. Environmental Obesogens: Organotins and Endocrine Disruption via Nuclear Receptor Signaling. Endocrinology 2006, 147, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Ulloa, V.; Gonzalez-Romero, R.; Eirin-Lopez, J.M. Environmental Epigenetics: A Promising Venue for Developing next-Generation Pollution Biomonitoring Tools in Marine Invertebrates. Mar. Pollut. Bull. 2015, 98, 5–13. [Google Scholar] [CrossRef]
- Axiak, V.; Vella, A.J.; Micallef, D.; Chircop, P.; Mintoff, B. Imposex in Hexaplex trunculus (Gastropoda: Muricidae): First Results from Biomonitoring of Tributyltin Contamination in the Mediterranean. Mar. Biol. 1995, 121, 685–691. [Google Scholar] [CrossRef]
- Erdelez, A.; Furdek Turk, M.; Štambuk, A.; Župan, I.; Peharda, M. Ecological Quality Status of the Adriatic Coastal Waters Evaluated by the Organotin Pollution Biomonitoring. Mar. Pollut. Bull. 2017, 123, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Waite, M.E.; Waldock, M.J.; Thain, J.E.; Smith, D.J.; Milton, S.M. Reductions in TBT Concentrations in UK Estuaries Following Legislation in 1986 and 1987. Mar. Environ. Res. 1991, 32, 89–111. [Google Scholar] [CrossRef]
- Terlizzi, A.; Geraci, S.; Gibbs, P.E. Tributyltin (TBT)-induced Imposex in the Neogastropod Hexaplex trunculus in Italian Coastal Waters: Morphological Aspects and Ecological Implications. Ital. J. Zool. 1999, 66, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Santos, J.A.; Galante-Oliveira, S.; Barroso, C. An Innovative Statistical Approach for Analysing Non-Continuous Variables in Environmental Monitoring: Assessing Temporal Trends of TBT Pollution. J. Environ. Monit. 2011, 13, 673. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Ramos-Martinez, J.I.; Bartolomé, T.R.; Pernas, R.V. Purification and Properties of Glutathione Reductase from Hepatopancreas of Mytilus edulis L. Comp. Biochem. Physiol. Part B Comp. Biochem. 1983, 75, 689–692. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases. The First Enzymatic Step in Mercapturic Acid Formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Williams, J.A.; Stadtman, E.R.; Shacter, E. Carbonyl Assays for Determination of Oxidatively Modified Proteins. Methods Enzymol. 1994, 233, 346–357. [Google Scholar]
- Buege, J.A.; Aust, S.D. Microsomal Lipid Peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Chiavarini, S.; Massanisso, P.; Nicolai, P.; Nobili, C.; Morabito, R. Butyltins Concentration Levels and Imposex Occurrence in Snails from the Sicilian Coasts (Italy). Chemosphere 2003, 50, 311–319. [Google Scholar] [CrossRef]
- Xu, M.; Li, X.; Korban, S.S. AFLP-Based Detection of DNA Methylation. Plant Mol. Biol. Report. 2000, 18, 361–368. [Google Scholar] [CrossRef]
- Herrera, C.M.; Bazaga, P. Quantifying the Genetic Component of Phenotypic Variation in Unpedigreed Wild Plants: Tailoring Genomic Scan for within-Population Use. Mol. Ecol. 2009, 18, 2602–2614. [Google Scholar] [CrossRef]
- Pérez-Figueroa, a. Msap: A Tool for the Statistical Analysis of Methylation-Sensitive Amplified Polymorphism Data. Mol. Ecol. Resour. 2013, 13, 522–527. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Antao, T.; Beaumont, M.A. Mcheza: A Workbench to Detect Selection Using Dominant Markers. Bioinformatics 2011, 27, 1717–1718. [Google Scholar] [CrossRef] [Green Version]
- Foll, M.; Gaggiotti, O. A Genome-Scan Method to Identify Selected Loci Appropriate for Both Dominant and Codominant Markers: A Bayesian Perspective. Genetics 2008, 180, 977–993. [Google Scholar] [CrossRef] [Green Version]
- Beaumont, M.A.; Nichols, R.A. Evaluating Loci for Use in the Genetic Analysis of Population Structure. Proc. R. Soc. B Biol. Sci. 1996, 263, 1619–1626. [Google Scholar] [CrossRef]
