EEG Resting Asymmetries and Frequency Oscillations in Approach/Avoidance Personality Traits: A Systematic Review



Department of Psychology, La Sapienza University of Rome, 00185 Rome, Italy
*
Authors to whom correspondence should be addressed.
Symmetry 2020, 12(10), 1712; https://doi.org/10.3390/sym12101712
Submission received: 24 September 2020
/
Revised: 12 October 2020
/
Accepted: 14 October 2020
/
Published: 16 October 2020
(This article belongs to the Special Issue Biological Psychology: Brain Asymmetry and Behavioral Brain)
Abstract
:Background: Brain cortical activity in resting electroencephalogram (EEG) recordings can be considered as measures of latent individual disposition to approach/avoidance behavior. This systematic review aims to provide an updated overview of the relationship between resting EEG cortical activity and approach/avoidance motivation personality traits. Methods: The review process was conducted according to the PRISMA-Statement, using PsycArticles, MEDLINE, Scopus, Science Citation Index, and Research Gate database. Restrictions were made by selecting EEG studies conducted in resting idling conditions, which included approach/avoidance personality traits or parallel measures, and an index of EEG brain activity. In the review 50 studies were selected, wherein 7120 healthy adult individuals participated. Results: The study of the relationship between resting EEG cortical activity and approach/avoidance personality traits provides controversial and unclear results. Therefore, the validity of resting asymmetry or frequency oscillations as a potential marker for approach/avoidance personality traits is not supported. Conclusions: There are important contextual and interactional factors not taken into account by researchers that could mediate or moderate this relationship or prove it scarcely replicable. Further, it would be necessary to conduct more sessions of EEG recordings in different seasons of the year to test the validity and the reliability of the neurobiological measures.
1. Introduction
Brain frequency oscillatory activity is defined as the real activity of the brain [1]. According to Klimesch [2], slow rhythms (delta and theta bands) have more ancient phylogenetic origin than fast rhythms (alpha, beta, gamma bands). Delta rhythm is dominant in reptiles, theta rhythm is dominant in the lower mammals, and alpha is defined as the dominant rhythm in adult humans.
The EEG alpha power at the frontal scalp is the gold standard measure used to evaluate functional inter-hemispheric asymmetry. According to Davidson [3] and Harmon-Jones [4], higher relative left frontal cortex activity is related to behavioral approach and positive emotions, whereas higher relative right frontal cortex activity is related to behavioral avoidance and negative emotions. Furthermore, according to Harmon-Jones and Gable [5], baseline electroencephalogram (EEG) measures in idling condition can be treated as personality dispositions. In this conceptual framework, the test–retest reliability of the resting EEG is comparable to the test–retest of self-reported personality trait measures, recorded in idling standard experimental conditions [6]. Thus, brain alpha oscillations can be considered as the measure of latent individual disposition to a specific style of behavior. However, the replicability of these associations with approach/avoidance personality traits is still unclear [7]. The same can be said for the function of the other oscillation frequency bands in the EEG spectrum [8], as well as the synchronization/desynchronization functions of brain rhythms, which are important issues that are sparsely discussed in terms of approach/avoidance personality traits.
This systematic review aims to provide an updated research overview of the relationship between resting EEG cortical activity in idling condition and individual differences in approach/avoidance motivation personality traits. A further aim of this review is to highlight the EEG frequency oscillation and its scalp-distributed lateral asymmetries as the most sensitive index of cortical excitability in approach/avoidance motivation personality traits.
2. Materials and Methods
This review study was conducted according to the PRISMA-Statement [9,10]. The protocol was registered on PROSPERO, CRD42020182795, and is available on the following link, https://www.crd.york.ac.uk/prospero/display_record.php?RecordID=182795.
2.1. Research Strategies
The research of literature was conducted on the international electronic databases, PsycArticles, MEDLINE, Scopus, and Science Citation Index. The last research of international literature was completed in June 2020.
In this work were included peer-reviewed full-text journal articles, written in English or Italian. The investigation was delimited to studies conducted on healthy adult human samples without restrictions to gender or ethnicity. These studies considered the relationship or association among EEG measure/s such as cortical asymmetry (e.g., frontal or posterior), frequency oscillations, synchronization or desynchronization of EEG rhythms, and personality traits such as approach/avoidance personality traits (behavioral inhibition system “BIS”, behavioral approach system “BAS”, fight-flight system “FFS” or fight-flight-freeze system “FFFS”) and/or their parallel measures (e.g., positive affect, extraversion, sensation seeking, negative affect, neuroticism, fear or state anxiety), in a resting state idling experimental condition. The research on electronic database was conducted including the following terms or keywords:
- Asymmetry and brain activity: “EEG asymmetry” OR left OR right OR lateral* OR front* OR posterior OR prefrontal OR parietal* OR electroenceph* OR oscill* OR rhythms OR coupling OR “frequency oscillations” OR synchronization OR desynchronization OR alpha OR delta OR theta OR beta OR gamma AND rest*;
- Approach/avoidance motivation: “approach motivation” OR motivation* OR approach* OR BAS OR reward* OR “positive affect” OR “avoidance motivation” OR avoid* OR “negative affect” OR BIS OR withdraw* OR inhibit* OR threat* OR fear OR FFS OR FFFS.
2.2. Eligibility Criteria
The results of the systematic research were examined by two authors (A.V.–Ph.D. student; V.D.P.–Ph.D. Tutor). A first articles’ exclusion was done by title and abstract reading, according to the following eligibility criteria: (i) EEG study conducted in resting state idling condition, only. This criterion led to the exclusion of the studies in which the participants were presented acoustic sounds [11], debated an oral presentation [12], and the resting-state EEG was recorded after physical exercise [13], after the induction of a negative mood [14], or stress condition by experimenter [15]. Measures of asymmetry and frequency oscillations recorded during hypnotic-state condition were excluded [16]. Furthermore, studies were excluded that (i) had considered resting-state EEG asymmetries or frequency oscillations as comparison measures to predict brain activity during a behavioral test [17,18,19]; (ii) report almost a self-report measure of approach/avoidance personality trait or parallel measures. This criterion excluded the studies that conceptualized EEG asymmetry as a latent state-latent trait in the absence of the self-report (e.g., [20]); (iii) involved healthy adult individuals. This criterion led to the exclusion of research conducted on a sample of preadolescents (e.g., [21]), and EEG studies conducted on clinically relevant mental disorders and other illnesses, such as Alzheimer’s and Parkinson’s disease, Down’s syndrome, chronic pain, mild cognitive impairment, and Williams’ syndrome.
In the first phase of screening, in order to not omit important research, the inclusion of studies by title and abstract readings were carried out independently by the two authors. Later, after the authors had reached a joint agreement, the first author (A.V.) examined in more depth the content of all articles that met the eligibility requirements, then moved on to the data extraction. The second author (V.D.P) supervised the data extraction to ensure it was performed correctly and carefully, and according to the eligibility criteria selected.
2.3. Data Collection and Quality Assessment
According to the PICOS approach [9,10], data collections were assessed by including in the selected studies: sample characteristics (i.e., sample size, gender, age, and education of participants); experimental design and used methods across studies; instruments used to measure personality traits of approach/avoidance motivation and parallel measures; the self-report scores; statistical analyses; and obtained results.
Two authors (A.V. and V.D.P.), independently, evaluated the risk of bias analysis. According to the criteria considered in the Cochrane Handbook for Systematic Reviews of Interventions [22,23], the second author (V.D.P.) blinded the articles selected by the first author (A.V). Therefore, the title of the study, the name of the journal, and the name of the authors were unknown to the first author (A.V.) who executed the evaluation of the articles. The quality assessment was conducted using the Joanna Briggs Institute (JBI) Critical Appraisal checklist for Analytical Cross-Sectional studies [24], modified ad hoc by considering the under-reported criteria and the aim of this review. According to Hagemann [25], different criteria adopted to EEG recording, such as referencing, analysis, and multiple sessions of measurement, represent some classical issues for resting-EEG measure validity and reliability. Thus, to evaluate the quality assessment for each selected study, the resting EEG recording method used and potential EEG fluctuations due to the state-condition changes were considered.
In line with this conceptual background, the final form of checklist consists of the six domains:
- (1)
- Adequacy of the criteria adopted for the inclusion of participants in the sample (absence of clinical psychological disorders or other diseases, suspension of drug or psychotropic substances in case used);
- (2)
- Sample and setting characteristics (mean age and standard deviation, gender, education, and handedness);
- (3)
- Methodological criteria used for the electrophysiological measures (open or closed eyes recording, reference, length, counterbalance between open/closed eyes order and participants);
- (4)
- Occurrence of contextual or interactional variables not considered in the study that could constitute confounding factors (time of day and season of EEG recording, measures of mood state before and during EEG recording, menstrual cycle for women, and gender of experimenters);
- (5)
- Reliability of electrophysiological measures (test–retest sessions of EEG recording);
- (6)
- Adequacy of the statistical analysis used (including the strategies to deal with confounding factors considered in the study).
For each research article, the methodological quality assessment was determined for each domain as low, partial, and high risk of bias (respectively, “0”, “1”, “2”), by calculating the mean score multiplied by 100. Then a cut-off level of 75% was established. The studies under or equal to 75% were considered as low risk of bias, while, the studies above 75% were considered as high risk of bias. Table 1 shows the data extracted and examined for each study included.
3. Results
3.1. Studies Selection
The flow chart (Figure 1) provides an accurate summary of the quality assessment of the articles identified through databases. The bibliographic research included all combinations of keywords and produced 5313 results. Furthermore, two additional articles were identified through other sources and included in this article. Later, duplicated studies were removed and after title and abstract readings, 2937 full-text articles were included. According to the eligibility criteria, 152 articles were reviewed by full-text screening and, consequently, 101 articles were excluded with reason. Finally, 47 articles, for a total of 50 studies, were passed to the quality assessment and were thus included in the systematic review.
3.2. Quality Assessment
Figure 2 shows the percentage of studies and articles included for the quality of the assessed criteria. Generally, 44 studies (88%) presented low scores on the risk of bias, while six studies (12%) showed high scores. A large percentage of the studies used valid methodological criteria for measuring EEG performance in idling condition and included an appropriate sample size. However, many researchers did not report controlled criteria for the inclusion of the participants in their studies (first domain), did not consider interactional or confounding variables that could influence the reliability of the evaluated relations (fourth domain) and, furthermore, they did not test–retest sessions of their EEG measures (fifth domain). These are the three domains in which risk of bias was subjectively evaluated as higher than the other domains. In contrast, statistical analysis was adequate to the outcome studied (see Figure 2).
3.3. Demographic Features
The literature research for this review was delimited by a period that ranged from January 1990 to June 2020. In the 47 articles selected, from a total of 50 studies, 7120 healthy adult participants, aged between 17 [26,33,35,43,56,71] and 82 years old [38] took part. The studies showed a percentage of men and women variable. In some cases, samples were composed by only women [26,27,57,58,60,66] and, in other cases, there were samples of only men [35], [45] (study 2 presented in the article), [52] (study 3 presented in the article), [53]. Only three studies were conducted on a sample equally balanced regarding gender [28,39,44]. However, in some studies, we did not find any information about gender [48] (study 1 presented in the article), [55,67], mean age or standard deviation [26,27,28,33,35,38,39,40,47,48,54,55,56,59,60,64,67], and education [38,44,51,52], (see Table 1). Assuming the heterogeneity of the constructs and the paradigms examined, some researchers included confounding variables (such as gender, age, education) and interactional variables (such as sex of experimenters, time of day and time of year, open and closed eyes, and EEG reference) in the statistical analyses.
3.4. Approach/Avoidance Personality Traits and Electrocortical Measures
Approach/avoidance personality traits represent long-term stable behavioral state patterns [118]. The first version of the reinforcement sensitivity theory (RST), conceptualized from animal behavior by Gray, was an extension of the theory originally postulated by Eysenck. This theory posited three systems responsible for behavior: (1) the behavioral inhibition system (BIS); (2) the behavioral approach system (BAS); (3) a not well-defined fight–flight system (FFS) activated by fear [119]. Later, Gray inserted the response of block or freeze into the FFS, and reconceptualized this system into the fight–flight–freeze system (FFFS) as the main system responsible for fear responses. The FFFS is activated only in the case of active avoidance of a threatening stimulus (escape) while, if the situation requires an attack on the threat, both the BIS and the FFFS are activated (fight).
According to the motivational model theory [119,120], a relatively greater left-frontal activity is associated with behavior that results in approaching or engaging a stimulus and is related to higher levels of behavioral approach (BAS), while a relatively greater right-frontal activity leads to the disengagement from a stimulus and is related to higher levels of withdrawal behavior.
The reinforcement sensitivity theory [121] and the motivational model [122] inspired many researchers to study the behavioral underpins in terms of neurobiological markers. Carver and White [82], using the systems conceptualized by Gray [123,124], structured the BIS/BAS scale [82] to assess the motivational model of frontal EEG asymmetry proposed by Davidson [122]. However, the problem with the BIS/BAS scales questionnaire was the lack of separation of the FFFS and the BIS, which may account for inconsistent findings obtained in past research when the BIS scale was related to resting frontal alpha activity. Recently, Neal and Gable [64] derived BIS and FFFS subscales from the original Carver and White’s scale [82], demonstrating that the BIS subscale, but not the FFFS, related to greater relative right frontal activity, and that a measure of impulsivity related to the smaller right frontal activity. However, in this vein, the most important revision was done by Corr [125] in his revised RST (r-RST), which produced the development of the Reinforcement Sensitivity Theory Personality Questionnaire “RST-PQ” [89]. In the r-RST, the function of the BIS is primarily to detect and resolve conflicts between the BAS and FFFS. In addition, in the r-RST, the BAS is reconceptualized as a multidimensional system [126] composed of the subcomponents reward interest (RI), goal-driven persistence (GDP), reward reactivity (RR), and impulsivity (Imp).
Another contribution to the definition of personality was conceptualized by Costa and McCrae who proposed the Big Five model [127]. This descriptive model defined the following five stable factors, considered as the basic vectors of the personality structure: conscientiousness; extraversion; emotional stability/instability; openness; agreeableness.
Although r-RST is a neurobiological theory and the Big Five model is descriptive, there are meeting points between the two. According to Smits and De Boeck [128], extraversion can be explained by the BAS trait, while, emotional instability (or neuroticism), can be explained by the BIS trait. Therefore, in this framework, a lot of individual disposition and state variables can be considered as parallel measures of the BIS, BAS, and FFFS motivational personality traits.
In line with this point of view, many studies have investigated the relationship between the hemispheric asymmetry of the EEG asymmetrical activation of the brain delta, theta, alpha, beta, and gamma rhythms and the approach/avoidance behavioral traits. All the studies included in this review assessed one or more measures of approach/avoidance personality traits and parallel, or related, measures. In particular, 16 articles considered the relation among resting-EEG activity and BIS/BAS personality traits [8,28,33,34,37,46], [48] (in study 2), [49,51,57,59,64,66,67,69,70].
Among studies testing the association between resting-EEG activity and BIS (or parallel measures), 11 articles have considered the role of state or trait anxiety [27,29,34,40,43,44,47,49,63,69,71], four articles studied the role of negative affect [26,28,29,57], 21 articles evaluated the role of neuroticism [29,30,31,34,35,36,37,38,39,40,43,49,51,53,54,55,56,60,61,62,70], three articles considered the role of defensiveness [27,30,43], seven articles studied the role of depression trait [27,40,42,43,44,63,71] and finally, one study considered the role of nostalgia [62].
Concerning the study of resting EEG activity and BAS parallel measures, generally, 19 articles considered the role of extraversion [29,30,31,34,35,36,37,38,39,43,51,53,54,55,56,60,61,62,70], but only in seven studies did the authors find a significant relation among this trait and EEG activity [38,43,51,52,53,54,70]. Moreover, in five articles authors studied the relation among EEG rhythms and the influence of positive affect [26,28,29,57,58], two articles considered the role of impulsive control [64,67], one study evaluated the role of positive urgency [59], and two studies considered the role of sensation seeking and risk-taking behaviors [45] (studies 1 and 2).
The association between resting-EEG activity and FFFS, was considered in four articles [64,66,67,69], but, of these, only two studies found significative results [64,66]. Furthermore, one study considered the relation among EEG gamma activity and autistic traits [65], one article studied the role of resilience [68], and two articles found a positive association between EEG alpha activity and emotional intelligence [39,50]. Some studies have observed a significant relation between autistic traits and approach/avoidance personality traits [129], as well as emotional intelligence [130] and resilience [131]. In particular, results have shown a negative correlation between resilience and the brain network flexibility for the delta, alpha, and beta bands, suggesting a robust relationship among the flexibility of human brain and resilience traits [68]. Finally, one study included in this review considered the relation between EEG activity and transliminality [41].
3.5. Approach/Avoidance Personality Traits and EEG Gamma Rhythm
Although there is still no univocal agreement among researchers regarding on the definition of EEG gamma band, in general it is referred to as the range of frequencies comprised between 30 and 70 Hz [132]. In particular, recent research classifies a slow gamma activity as being around 20–40 Hz, and a fast gamma activity as around 40–70 Hz, generated from excitatory–inhibitory interactions of pyramidal cell and interneuron networks [133] in posterior brain regions [134], visual cortex [135], temporal-parietal regions [136], and the hippocampus [137]. Furthermore, according to Fries [138], the synchronization of the gamma frequency would be involved in the synchronization of the alpha-beta feedback signal in the cortical networks.
