

Antibacterial Activity of Ferulic Acid Ester against Ralstonia solanacearum and Its Synergy with Essential Oils

1
School of Biotechnology, Jiangsu University of Science and Technology, Zhenjiang 212018, China
2
School of Pharmacy, Nanjing University of Chinese Medicine Hanlin College, Taizhou 225300, China
3
Key Laboratory of Silkworm and Mulberry Genetic Improvement, Ministry of Agricultural and Rural Affairs, Sericultural Research Institute, Chinese Academy of Agricultural Sciences, Zhenjiang 212018, China
*
Author to whom correspondence should be addressed.
Sustainability 2022, 14(24), 16348; https://doi.org/10.3390/su142416348
Submission received: 25 September 2022
/
Revised: 16 November 2022
/
Accepted: 25 November 2022
/
Published: 7 December 2022
(This article belongs to the Special Issue Recent Advances in Biomass Conversion, Renewable Energy, and Biomaterials)
Abstract
:Ralstonia solanacearum is one of the ten most harmful plant bacteria worldwide, and traditional agrochemicals are not very effective in controlling this pathogen. Moreover, excessive pesticides always bring organic residues and resistant strains, which cause the unsustainability of the environment. In this paper, ferulic acid and essential oils are used as antibacterial materials. These compounds are natural substances with low toxicity and environmental safety. Through the structural optimization and the analysis of binary combined bacteriostatic efficiency, the MIC values of chlorobutyl ferulate (2e) and peppermint essential oil (EO1) were 0.64 mg/mL and 2.02 mg/mL, respectively, and the MIC value of 2e-EO1 (mass ratio 1:1.5) was 0.40 mg/mL. The growth rate of bacteria treated with 2e-EO1 was inhibited, the OD590nm value of cell membrane decreased by 57.83%, and the expression levels of hrpB, pehC, pilT, polA, aceE, egl, and phcA were downregulated to 18.81%, 30.50%, 14.00%, 44.90%, 86.79%, 23.90%, and 27.56%, respectively. The results showed that 2e-EO1 had a synergistic inhibitory effect against R. solanacearum. It significantly affected the formation of the bacterial cell membrane and the expression of pathogenic genes. Consequently, 2e-EO1 provides the potential to become a green pesticide and can promote the sustainability of the agricultural ecological environment.
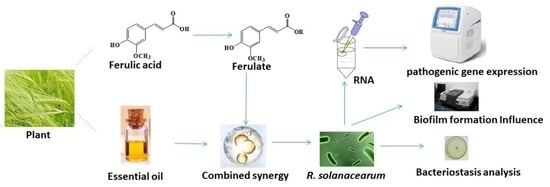
1. Introduction
The sustainability of agriculture is related to the development of the national economy and society’s stability. By 2050, the crop output will be far lower than the needs of the growing population [1]. Plant diseases are the main factors affecting crop losses, reducing the yield by 14% [2]. It is considered that soil-borne diseases are more restrictive than seed-borne or air-borne diseases, accounting for 10–20% of the annual yield loss [3]. Ralstonia solanacearum is one of the ten most harmful plant bacteria and is distributed all over the world, especially in subtropical and tropical regions [4]. This soil-borne bacterium can infect more than 450 plant species of 54 botanical families [5] and cause huge direct economic losses every year [6]. The pathogenic genes of R. solanacearum have coevolved with the external environment and are characterized by multiple toxic factors [7]. Difficulties are associated with controlling this pathogen due to its ability to grow endophytically, its relationship with weeds, and its survival in soil [6]. Therefore, it is of great significance to research how to effectively inhibit R. solanacearum for agricultural sustainability.
Generally, a 1% increase in crop output per hectare is associated with a 1.8% increase in pesticide use per hectare [8]. However, it is not significant for many traditional agrochemicals against R. solanacearum [9]. Moreover, excessive chemical pesticides always bring organic residues and the production of resistant strains [10], which cause the unsustainability of the environment. Several effective fumigants are restricted due to their serious environmental hazards, such as metam sodium and chloropicrin. It is reported that many plant residues derived from, e.g., chili [11], Chinese gall [12], and clove [13], have antimicrobial activities and the indirect suppression of pathogens through improved soil properties [14]. Biopesticides have emerged as a sustainable alternative leading to safe organic farming. At the global level, the environment-friendliness and target-specificity of biopesticides are gaining wide popularity [15].