- Zhivotovsky, L.A. Estimating Population Structure in Diploids with Multilocus Dominant DNA Markers. Mol. Ecol. 1999, 8, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Egusquiza, R.J.; Blumberg, B. Environmental Obesogens and Their Impact on Susceptibility to Obesity: New Mechanisms and Chemicals. Endocrinology 2020, 161, bqaa024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.-C.; Fonseca, E.; Park, J.C.; Yoon, D.-S.; Choi, H.; Kim, M.; Han, J.; Cho, H.-S.; Shin, K.-H.; Santos, M.L.; et al. Tributyltin Affects Retinoid X Receptor-Mediated Lipid Metabolism in the Marine Rotifer Brachionus koreanus. Environ. Sci. Technol. 2019, 53, 7830–7839. [Google Scholar] [CrossRef]
- Bigatti, G.; Carranza, A. Phenotypic Variability Associated with the Occurrence of Imposex in Odontocymbiola magellanica from Golfo Nuevo, Patagonia. J. Mar. Biol. Assoc. U. K. 2007, 87, 755. [Google Scholar] [CrossRef]
- Langston, W.J. Endocrine Disruption and Altered Sexual Development in Aquatic Organisms: An Invertebrate Perspective. J. Mar. Biol. Assoc. U. K. 2020, 100, 495–515. [Google Scholar] [CrossRef]
- Pascoal, S.; Carvalho, G.; Vasieva, O.; Hughes, R.; Cossins, A.; Fang, Y.; Ashelford, K.; Olohan, L.; Barroso, C.; Mendo, S.; et al. Transcriptomics and in Vivo Tests Reveal Novel Mechanisms Underlying Endocrine Disruption in an Ecological Sentinel, Nucella lapillus. Mol. Ecol. 2013, 22, 1589–1608. [Google Scholar] [CrossRef] [PubMed]
- Bettin, C.; Oehlmann, J.; Stroben, E. TBT-Induced Imposex in Marine Neogastropods Is Mediated by an Increasing Androgen Level. Helgoländer Meeresunters. 1996, 50, 299–317. [Google Scholar] [CrossRef] [Green Version]
- Oberdörster, E.; McClellan-Green, P. Mechanisms of Imposex Induction in the Mud Snail, Ilyanassa obsoleta: TBT as a Neurotoxin and Aromatase Inhibitor. Mar. Environ. Res. 2002, 54, 715–718. [Google Scholar] [CrossRef]
- Castro, L.; Lima, D.; Machado, A.; Melo, C.; Hiromori, Y.; Nishikawa, J.; Nakanishi, T.; Reis-Henriques, M.A.; Santos, M.M. Imposex Induction Is Mediated through the Retinoid X Receptor Signalling Pathway in the Neogastropod Nucella lapillus. Aquat. Toxicol. 2007, 85, 57–66. [Google Scholar] [CrossRef]
- Stange, D.; Sieratowicz, A.; Oehlmann, J. Imposex Development in Nucella Lapillus—Evidence for the Involvement of Retinoid X Receptor and Androgen Signalling Pathways in Vivo. Aquat. Toxicol. 2012, 106–107, 20–24. [Google Scholar] [CrossRef]
- Giulianelli, S.; Primost, M.A.; Lanari, C.; Bigatti, G. RXR Expression in Marine Gastropods with Different Sensitivity to Imposex Development. Sci. Rep. 2020, 10, 9507. [Google Scholar] [CrossRef]
- Lyssimachou, A.; Santos, J.G.; André, A.; Soares, J.; Lima, D.; Guimarães, L.; Almeida, C.M.R.; Teixeira, C.; Castro, L.F.C.; Santos, M.M. The Mammalian “Obesogen” Tributyltin Targets Hepatic Triglyceride Accumulation and the Transcriptional Regulation of Lipid Metabolism in the Liver and Brain of Zebrafish. PLoS ONE 2015, 10, e0143911. [Google Scholar] [CrossRef]
- Rey, O.; Eizaguirre, C.; Angers, B.; Baltazar-Soares, M.; Sagonas, K.; Prunier, J.G.; Blanchet, S. Linking Epigenetics and Biological Conservation: Towards a Conservation Epigenetics Perspective. Funct. Ecol. 2020, 34, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Ardura, A.; Clusa, L.; Zaiko, A.; Garcia-Vazquez, E.; Miralles, L. Stress Related Epigenetic Changes May Explain Opportunistic Success in Biological Invasions in Antipode Mussels. Sci. Rep. 2018, 8, 10793. [Google Scholar] [CrossRef] [Green Version]
- Thorson, J.L.M.; Smithson, M.; Beck, D.; Sadler-Riggleman, I.; Nilsson, E.; Dybdahl, M.; Skinner, M.K. Epigenetics and Adaptive Phenotypic Variation between Habitats in an Asexual Snail. Sci. Rep. 2017, 7, 14139. [Google Scholar] [CrossRef] [Green Version]