With regard to the study of brain oscillations and personality, Jaušovec and Jaušovec [39] investigated the relationship among Big Five personality traits [94], emotional intelligence, and EEG activity in right-handed participants during an eyes-closed resting period. In this study, these authors, taking into account the role of gender, analyzed the EEG rhythms using entropy measures, fast Fourier transform (FFT), and low-resolution electromagnetic tomography (LORETA) techniques. Results highlighted that brain activity was increased in the parieto-occipital areas rather in the frontal area only for compounds among extreme personality styles (neurotic type, low emotional intelligence and agreeableness; high neuroticism, or specific combinations of personality dimensions, e.g., introverts with high emotional IQ, versus extraverts with low to average IQ). These compounds were related to the spectrum of gamma-band activity and gender [39].
Openness to experience (O—Big Five trait) has been found to be associated with transiliminality [139]. This trait has been defined as “susceptibility to, and awareness of, large volumes of imagery, ideation, and affect. These phenomena can be generated by subliminal, supraliminal, or external input” [140] (p. 327). In a recent study, Fleck and colleagues [41] studied the association among transliminality and frequency oscillations, suggesting that higher levels of transliminality are related to lower slow alpha, beta, and gamma-band activity in the left posterior cortex and lower fast alpha, lower beta, and gamma activity in the right superior temporal areas. Moreover, lower levels of transliminality were related to increased gamma-band in the mid frontal areas, than higher levels of this trait [41].
In addition to studying the relationship among personality traits and gamma-band activity, many researchers have been interested in studying the relationship within brain traits and the development of cognitive processes between personality traits. Many studies found that gamma-band activity is related to cognitive functions [141], such as perceptual binding [142], attention [143], working memory [144], language [145], and social interaction [146]. According to Groot and Van Strien [65], these domains are altered in the autism spectrum disorder (ASD). Recent studies assess that individuals with ASD have increased spontaneous gamma oscillations [147]. Thus, according to these studies and the autism spectrum hypothesis [74], De Groot and Van Strien [65] hypothesized that enhanced gamma-band activity could be considered as a biomarker of ASD—an endophenotype present in people with higher levels of autistic traits. To confute this supposition, the authors administered the Autism-Spectrum Quotient (AQ) questionnaire [74] to right-handed male and female university students and recorded their EEG activity during a resting idling condition. In this sample, results suggested that gamma power was not related to the autistic quotient “AQ” score. This result may be due to the fact that autistic traits in the general population are not strong enough to be detected [65].
In sum, the number of reports referring to the relationship between resting EEG gamma-band activity and personality traits is so limited that they cannot be discussed in depth within a context of motivational theory [3,4,89]. Thus, among the articles included in this review, only three [39,41,65] analyzed the relation between personality traits and EEG gamma-band activity. However, we believe it is important to highlight and reflect on these poor results to inspire future research exploring this relationship further.
3.6. Approach/Avoidance Personality Traits and EEG Beta Rhythm
Beta rhythm is the oscillatory neural activity in the frequency range of 14–30 Hz, generated by dipole located in the deep layer networks [148] of the pre-central and post-central cortical sites [149], and in the parietal regions [150,151]. It is the rhythm dominant in normal waking consciousness, traditionally associated with sensorimotor processing [152]. However, recent research emphasizes that beta rhythm is related to sensorial and cognitive processes, such as alerting [153], working memory [154], decision making [155], and focused mental activity [156]. Specifically, slow beta activity (12–15 Hz) is associated with coordination [157], central beta activity (15–22 Hz) is associated with mental active engagement [158], while high beta activity (22–38 Hz) is associated with metacognition [159] and integration of new experiences and semantic memory [160]. Furthermore, recent EEG studies have introduced beta (13–30 Hz) oscillations as a cortical brain rhythm that may reflect active neural inhibition [161] and excitement or trait anxiety [56].
In terms of the anxiety trait, Pavlenko and colleagues [47] studied the relation among resting EEG oscillations in healthy adult male/female participants, and personality measures of state and trait anxiety. Results highlighted, in only two studies, that state anxiety was positively correlated with the spectral power density (SPD) of central beta-band in the temporal and occipital regions of the right-hemisphere during the eyes-open recording. Moreover, the correlations of the estimates of trait anxiety with the SPDs of the beta rhythm were found in frontal and central areas of both hemispheres and parietal and occipital loci of the right hemisphere. Anxiety trait was positively correlated with the SPDs of the slow and central beta-band oscillations [47]. These results are partially in line with approach/avoidance motivational theory in which the behavioral inhibition, or its parallel anxiety measures, are associated with a greater relative right frontal cortical activation [28,64].
According to Threadgill and Gable [67], the beta activity during resting state can be assumed as a neurophysiological marker of motivated motor-action preparation. In their experiment, the authors first assessed, in a sample of university students, the behavioral approach/avoidance motivational traits (BIS/BAS Scale [82]) and the Impulsive Behavior Scale of the Regulatory Control Questionnaire (UPPS-P [90]), to evaluate the influence of behavioral disinhibition. They then recorded the EEG in a resting idling condition. In this sample, results emphasized that a greater trait approach was negatively associated with resting beta activity, while greater trait impulsivity was associated with a greater resting beta activity. Lower levels of resting beta activity in the motor cortex was found associated with traits related to motivated motor behaviors. Furthermore, according to Schutter and colleagues [8], frontal EEG asymmetry of beta activity (13–30 Hz) reflects the brain cortical excitability and approach–avoidance motivational predispositions. In this study, frontal asymmetry is a direct measure of cortical excitability and is seen in line with the approach/avoidance motivational model proposed by Davidson [3], wherein self-reported emotional tendencies for approach or avoidance are associated, respectively, with left- or right frontal asymmetry.
In sum, resting EEG beta asymmetry findings from the above-mentioned three studies included in this review are in line with our initial hypotheses. These findings indicated that in resting idling conditions, the beta frontal asymmetry [8,47,67] and the scalp-distributed beta activity can be defined as potential markers of the approach/avoidance motivation personality traits.
3.7. Approach/Avoidance Personality Trait, Interactional Variables, and EEG Alpha Rhythm
Alpha rhythm, or Berger’s rhythm, is a brain activity with a frequency ranging from 8 to 13 Hz, associated with a state of wakeful relaxation [162]. This rhythm is classified in slow alpha (8–10 Hz), generated in the anterior brain regions, and fast alpha (11–13 Hz), generated in the posterior regions [163]. The anterior and posterior systems constitute a single alpha network, distributed over the whole brain surface [164]. Moreover, the alpha rhythm is considered as a mechanism of surrounding inhibition [165], useful in increasing the signal–noise ratio and to inhibit ongoing conflicting processes [2]. According to the “idling condition hypothesis”, alpha activity reflects both the ongoing cognitive processes and the proper inhibitory mechanisms of this condition. This characterizes alpha activity as a good biological index sensitive to personality and behavior differences among individuals [166].
The association between alpha rhythm and the approach/avoidance behavioral traits was studied in more depth than other EEG rhythms. In this review of the literature, 33 studies that found a relationship between resting-EEG alpha rhythm and approach/avoidance behavioral traits, or other parallel trait measures were included. Of these studies, 27 analyzed the relationship among EEG alpha activity and approach/avoidance personality traits [26,27,28,29,30,33,35,36,37,38,39,40,42,43,44], [45] (studies 1 and 2), [46,50], [52] (studies 3 and 4), [57,59,62,63,64,71], while, the remaining six studies analyzed the influence of the interactional or contextual variables, on the relationship between approach/avoidance personality traits and resting EEG alpha activity [31,32], [48] (studies 1 and 2), [60,66]. For this reason, we decided to discuss the results highlighted by this research line in two separate sections. In the first section, we review findings on the relationship between resting EEG alpha activity and approach/avoidance personality traits. In the second section of this paragraph, we review findings supporting the influence of interactional or contextual variables on this relationship.
3.7.1. Approach/Avoidance Personality Traits and EEG Alpha Rhythm
The measure of EEG alpha spectral power, in the frontal area, is used to calculate an index of inter-hemispheric frontal asymmetry in resting condition. This index is usually calculated by subtracting the resting EEG alpha power of the cortical area of interest in the left hemisphere from that of the homologous area in the right hemisphere [167].
In a pioneer research, Tomarken and colleagues [26] found that resting EEG anterior alpha asymmetry, recorded in a sample of adult women, was related to individual differences in positive and negative affect (PA, NA) traits. In particular, they found that the activation of the anterior left hemisphere was related to relatively frontal higher levels of PA scores, while lower levels of NA were associated with an increased right hemisphere anterior activation [26]. The relation among resting EEG alpha asymmetry and positive/negative emotional traits (PA, NA) was labelled as the “emotional model” of hemispheric asymmetry. However, in other studies, the validity of this model was partially supported. Hagemann and colleagues [29] examined the relation between resting EEG alpha asymmetry and personality traits of PA and NA, neuroticism (N), and extraversion (E). In this study a significant association was highlighted between the right hemisphere activation and NA trait, but no significant association was found between alpha rhythm and PA trait. However, these authors reported, in contrast with the emotional model, a left anterior temporal activation in subjects with higher NA scores, while they did not find any significant association between NA and N or PA and E. These results suggested that the biological bases of N and NA are different [29], disconfirming the hypotheses of Eysenck and Eysenck [168], according to which N trait should be positively associated with negative affect, whereas E trait is positively associated with positive affect. Minnix and Kline [36], in contrast with Hagemann and colleagues [29], observed that higher N or emotional lability levels were associated with greater variability of mid frontal asymmetry. These authors proposed the inconsistency of the outlined relationship between neuroticism and right frontal activation, and suggested the necessity to provide a novel neurobiological index sensitive to individual differences in N and able to predict psychopathologies linked to this trait.
In general, a left hemisphere hypoactivation in anterior regions is a biological marker of affective style and related to the risk of psychopathology [27]. To assess whether resting anterior asymmetry discriminates individual differences in repressive coping styles, Tomarken and Davidson [27], in a sample of university students, studied the relationship between defensiveness copying style and EEG alpha asymmetry. In their experiment the authors administered the Marlowe-Crowne Social Desirability Scale [77] and the State-Trait Anxiety Inventory [78], before recording the EEG in resting condition. Defensiveness trait was associated with greater right frontal activation in the presence of an opposite-sex experimenter, but not with a same-sex one. In other EEG alpha asymmetry studies, in the presence of opposite-sex experimenters, but not same-sex experimenters, high-defensive participants had a relative frontal left hemisphere activation, while, low-defensive participants had a frontal right hemisphere activation [30,169]. Further, in another study, the defensiveness trait resulted in being related to the retrospective quality of parental caring [170], suggesting that repressed coping style, or defensiveness trait, and perceived maternal caring predicted left lateral frontal alpha activation [30].
According to other authors, frontal EEG asymmetry reflects not only emotion, but also individual differences in motivational personality traits [171,172]. In their pioneer experiment, Sutton and Davidson [28] showed that EEG alpha asymmetry explained more than 25% of the variance in the self-report measure of BAS and BIS traits, but prefrontal EEG asymmetry, however, was not significantly correlated with PA or NA. This research demonstrated that resting EEG alpha asymmetry can be predicted by BIS and BAS motivational traits. In this study, participants with greater prefrontal left hemisphere activation reported higher levels of BAS, whereas those with greater prefrontal right hemisphere activation reported higher levels of BIS. In this vein, the approach/withdrawal, or motivational model, of frontal EEG alpha asymmetry highlights that frontal brain activity corresponds to motivational propensities to approach versus withdraw behavioral tendencies [120,122]. Later, Coan and Allen [33], in an attempt to replicate Sutton and Davidson’s findings [28], reported that approach motivation was a valid construct associated with EEG asymmetry findings in the temperament, emotion, and psychopathological domains, but, in contrast with the prediction of the model, the right hemisphere asymmetry in higher BIS scorers was partially confirmed. Although Sutton and Davidson [28] argued that higher BIS scores were related to a greater right frontal activity, Coan and Allen [33] found only a weakly significant relationship between BIS and right hemisphere activation in the mid frontal region rather the frontal region. Probably, in this study, the discrepancy was due to the different conceptualization of withdrawal and BIS constructs used in the two studies. Davidson [120] conceptualized a withdrawal construct as the system that motivates, or potentially motivates, organisms to withdraw from sources of aversive stimulation, whereas Gray [173] conceptualized the BIS as the system that, among other things, interrupts ongoing behavior, increases arousal, and increases attention, none of which inevitably leads to a withdrawal response.
Hewig and colleagues [37] suggested that the problem of replicability could be due to the different conceptualization among the withdrawal and approach systems that could both be subsystems of the behavioral activation system. Authors proposed a new view of the approach/withdrawal model of anterior asymmetry, in which the behavioral activation system is related to bilateral frontal cortical activity, and right and left activations related, respectively, to withdrawal and approach behavior [174]. This model is in line with results of Harmon-Jones and Allen [175], who reported a positive relation among bilateral frontal cortical activity and BAS. In this vein, behavioral activation is the product of both approach and withdrawal motivational traits [37].
Research on personality correlates of right frontal alpha asymmetry has demonstrated that a higher dispositional tendency to experience withdrawal-related behaviors was related to increased proneness to nostalgia or sadness [62]. Adolph and Margraf [63] studied the relationship among symptoms of anxiety, depression, and frontal asymmetry in a sample of healthy individuals. Results indicate that higher symptom severity of depression and anxiety were correlated to a larger right frontal cortical activity. Furthermore, a larger right frontal cortical activity was influenced by anxiety symptoms [63]. In sum, frontal alpha asymmetry can be considered a biological marker for the risk of anxiety and depression [3].
From a genetic point of view, research affirmed that the relation between frontal alpha asymmetry and the risk for anxiety and depression is heritable only in young adults (males 32% and females 37%), but not in middle-aged adults [40]. In particular, the BDNF Val66Met polymorphism would be associated with the depression trait and mediated by EEG alpha power [42]. These findings highlight the utility of studying the relation among EEG measures and genotype to elucidate the pathway that elapses between the expression of an endophenotype and individual dispositions in personality traits [42]. According to Mathersul and colleagues [44], increased activity of alpha rhythm in the right parietal-temporal cortex is related to higher comorbidity of anxiety and depression, while increased right parietal-temporal alpha activity is related to anxious apprehension. These data support predictions for frontal, but not posterior regions and are in line with the motivational model of Davidson [120], in which depressed individuals differ from controls by an increase in withdrawal trait and negative affective valence, but in contrast to the valence-arousal model [176]. According to Heller [176], depressed mood is characterized by an asymmetrical profile associated with increased right frontal activity, due to a dissociation among lower and higher right parietal-temporal activity. Moreover, in this experiment, individuals that used a higher coping style oriented toward an emotional approach (through emotional expressions), had neural activities indicative of greater approach motivation [44]. Positive emotional expression traits are significantly related to greater left-sided frontal alpha asymmetry in the resting EEG condition [46].
According to Zhang and colleagues [71], participants who showed a higher relative left frontal activity during a resting state condition, exhibited fewer difficulties in everyday emotion regulation, especially in the dimension of impulsive control. Among personality traits related to impulsive control, the positive urgency (defined by Cyders and colleagues [108] as the tendency towards rash action in response to extreme positive emotional states) is related to left frontal alpha asymmetry. Gable and colleagues [59] found that higher levels of positive urgency trait were associated with a greater left frontal EEG activity, which originates from reduced right frontal activity in the inferior frontal gyrus [59]. According to these authors, a decreased right frontal activity could be considered as a potential neurobiological trait of impulsivity, related to the higher activity of the supervisory control system. This result was later confirmed by Neal and Gable [64], who suggested that impulsivity is related to reduced right frontal brain activity. Furthermore, higher BIS levels were related to greater right frontal activity, while BAS and FFFS traits (motivation to approach and motivation to withdrawal, respectively) were not associated with frontal alpha asymmetry. The authors state that regulatory control processes are associated with BIS and impulsivity and related to the right frontal activity rather than to withdrawal motivation expressed by the FFFS [64].
Another personality trait associated with impulsive control and the predisposition to approach is sensation seeking [177]. In two studies conducted in separated laboratories with different samples, Santesso and colleagues [45] examined whether the pattern of left frontal resting EEG activity related to approach-related behaviors and sensation seeking. Both studies highlighted that sensation seeking is associated with a greater left frontal alpha asymmetry in resting EEG, specifically in male participants. These authors suggested that this pattern could reflect the predisposition of sensation seekers to search out novelty stimulus or engage in risky behaviors to reach the reward.
Concerning the parallel dispositional measures within the framework of the behavioral approach construct, De Pascalis, Cozzuto, Caprara, and Alessandri [57] observed that both dispositional optimism and BAS traits are related to EEG alpha asymmetry. In this study, findings on power spectral density in the alpha band have shown a robust relationship between higher cortical activity in the left middle frontal gyrus (BA11) and BAS. Optimism was related to both anterior left frontal cortical activation in the superior frontal gyrus (BA10), and a higher right-sided cortical activation in the posterior cingulate (BA31). In particular, alpha asymmetry in the posterior cingulate cortex, BA23 and BA31 regions, was uniquely associated with positivity trait, a basic disposition necessary to integrate self-referential thought and autobiographical memories [58]. In this vein, approach behavioral trait would be associated with greater left frontal activity, while behavioral avoidance trait would be associated with the greater right frontal alpha activity [59]. However, several findings do not support the relationship between motivational theory [120] and frontal alpha asymmetry. For example, another recent study conducted by Wacker and colleague [51], did not support the predicted left frontal asymmetry relation with the trait BAS/agentic extraversion (BAS/AE), but a positive relationship among consciousness (C) and frontal alpha asymmetry was observed [51]. Furthermore, within the frame of the Big Five theory [94], it was found that the personality traits of the NEO-Five-Factor Inventory (NEO–FFI) were related to alpha-beta coupling only in male participants [38]. Mainly, men with extreme trait levels differed in slow brain alpha activity compared to women [39]. However, according to Korjus and colleagues [61], the five dimensions of NEO–FFI personality traits, as well as their subordinate measures, could not be predicted from the resting state EEG data.