Ferulic acid (1) and Essential oils (EO) are common natural metabolites with diverse bioactivities [16]. So far, there have been many reports on the effect of these substances and their analogs against R. solanacearum. Endophytes from the roots of Solanum surattense secrete a vast array of bioactive metabolites containing 1 to protect host plants against R. solanacearum [17]. Lansiumamide B, isolated from the seeds of Clausena lansium, the minimum inhibitory concentration (MIC) of which is 0.13 mg/mL, and the control efficiency against tobacco bacterial wilt is nearly 40 times higher than streptomycin [18]. EOs contain a variety of chemicals that have inhibitory activity against R. solanacearum [19]. Thyme oil, marjoram oil, and caraway oil have a significant antibacterial effect against R. solanacearum of tomato. Under field experiments, thyme oil treatment against tomato bacterial wilt reduces it by 94.8% [20]. Bacterial wilt of sweet pepper in a field treated with palmarosa oil, the latency period is increased (38%), the bacterial wilt index (36%), and the area under the disease progress curve (38%) are reduced [21]. In addition, due to low toxicity [22] and environmental friendliness [23], it is sustainable for 1 and EO against R. solanacearum compared with chemical pesticides.
Essentially, 1 belongs to the group of hydroxycinnamic acids (HCAs), and they are known to play multifunctional roles in rhizospheric plant-microbe interactions. In response to root pathogens, natural compounds that form chemical barriers play a key role in preventing pathogens from infecting plant roots, and many plants release de novo synthesized HCAs into the rhizosphere [24]. EO is a complex, volatile mixture that demonstrates antibacterial activity individually or as mixtures; its mechanism of bacteriostasis is very complicated and may be described as EO acting on membrane integrity [25]. The combined use of 1 and EO can improve their antimicrobial activity and may eliminate L. monocytogenes from mildly acid food products [26]. Although the combined effects of EO changed depending on the strain and the type of EO used, generally, the use of combinations increased the efficacy of EO [27].
In this study, ferulic acid ester (2) was synthesized in order to enhance biological activity, the combined antimicrobial efficiency of a binary mixture was determined, and the mechanism of bacteriostasis was discussed. This research was ready for the development of potential green antimicrobial agents, thereby reducing the pollution of chemical pesticides and promoting the sustainability of agriculture.
2. Materials and Methods
2.1. Materials and Bacterial
Peppermint EO (EO1), Artemisia EO (EO2), Citronella EO (EO3), Chamomile EO (EO4), Fennel EO (EO5) and Patchouli EO (EO6) were purchased from Jishui Lianxing Spice Oil Co. Ltd. (Jian, China). Other reagents were purchased from Aladdin Reagent (Shanghai) Co., Ltd. (Shanghai, China). The R. solanacearum of mulberry (PRJNA782242) was previously isolated by the lab members. It was cultured at 30 °C in a medium I (purified water with 0.1% of casamino acids, 0.5% of glucose, and 1% of tryptone). Medium II was prepared by adding 0.017 g of agar powder to 1mL Medium I. Bacterial suspensions of R. solanacearum were approximately 108 CFU/mL at the optical density (OD) of 0.3–0.5 at 600 nm.
2.2. Gas Chromatography Analysis
The EO was analyzed by gas chromatography-mass spectrometry [28]. HP-INNOWax polyethylene glycol (30 m × 0.32 mm × 0.25 μm) was used. The carrier gas was helium, with a flow rate of 2.0 mL/min. The chromatographic analysis started at 40 °C and was maintained for 2 min. The temperature was raised to 240 °C at a rate of 5 °C/min and maintained for 18 min. The injector temperature was 250 °C with a split ratio of 20:1. The temperatures of the ionization source and transfer lines were 230 °C and 250 °C, respectively. The ionic energy was 70 eV, and the mass range was 40–400 Da.
2.3. Synthesis of Ferulic Acid Ester
Based on this esterification method [29] with minor modifications and following the general procedure as shown in Scheme 1, in brief, 1 (5 mmol), 4-dimethylaminopyridine (0.25 mmol), and N, N′-dicyclohexylcarbodiimide (6 mmol) was dissolved in 20 mL of tetrahydrofuran, alcohol (30 mmol) was slowly added dropwise to the mixed solution. Then it was stirred for 12 h under nitrogen flow at room temperature. After filtration and concentration, the required product was purified by column chromatography (petroleum ether: ethyl acetate = 6:1), and their structures were characterized by FT–IR NMR and MS.
2.4. Determination of the Inhibition Rate (IR)
Based on this spectrophotometric method with slight modifications [30], briefly, 40 μL of bacterial suspension was added to 156 μL of Medium I and then replenished with different concentrations of 4 μL of the sample (1, 2a–2g and EO1–6). The resulting solution was incubated at 30 °C for 24 h. The control check group (CK) used 4 μL of ethanol. The IR was calculated with Equation (1), and the half-maximal effective concentration (EC50) value was obtained from the calculation of the fitted curve.