- Wenzel, M.A.; Piertney, S.B. Fine-Scale Population Epigenetic Structure in Relation to Gastro-Intestinal Parasite Load in Red Grouse (Lagopus lagopus scotica). Mol. Ecol. 2014, 23, 4256–4273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wogan, G.O.U.; Yuan, M.L.; Mahler, D.L.; Wang, I.J. Genome-Wide Epigenetic Isolation by Environment in a Widespread Anolis Lizard. Mol. Ecol. 2020, 29, 40–55. [Google Scholar] [CrossRef]
- Caizergues, A.E.; Le Luyer, J.; Grégoire, A.; Szulkin, M.; Senar, J.C.; Charmantier, A.; Perrier, C. Epigenetics and the City: Non-Parallel DNA Methylation Modifications across Pairs of Urban-Forest Great Tit Populations. Evol. Appl. 2022, 15, 149–165. [Google Scholar] [CrossRef]
- Schulz, B.; Eckstein, R.L.; Durka, W. Epigenetic Variation Reflects Dynamic Habitat Conditions in a Rare Floodplain Herb. Mol. Ecol. 2014, 23, 3523–3537. [Google Scholar] [CrossRef]
- Shea, N.; Pen, I.; Uller, T. Three Epigenetic Information Channels and Their Different Roles in Evolution. J. Evol. Biol. 2011, 24, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.J.; Spencer, H.G.; Donohue, K.; Sultan, S.E. How Stable “Should” Epigenetic Modifications Be? Insights from Adaptive Plasticity and Bet Hedging. Evolution 2014, 68, 632–643. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.M.; Medrano, M.; Bazaga, P. Variation in DNA Methylation Transmissibility, Genetic Heterogeneity and Fecundity-Related Traits in Natural Populations of the Perennial Herb Helleborus Foetidus. Mol. Ecol. 2014, 23, 1085–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stajic, D.; Jansen, L.E.T. Empirical Evidence for Epigenetic Inheritance Driving Evolutionary Adaptation. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200121. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, X.; Cai, Z. Tributyltin Toxicity in Abalone (Haliotis Diversicolor Supertexta) Assessed by Antioxidant Enzyme Activity, Metabolic Response, and Histopathology. J. Hazard. Mater. 2010, 183, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Ferraz da Silva, I.; Merlo, E.; Costa, C.S.; Graceli, J.B.; Rodrigues, L.C.M. Tributyltin Exposure Is Associated with Recognition Memory Impairments, Alterations in Estrogen Receptor α Protein Levels, and Oxidative Stress in the Brain of Female Mice. Front. Toxicol. 2021, 3, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Chen, C.; Li, M.; Liu, L.; Dong, K.; Chen, K.; Qiu, X. Oral Exposure to Tributyltin Induced Behavioral Abnormality and Oxidative Stress in the Eyes and Brains of Juvenile Japanese Medaka (Oryzias Latipes). Antioxidants 2021, 10, 1647. [Google Scholar] [CrossRef] [PubMed]
- Primost, M.A.; Sabatini, S.E.; Di Salvatore, P.; Ríos De Molina, M.C.; Bigatti, G. Oxidative Stress Indicators in Populations of the Gastropod Buccinanops Globulosus Affected by Imposex. J. Mar. Biol. Assoc. U. K. 2017, 97, 35–42. [Google Scholar] [CrossRef]
- Hamilton, P.B.; Rolshausen, G.; Uren Webster, T.M.; Tyler, C.R. Adaptive Capabilities and Fitness Consequences Associated with Pollution Exposure in Fish. Philos. Trans. R. Soc. B Biol. Sci. 2016, 372, 20160042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, D.; Mendonça, V.; Madeira, C.; Gaiteiro, C.; Vinagre, C.; Diniz, M.S. Molecular Assessment of Wild Populations in the Marine Realm: Importance of Taxonomic, Seasonal and Habitat Patterns in Environmental Monitoring. Sci. Total Environ. 2019, 654, 250–263. [Google Scholar] [CrossRef]
- Márquez, F.; González-José, R.; Bigatti, G. Combined Methods to Detect Pollution Effects on Shell Shape and Structure in Neogastropods. Ecol. Indic. 2011, 11, 248–254. [Google Scholar] [CrossRef]