At a neurobiological level, Pavlenko and colleagues [47] hypothesized that a well-developed alpha rhythm is characterized by an active and stable functioning of the cerebral dopaminergic system since they found that this pattern simultaneously serves as a prerequisite of high emotional stability and social adaptability. These findings are in line with the evolutionary assumption of Knyazev and Slobodskaya [34], suggesting that alpha rhythm reflects the adequacy of descending inhibitory control generated by the thalamocortical system, associated with cognitive performance [47]. Further, Razoumnikova’s [35] findings indicate that, in male participants, higher levels of cognitive performance would be related to enhanced cortical connectivity of fast-frequency alpha rhythm [35].
Emotional intelligence is another trait related to cognitive abilities [178] and motivational neurobiology [130]. In terms of EEG frequency oscillations and individual differences, higher levels of emotional intelligence were associated with higher levels of positive affect and lower levels of negative affect [179], and positively related to a frontal left-sided alpha EEG asymmetry [50,180].
In conclusion, the present research review examined the relationship between interhemispheric EEG asymmetry and personality traits. In this context, alpha activity, in resting condition, has been the most used to derive an index of hemispheric asymmetry. Thus, this brain oscillatory activity was the most discussed in this review. However, this does not mean that alpha rhythm has produced the most stable and reliable results. Unfortunately, the study of this relationship provides controversial and unclear results. Several studies have supported the motivational model [28,43,44,45,57,59,62,63,71], many others have supported it partially [59,64,67], and others have disconfirmed it [29,33,37,52,64,66,175].
3.7.2. Interactional and Contextual Variables in the Relationship between Personality Traits and EEG Alpha Asymmetry
Recent research has studied the influence of interactional and contextual variables in the relationship between interhemispheric EEG asymmetry and personality traits. In particular, contextual variables, such as time of day and season [48], mood state and pre- and post-cap preparation [32], and experimenter sex [31,66], can influence the relation among EEG alpha asymmetry and personality traits. To demonstrate that a relationship is reliable, multiple recording sessions should be necessary, because only half of the variance in a resting session is due to the trait influences [25]. In this framework, Peterson and Harmon-Jones [48] studied, in two different samples, the role of different seasons and time of day in the relation of resting EEG alpha asymmetry and personality traits of approach–avoidance, nurturance, and dominance. In the first experiment, the EEG baseline of participants was recorded before noon and in the afternoon, both in the spring and summer months, and in the autumn and winter months. Results highlighted that frontal alpha asymmetry in resting EEG reflects circadian and seasonal influences. The right frontal activity increased during autumn mornings. These results could explain why the relation between resting alpha asymmetry and personality traits is not replicable across studies [181].
With regard to the influence of experimental context on EEG performance, Blackhart and colleagues [32] assessed that EEG cap preparation leads to a less positive mood. In this experiment, the measures of mood (evaluated with the self-assessment manikin) most proximate to the EEG recordings were associated with asymmetry when the results of pre-cap preparation mood ratings were statistically controlled. Men and women showed a shift toward a more negative mood state post-preparation. Negative mood post-preparation, but not pre-preparation, predicted relative left frontal activation in women.
For women participants, another important intervening variable to consider before the EEG recording measures is the menstrual cycle. In particular, higher levels of neuroticism (a parallel measure of BIS) were related to lower left prefrontal activity than lower levels during the mid-late luteal phase of the woman. This relation was identified as indexed by slow alpha component, and alpha-total asymmetry scores in the prefrontal regions. Therefore, the relation among resting frontal alpha asymmetry and high/low neuroticism levels in females is moderated by the menstrual cycle [60].
Finally, in a recent study, conducted by De Pascalis, Sommer, and Scacchia [66], on a sample of right-handed female university students, the authors discussed the relevance of taking into account the gender of experimenter. In this experiment, the authors examined the association among Reinforcement Sensitivity Theory-Personality Questionnaire (RST-PQ) traits [89] and alpha asymmetry in resting EEG. Results indicate that in the total group, which included two subgroups with experimenters of different gender (male, female), FFFS was related to the greater left rather than right frontal activity, while BIS was related to the greater right frontocentral activity. These associations remained significant for the subgroup with a young same-sex experimenter, but not with an opposite-sex experimenter.
In conclusion, it can be affirmed that besides the different conceptualization models, the interactional and contextual variables can also lead to a lack of replicability of the results in relation to EEG alpha rhythm and approach/avoidance personality traits.
3.8. Approach/Avoidance Personality Traits and EEG Theta Rhythm
The theta rhythm is a slow brain activity within a typical frequency range of 4–7 Hz [182]. According to Kramis, Vanderwolf, and Land [183], this rhythm is classified as type 1 theta (8 Hz), associated with locomotion and voluntary behavior, and type 2 theta (4–7 Hz), associated with motionlessness. The main brain dipoles that generate theta rhythm are sited in the midline prefrontal region of the cortex [184], the hippocampus, and the limbic system [185].
Theta oscillations are involved in various cognitive abilities, such as associative thinking [186], the encoding of information, active exploratory movements, spatial navigation of the environment, and memory [187]. In a study, Razoumnikova [35], in a sample of adult males, studied the relationships among EEG power and coherence measures of brain oscillations, and personality traits of extraversion, neuroticism, psychoticism, sensation–intuition, thinking–feeling, judging–perceiving, and emotional intelligence.
Results showed that higher levels of emotional intelligence were characterized by an increase in theta power in the right hemisphere, while lower levels of this trait were related to increase in theta power in the left hemisphere. In the subgroup with high levels of emotional intelligence, as compared to the subgroups with low levels, there were lower levels of power of type 1 and 2 theta-bands, and slow alpha bands, while there were greater levels of power in the beta-band. Furthermore, the groups with high emotional ability, compared to low groups, were related to higher interhemispheric coherence. According to the author, the results highlighted that EEG spectral parameters, in resting EEG conditions, reflect the relationships between neuronal integration (memory) and personality/intelligence variables.
With regard to the primary personality traits, several studies suggest that the posterior–anterior distribution of resting EEG activity [37,175,188], in the delta and theta frequency range rather than alpha range [51,52,54], is associated with the extraversion personality trait. In line with this evidence, Chavanon, Wacker, and Stemmler [53] observed an association among agentic extraversion (AE), and posterior versus anterior resting EEG theta activity in the rostral anterior cingulate cortex. According to Knyazev, Bocharov, and Pylkova [56], AE trait is related to higher theta activity in the posterior default mode network and lower theta activity in the orbitofrontal cortex. These results suggest higher tonic activity in the orbitofrontal cortex and lower activity in the default mode network in extraverts compared to introvert participants.
In sum, from among the four articles included in this section [35,53,55,56], it emerges that emotional intelligence [35] and personality traits modulate the activity of theta EEG activity [55]. In particular, for AE it was highlighted that rostral anterior cingulate activity generates posterior versus anterior theta activity [53]. On this basis, it has been suggested that frontal-posterior EEG theta spectral power gradient can be considered as a marker of extraversion personality trait [56].
3.9. Approach/Avoidance Personality Traits and EEG Delta Rhythm
Delta rhythm is a slow brain activity within a frequency range of around 0.5–3 Hz, associated with regenerative processes such as deep dreamless and sleep [189]. In particular, delta activity is supposed to reflect cortical reorganization of waking circuits and it is related to the activity of the parasympathetic nervous system, and the activation of ascending cholinergic projections from the thalamus [190]. According to Knyazev [164], although the origin of delta rhythm is uncertain, the dipole generators of these oscillations are located in the anterior medial frontal cortex [191], the subcortical regions linked to the brainstem [192], the nucleus accumbens [193], and the ventral tegmental area [194]. These brain regions are associated with dopaminergic activity and the approach/avoidance motivational traits [195]. Within an evolutionary framework of the brain oscillation systems, Knyazev and Slobodskaya [34] evaluated the relationship between resting-state EEG of adult right-handed male and female university students and BIS trait (Gray–Wilson Personality Questionnaire [88]). The strength of descending noradrenergic fibres of the locus coeruleus [196] was measured by the negative correlations among delta, theta, and alpha powers, separately estimated in each EEG band [2]. Their results confirm that higher BIS scores were associated with the enhanced negative coupling alpha to the delta frequency oscillations. These results were confirmed in other research in which increased levels of delta beta coupling were associated with state anxiety [49], indicating a predominantly cortical origin of the trait anxiety. Furthermore, higher levels of state anxiety increased the alpha–delta anticorrelation and were positively related to the power of alpha oscillations, and negatively related to the power of delta oscillations [34].
In terms of anxiety trait, Eysenck [197] explained neuroticism as the product of activation of the sympathetic nervous system, so that higher scores in neuroticism (N) are positively related with greater activation levels, i.e., lower activation thresholds, within this subcortical structure. Tran and colleagues [38] investigated the relationship among individual personality differences [94] and eyes-closed EEG resting brain activity. The authors suggested that there is a significative effect of gender on N trait—women exhibited higher levels of anxiety trait than men, while, extraversion (E) and consciousness (C) traits were associated with delta and theta rhythms in all cortical regions. Generally, NEO-FFI personality traits were associated with the amplitude of alpha and beta oscillations in men [38].
The approach/avoidance personality traits and EEG activity in the delta range were also found associated with reward and stimulus salience processing in the reward circuit [164]. In a recent research, De Pascalis, Vecchio and Cirillo [69] tested whether cortical–subcortical coupling would increase as a function of decreased delta (theta) or higher beta (gamma) activity in a sample of right-handed university students during a resting anxiogenic situation and a relaxation situation. To evaluate the influence of state anxiety and approach/avoidance personality traits on these neurobiological processes, participants completed the State Anxiety Inventory [198] and the Reinforcement Sensitivity Theory Personality Questionnaire [89]. During the resting anxiety condition, a significant positive between-subject delta–beta correlation was observed. This association was significantly higher than the association observed during the relaxation condition. In the anxiety, but not in the relaxation group, a delta–beta coupling for the low delta activity was observed. In addition, in the anxiety condition, BIS was significantly associated with a higher strength of within-subject delta–beta coupling, while, in the relaxation group, BIS was positively associated with delta–theta coupling. In both groups, BAS goal-driven persistence sub-trait (BAS-GDP) was positively associated with higher delta–gamma coupling [69]. These results suggested that the coupling between slow and fast EEG frequency oscillations reflects cortical–subcortical interaction [164,199]. However, these findings should be interpreted with caution, because data findings relied on two separate samples of the university students and not on the general population.
In terms of approach motivation behavior, the AE trait can be defined as the major expression of behavior and is associated with dopamine neural activity [200]. Wacker and Gatt [51] stated that resting posterior versus frontal EEG delta–theta activity is both sensitive to pharmacological manipulations of neural dopamine and associated with the AE. Furthermore, posterior versus frontal resting EEG delta/theta activity represents the molecular genetic basis of agentic extraversion associated with Catechol-O-methyltransferase Val158Met (COMT VAL/MET) polymorphism [52], and is sensitive to dopamine D2 receptor antagonist-induced changes in dopaminergic activity [54]. Studying the relationship among the posterior–frontal distribution of slow oscillations, Koehler and colleagues [54] confirmed Depue’s and Collins’ statement that agentic extraversion is linked to individual differences in dopaminergic activity, although these authors did not emphasize a significant association among extraversion and DRD2.
In sum, the eight studies reviewed and analyzed in this section have demonstrated that the psychological traits of personality modulate the activity of delta as well as theta EEG activity [55]. In terms of the BAS/E trait, some of these studies highlighted that this trait was related to delta–theta coupling at posterior versus frontal brain regions [51,54]. Interesting, BAS-GDP was positively related to higher delta–gamma coupling [69]. Using the NEO-FFI, Tran and colleagues [38] found that delta–theta activity was related to E and C, while the alpha–beta coupling was related to personality traits in males only. In terms of BIS/N, higher delta–alpha coupling has been related to increased behavioral inhibition [34], while the increased delta–beta coupling is associated with state anxiety [49]. Finally, the BIS trait was related to higher delta–beta coupling during a state of resting anxiety and with higher delta–theta coupling during a neutral resting condition [69]. These results disconfirmed the frontal alpha asymmetry hypotheses for the BAS trait [52]. Findings from the above-mentioned studies suggest that the evaluation of coupling among slow and fast rhythms is a good method for evaluating cortical–subcortical excitability in behavioral processes.
4. Discussion
This review mainly aimed at studying the relationship among resting EEG cortical activity in resting condition, and individual differences in approach/avoidance motivation personality traits. Another aim was to understand which EEG frequency oscillation and the associated scalp-distributed lateral asymmetries can be defined as an index of cortical excitability sensitive to the approach/avoidance motivational personality traits.
Research on databases has shown that many studies have satisfied the inclusion criteria, confirming the high interest of the researchers on this topic. This interest was opened to many factors, such as the study of the neurobiological underpinning of behavioral individual differences [26,27,28,29], the validation of the motivational personality theory [8,33,46,52,56,57,59,64,67], and the investigation of the neurobiological cortical markers related to the risk of psychopathological disease such as anxiety and depression [30,31,37,41,43,44,62,63], or behavioral disinhibition [45,64,71].
In accordance with the motivational model theory [121], research showed that the EEG gamma frequency oscillation, or the associated scalp-distributed lateral asymmetries, is not a good index of approach/avoidance motivation personality traits. Interesting results were found from the study of resting-EEG beta-band activity and approach/avoidance personality traits, wherein the results highlighted that resting beta asymmetry is a neurophysiological marker of approach/avoidance personality traits [8,67]. With regard to the study of EEG alpha asymmetry and the affective/motivational dispositions, some research has supported the motivational model theory [28,44,45,57,59,62], others have partially supported it [5,33,43,64,66,71], or disconfirmed it [29,37,52,176].
A number of studies on the relationship between brain oscillations and personality have outlined that frontal–posterior EEG theta spectral power gradient is a good index of cortical excitability in the approach/avoidance motivational personality traits [51,53,54], and a stable individual measure related to extraversion personality trait [56]. Regarding to the BIS functions, higher delta–alpha coupling is related to increased behavioral inhibition [34], while an increased delta–beta coupling is associated with state anxiety [49]. In sum, all these findings indicate that the coupling among fast and slow rhythms can be considered as indexes of cortical–-subcortical interactive influences on personality traits [69].
However, the current review suggests that these results should be interpreted with caution, because several methodological problems persist in this field of research. The qualitative assessment of the selected articles emphasized a medium risk of bias (see, the last column” Total” in Figure 2). This result could depend on several factors. Many researchers did not conduct any evaluation referring to the anamnesis of the participants enrolled in their research. They did not consider the possible influence of organic, psychological, psychopathological, neurological disorder, or brain trauma, and the eventual intake of drugs or psychostimulants before the resting-EEG recording was done [26,27,29,30,31,32,33,34,35,36,37,38,39,40,45,47,48,49,50,52,54,55,56,59,60,61,62,64,65,67]. Further, although in some studies researchers have evaluated the history of diseases and the eventual use of drugs, they did not control for the limitation of the assumption of psychotropic substance intake, like caffeine or nicotine, two hours before the rest-EEG recording [18,28,39,68,70]. It is known that the taking of drugs or psychotropic substances by the participant may impair the brain activity during resting-EEG recording [201,202], and that also, the presence of an organic or neuropsychological disease [203] may alter the brain rhythms of the participants, confounding the validity of the results obtained. Moreover, many authors did not take into consideration the handedness of the participants in the relationship studied [33,34,35,38,39,41,42,47,48,49,50,51,53,54,55,61,62,65,70,71]. Although some authors did not consider this factor as relevant [40], we think it is important to do so [72,73].
From a methodological point of view, the studies selected had not always adopted adequate criteria for EEG measurement, nor for counterbalancing the order of the opened-/closed-eye sequences of EEG recordings [31,36,41,42,44,45,46,47,49,50,51,62], or these data were not clearly reported [28,30,32,39,56,57,58,64,69,70,71]. For example, in the study by Konareva, a counterbalancing between the eye conditions was not adopted [55], while Korjus and colleagues [61] generalized their results obtained from five resting-state experiments conducted using different samples and different recording times (i.e., 1 min, 2 min, and three separate sessions of 1 min, respectively). In particular, in these studies, the method used produced several limitations regarding the validity and reliability of the significant relations obtained. Additionally, the use of counterbalance strategies is necessary to reduce the order effect and the sequence effect, which increases the validity of the EEG measures.
In addition to the appropriate reference electrode placement and the length of EEG recording, there are other important factors to consider for reliable asymmetry or frequency oscillation measures. According to Hagemann [25], the good reliability of the asymmetry measure in a resting-state can only be achieved if the EEG is recorded for at least 4 min. However, in some reviewed studies, many researchers conducted EEG recordings for less than 4 min [30,31,32,36,40,43,49,56,65,69], others for 2 min [42,44,47,51,57,58,60] and, finally, some for 2 min or less [45,71]. Moreover, some other researchers used short recording segments, such as 15 [60] or 30 s [62,71]. Although some studies showed that alpha power at single sites shows good reliability estimates for EEG segments as short as 20 or 30 s [204,205], this observation may not be generalizable to other asymmetry measures obtained for other brain oscillations [25]. This could produce a lack of relation between brain rhythms and personality traits that would otherwise be significant in longer EEG recording segments.