T0sample and TFsample indicate the absorbance values of the bacterial suspension before and after adding the sample, respectively. T0blank and TFblank indicate the absorbance values of the bacterial suspension before and after adding the control solution, respectively.
2.5. Determination of the MIC Values
Based on the serial microdilution method [31], briefly, 1 μL of 1% 2,3,5-Triphenyl-tetrazolium chloride (TTC) and 1 μL of bacteria suspension were inoculated into 198 μL of Medium I containing the samples (2e and EO1–6, 0.12–9.6 mg/mL). The mixture was incubated at 30 °C for 24 h. The MIC value is defined as the minimum concentration of the sample that does not produce a pink color.
2.6. Determination of Combined Antimicrobial Efficiency
Based on this checkerboard method [32] with minor modifications, briefly, 1 μL of 1% TTC and 1 μL of bacteria suspension were inoculated into 198 μL of Medium I containing equal volumes of different concentrations of 2e and EO1–6 (2, 1, 0.5 and 0.25 times MIC). The mixture was incubated at 30 °C for 24 h. The fractional inhibitory concentration index (FIC) was calculated using Equation (2).
The MICA and MICB values represented the lowest concentrations of component A and component B when used alone, and the MICmixA and MICmixB values represented the lowest concentrations of A and B when combined.
2.7. Determination of the Minimum Bactericidal Concentration (MBC)
Based on this method [33] with minor modifications, briefly, the ethanol solution of the samples (2e, EO1, and 2e-EO1) was added separately to plates with freshly sterilized Medium II and the final concentration of the sample was 0.1–3.2 mg/mL. The plate was immediately placed on the super-clean table and cooled for half an hour. Then, 1 μL of bacterial suspension was dropped in the center of the culture dish. The MBC value is defined as concentration without bacterial growth after 96 h at 30 °C.
2.8. Growth Curve Assay
Based on this method [34] with modifications, briefly, the bacterial suspension was diluted by one-thousandth with Medium I, the samples (2e, EO1, and 2e-EO1) were added with a final concentration of 0.2 mg/mL, and the resulting solution was incubated in a shaking table at 30 °C. The optical density was read spectrophotometrically at 600 nm with a time interval of 4 h. The growth of R. solanacearum was observed by the change in the absorbance value.
2.9. Biofilm Assay
Bacteria form biofilms on a wide range of abiotic surfaces [35]; the polyvinylchloride (PVC) microtiter plate assay [31] was used for the quantification of the biofilm with minor modifications. Briefly, 40 μL of bacterial suspension was added to 160 μL of Medium I and then supplemented with the samples (2e, EO1, and 2e-EO1) at final concentrations of 0.1–1.0 mg/mL. The resulting solution was removed carefully after cultivating for 24 h. The residue was cleaned with 200 μL of purified water and was fixed for 15 min with 200 μL of methanol. Then, 220 μL of 0.1% crystal violet was mixed with the biofilms for 30 min and was removed. The floating color was washed twice with 200 μL of purified water and was removed. The residue was dried at 25 °C for 30 min. The crystal violet adsorbed on the biofilms was dissolved with 200 μL of 95% ethanol for 30 min, and the OD590nm value of the solution was measured.
2.10. Influence of Pathogenic Gene Expression
The influence of pathogenic gene expression was analyzed according to this early report [36]. Briefly, 0.8 mg of the samples (2e, EO1, and 2e-EO1) were added to 4 mL of the bacterial suspension. The mixture was incubated at 30 °C for 24 h and centrifuged at 12,000 rpm for 3 min to collect the precipitates. The total RNA extraction process was carried out according to the protocol modifications. Briefly, the total RNA was extracted by the Trizol reagent (Invitrogen, Waltham, MA, USA) and was purified by DNase I (Promega, Madison, WI, USA). The Prime Script RT Reagent Kit with gDNA Eraser (Takara Biotechnology, Otsu, Japan) was used to synthesize the cDNA, and the relative expression of pathogenicity-related genes (hrpB, pehC, pilT, polA, aceE, egl and phcA) was determined by the Light Cycler 96 real-time PCR system. Then, 20 mL of the reaction system was made up of 10 mL of TB Green, 6.4 mL of RNase-free double-distilled water, 2 mL of cDNA, 0.8 mL of the forward primer (F), and 0.8 mL of the reverse primer (R). The specific primers were synthesized as shown in Table 1, and the 16S rRNA of R. solanacearum was used as CK.