- Primost, M.A.; Bigatti, G.; Márquez, F. Shell Shape as Indicator of Pollution in Marine Gastropods Affected by Imposex. Mar. Freshw. Res. 2015, 67, 1948–1954. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imposex Stage | Description | |
|---|---|---|
| 1 | 0 | Normal females |
| 2 | 1 | Presence of a tiny penis behind the right ocular tentacle |
| 3 | 2 | Both penis and penis duct are present |
| 4 | 3 | Development of a penis and a short vas deferens |
| 5 | 4 | Presence of a penis and a vas deferens which reaches the vagina |
| 6 | 4.3 | The vas deferens passes the vaginal opening |
| 7 | 4.7 | The vas deferens runs along 30% of the capsule gland |
| 8 | 5 | No vulva is visible, and the capsule gland may be split |
| (A) | Value | SE | t-Value | p-Value | OR | CI | AIC | |
|---|---|---|---|---|---|---|---|---|
| TBT | Coefficient | 0.03 | 0.01 | 2.47 | 0.01345 * | 1.03 | 1.01–1.05 | 236.49 |
| 1|2 | Intercepts | −2.32 | 0.53 | −4.35 | 0.00001 *** | |||
| 2|3 | −1.53 | 0.41 | −3.77 | 0.00017 *** | ||||
| 3|4 | −1.26 | 0.38 | −3.32 | 0.00091 *** | ||||
| 4|5 | −0.62 | 0.34 | −1.83 | 0.06750 | ||||
| 5|6 | 0.03 | 0.33 | 0.10 | 0.92236 | ||||
| 6|7 | 0.73 | 0.34 | 2.15 | 0.03153 * | ||||
| 7|8 | 1.07 | 0.35 | 3.06 | 0.00220 ** | ||||
| (B) | Value | SE | t-Value | p-Value | OR | CI | AIC | |
| Methylation | Coefficient | −47.17 | 12.69 | −3.72 | 0.00020 *** | 3.28 × 10−21 | 2.36 × 10−32–1.17 × 10−10 | 406.93 |
| 1|2 | Intercepts | −45.00 | 11.30 | −3.98 | 0.00007 *** | |||
| 2|3 | −43.95 | 11.29 | −3.89 | 0.00010 *** | ||||
| 3|4 | −43.36 | 11.28 | −3.84 | 0.00012 *** | ||||
| 4|5 | −42.80 | 11.26 | −3.80 | 0.00014 *** | ||||
| 5|6 | −42.37 | 11.24 | −3.77 | 0.00016 *** | ||||
| 6|7 | −41.85 | 11.22 | −3.73 | 0.00019 *** | ||||
| 7|8 | −41.44 | 11.21 | −3.70 | 0.00022 *** |
| EPIGENETIC (MSL) | GENETIC (NML) | |||
|---|---|---|---|---|
| Source of Variation | Percentage Variation | Fixation Index (* p < 0.05) | Percentage Variation | Fixation Index (* p < 0.05) |
| Among populations | 2.58 | FST: 0.02581 * | 0.96 | FST: 0.00964 * |
| Within populations | 97.42 | 99.04 | ||
| Čiovo | Ston | Marina | Trogir | Vranjic | Split-Špinut | Split-Harbour | |
| Čiovo | - | 0.006 | 0.025 * | 0 | 0.005 | 0.017 * | 0.024 * |
| Ston | 0.036 * | - | 0.028 * | 0.008 | 0.014 * | 0 | 0.018 |
| Marina | 0.048 * | 0.061 * | - | 0.028 * | 0.005 | 0.014 | 0.001 |
| Trogir | 0.016 * | 0.015 * | 0.028 * | - | 0 | 0.016 * | 0.014 |
| Vranjic | 0.019 * | 0.030 * | 0.056 * | 0.011 * | - | 0.001 | 0 |
| Split-Špinut | 0.026 * | 0.015 * | 0.049 * | 0.015 * | 0.009 * | - | 0 |
| Split-Harbour | 0.028 * | 0.022 * | 0.049 * | 0.002 | 0.003 | 0.012* | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šrut, M.; Sabolić, I.; Erdelez, A.; Grbin, D.; Furdek Turk, M.; Bakarić, R.; Peharda, M.; Štambuk, A. Marine Pollutant Tributyltin Affects DNA Methylation and Fitness of Banded Murex (Hexaplex trunculus) Populations. Toxics 2023, 11, 276. https://doi.org/10.3390/toxics11030276
Šrut M, Sabolić I, Erdelez A, Grbin D, Furdek Turk M, Bakarić R, Peharda M, Štambuk A. Marine Pollutant Tributyltin Affects DNA Methylation and Fitness of Banded Murex (Hexaplex trunculus) Populations. Toxics. 2023; 11(3):276. https://doi.org/10.3390/toxics11030276
Chicago/Turabian StyleŠrut, Maja, Iva Sabolić, Anita Erdelez, Dorotea Grbin, Martina Furdek Turk, Robert Bakarić, Melita Peharda, and Anamaria Štambuk. 2023. "Marine Pollutant Tributyltin Affects DNA Methylation and Fitness of Banded Murex (Hexaplex trunculus) Populations" Toxics 11, no. 3: 276. https://doi.org/10.3390/toxics11030276