The method used for EEG recordings in eyes-open and eyes-closed conditions should be carefully reconsidered. Many researchers have recorded resting-EEG by asking the participants to refrain from blinking and/or moving their eyes by fixating on a cross to minimize ocular artefacts [51,57,58,61,66], or they did resting-EEG recordings in an eyes-closed condition [35,40,42,54,68]. In the first case, the method is confounding, because blinks and spontaneous eye movements are controlled by several autonomic brain systems [206], and the instruction to suppress these systems may act as a secondary task [207], while in the second case, to study the brain activity only in a closed-eyes state leads to a poor external validity of the EEG measures obtained. Further, the use of different reference schemes is also another important factor to consider in EEG asymmetry research.
In the studies selected, different EEG reference schemes were used, such as the average to overall electrodes [43,44,45,54,67], the reference to average voltage [8], the link to an active electrode [47], or the link to the left earlobe [46,48,59,71], but among these, the average over two electrodes positioned on the earlobes or mastoids [28,30,31,32,34,35,36,38,39,40,41,42,43,49,50,51,56,60,61,65,69], and the reference electrode positioned in the middle of the scalp between Fz and Cz sites [54], FCz site [70], or on the Cz site [26,27,29,37,45,52,57,58] were the most commonly used. These differences undermine the generalizability of the findings. However, despite the fact that many researchers were using the Cz electrode, positioned in the middle of the scalp as a reference, this could lead to numerous problems regarding the validity of the EEG measurement. According to Hagemann [208], the vertex reference at Cz has an unfavorable signal-to-noise ratio, because Cz is an active electrical site, and depending on the amplitude and phase relations between the two target sites and the reference site, the true amplitude asymmetry of the target sites may be enhanced, mitigated, or even reverted. Moreover, the same considerations are true to any other active electrode used as a reference scheme for the EEG measurement. For this reason, the linked earlobes/mastoids reference scheme would seem the better solution to this problem—the average of A1 and A2 is substantially less active than the cephalic target sites [25]. However, although this scheme has been repeatedly indicated and considered as the best solution in this EEG research field [25,208], today still no consensus has been reached among researchers. Another solution, as suggested by Coan and Allen [33], could be to use a general linear model of analysis, which allows the inclusion of a repeated measures factor by considering as factor the reference schemes used for EEG recordings.
The relationship between brain activity and approach/avoidance behavioral traits could be moderated not only by the EEG reference schemes, but also by the interactional or contextual variables (e.g., participant’s mood, sex of experimenter, or time of day and time of year in which the EEG is recorded). Two studies have reported that time of day and time of year are correlated with alpha asymmetric frontal cortical activity (studies 1 and 2 [48]), revealing that the right frontal activity is highest during autumn mornings. These results had important health implications and suggested the EEG as an endophenotype of the risk of depression. Research has argued that the time of year is associated with an increased depression state [209], the time of day is associated with higher cortisol levels [210], and the cortisol level has been linked with withdrawal motivation. Thus, a greater relative right frontal activity at rest in the autumn mornings could be due to the combination of variables associated with a decreased approach motivation and increased withdrawal motivation [5].
Several studies have suggested that controlling the mood before EEG recordings might increase the predictive value of the studied relationship, because the procedure of fitting an EEG cap, such as the abrasion of the scalp, and the application of gel during the EEG preparation, is aversive and, thus, may induce a negative affective state or avoidance. Therefore, differences in transient mood may contribute to the state variance of resting asymmetry [32]. Perhaps another factor that contributes to the state variance of resting asymmetry is a transient state of approach and withdrawal motivation. Higher motivation to approach may respond to the novel lab situation with greater relative left frontal activity, whereas individuals with higher withdrawal and motivation to inhibition may respond to the novel lab situation with greater right frontal activation [5,66]. Therefore, it is very important to consider the influence of interactional and contextual variables that can confound the relationship between EEG activity and motivational personality traits. This influence can be reduced by conducting multiple EEG recordings [3]. However, among the studies reviewed, only a few have evaluated the construct validity of EEG measurements in multiple recording sessions with a total of three EEG measurements 3 weeks apart [26,27,28,37,43,60], and only one study analyzed the stability of the relation, over a long time of retest, for a total of three EEG measurements 1 year apart (study 3 [52]). Thus, it is to be assumed that the construct validity of the studies included in this review is poor. The measures conducted in multiple EEG sessions across time can constitute better reliability and construct validity of the measured relation [25,27].
Finally, in terms of the quality assessment of the studies included in this review (Figure 1), we think that the statistical analysis was adequate to the studied outcome. The studies used valid and reliable methods to evaluate the influence of the brain cortical activity on the individual disposition of approach/avoidance personality domains, including appropriate analysis for the used sample size and adequate control for the confounding variables considered by the authors. However, some studies used statistical models without considering the role of education [44,52], gender [18,28,30,33,36,37,41,47,48,50,51,55,59,62,68], and age [38,42,54,68], in the relation studied. Gender and age can modulate the relationship between brain activity and motivational traits of personality. In the studies by Santesso and colleagues [45], for example, the authors highlighted that higher sensation-seeking levels were related to a greater left frontal activity at rest in male participants only, while other studies have observed higher neuroticism levels in women than in men. In particular, Huang and colleagues [60] demonstrated that the menstrual cycle is an interactional variable that can alter the resting frontal alpha asymmetry. Unfortunately, only five articles selected in this review considered the influence of the menstrual cycle in personality traits as an interactional variable [57,58,60,66,69].
In terms of age influence, results appear to be more consistent. However, some studies evaluating the role of age on the frontal asymmetry heritability, found that frontal-alpha asymmetry is heritable only in young adults, but not in middle-aged adults [40].
Although we checked for the method used in the reviewed studies, this work presents some limitations that could undermine the generalizability of the findings. The first limitation is related to the heterogeneity of samples and methods used for the EEG measurements. The second is due to the heterogeneity of the EEG phenomena studied (asymmetry, and/or brain rhythms), which have determined a lack of a quantitative analysis in the meta-analysis. This would have given a greater strength to the inferences by examining the size of the effects studied.
Another limitation is related to the choice to include only academic articles published in peer-review journals. This aspect could have limited the selection of only those studies that have obtained results in line with the literature, and consequently have influenced the publication bias. Therefore, the presented results could have an overestimation of the relationship observed. In addition, the choice to select only the studies published in English and in Italian could have led to the deletion of studies conducted in other populations.
In conclusion, it can be assumed that research on the relationship of resting EEG cortical activity in idling condition and the approach/withdrawal motivational model has provided controversial and unclear results. Findings have shown that gamma, delta, and alpha frequencies are not good indicators of cortical excitability that can be associated with approach/avoidance motivation personality traits, and although in some studies beta and theta frequencies have resulted as good markers of approach/avoidance motivational behavior, the number of studies is scarce. Finally, to confirm these promising but “preliminary” results, and to give greater validity, future research should consider the role of gender contextual interactional variables, discussed above, in these relations, and conducting multiple sessions of resting-EEG recording.
5. Conclusions
Observations derived from this review are in accordance with Harmon-Jones and Gable’s [5] considerations that the manifestation of trait frontal asymmetry is until today an unknown phenomenon, as well as the association of scalp-distributed lateral asymmetries with approach/avoidance motivation personality traits. Although the reliability of resting EEG measures in idling condition is hypothetically comparable with the reliability of self-reported personality traits and can be used as a valid signature of a person, the trait frontal asymmetry could simply reflect individual differences of frontal asymmetry in an idling rest condition [6]. This endophenotype can be, in fact, influenced by situational and interactional variables related to the experimental contest, such as the psychological state of the individual, and the unconsciously cognitive processes performed by the participant during the resting EEG recording, which are impossible to control [211]. Besides, it is necessary to remember that the study of electrocortical correlates should be interpreted with the utmost caution, because genetic factors [51,54], hereditary [33,40,42], and situational factors [212] can all interact to modify behavior and brain dynamics, and expressions of personality traits [213].
Author Contributions
V.D.P. and A.V.: Conception of review, literature research, and evaluation of quality assessment. A.V.: wrote the manuscript. V.D.P.: Supervised the manuscript. All authors read and agreed with the published version of the manuscript.
Funding
Funding for Research from Ph.D. program in Psychology and Cognitive Science. Department of Psychology, Sapienza University of Rome (Italy).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Başar, E.; Schürmann, M.; Demiralp, T.; Başar-Eroglu, C.; Ademoglu, A. Event-related oscillations are ‘real brain responses’—Wavelet analysis and new strategies. Int. J. Psychophysiol. 2001, 39, 91–127. [Google Scholar] [CrossRef]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Davidson, R.J. Anterior cerebral asymmetry and the nature of emotion. Brain Cogn. 1992, 20, 125–151. [Google Scholar] [CrossRef]
- Harmon-Jones, E. Clarifying the emotive functions of asymmetrical frontal cortical activity. Psychophysiology 2003, 40, 838–848. [Google Scholar] [CrossRef] [PubMed]
- Harmon-Jones, E.; Gable, P.A. On the role of asymmetric frontal cortical activity in approach and withdrawal motivation: An updated review of the evidence. Psychophysiology 2018, 55, e12879. [Google Scholar] [CrossRef] [PubMed]
- Näpflin, M.; Wildi, M.; Sarnthein, J. Test–retest reliability of resting EEG spectra validates a statistical signature of persons. Clin. Neurophysiol. 2007, 118, 2519–2524. [Google Scholar] [CrossRef] [PubMed]
- Kuper, N.; Käckenmester, W.; Wacker, J. Resting frontal EEG asymmetry and personality traits: A meta-analysis. Eur. J. Pers. 2019, 33, 154–175. [Google Scholar] [CrossRef]
- Schutter, D.J.; de Weijer, A.D.; Meuwese, J.D.; Morgan, B.; van Honk, J. Interrelations between motivational stance, cortical excitability, and the frontal electroencephalogram asymmetry of emotion: A transcranial magnetic stimulation study. Hum. Brain Mapp. 2008, 29, 574–580. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 6, e1000100. [Google Scholar]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Prisma Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Knyazev, G.G.; Savostyanov, A.N.; Levin, E.A. Alpha oscillations as a correlate of trait anxiety. Int. J. Psychophysiol. 2004, 53, 147–160. [Google Scholar] [CrossRef]
- Beaton, E.A.; Schmidt, L.A.; Ashbaugh, A.R.; Santesso, D.L.; Antony, M.M.; McCabe, R.E. Resting and reactive frontal brain electrical activity (EEG) among a non-clinical sample of socially anxious adults: Does concurrent depressive mood matter? Neuropsychiatr. Dis. Treat. 2008, 4, 187. [Google Scholar]
- Hall, E.E.; Ekkekakis, P.; Petruzzello, S.J. Predicting affective responses to exercise using resting EEG frontal asymmetry: Does intensity matter? Biol. Psychol. 2010, 83, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Keune, P.M.; Bostanov, V.; Kotchoubey, B.; Hautzinger, M. Mindfulness versus rumination and behavioral inhibition: A perspective from research on frontal brain asymmetry. Pers. Individ. Differ. 2012, 53, 323–328. [Google Scholar] [CrossRef]
- Pérez-Edgar, K.; Kujawa, A.; Nelson, S.K.; Cole, C.; Zapp, D.J. The relation between electroencephalogram asymmetry and attention biases to threat at baseline and under stress. Brain Cogn. 2013, 82, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamieson, G.A.; Burgess, A.P. Hypnotic induction is followed by state-like changes in the organization of EEG functional connectivity in the theta and beta frequency bands in high-hypnotically susceptible individuals. Front. Hum. Neurosci. 2014, 8, 528. [Google Scholar] [CrossRef] [Green Version]
- Käckenmester, W.; Kroencke, L.; Wacker, J. Frontal asymmetry predicts the incentive value of perceptual information. Int. J. Psychophysiol. 2018, 134, 22–29. [Google Scholar] [CrossRef]
- Adolph, D.; von Glischinski, M.; Wannemüller, A.; Margraf, J. The influence of frontal alpha-asymmetry on the processing of approach-and withdrawal-related stimuli—A multichannel psychophysiology study. Psychophysiology 2017, 54, 1295–1310. [Google Scholar] [CrossRef]
- Massar, S.A.A.; Rossi, V.; Schutter, D.J.L.G.; Kenemans, J.L. Baseline EEG theta/beta ratio and punishment sensitivity as biomarkers for feedback-related negativity (FRN) and risk-taking. Clin. Neurophysiol. 2012, 123, 1958–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagemann, D.; Naumann, E.; Thayer, J.F.; Bartussek, D. Does resting electroencephalograph asymmetry reflect a trait? An application of latent state-trait theory. J. Pers. Soc. Psychol. 2002, 82, 619. [Google Scholar] [CrossRef]
- Harmon-Jones, E.; Allen, J.J. Anger and frontal brain activity: EEG asymmetry consistent with approach motivation despite negative affective valence. J. Pers. Soc. Psychol. 1998, 74, 1310. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Sterne, J.A. The Cochrane Collaboration’s tool for assessing risk of bias in randomised trials. BMJ 2011, 343, d5928. [Google Scholar] [CrossRef] [Green Version]
- Jadad, A.R. Blind assessment of the quality of trial reports. Control. Clin. Trials 1996, 17, 1–12. [Google Scholar] [CrossRef]
- Institute, J.B. The Joanna Briggs Institute Critical Appraisal Tools for Use in JBI Systematic Reviews Checklist for Analytical Cross Sectional Studies; The Joanna Briggs Institute: North Adelaide, Australia, 2017. [Google Scholar]
- Hagemann, D. Individual differences in anterior EEG asymmetry: Methodological problems and solutions. Biol. Psychol. 2004, 67, 157–182. [Google Scholar] [CrossRef]
- Tomarken, A.J.; Davidson, R.J.; Wheeler, R.E.; Doss, R.C. Individual differences in anterior brain asymmetry and fundamental dimensions of emotion. J. Pers. Soc. Psychol. 1992, 62, 676. [Google Scholar] [CrossRef]
- Tomarken, A.J.; Davidson, R.J. Frontal brain activation in repressors and nonrepressors. J. Abnorm. Psychol. 1994, 103, 339. [Google Scholar] [CrossRef] [PubMed]
- Sutton, S.K.; Davidson, R.J. Prefrontal brain asymmetry: A biological substrate of the behavioral approach and inhibition systems. Psychol. Sci. 1997, 8, 204–210. [Google Scholar] [CrossRef]
- Hagemann, D.; Naumann, E.; Lürken, A.; Becker, G.; Maier, S.; Bartussek, D. EEG asymmetry, dispositional mood and personality. Pers. Individ. Differ. 1999, 27, 541–568. [Google Scholar] [CrossRef]
- Kline, J.P.; Knapp-Kline, K.; Schwartz, G.E.; Russek, L.G. Anterior asymmetry, defensiveness, and perceptions of parental caring. Pers. Individ. Differ. 2001, 31, 1135–1145. [Google Scholar] [CrossRef]
- Kline, J.P.; Blackhart, G.C.; Joiner, T.E. Sex, lie scales, and electrode caps: An interpersonal context for defensiveness and anterior electroencephalographic asymmetry. Pers. Individ. Differ. 2002, 33, 459–478. [Google Scholar] [CrossRef]
- Blackhart, G.C.; Kline, J.P.; Donohue, K.F.; LaRowe, S.D.; Joiner, T.E. Affective responses to EEG preparation and their link to resting anterior EEG asymmetry. Pers. Individ. Differ. 2002, 32, 167–174. [Google Scholar] [CrossRef]