2.11. Statistical Analysis
All tests were performed in triplicate, and the experimental data was recorded as the average value and stdev (SD). Variance analysis performed was one-way analysis and Tukey’s test.
3. Results and Discussion
3.1. Synthesis of Ferulic Acid Ester
The yields of 2a–2g were 74.57 ± 1.45%, 72.31 ± 2.38%, 69.75 ± 1.48%, 65.42 ± 2.49%, 67 ± 3.38%, 69 ± 3.29%, and 67 ± 3.36%, respectively. The spectral data of Figures S1, S2, and Figure 1 confirmed the chemical structure of 2a–2g. In the ESI-MS analysis, 223, 237, 237, 251, 285, 265, and 293 were molecular ion peaks of 2a–2g, respectively.
The characteristic structure of this series is similar. 2e is an example of the structural analysis of this series. For the 1H NMR analysis, 1H chemical shifts of the protons of the aromatic ring were assigned at δ 7.17, δ 7.05, and δ 6.79 ppm, and a shift signal at δ 4.15 ppm corresponded to the hydrogens of the first carbon in the ester alkyl chain, which suggested that the ester group was obtained. Besides, the other signals of chemical shifts between δ 1.68 and 0.93 δ ppm were assigned to the hydrogens of the other carbons in the alkyl chain.
For the FT–IR analysis, 3450 cm−1 was the stretching vibration peak of –OH, 2970 cm−1 was the stretching vibration peak of C–H, 1684 cm−1 was the stretching vibration peak of C=O, 1634 cm−1, 1624 cm−1, 1600 cm−1, and 1509 cm−1 were the vibration peaks of conjugated double bonds of a benzene ring and its substituents, 1464–1427 cm−1 were the bending vibration peaks of C–H, 1323–1269 cm−1 were the C–O vibration peaks on a benzene ring, and the wide peak near 1156 cm−1 was the C–O–C vibration peak of an ester.
3.2. Antimicrobial Activity of Ferulic Acid Ester against R. solanacearum
Figure 2 showed that the inhibition rate of 2 was proportional to the concentration, and the antibacterial activity was stronger than 1. This was consistent with the report that electron-donating substituents on the benzene ring might enhance the antibacterial activity [37]. At the concentration of 0.32 mg / mL, the inhibition rates of 2a, 2b, 2c, 2d, 2e, 2f, and 2g were 74.62 ± 2.42%, 79.26 ± 3.13%, 82.04 ± 4.41%, 84.13 ± 4.38%, 89.51 ± 4.84%, 72.13 ± 6.30% and 70.25 ± 4.83%, respectively. Their EC50 values were calculated to be 0.12 ± 0.002 mg/mL, 0.12 ± 0.004 mg/mL, 0.12 ± 0.004 mg/mL, 0.10 ± 0.005 mg/mL, 0.07 ± 0.003 mg/mL, 0.13 ± 0.005 mg/mL, and 0.13 ± 0.002 mg/mL respectively. Therefore, the activity of short-chain ferulate varies little with different alkyl numbers, and the activity of butyl ester (2d) is slightly stronger than that of other short-chain alkyl esters (2a, 2b, 2c, 2f, 2g). Butyl chloride (2e) is larger than butyl ester (2d), which may be related to the bacteriostatic effect of chlorine [38].
3.3. Combined Antimicrobial Efficiency of 2e and EO
Table 2 shows that the MIC value of 2e against R. solanacearum was 0.64 ± 0.03 mg/mL. According to the FIC index, a mixture of 2e and EO1 showed a synergistic effect, and a mixture of 2e and other EOs showed an additive effect. The bacteriostatic effect of 2e and six EOs were stronger than that of either alone. This may be because of the multi-target effect and pharmacokinetic effects [39], and there may be synergy between the major and minor components of EO [40] and between EO and bacteriostatic agents [41]. Among six combinations, 2e-EO1 was the most significant; the MIC value of 2e-EO1 was 0.4 ± 0.05 mg/mL, and their optimal mass ratio of 2e and EO1 was 1:1.5. The amount of 2e in the composition was reduced to 25%. Figure 3 showed that the MBC values of 2e, EO1, and 2e-EO1 were 1.60 ± 0.08 mg/mL, 3.20 ± 0.14 mg/mL, and 0.80 ± 0.06 mg/mL, respectively. The EC50 value of 2e EO1 was 0.05 ± 0.004 mg/mL, as shown in Figure S3. The main component of EO1 was menthol (65.38%), as shown in Table 3. These results verified that the bacteriostatic synergy of 2e-EO1 was significant. According to another report [42], EO1 can increase membrane permeability and inhibition of bacterial quorum sensing ability in multidrug-resistant E. coli, aiding in the reversal of antibiotic resistance.