- Coan, J.A.; Allen, J.J. Frontal EEG asymmetry and the behavioral activation and inhibition systems. Psychophysiology 2003, 40, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Knyazev, G.G.; Slobodskaya, H.R. Personality trait of behavioral inhibition is associated with oscillatory systems reciprocal relationships. Int. J. Psychophysiol. 2003, 48, 247–261. [Google Scholar] [CrossRef]
- Razoumnikova, O. Interaction of personality and intelligence factors in cortex activity modulation. Pers. Individ. Differ. 2003, 35, 135–162. [Google Scholar] [CrossRef]
- Minnix, J.A.; Kline, J.P. Neuroticism predicts resting frontal EEG asymmetry variability. Pers. Individ. Differ. 2004, 36, 823–832. [Google Scholar] [CrossRef]
- Hewig, J.; Hagemann, D.; Seifert, J.; Naumann, E.; Bartussek, D. The relation of cortical activity and BIS/BAS on the trait level. Biol. Psychol. 2006, 71, 42–53. [Google Scholar] [CrossRef]
- Tran, Y.; Craig, A.; Boord, P.; Connell, K.; Cooper, N.; Gordon, E. Personality traits and its association with resting regional brain activity. Int. J. Psychophysiol. 2006, 60, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Jaušovec, N.; Jaušovec, K. Personality, gender and brain oscillations. Int. J. Psychophysiol. 2007, 66, 215–224. [Google Scholar] [CrossRef]
- Smit, D.J.A.; Posthuma, D.; Boomsma, D.I.; De Geus, E.J.C. The relation between frontal EEG asymmetry and the risk for anxiety and depression. Biol. Psychol. 2007, 74, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Fleck, J.I.; Green, D.L.; Stevenson, J.L.; Payne, L.; Bowden, E.M.; Jung-Beeman, M.; Kounios, J. The transliminal brain at rest: Baseline EEG, unusual experiences, and access to unconscious mental activity. Cortex 2008, 44, 1353–1363. [Google Scholar] [CrossRef] [Green Version]
- Gatt, J.M.; Kuan, S.A.; Dobson-Stone, C.; Paul, R.H.; Joffe, R.T.; Kemp, A.H.; Williams, L.M. Association between BDNF Val66Met polymorphism and trait depression is mediated via resting EEG alpha band activity. Biol. Psychol. 2008, 79, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Kline, J.P.; Allen, S. The failed repressor: EEG asymmetry as a moderator of the relation between defensiveness and depressive symptoms. Int. J. Psychophysiol. 2008, 68, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Mathersul, D.; Williams, L.M.; Hopkinson, P.J.; Kemp, A.H. Investigating models of affect: Relationships among EEG alpha asymmetry, depression, and anxiety. Emotion 2008, 8, 560. [Google Scholar] [CrossRef]
- Santesso, D.L.; Segalowitz, S.J.; Ashbaugh, A.R.; Antony, M.M.; McCabe, R.E.; Schmidt, L.A. Frontal EEG asymmetry and sensation seeking in young adults. Biol. Psychol. 2008, 78, 164–172. [Google Scholar] [CrossRef]
- Master, S.L.; Amodio, D.M.; Stanton, A.L.; Yee, C.M.; Hilmert, C.J.; Taylor, S.E. Neurobiological correlates of coping through emotional approach. Brain Behav. Immun. 2009, 23, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Pavlenko, V.B.; Chernyi, S.V.; Goubkina, D.G. EEG correlates of anxiety and emotional stability in adult healthy subjects. Neurophysiology 2009, 41, 337–345. [Google Scholar] [CrossRef]
- Peterson, C.K.; Harmon-Jones, E. Circadian and seasonal variability of resting frontal EEG asymmetry. Biol. Psychol. 2009, 80, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Knyazev, G.G.; Slobodskoi-Plyusnin, Y.Y.; Savost’yanov, A.N.; Levin, E.A.; Bocharov, A.V. Reciprocal relationships between the oscillatory systems of the brain. Neurosci. Behav. Physiol. 2010, 40, 29. [Google Scholar] [CrossRef]
- Mikolajczak, M.; Bodarwé, K.; Laloyaux, O.; Hansenne, M.; Nelis, D. Association between frontal EEG asymmetries and emotional intelligence among adults. Pers. Individ. Differ. 2010, 48, 177–181. [Google Scholar] [CrossRef]
- Wacker, J.; Gatt, J.M. Resting posterior versus frontal delta/theta EEG activity is associated with extraversion and the COMT VAL158MET polymorphism. Neurosci. Lett. 2010, 478, 88–92. [Google Scholar] [CrossRef]
- Wacker, J.; Chavanon, M.L.; Stemmler, G. Resting EEG signatures of agentic extraversion: New results and meta-analytic integration. J. Res. Pers. 2010, 44, 167–179. [Google Scholar] [CrossRef]
- Chavanon, M.L.; Wacker, J.; Stemmler, G. Rostral anterior cingulate activity generates posterior versus anterior theta activity linked to agentic extraversion. Cogn. Affect. Behav. Neurosci. 2011, 11, 172–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehler, S.; Wacker, J.; Odorfer, T.; Reif, A.; Gallinat, J.; Fallgatter, A.J.; Herrmann, M.J. Resting posterior minus frontal EEG slow oscillations is associated with extraversion and DRD2 genotype. Biol. Psychol. 2011, 87, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Konareva, I.N. Modulation of low-frequency EEG rhythms under conditions of an activation reaction: Dependence on psychological characteristics of personality. Neurophysiology 2011, 42, 434–445. [Google Scholar] [CrossRef]
- Knyazev, G.G.; Bocharov, A.V.; Pylkova, L.V. Extraversion and fronto-posterior EEG spectral power gradient: An independent component analysis. Biol. Psychol. 2012, 89, 515–524. [Google Scholar] [CrossRef] [PubMed]
- De Pascalis, V.; Cozzuto, G.; Caprara, G.V.; Alessandri, G. Relations among EEG-alpha asymmetry, BIS/BAS, and dispositional optimism. Biol. Psychol. 2013, 94, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Alessandri, G.; Caprara, G.V.; De Pascalis, V. Relations among EEG-alpha asymmetry and positivity personality trait. Brain Cogn. 2015, 97, 10–21. [Google Scholar] [CrossRef]
- Gable, P.A.; Mechin, N.C.; Hicks, J.A.; Adams, D.L. Supervisory control system and frontal asymmetry: Neurophysiological traits of emotion-based impulsivity. Soc. Cogn. Affect. Neurosci. 2015, 10, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhou, R.; Cui, H.; Wu, M.; Wang, Q.; Zhao, Y.; Liu, Y. Variations in resting frontal alpha asymmetry between high-and low-neuroticism females across the menstrual cycle. Psychophysiology 2015, 52, 182–191. [Google Scholar] [CrossRef]
- Korjus, K.; Uusberg, A.; Uusberg, H.; Kuldkepp, N.; Kreegipuu, K.; Allik, J.; Aru, J. Personality cannot be predicted from the power of resting state EEG. Front. Hum. Neurosci. 2015, 9, 63. [Google Scholar] [CrossRef] [Green Version]
- Tullett, A.M.; Wildschut, T.; Sedikides, C.; Inzlicht, M. Right-frontal cortical asymmetry predicts increased proneness to nostalgia. Psychophysiology 2015, 52, 990–996. [Google Scholar] [CrossRef] [Green Version]
- Adolph, D.; Margraf, J. The differential relationship between trait anxiety, depression, and resting frontal α-asymmetry. J. Neural Transm. 2017, 124, 379–386. [Google Scholar] [CrossRef]
- Neal, L.B.; Gable, P.A. Regulatory control and impulsivity relate to resting frontal activity. Soc. Cogn. Affect. Neurosci. 2017, 12, 1377–1383. [Google Scholar] [CrossRef]
- De Groot, K.; Van Strien, J.W. Spontaneous resting-state gamma oscillations are not predictive of autistic traits in the general population. Eur. J. Neurosci. 2018, 48, 2928–2937. [Google Scholar] [CrossRef]
- De Pascalis, V.; Sommer, K.; Scacchia, P. Resting frontal asymmetry and reward sensitivity theory motivational traits. Sci. Rep. 2018, 8, 13154. [Google Scholar] [CrossRef]
- Threadgill, A.H.; Gable, P.A. Resting beta activation and trait motivation: Neurophysiological markers of motivated motor-action preparation. Int. J. Psychophysiol. 2018, 127, 46–51. [Google Scholar] [CrossRef]
- Paban, V.; Modolo, J.; Mheich, A.; Hassan, M. Psychological resilience correlates with EEG source-space brain network flexibility. Network Neurosci. 2019, 3, 539–550. [Google Scholar] [CrossRef]
- De Pascalis, V.; Vecchio, A.; Cirillo, G. Resting anxiety increases EEG delta–beta correlation: Relationships with the Reinforcement Sensitivity Theory Personality traits. Pers. Individ. Differ. 2020, 156, 109796. [Google Scholar] [CrossRef]
- Zanesco, A.P.; King, B.G.; Skwara, A.C.; Saron, C.D. Within and between-person correlates of the temporal dynamics of resting EEG microstates. NeuroImage 2020, 211, 116631. [Google Scholar] [CrossRef]
- Zhang, J.; Hua, Y.; Xiu, L.; Oei, T.P.; Hu, P. Resting state frontal alpha asymmetry predicts emotion regulation difficulties in impulse control. Pers. Individ. Differ. 2020, 159, 109870. [Google Scholar] [CrossRef]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Chapman, L.J.; Chapman, J.P. The measurement of handedness. Brain Cogn. 1987, 6, 175–183. [Google Scholar] [CrossRef]
- Baron-Cohen, S.; Wheelwright, S.; Skinner, R.; Martin, J.; Clubley, E. The autism-spectrum quotient (AQ): Evidence from asperger syndrome/high-functioning autism, malesand females, scientists and mathematicians. J. Autism Dev. Disord. 2001, 31, 5–17. [Google Scholar] [CrossRef]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and validation of brief measures of positive and negative affect: The PANAS scales. J. Pers. Soc. Psychol. 1988, 54, 1063. [Google Scholar] [CrossRef]
- Larsen, R.J.; Diener, E.; Cropanzano, R.S. Cognitive operations associated with individual differences in affect intensity. J. Pers. Soc. Psychol. 1987, 53, 767. [Google Scholar] [CrossRef]
- Crowne, D.P.; Marlowe, D. Social Desirability Scale; John Wiley: Hoboken, NJ, USA, 1964. [Google Scholar]
- Spielberger, C.D.; Gorsuch, R.L.; Lushene, R.E. State-Trait Anxiety Inventory Manual; Mind Garden, Inc.: Palo Alto, CA, USA, 1970. [Google Scholar]
- Bendig, A.W. The development of a short form of the Manifest Anxiety Scale. J. Consult. Clin. Psychol. 1956, 20, 384. [Google Scholar] [CrossRef]
- Beck, A.T.; Ward, C.; Mendelson, M.; Mock, J.; Erbaugh, J. Beck depression inventory (BDI). Arch. Gen. Psychiatry 1961, 4, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Zerssen, D.; Koeller, D.M. Depressivitätsskala: Manual [Depression Scale: Manual]; Weinheim: Göttingen, Germany, 1976. [Google Scholar]
- Carver, C.S.; White, T.L. Behavioral inhibition, behavioral activation, and affective responses to impending reward and punishment: The BIS/BAS scales. J. Pers. Soc. Psychol. 1994, 67, 319. [Google Scholar] [CrossRef]
- Hartig, J.; Moosbrugger, H. Die “ARES-Skalen” zurErfassung der individuellen BIS-und BAS-Sensitivität. ZDDP 2003, 24, 293–310. [Google Scholar]
- Rosenberg, M. Rosenberg self-esteem scale (RSE). Accept. Commit. Ther. Meas. Package 1965, 61, 18. [Google Scholar]
- Eysenck, H.J.; SBG, E. Eysenck Personality Questionnaire-Junior (EPQ-J) & Adult (EPQ-A); Hodder and Stoughton Educational: London, UK, 1975. [Google Scholar]
- Russek, L.G.; Schwartz, G.E. Perceptions of parental caring predict health status in midlife: A 35-year follow-up of the Harvard Mastery of Stress Study. Psychosom. Med. 1997, 59, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Mehrabian, A.; Russell, J.A. An Approach to Environmental Psychology; The MIT Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Wilson, G.D.; Gray, J.A.; Barrett, P.T. A factor analysis of the Gray-Wilson personality questionnaire. Pers. Individ. Differ. 1990, 11, 1037–1044. [Google Scholar] [CrossRef]
- Corr, P.J.; Cooper, A.J. The Reinforcement Sensitivity Theory of Personality Questionnaire (RST-PQ): Development and validation. Psychol. Assess. 2016, 28, 1427. [Google Scholar] [CrossRef]
- Whiteside, S.P.; Lynam, D.R.; Miller, J.D.; Reynolds, S.K. Validation of the UPPS impulsive behaviour scale: A four-factor model of impulsivity. Eur. J. Pers. 2005, 19, 559–574. [Google Scholar] [CrossRef]
- Ovchinnikov, B.V.; Pavlov, K.V.; Vladimirova, I.M. Vashpsikhologicheskiy Tip [Your Psychological Type]; Andreev Isynov’ya Publ.: St. Petersburg, Russia, 1994. [Google Scholar]
- Diener, E.D.; Emmons, R.A.; Larsen, R.J.; Griffin, S. The satisfaction with life scale. J. Pers. Assess. 1985, 49, 71–75. [Google Scholar] [CrossRef]
- Amthauer, R. Intelligenz-Struktur-Test.: IST 70; Verlag für Psychologie Hogrefe: Göttingen, Germany, 1970. [Google Scholar]
- Costa, P.T.; McCrae, R.R. Normal personality assessment in clinical practice: The NEO Personality Inventory. J. Pers. Assess. 1992, 4, 5. [Google Scholar] [CrossRef]
- Costa, P.T.; McCrae, R.R. Revised NEO Personality Inventory (NEO-PI-R) and Neo Five-Factor Inventory (NEO-FFI); Psychological Assessment Resources: Odessa, FL, USA, 1992. [Google Scholar]
- John, O.P.; Naumann, L.P.; Soto, C.J. Handbook of Personality: Theory and Research, 3rd ed.; Guilford Press: New York, NY, USA, 2008; pp. 114–158. [Google Scholar]
- Batcho, K.I. Nostalgia: A psychological perspective. Percept. Mot. Ski. 1995, 80, 131–143. [Google Scholar] [CrossRef]
- Barrett, F.S.; Grimm, K.J.; Robins, R.W.; Wildschut, T.; Sedikides, C.; Janata, P. Music-evoked nostalgia: Affect, memory, and personality. Emotion 2010, 10, 390–403. [Google Scholar] [CrossRef] [Green Version]
- Mayer, J.D.; Salovey, P.; Caruso, D.R. Mayer-Salovey-Caruso Emotional Intelligence Test (MSCEIT) Item Booklet; University of New Hampshire: Durham, NC, USA, 2002. [Google Scholar]
- Achenbach, T.M. Conceptualization of developmental psychopathology. In Handbook of Developmental Psychopathology; Springer: Boston, MA, USA, 1990; pp. 3–14. [Google Scholar]
- De Wilde, G.J.S. Nenrotische Labiliteit, Gemeten Volgens de Vragenlijst Methode [Neuroticism, Measured by Means of a Self Report Inventory]; Van Rossen: Amsterdam, The Netherlands, 1970. [Google Scholar]
- Lange, R.; Thalbourne, M.A.; Houran, J.; Storm, L. The Revised Transliminality Scale: Reliability and validity data from a Rasch top-down purification procedure. Conscious. Cogn. 2000, 9, 591–617. [Google Scholar] [CrossRef]
- Connor, K.M.; Davidson, J.R. Development of a new resilience scale: The Connor-Davidson resilience scale (CD-RISC). Depress. Anxiety 2003, 18, 76–82. [Google Scholar] [CrossRef]
- Lovibond, P.F.; Lovibond, S.H. The structure of negative emotional states: Comparison of the Depression Anxiety Stress Scales (DASS) with the Beck Depression and Anxiety Inventories. Behav. Res. Ther. 1995, 33, 335–343. [Google Scholar] [CrossRef]
- Gratz, K.L.; Roemer, L. Multidimensional assessment of emotion regulation and dysregulation: Development, factor structure, and initial validation of the difficulties in emotion regulation scale. J. Psychopathol. Behav. Assess. 2004, 26, 41–54. [Google Scholar] [CrossRef]
- Scheier, M.F.; Carver, C.S.; Bridges, M.W. Distinguishing optimism from neuroticism (and trait anxiety, self-mastery, and self-esteem): A reevaluation of the Life Orientation Test. J. Pers. Soc. Psychol. 1994, 67, 1063. [Google Scholar] [CrossRef]
- Caprara, G.V.; Alessandri, G.; Eisenberg, N.; Kupfer, A.; Steca, P.; Caprara, M.G.; Abela, J. The positivity scale. Psychol. Assess. 2012, 24, 701. [Google Scholar] [CrossRef] [PubMed]
- Cyders, M.A.; Smith, G.T.; Spillane, N.S.; Fischer, S.; Annus, A.M.; Peterson, C. Integration of impulsivity and positive mood to predict risky behavior: Development and validation of a measure of positive urgency. Psychol. Assess. 2007, 19, 107. [Google Scholar] [CrossRef]
- Zuckerman, M. Behavioral Expressions and Biosocial Bases of Sensation Seeking; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Fromme, K.; Katz, E.C.; Rivet, K. Outcome expectancies and risk-taking behavior. Cogn. Ther. Res. 1997, 21, 421–442. [Google Scholar] [CrossRef]
- Stanton, A.L.; Danoff-Burg, S.; Cameron, C.L.; Ellis, A.P. Coping through emotional approach: Problems of conceptualizaton and confounding. J. Pers. Soc. Psychol. 1994, 66, 350. [Google Scholar] [CrossRef]
- Steyer, R.; Schwenkmezger, P.; Notz, P.; Eid, M. Multidimensional Mood State Questionnaire (MDBF); Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- Cattell, H.E.; Mead, A.D. The Sixteen Personality Factor Questionnaire (16PF). In The SAGE Handbook of Personality Theory and Assessment; Boyle, G.J., Matthews, G., Saklofske, D.H., Eds.; Personality Measurement and Testing; Sage Publications, Inc.: Thousand Oaks, CA, USA, 2008; Volume 2, pp. 135–159. [Google Scholar] [CrossRef]
- Ruch, W.; Angleitner, A.; Strelau, J. The Strelau Temperament Inventory—Revised (STI-R): Validity studies. Eur. J. Pers. 1991, 5, 287–308. [Google Scholar] [CrossRef]
- Rusalov, V.M. Object-related and communicative aspects of human temperament: A new questionnaire of the structure of temperament. Pers. Individ. Differ. 1989, 10, 817–827. [Google Scholar] [CrossRef]
- Wiggins, J.S.; Trapnell, P.; Phillips, N. Psychometric and geometric characteristics of the Revised Interpersonal Adjective Scales (IAS-R). Multivar. Behav. Res. 1988, 23, 517–530. [Google Scholar] [CrossRef]
- Petrides, K.V.; Pérez, J.C.; Furnham, A. The Trait Emotional Intelligence Questionnaire (TEIQue). A measure of emotional self-efficacy. In Proceedings of the XI Biennial Meeting of the International Society for the Study of the Individual Differences, Graz, Austria, 13 July 2003. [Google Scholar]
- McNaughton, N.; DeYoung, C.G.; Corr, P.J. Neuroimaging Personality, Social Cognition, and Character; Academic Press: Cambridge, MA, USA, 2016; pp. 25–49. [Google Scholar]
- Gray, J.A. A Model for Personality; Springer: Berlin/Heidelberg, Germany, 1981; pp. 246–276. [Google Scholar]
- Davidson, R.J. Anterior electrophysiological asymmetries, emotion, and depression: Conceptual and methodological conundrums. Psychophysiology 1998, 35, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.A. Explorations in Temperament; Springer: Boston, MA, USA, 1991; pp. 105–128. [Google Scholar]
- Davidson, R.J. Parsing affective space: Perspectives from neuropsychology and psychophysiology. Neuropsychology 1993, 7, 464. [Google Scholar] [CrossRef]
- Gray, J.A.; Nebylitsyn, V.D. Biological Bases of Individual Behavior; Academic Press: Cambridge, MA, USA, 1972; pp. 182–205. [Google Scholar]
- Gray, J.A. Brain systems that mediate both emotion and cognition. Cogn. Emot. 1990, 4, 269–288. [Google Scholar] [CrossRef]
- Corr, P.J. Reinforcement Sensitivity Theory (RST): Introduction. In The Reinforcement Sensitivity Theory of Personality; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Corr, P.J. Reinforcement sensitivity theory of personality questionnaires: Structural survey with recommendations. Pers. Individ. Differ. 2016, 89, 60–64. [Google Scholar] [CrossRef]
- Costa, P.T.; McCrae, R.R. The NEO Personalityinventory; Psychological Assessment Resources: Odessa, FL, USA, 1985. [Google Scholar]
- Smits, D.J.; Boeck, P.D. From BIS/BAS to the big five. EPJ 2006, 20, 255–270. [Google Scholar] [CrossRef]
- Matyjek, M.; Bayer, M.; Dziobek, I. Autistic Traits Affect Reward Anticipation but not Reception. Sci. Rep. 2020, 10, 8396. [Google Scholar] [CrossRef]
- Bacon, A.M.; Corr, P.J. Motivating emotional intelligence: A reinforcement sensitivity theory (RST) perspective. Motiv. Emot. 2017, 41, 254–264. [Google Scholar] [CrossRef]
- Taubitz, L.E.; Pedersen, W.S.; Larson, C.L. BAS Reward Responsiveness: A unique predictor of positive psychological functioning. Pers. Individ. Differ. 2015, 80, 107–112. [Google Scholar] [CrossRef]
- Jasper, H.H.; Andrews, H.L. Electro-encephalography: III. Normal differentiation of occipital and precentral regions in man. Arch. Neurol. Psychiatry 1938, 39, 96–115. [Google Scholar] [CrossRef] [Green Version]
- Buzsáki, G.; Wang, X.J. Mechanisms of gamma oscillations. Annu. Rev. Neurosci. 2012, 35, 203–225. [Google Scholar] [CrossRef] [Green Version]
- Murty, D.V.; Shirhatti, V.; Ravishankar, P.; Ray, S. Large visual stimuli induce two distinct gamma oscillations in primate visual cortex. J. Neurosci. 2018, 38, 2730–2744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veit, J.; Hakim, R.; Jadi, M.P.; Sejnowski, T.J.; Adesnik, H. Cortical gamma band synchronization through somatostatin interneurons. Nat. Neurosci. 2017, 20, 951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinreich, S.; Djalovski, A.; Kraus, L.; Louzoun, Y.; Feldman, R. Brain-to-brain synchrony during naturalistic social interactions. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Colgin, L.L.; Denninger, T.; Fyhn, M.; Hafting, T.; Bonnevie, T.; Jensen, O.; Moser, E.I. Frequency of gamma oscillations routes flow of information in the hippocampus. Nature 2009, 462, 353–357. [Google Scholar] [CrossRef]
- Fries, P. Rhythms for cognition: Communication through coherence. Neuron 2015, 88, 220–235. [Google Scholar] [CrossRef] [Green Version]
- Thalbourne, M.A. Transliminality: A review. IJP 2000, 11, 1–34. [Google Scholar]
- Thalbourne, M.A.; Bartemucci, L.; Delin, P.S.; Fox, B.; Nofi, O. Transliminality: Its nature and correlates. J. Am. Soc. Psych. Res. 1997, 91, 305–331. [Google Scholar]
- Das, N.; Gastaut, H. Variations in the electrical activity of the brain, heart, and skeletal muscles during yogic meditation and trance. Clin. Neurophysiol. 1955, 6, 211–219. [Google Scholar]
- Gray, C.M.; König, P.; Engel, A.K.; Singer, W. Oscillatory responses in cat visual cortex exhibit inter-columnar synchronization which reflects global stimulus properties. Nature 1989, 338, 334–337. [Google Scholar] [CrossRef]
- Gregoriou, G.G.; Gotts, S.J.; Zhou, H.; Desimone, R. High-frequency, long-range coupling between prefrontal and visual cortex during attention. Science 2009, 324, 1207–1210. [Google Scholar] [CrossRef] [Green Version]
- Pesaran, B.; Pezaris, J.S.; Sahani, M.; Mitra, P.P.; Andersen, R.A. Temporal structure in neuronal activity during working memory in macaque parietal cortex. Nat. Neurosci. 2002, 5, 805–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Hagoort, P.; Jensen, O. Language prediction is reflected by coupling between frontal gamma and posterior alpha oscillations. J. Cogn. Neurosci. 2018, 30, 432–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, M.X.; David, N.; Vogeley, K.; Elger, C.E. Gamma-band activity in the human superior temporal sulcus during mentalizing from nonverbal social cues. Psychophysiology 2009, 46, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Van Diessen, E.; Senders, J.; Jansen, F.E.; Boersma, M.; Bruining, H. Increased power of resting-state gamma oscillations in autism spectrum disorder detected by routine electroencephalography. Eur. Arch. Psychiatry Clin. Neurosci. 2015, 265, 537–540. [Google Scholar] [CrossRef] [Green Version]
- Roopun, A.K.; Middleton, S.J.; Cunningham, M.O.; LeBeau, F.E.N.; Bibbig, A.; Whittington, M.A.; Traub, R.D. A beta2-frequency (20–30 Hz) oscillation in nonsynaptic networks of somatosensory cortex. Proc. Natl. Acad. Sci. USA 2006, 103, 15646–15650. [Google Scholar] [CrossRef] [Green Version]
- Murthy, V.N.; Fetz, E.E. Coherent 25-to 35-Hz oscillations in the sensorimotor cortex of awake behaving monkeys. Proc. Natl. Acad. Sci. USA 1992, 89, 5670–5674. [Google Scholar] [CrossRef] [Green Version]
- Canolty, R.T.; Cadieu, C.F.; Koepsell, K.; Ganguly, K.; Knight, R.T.; Carmena, J.M. Detecting event-related changes of multivariate phase coupling in dynamic brain networks. J. Neurophys. 2012, 107, 2020–2031. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.K.; Senkowski, D.; Schneider, T.R. The Neural Bases of Multisensory Processes; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2012. [Google Scholar]
- Pfurtscheller, G.; Stancak, A., Jr.; Edlinger, G. On the existence of different types of central beta rhythms below 30 Hz. Clin. Neurophysiol. 1997, 102, 316–325. [Google Scholar] [CrossRef]
- Kamiński, J.; Brzezicka, A.; Gola, M.; Wróbel, A. Beta band oscillations engagement in human alertness process. Int. J. Psychophysiol. 2012, 85, 125–128. [Google Scholar] [CrossRef]
- Miller, E.K.; Lundqvist, M.; Bastos, A.M. Working Memory 2.0. Neuron 2018, 100, 463–475. [Google Scholar] [CrossRef] [Green Version]
- Siegel, M.; Engel, A.K.; Donner, T.H. Cortical network dynamics of perceptual decision-making in the human brain. Front. Hum. Neurosci. 2011, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarland, D.J.; Miner, L.A.; Vaughan, T.M.; Wolpaw, J.R. Mu and beta rhythm topographies during motor imagery and actual movements. Brain Topogr. 2000, 12, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Von Stein, A.; Rappelsberger, P.; Sarnthein, J.; Petsche, H. Synchronization between temporal and parietal cortex during multimodal object processing in man. Cereb. Cortex 1999, 9, 137–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, A.K.; Fries, P. Beta-band oscillations—Signalling the status quo? Curr. Opin. Neurobiol. 2010, 20, 156–165. [Google Scholar] [CrossRef]
- Kononowicz, T.W.; Roger, C.; VanWassenhove, V. Temporal metacognition as the decoding of self-generated brain dynamics. Cereb. Cortex 2019, 29, 4366–4380. [Google Scholar] [CrossRef]
- Hanslmayr, S.; Spitzer, B.; Bäuml, K.H. Brain oscillations dissociate between semantic and nonsemantic encoding of episodic memories. Cereb. Cortex 2009, 19, 1631–1640. [Google Scholar] [CrossRef] [Green Version]
- Jensen, O.; Goel, P.; Kopell, N.; Pohja, M.; Hari, R.; Ermentrout, B. On the human sensorimotor-cortex beta rhythm: Sources and modeling. Neuroimage 2005, 26, 347–355. [Google Scholar] [CrossRef]
- Berger, H. Über das elektroenkephalogramm des menschen. Arch. Psychiatr. Nervenkrankh. 1929, 87, 527–570. [Google Scholar] [CrossRef]
- Katznelson, R.D. Chapter 6 in Nunez, P.L. Electric Fields of the Brain: The Neurophysics of EEG; Oxford University Press: New York, NY, USA, 1981. [Google Scholar]
- Knyazev, G.G. Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci. Biobehav. Rev. 2007, 31, 377–395. [Google Scholar] [CrossRef]
- Pfurtscheller, G. Induced oscillations in the alpha band: Functional meaning. Epilepsia 2003, 44, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Clark, J.M. Contributions of inhibitory mechanisms to unified theory in neuroscience and psychology. Brain Cogn. 1996, 30, 127–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, R.J. EEG measures of cerebral asymmetry: Conceptual and methodological issues. Int. J. Neurosci. 1988, 39, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Eysenck, H.J.; Eysenck, M.W. Personality and Individual Differences; Plenum: New York, NY, USA, 1987. [Google Scholar]
- Kline, J.P.; Blackhart, G.C.; Woodward, K.M.; Williams, S.R.; Schwartz, G.E. Anterior electroencephalographic asymmetry changes in elderly women in response to a pleasant and an unpleasant odour. Biol. Psychol. 2000, 52, 241–250. [Google Scholar] [CrossRef]
- Myers, L.B.; Brewin, C.R.; Winter, D.A. Repressive coping and self-reports of parenting. Br. J. Clin. Psychol. 1999, 38, 73–82. [Google Scholar] [CrossRef]
- Coan, J.A.; Allen, J.J.; Harmon-Jones, E. Voluntary facial expression and hemispheric asymmetry over the frontal cortex. Psychophysiology 2001, 38, 912–925. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.J.; Ekman, P.; Saron, C.D.; Senulis, J.A.; Friesen, W.V. Approach-withdrawal and cerebral asymmetry: Emotional expression and brain physiology: I. J. Pers. Soc. Psychol. 1990, 58, 330. [Google Scholar] [CrossRef]
- Gray, J.A. The Nature of Emotion: Fundamental Questions; Oxford University Press: Oxford, UK, 1994; pp. 329–331. [Google Scholar]
- Hewig, J.; Hagemann, D.; Seifert, J.; Naumann, E.; Bartussek, D. On the selective relation of frontal cortical asymmetry and anger-out versus anger-control. J. Pers. Soc. Psychol. 2004, 87, 926. [Google Scholar] [CrossRef]
- Harmon-Jones, E.; Allen, J.J. Behavioral activation sensitivity and resting frontal EEG asymmetry: Covariation of putative indicators related to risk for mood disorders. J. Abnorm. Psychol. 1997, 106, 159. [Google Scholar] [CrossRef]
- Heller, W. Neuropsychological mechanisms of individual differences in emotion, personality, and arousal. Neuropsychology 1993, 7, 476. [Google Scholar] [CrossRef]
- Zuckerman, M.; Kuhlman, D.M. Personality and risk-taking: Common biosocial factors. J. Pers. 2000, 68, 999–1029. [Google Scholar] [CrossRef]
- Mayer, J.D.; Salovey, P. The Intelligence of Emotional Intelligence; Elsevier: Amsterdam, The Netherlands, 1993. [Google Scholar]
- Mikolajczak, M.; Nelis, D.; Hansenne, M.; Quoidbach, J. If you can regulate sadness, you can probably regulate shame: Associations between trait emotional intelligence, emotion regulation and coping efficiency across discrete emotions. Pers. Individ. Differ. 2008, 44, 1356–1368. [Google Scholar] [CrossRef]
- Kemp, A.H.; Cooper, N.J.; Hermens, G.; Gordon, E.; Bryant, R.; Williams, L.M. Toward an integrated profile of emotional intelligence: Introducing a brief measure. J. Integr. Neurosci. 2005, 4, 41–61. [Google Scholar] [CrossRef] [PubMed]
- Reid, S.A.; Duke, L.M.; Allen, J.J. Resting frontal electroencephalographic asymmetry in depression: Inconsistencies suggest the need to identify mediating factors. Psychophysiology 1998, 35, 389–404. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G.; Logothetis, N.; Singer, W. Scaling brain size, keeping timing: Evolutionary preservation of brain rhythms. Neuron 2013, 80, 751–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramis, R.; Vanderwolf, C.H.; Bland, B.H. Two types of hippocampal rhythmical slow activity in both the rabbit and the rat: Relations to behavior and effects of atropine, diethyl ether, urethane, and pentobarbital. Exp. Neurol. 1975, 49, 58–85. [Google Scholar] [CrossRef]
- Lang, M.; Lang, W.; Diekmann, V.; Kornhuber, H.H. The frontal theta rhythm indicating motor and cognitive learning. Clin. Neurophysiol. Supplement. 1987, 40, 322–327. [Google Scholar]
- Buzsáki, G. Theta oscillations in the hippocampus. Neuron 2002, 33, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Berry, S.D.; Thompson, R.F. Prediction of learning rate from the hippocampal electroencephalogram. Science 1978, 200, 1298–1300. [Google Scholar] [CrossRef]
- Buzsáki, G. Theta rhythm of navigation: Link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 2005, 15, 827–840. [Google Scholar] [CrossRef] [Green Version]
- Knyazev, G.G.; Slobodskoy-Plusnin, J.Y. Substance use underlying behavior: Investigation of theta and high frequency oscillations in emotionally relevant situations. Clin. EEG Neurosci. 2009, 40, 1–4. [Google Scholar] [CrossRef]
- Krishnan, M.; Mariappan, M. EEG-based brain-machine interface (BMI) for controlling mobile robots: The trend of prior studies. J. Electron. 2015, 3, 159–165. [Google Scholar]
- Steriade, M.; Contreras, D.; Dossi, R.C.; Nunez, A. The slow (<1 Hz) oscillation in reticular thalamic and thalamocortical neurons: Scenario of sleep rhythm generation in interacting thalamic and neocortical networks. J. Neurosci. 1993, 13, 3284–3299. [Google Scholar] [PubMed] [Green Version]
- Michel, C.M.; Henggeler, B.; Brandeis, D.; Lehmann, D. Localization of sources of brain alpha/theta/delta activity and the influence of the mode of spontaneous mentation. Physiol. Measur. 1993, 14, A21. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, M.; Langhorst, P. Simultaneous changes of rhythmic organization in brainstem neurons, respiration, cardiovascular system and EEG between 0.05 Hz and 0.5 Hz. J. Auton. Nerv. Syst. 1998, 68, 58–77. [Google Scholar] [CrossRef]
- Leung, L.S.; Yim, C.C. Rhythmic delta-frequency activities in the nucleus accumbens of anesthetized and freely moving rats. Can. J. Phisiol. Pharmacol. 1993, 71, 311–320. [Google Scholar] [CrossRef]
- Grace, A.A. The tonic/phasic model of dopamine system regulation: Its relevance for understanding how stimulant abuse can alter basal ganglia function. Drug Alcohol Depend. 1995, 37, 111–129. [Google Scholar] [CrossRef]
- Berridge, K.C.; Robinson, T.E.; Aldridge, J.W. Dissecting components of reward: ‘liking’, ‘wanting’, and learning. Curr. Opin. Pharmacol. 2009, 9, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.A. The Neuropsychology of Anxiety; Clarendon Press/Oxford University Press: Oxford, UK, 1982. [Google Scholar]
- Eysenck, H.J. Biological Dimensions of Personality; The Guilford Press: New York, NY, USA, 1990. [Google Scholar]
- Spielberger, C.D.; Sydeman, S.J.; Owen, A.E.; Marsh, B.J. Measuring Anxiety and Anger with the State-Trait. Anxiety Inventory (STAI) and the State-Trait. Anger Expression Inventory (STAXI); Lawrence Erlbaum Associates Publishers: Mahwah, NJ, 1999. [Google Scholar]
- Robinson, D.L. Properties of the diffuse thalamocortical system and human personality: A direct test of Pavlovian/Eysenckian theory. Pers. Individ. Differ. 1982, 3, 1–16. [Google Scholar] [CrossRef]
- Depue, R.A.; Collins, P.F. Neurobiology of the structure of personality: Dopamine, facilitation of incentive motivation, and extraversion. Behav. Brain Sci. 1999, 22, 491–517. [Google Scholar] [CrossRef]
- Sawyer, D.A.; Julia, H.L.; Turin, A.C. Caffeine and human behavior: Arousal, anxiety, and performance effects. J. Behav. Med. 1982, 5, 415–439. [Google Scholar] [CrossRef]
- Gibbs, F.A.; Gibbs, E.L.; Lennox, W.G. Effect on the electro-encephalogram of certain drugs which influence nervous activity. Arch. Intern. Med. 1937, 60, 154–166. [Google Scholar] [CrossRef]
- Buzsáki, G.; Watson, B.O. Brain rhythms and neural syntax: Implications for efficient coding of cognitive content and neuropsychiatric disease. Dialogues Clin. Neurosci. 2012, 14, 345. [Google Scholar] [PubMed]
- Gasser, T.; Bächer, P.; Steinberg, H. Test-retest reliability of spectral parameters of the EEG. Clin. Neurophysiol. 1985, 60, 312–319. [Google Scholar] [CrossRef]
- Hagemann, D.; Naumann, E.; Becker, G.; Maier, S.; Bartussek, D. Frontal brain asymmetry and affective style: A conceptual replication. Psychophysiology 1998, 35, 372–388. [Google Scholar] [CrossRef]
- Brodal, P. The Central Nervous System: Structure and Function; Oxford University Press: Oxford, UK, 1992; pp. 246–261. [Google Scholar]
- Verleger, R. The instruction to refrain from blinking affects auditory P3 and N1 amplitudes. Clin. Neurophysiol. 1991, 78, 240–251. [Google Scholar] [CrossRef]
- Hagemann, D.; Naumann, E.; Thayer, J.F. The quest for the EEG reference revisited: A glance from brain asymmetry research. Psychophysiology 2001, 38, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Melrose, S. Seasonal affective disorder: An overview of assessment and treatment approaches. Depress. Res. Treat. 2015. [Google Scholar] [CrossRef] [Green Version]
- King, J.A.; Rosal, M.C.; Ma, Y.; Reed, G.; Kelly, T.A.; Ockene, I.S. Sequence and seasonal effects of salivary cortisol. J. Behav. Med. 2000, 26, 67–73. [Google Scholar] [CrossRef]
- Coan, J.A.; Allen, J.J.; McKnight, P.E. A capability model of individual differences in frontal EEG asymmetry. Biol. Psychol. 2006, 72, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J. An abilities conception of personality: Some implications for personality measurement. Am. Psychol. 1966, 21, 132. [Google Scholar] [CrossRef]
- Cloninger, C.R.; Przybeck, T.R.; Svrakic, D.M. The tridimensional personality questionnaire: US normative data. Psychol. Rep. 1991, 69, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PRISMA flow chart of the selection process.

Figure 2.
Risk of bias for each domain considered in the selected studies.


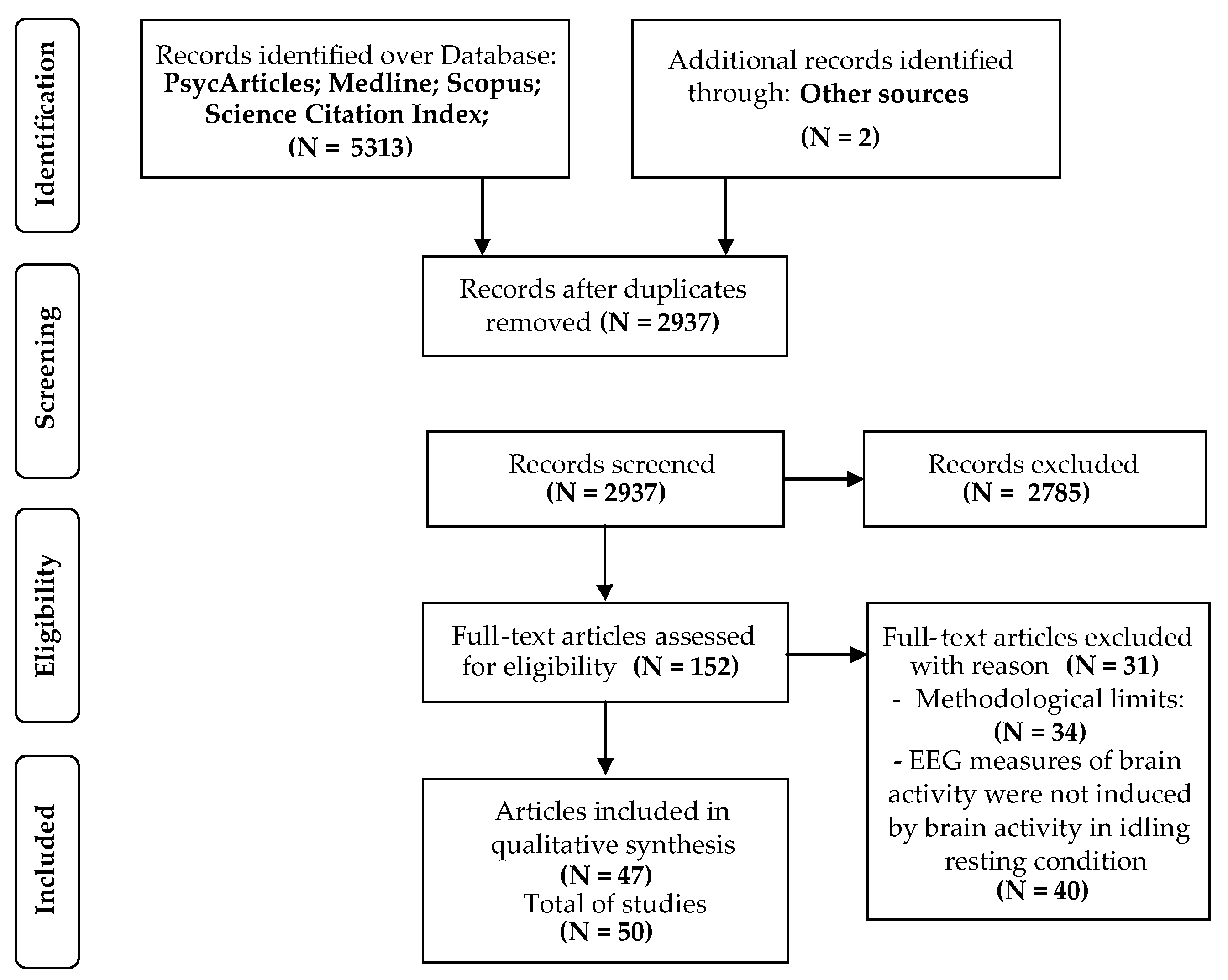{kind=link}
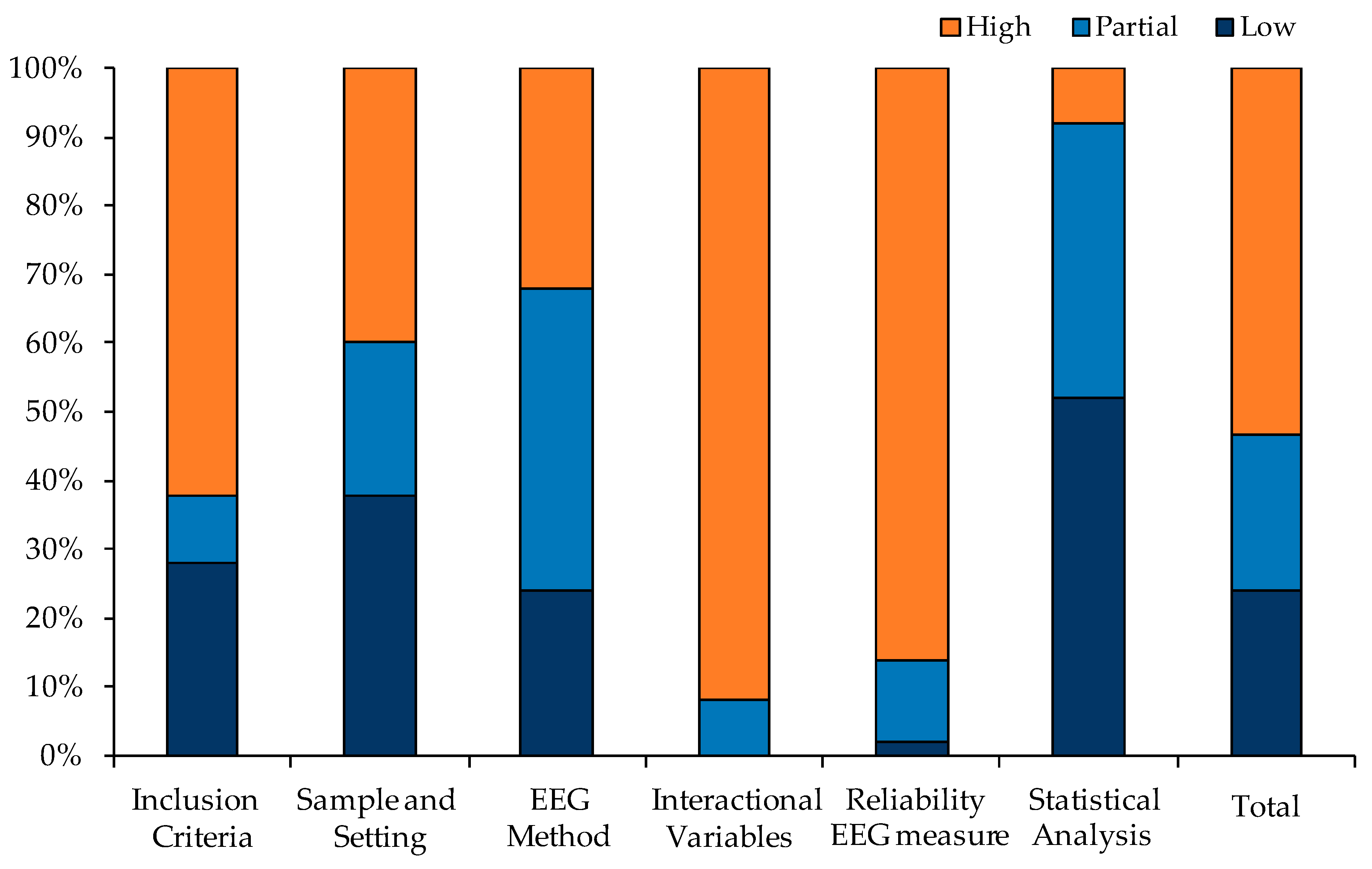{kind=link}
Table 1.
Data selected for each study.
| Participants | Personality Domain | Electrophysiological Domain | Bias | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Authors | N | Age Mean (SD) | Sex Men % | Self-Report | Trait/s Considered | Resting State Recording Condition (EEG Reference, Length) | Results and Indexes of Brain Activation | EEG Test–Retest | Risk % | |
| Tomarken et al., 1992 [26] | 90 | 19 (-) | 0 | EHI a PANAS b AIM c | Handedness Positive Affect (PA), Negative Affect (NA) Intensity of individual experiences emotion | Eight of 1-min baseline, balanced for four opened (O) and four closed (C)-eyes order, (O-C-C-O-C-O-O-C and C-O-O-C-O-C-C-O), participants, and sessions. Online ref. linked CZ site. Offline re-ref. average earlobes. | Left anterior α-asymmetry is positively related to PA, and negatively related to NA. This pattern of EEG activation is stable over time. | Two measures 2-after 3 weeks | 58.4 | |
| Tomarken and Davidson, 1994 [27] | 90 | 19 (-) | 0 | EHI a MCSD d STAI e BDI f | Handedness Defensiveness Trait Anxiety (TA) Depression Trait (TD) | 8 of 1-min baseline, balanced for 4 O and 4 C eyes order, (O-C-C-O-C-O-O-C and C-O-O-C-O-C-C-O), participants, and sessions. Online ref. linked CZ site. Offline re-ref. average earlobes. | The α activity in mid-frontal and left lateral frontal sites is related to a higher level of defensiveness than a lower level. | Two measures 2-after 3 weeks | 58.4 | |
| Sutton and Davidson, 1997 [28] | 46 | 20 (-) | 50 | EHI a PANAS b BIS/BAS Scale g | Handedness PA, NA Behavioral Inhibition System (BIS), Behavioral Approach System (BAS) | Eight of 1-min baseline, balanced for four O and four C eyes order, participants, and sessions. Online ref. linked earlobes. Offline re-ref. average earlobes. | Resting prefrontal activation is an index of EEG α-asymmetry related to Gray’s BAS and BIS rather than PA and NA. | Two measures two after 3 months | 66.7 | |
| Hagemann et al., 1999 [29] | 36 | 23.5 (4.0) | 36 | EHI a PANAS b EPQ h | Handedness PA, NA E, N | Eight of 1-min baseline, balanced for four O and four C eyes order (O-C-C-O-C-O-O or C-O-O-CO-C-CO), and participants. Online ref. linked mastoids. Offline re-ref. CZ site. | Lateral α-asymmetry in resting EEG is related to NA but is not significantly related to PA. | One Measure | 50.0 | |
| Kline et al., 2001 [30] | 72 | 24.4 (4.6) | 42 | EHI a EPQ h HPCS i MCSD d | Handedness Neuroticism (N), Psychoticism (P), Extraversion (E), Lie (L) Maternal and Paternal caring Defensiveness | Six of 1-min baseline, balanced for three O and three C eyes order, and participants. Online ref. linked earlobes. | Defensiveness and perceived maternal (but not paternal) caring, separately predict left lateral frontal activation of the EEG α-asymmetry. | One Measure | 66.7 | |
| Kline et al., 2002 [31] | 235 | 20.4 (4.1) | 67 | EHI a EPQ h | Handedness N, E, P, L | Six of 1-min baseline, three O eyes, three C eyes. Online ref. linked earlobes. | Defensiveness is related with right frontal α-activity, in the presence of opposite-sex experimenters but not same-sex. | One Measure | 58.4 | |
| Blackhart et al., 2002 [32] | 77 | 20.5 (3.7) | 47 | EHI a SAM j | Handedness Pleasure–Displeasure, Arousal–Non arousal, Dominance–Submissive. | Six of 1-min baseline, three O eyes, three C eyes. Online ref. linked earlobes. | Negative mood post-preparation, predicts left frontal α-asymmetry during resting EEG, in women. | One Measure | 66.7 | |
| Coan and Allen, 2003 [33] | 32 | 20.5 (-) | 28 | BIS/BAS Scale g | BIS, BAS | Eight of 1-min baseline balanced for four O and four C eyes order. Online ref. Cz site. Offline re-ref. linked mastoids. Offline re-ref. average brain activity. | Resting EEG left frontal -asymmetry is related to greater BAS scores; higher BIS scores are related to right frontal activity in the mid-frontal region, only. | One Measure | 83.4 | |
| Knyazev and Slobodskaya, 2003 [34] | 47 | 23.9 (5.3) | 13 | GWPQ k EPQ h STAI e | BIS, BAS E, N, P TA | Six of 60-s baseline, balanced for a pseudo-random O/C-eyes order, and participants. Online ref. linked earlobes. | Higher δ–α EEG frequency oscillations coupling, are related to increased behavioral inhibition. | One Measure | 75.0 | |
| Razoumnikova, 2003 [35] | 46 | 18 (-) | 100 | EPQ h AIS m KTI l | E, N, P Emotional Intelligence: Verbal, Numerical, Figural Sensation–Intuition (S), Thinking–Feeling (T), Judging–Perceiving (J) | 3 min resting period with C eyes. Online ref. linked earlobes. | EEG rhythms are relating to IQ, S, E, and N. Cognitive abilities in higher IQ subjects are related with the cortex connectivity, and θ1–θ2–α1 EEG rhythms. The β2 rhythm is predominant in right hemisphere. | One Measure | 83.4 | |
| Minnix and Kline, 2004 [36] | 140 | 20.4 (4.1) | 30 | EHI a EPQ h | Handedness N, E, P, L | Six of 1-min baseline, three O eyes, three C eyes. Online ref. linked earlobes. | EEG Resting Mid-Frontal α–band activity is related with higher N. | One Measure | 75.0 | |
| Hewig et al., 2006 [37] | 59 | 24 (3.3) | 49 | EHI a BIS/BAS Scale g EPQ h | Handedness BIS, BAS E, N | 12 of 1-min baseline, six O and six C eyes, balanced for two orders (OCCO-COOC-OCCO; COOC-OCCO-COOC) and participants. Online ref. linked CZ. | There is no relation between resting frontal α–activity of EEG, and BIS/BAS traits. | Four measures one every 4 weeks | 58.4 | |
| Tran et al., 2006 [38] | 699 | 36 (-) | 49 | NEO-FFI n | E, N, Openness (O), Agreeableness (A), Consciousness (C). | 3 min resting period with C eyes. Offline re-ref. linked mastoids. | N scores are higher in women than men. In both, δ–θ activity is related to E and C, while, α–β activity is related to NEO–FFI traits in males. | One Measure | 75.0 | |
| Jaušovec and Jaušovec, 2007 [39] | 110 | 21 (-) | 50 | NEO-FFI n MSCEIT o | N, E, O, A, C Emotional Intelligence: Verbal, Performance, Experiential–Emotional, Strategic–Emotional | 5 min resting period with C eyes and 5 min resting period with O eyes. Repeating O/C eyes order after each minute of recording. Online ref. linked mastoids. | Women with extreme traits levels differ in the γ band activity than males; vice-versa in the α1 band. Brain activity increased in parietal-occipital areas rather in frontal. | One Measure | 66.7 | |
| Smit et al., 2007 [40] | Young Mid-Aged | 381 380 | 26.2 (4.1) 49.4 (7.2) | - - | No specified STAI e YARS p ABV q | Handedness TA Depression Trait (TD) N, Somatic-Anxiety | 3 min resting period with C eyes. Online ref. linked earlobes. | Frontal α-Asymmetry (FA) in rest-EEG, is heritable in young adults but not in middle-aged. FA is related to the risk of anxiety and depression in young women. | One Measure | 83.4 |
| Fleck et al., 2008 [41] | 34 | 22.4 (4.7) | 47 | RTS r | Transliminality (Tr) | 3.5 min resting period with C eyes and 3.5 min resting period with O eyes. Online ref. linked mastoids. | Higher Tr scores related with lower α1, β1, γ1 activity in left posterior cortex, and lower α2, β1, γ activity in right superior temporal areas, than lower scores. Lower levels of Tr, related to increased γ in the mid-frontal areas, than higher. | One Measure | 83.4 | |
| Gatt et al., 2008 [42] | 117 | 36.9 (12.6) | 38 | DASS-21 s | TD | 2 min resting period with C eyes followed by 2 min O eyes. Offline re-ref. linked earlobes. | BDNF M/M genotype is a predictor of working memory activity. The EEG α-power mediates the effect of this phenotype on TD expression. | One Measure | 66.7 | |
| Kline and Allen, 2008 [43] | 71 | 18.9 (1.5) | 30 | EHI a EPQ h MCSD d BDI f TMAS ee | Handedness N, E, P, L Defensiveness TD TA | Six of 1-min baseline, balanced for 3 O eyes and 3 C eyes orders (O–C–C–O– C–O; C–O–O–C–O–C). Online ref. linked earlobes. Offline re-ref. average brain activity. | In the context of increased α-band activity in the right lateral frontal asymmetry EPQL traits are positively related to BDI. | Two measures two after 3 weeks | 41.7 | |
| Mathersul et al., 2008 [44] | 428 | 34.9 (12.6) | 50 | No specified DASS-21 s | Handedness TD, TA | 2 min resting period with O eyes and 2 min with C eyes. Offline re-ref. average brain activity. | An increased α-asymmetry in the right parietal-temporal areas related to higher anxiety comorbidity and depression trait. Right parietal-temporal α-activity related to anxiety apprehension. | One Measure | 58.4 | |
| Santesso et al., 2008 [45] | Study1 Study2 | 37 44 | 19.5 (1.4) 18.6 (0.7) | 24 100 | EHI a SSS-V u FIS v | Handedness Sensation Seeking Risk-taking behaviors | 1 min resting period with O eyes and 1 min with C eyes. Online ref. linked CZ. Offline re-ref. average brain activity. | Sensation seeking is associated with a greater left frontal -asymmetry in resting EEG. | One Meas. One Meas. | 66.7 66.7 |
| Schutter et al., 2008 [8] | 24 | 22.2 (2.0) | 46 | BIS/BAS Scale g | BIS, BAS | Four of 1-min baseline balanced for two O and two C eyes order (O–C–O–C). Online ref. voltage. Offline re-ref. CZ. | Β-asymmetry in resting EEG reflects approach–avoidance motivational predispositions. | One Measure | 41.7 | |
| Master et al., 2009 [46] | 46 | 20.1 (1.5) | 43 | No specified EAC w BIS/BAS Scale g | Handedness Emotional Approach BIS, BAS | Eight of 1-min baseline balanced for four O and four C eyes order. Online ref. Left-lobe. Offline re-ref. lobes average. | Emotional expression is significantly related to greater left-sided frontal A-asymmetry in the rest-EEG state. | One Measure | 58.4 | |
| Pavlenko et al., 2009 [47] | 111 | 21.5 (-) | 42 | STAI e Cattell 16PFx | State and Trait Anxiety A, B, C, E, F, G, H, I, L, M, N, O, Q1, Q2, Q3, Q4.+ | Two of 1-min baseline, one with O eyes and one with C eyes. Online ref. linked an active electrode n.a. | State anxiety is positively related to β2 spectral power density in the temporal occipital areas of the right hemisphere (O-eyes). Trait anxiety is positively related to β1, β2. | One Measure | 75.0 | |
| Peterson and Harmon-Jones, 2009 [48] | Study1 Study2 | 72 65 | Stud. (-) Stud. (-) | - 58 | IAS-R y Time Variables BIS/BAS Scale g | Nurturance and Dominance Day Time, Year season BIS, BAS | Four of 1-min baseline balanced for two O and two C eyes order (O–C–C–O or C–O–O–C) and participants. Online ref. eft-earlobe. Offline re-ref. earlobes average. | Frontal α-asymmetry in resting EEG reflects circadian and seasonal influences. The right frontal activity is increased during fall mornings. | One Measure One Measure | 66.7 50.0 |
| Knyazev et al., 2010 [49] | 132 | 27.5 (-) | 39 | EPQ h GWPQ k STAI e | N BIS TA | 3-min rest period with C eyes, 3- min with O eyes balanced for O/C-eyes order. Online ref. lobes. | Increased coupling δ–β frequency oscillations is related with state anxiety. | One Measure | 75.0 | |
| Mikolajczak et al., 2010 [50] | 31 | 22.4 (3.8) | 20 | TEI–Que z | Well-Being, Self-Control, Emotionality, Sociability | Eight of 1-min baseline balanced for four O and four C eyes order. Online ref. linked earlobe. | Resting EEG α–frontal asymmetry is related to emotional intelligence. | One Measure | 83.3 | |
| Wacker and Gatt, 2010 [51] | 1093 | 39.3 (18.7) | 46 | NEO–FFI n | N, E, O, A, C | 2-min resting period with O eyes followed by 2-min C eyes. Online ref. linked mastoids. | The δ–θ activity in posterior vs. frontal areas related to E and COMT VAL-158 MET polymorphism. | One Measure | 66.7 | |
| Wacker et al., 2010 [52] | Study3 Study4 | 112 35 | 23.8 (3.4) 22.9 (3.4) | 100 62 | ARES gg BIS/BAS Scale g | BAS, BIS BAS, BIS | Five for 90-s of resting period with C eyes. Online ref. linked CZ site. Offline re-ref. linked mastoids. | Resting posterior vs. frontal δ–θ EEG activity, is related to BAS/AE but BAS/AE, not related to frontal α -asymmetry. | Three meas. two after 1 year, three n.a. One Meas. | 66.7 91.7 |
| Chavanon et al., 2011 [53] | 78 | 23.2 (2.7) | 100 | MPQ–NE EPQ h | Reactivity, Alienation, and Aggression E, N, P | Five for 1-min resting period with C eyes. Online ref. linked CZ site. Offline re-ref. avrage brain activity. | Rostral anterior cingulate activity generates the posterior vs. anterior θ activity, related to AE. | One Measure | 66.7 | |
| Koehler et al., 2011 [54] | 141 | 27.8 (-) | 52 | NEO–PI–R nn | N, E, O, A, C | 5-min rest period with closed eyes. Online ref. linked between Fz-Cz sites. Offline re-ref. average brain activity. | Resting posterior minus frontal EEG δ–θ activity is related with E and DRD2 genotype. | One Measure | 83.4 | |
| Konareva, 2011 [55] | 118 | 27 (-) | - | Cattell 16 PF x EPQ h STI xx OST xxx | A, B, C, E, F, G, H, I, L, M, N, O, Q1, Q2, Q3, Q4.+ E, N, P Objective plasticity, Social ergoicity Ergonicity, Plasticity, Tempo, Emotionality. | n.a. resting period with O eyes followed by n.a. resting period with C eyes. Online ref. n.a. Offline ref n.a. | Psychological traits of personality modulated the activity of δ and θ EEG rhythms. | One Measure | 91.7 | |
| Knyazev et al., 2012 [56] | 88 | 24.5 (-) | 51 | Self-report ad hoc EPP hh | Positive emotions, mood state E, N, P | 3-min rest period with C eyes and 3-min with O eyes balanced for O/C-eyes order. Online ref. linked lobes. | AE related to higher θ activity in the posterior default mode network and lower θ in the orbitofrontal cortex. | One Measure | 66.7 | |
| De Pascalis et al., 2013 [57] | 51 | 24.1 (3.7) | 0 | EHI a BIS/BAS Scale g LOT–R t PANAS b | Handedness BIS, BAS Optimism PA, NA | Four of 1-min baseline balanced for two O and two C eyes order and participants. Online ref. linked CZ site. Offline re-ref linked earlobes. | Higher BAS scores are related to greater left-sided α-activity in the BA11 area. Optimism is related to higher activity in the BA10 and BA31 areas. | One Measure | 42.8 | |
| Alessandri et al., 2015 [58] | 51 | 24.1 (3.7) | 0 | EHI a GSS gs LOT–R t LSS ls P–OR tt | Handedness Self-esteem Optimism Life Satisfaction Positive orientation | Four of 1-min baseline balanced for two O and two C eyes order and participants. Online ref. linked CZ site. Offline re-ref linked earlobes. | The α-asymmetry in the posterior cingulate cortex is uniquely associated with both positivity scores. | One Measure | 42.8 | |
| Gable et al., 2015 [59] | 126 | Stud. (-) | 46 | Self-report ad hoc PUM ttt BIS/BAS Scale g | Handedness Positive Urgency BIS, BAS | Eight of 1-min baseline, balanced among four O and four C eyes order (C-O-O-C-O-C-C-O or O-C-C-O-C-O-O-C) and participants. Online ref. Left-lobe. Offline re-ref. average brain activity. | BAS trait related to greater left-frontal α-asymmetry, while BIS trait is related to greater right-frontal α-asymmetry. | One Measure | 75.0 | |
| Huang et al., 2015 [60] | 94 | 26.5 (-) | 0 | CC-scale aa EPQ h | Handedness E, N, P, L | 16 of 15-s baseline, eight O and eight C eyes balanced for two groups (O–C–O–C–O–C–O–C and C–O–C–O–C–O–C–O). Online ref. linked mastoids. Offline re-ref. CZ. | Women with higher N scores, exhibit lower-left prefrontal α1– αTotal– asymmetry, than women with lower N during the mid-late luteal phase. | Three measures two after 11–13 days three after 22–24 days | 41.7 | |
| Korjus et al., 2015 [61] | 289 | 22.0 (3.6) | 35 | NEO-PI-R nn | N, E, O, A, C | 1-min; two of 1-min and three of 1-min baseline one O eyes and one C eyes balanced for O/C order. Online ref. linked lobes. Offline re-ref. lobes. | Personality traits cannot be predicted from the power of resting-state EEG. | One Measure | 75.0 | |
| Tullett et al., 2015 [62] | 56 | 19.8 (3.8) | 46 | NI ni SNS ns BFQ nnn | Degree of Nostalgia Frequency of Nostalgia N, E, O, A, C | Eight of 30-s baseline, balanced for four O and four C eyes order. Online ref. digital average. Offline re-ref. lobes. | The right-frontal α-asymmetry predicts an increased proneness to nostalgia scores. | One Measure | 75.0 | |
| Adolph and Margraf, 2017 [63] | 37 | 24.2 (3.8) | 51 | No specified STAI e D–S ff | Handedness TA TD | 16 of 1-min baseline, balanced for eight O and eight C eyes order, and participants (O–C–C–O–C–O–O–C or C–O–O–C–O–C–C–O). Online ref. left mastoid. Offline re-ref. mastoids. | Depression and anxiety are both significantly related to larger right frontal α-asymmetry. | One Measure | 50.0 | |
| Neal and Gable, 2017 [64] | 182 | Stud. (-) | 40 | Checklist ad hoc UPPS-P imp BIS/BAS Scale g | Handedness Impulsivity (Imp) BIS, BAS | Eight of 1-min baseline, balanced for four O and four C eyes order. Online ref. linked left earlobe. Offline re-ref. to earlobes. | Regulatory control and impulsivity are related to BIS. Withdrawal is related to FFFS, and right frontal A-asymmetry. | One Measure | 75.0 | |
| De Groot and Van-Strien, 2018 [65] | 93 | 21.03 (2.4) | 39 | AQ aq | Autistic Traits | 3 min rest period with C eyes, 3 min with O eyes balanced for O/C order. Online ref. linked mastoids. Offline re-ref. to mastoids average. | Spontaneous γ1–γ2 oscillations during resting-state are not predictive of autistic traits in the general population. | One Measure | 58.3 | |
| De Pascalis et al., 2018 [66] | 162 | 23.6 (2.4) | 0 | EHI a RST–PQ kk | Handedness BIS, BAS, FFFS | 4-min rest period with C eyes, 4-min with O eyes balanced among O/C order and participants. Online ref. linked left earlobe. Offline re-ref. to earlobes. | In the subgroup with a young same-sex experimenter, BAS-impulsivity is related to a greater left-frontal α-activity, and FFFS. BIS trait, not related to the greater right-frontal-central activity. | One Measure | 33.4 | |
| Threadgill and Gable, 2018 [67] | 125 | Stud. (-) | - | EHI a BIS/BAS Scale g UPPS-P imp | Handedness BAS, FFFS Impulsivity (Impu) | 4 min rest period with C eyes, 4 min with O eyes balanced between participants. Online ref. linked left earlobe. Offline re-ref. average brain activity. | BAS trait is negatively related to resting β activity. Impu. is related to greater resting β activity. Lower levels of β activity in the motor cortex related to motivational traits. | One Measure | 75.0 | |
| Paban et al., 2019 [68] | 45 | 34.7 (9.1) | 51 | CD–RISC rt | Resilience Trait | 10 min rest period with C eyes. Online ref. individual vertex site. | Resilience is related to resting EEG source-space brain network flexibility. | One Measure | 75.0 | |
| De Pascalis et al., 2020 [69] | 125 | 22.8 (2.8) | 39 | No specified RST–PQkk STAI–State e | Handedness BIS, BAS, Fight–Flight–Freeze System (FFFS) State Anxiety | 3 min rest period with C eyes, 3 min with O eyes balanced among O/C eyes order and participants. Online ref. linked earlobes. Offline re-ref. average brain activity. | BIS trait related to higher δ–β coupling in Anxiety group, and high δ–θ coupling in Relaxation group. In both groups, BAS-GDP is positively related to higher δ–γ coupling. | One Measure | 41.7 | |
| Zanesco et al., 2020 [70] | 187 | 36.1 (20.1) | 64 | BIS/BAS Scale g NEO–FFI p MDMQ ww | BIS, BAS N, E, O, A, C Current Mood state | 16 of 60-s baseline, balanced for eight O, and eight C eyes order and participants. Online ref. linked FCz site. Offline re-ref. average brain activity. | Personality traits can be reflected in microstate dynamics present during periods of quiet rest. | One Measure | 58.4 | |
| Zhang et al., 2020 [71] | 80 | 21.49 (2.5) | 55 | DERS ss STAI–State e STAI–Trait e BDI f | Emotional-Dysregulation State Anxiety TA TD | 12 of 15-s baseline, six O eyes, six C eyes balanced for O/C eyes order and participants. Online ref. linked left mastoid. Offline re-ref. average of mastoids. | The resting-state frontal α-asymmetry predicts the regulation of emotion, mainly in impulse control. | One Measure | 58.4 | |
a Edinburgh Handedness Inventory [72]; aa Chapman and Chapman’s scale [73];aq Autism-Spectrum Quotient [74];b Positive and Negative Affect Schedule [75]; c Affect Intensity Measure [76]; d Marlowe–Crowne Social Desiderability Scale [77]; e Spielberger State Trait Anxiety Inventory [78]; ee Taylor Manifest Anxiety Scale [79]; f Beck Depression Inventory [80]; ff Depressions Scale [81]; g BIS/BAS scales [82]; gg Action Regulation Emotion Systems [83];gs General Self-Esteem Scale [84]; h Eysenck Personality Questionnaire [85]; i Harvard Parental Caring Scale [86]; j Self-Assessment Manikin [87]; k Gray–Wilson Personality Questionnaire [88]; kk Reinforcement Sensitivity Theory of Personality Questionnaire [89]; imp Impulsive Behavior Scale [90]; l Keirser’s Type Inventory [91]; ls Satisfaction with Life Scale [92]; m Amthauer’s Intelligence Scale [93]; n NEO-Five-Factor Inventory [94]; nn Revised NEO Personality Inventory [95]; nnn Big Five Inventory [96]; ni Nostalgia Inventory [97]; ns Southampton Nostalgia Scale [98]; o Mayer–Salovey–Caruso Emotional Intelligence Test [99]; p Young Adult Self Report scale [100]; q AmsterdamseBiografischeVragenlijst [101]; r Revised Transliminality Scale [102]; rt Connor–Davidson Resilience Scale [103]; s Shortened version of the Depression Anxiety Stress Scale [104]; ssDifficulties in Emotion Regulation Scale [105]; t Life Orientation Test Revised (LOT-R, [106]); tt Positive Orientation Scale [107];ttt Positive Urgency Measure [108]; u Sensation Seeking Scale Form V [109]; v Frequency of Involvement Scale [110] w Emotional Approach Coping [111];ww Multidimensional Mood State Questionnaire [112]; x Cattel’s technique 16 Personality Factor: + Warmth (A), Reasoning (B), Emotional Stability (C), Dominance (E), Liveliness (F), Rule-Consciousness (G), Social Boldness (H), Sensitivity (I), Vigilance (L), Abstractedness (M), Privateness (N), Apprehension (O), Openness to Change (Q1), Self-Reliance (Q2), Perfectionism (Q3), Tension (Q4) [113]; xx Strelau Temperament Inventory [114]; xxx Rusalov Structure of Temperament Questionnaire [115]; y Interpersonal Adjective Scales (IAS-R, [116]); z Trait Emotional Intelligence Questionnaire [117].
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vecchio, A.; De Pascalis, V. EEG Resting Asymmetries and Frequency Oscillations in Approach/Avoidance Personality Traits: A Systematic Review. Symmetry 2020, 12, 1712. https://doi.org/10.3390/sym12101712
AMA Style
Vecchio A, De Pascalis V. EEG Resting Asymmetries and Frequency Oscillations in Approach/Avoidance Personality Traits: A Systematic Review. Symmetry. 2020; 12(10):1712. https://doi.org/10.3390/sym12101712
Chicago/Turabian StyleVecchio, Arianna, and Vilfredo De Pascalis. 2020. "EEG Resting Asymmetries and Frequency Oscillations in Approach/Avoidance Personality Traits: A Systematic Review" Symmetry 12, no. 10: 1712. https://doi.org/10.3390/sym12101712
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.