3.4. The Growth Curve of R. solanacearum
The effect of 2e-EO1 on the growth curve of R. solanacearum is shown in Figure 4. The growth process of CK groups showed that the lag phase of R. solanacearum was 0–8 h, where there was a lesser growth of bacteria, and the log phase was 8–24 h, where the growth rate was rapid. This result was consistent with another report [43]. In the growth process of 2e, EO1, and 2e-EO1, the absorbance value of R. solanacearum changed slowly at 0–12 h, only increased by 0.027 ± 0.013, 0.045 ± 0.016 and 0.023 ± 0.009 respectively, and their growth rate was significantly slower than that of the control group (0.127 ± 0.02). Thus, 2e, EO1, and 2e-EO1 have good antimicrobial effects in the first 12 h. The absorbance value of R. solanacearum treated with 2e, EO1, and 2e-EO1 increased significantly from 12 to 24 h, were 0.153 ± 0.006, 0.292 ± 0.005, and 0.126 ± 0.004, respectively. Among them, the change in the 2e-EO1 treatment group was relatively little, and the antimicrobial effect was stronger than that of the 2e and EO1 treatment groups.
3.5. Biofilm Assay of R. solanacearum
Figure 5 showed that 2e, EO1, and 2e-EO1 had different degrees of inhibition for the biofilm formation of R. solanacearum, and 2e-EO1 was especially significant. The OD590nm value of bacteria treated with 1.0 mg/mL of 2e-EO1 was 0.35 ± 0.03, and the OD590nm value was decreased by 57.83 ± 6.46%. Similar to many plant pathogens, bacterial biofilms formed in the roots of host plants can effectively colonize the xylem walls of host plants [44] and promote bacterial invasion and infection [45]. According to another report [46], the specific phenolic exudates in plants infected with R. solanacearum that exhibit promote plant resistance against pathogens and antibacterial activity. In addition, the mechanism of action of EO against R. solanacearum might be described as acting on membrane integrity [47]. The reason was that the membrane structure of the pathogen was damaged, resulting in the thinning of the cell membrane and irregular cavities in cells.
3.6. Influence of Pathogenic Gene Expression
Figure 6 shows the expression of these pathogenic genes of R. solanacearum after treatment with 2e, EO1, and 2e-EO1. Compared with CK, the expression levels of pilT, polA, aceE, egl, and phcA treated with 2e were downregulated to 47.00 ± 9.90%, 89.78 ± 7.83%, 93.17 ± 9.19%, 85.86 ± 8.42% and 54.48 ± 3.92%, respectively. The expression levels of pilT, polA, aceE, and phcA treated with EO1 were downregulated to 23.53 ± 2.41%, 75.44 ± 7.03%, 26.30 ± 3.83% and 12.81% ± 1.56%, respectively. The expression levels of hrpB, pehC, pilT, polA, aceE, egl, and phcA treated with 2e-EO1 were downregulated to 18.81 ± 4.79%, 30.50 ± 4.03%, 14.00 ± 1.97%, 44.90 ± 8.00%, 86.79 ± 12.48%, 23.90 ± 2.01%, and 27.56 ± 4.14%, respectively. Among these pathogenic genes, hrpB is a core type II and type III secretion system regulator gene [48]. pehC, pilT, polA, and aceE are important factors in the early stage of bacterial blight, and egl, along with phcA, are important factors in the late stage of bacterial blight [21]. This result indicated that the drug has a multi-level inhibitory effect on the infection of R. solanacearum. Under the synergistic effect of 2e and EO1, the target of action was expanded. The multi-target mechanism is beneficial in reducing the drug resistance of bacteria.
4. Conclusions
In this research, ferulic acid esters (2a–2g) were synthesized, the inhibitory activity against R. solanacearum was determined, and the antibacterial combination of 2 and EO was optimized. The antibacterial curve of the composition and the expression of pathogenic factors were discussed. This paper showed that the EC50 value of chlorobutyl ferulate (2e) was 0.07 mg/mL, and the composition of 2e and EO1 had a synergistic effect. The MIC and MBC values of 2e-EO1 were 0.40 mg/mL and 0.80 mg/mL, respectively. In the growth curve of R. solanacearum, the 2e-EO1 treatment group grew slowly, and the multiple pathogenic genes (hrpB, pehC, pilT, polA, aceE, egl, and phcA) of R. solanacearum treated with 2e-EO1 were significantly downregulated. These results show that 2e-EO1 has significant bacteriostatic properties against R. solanacearum. In addition, 2 and EO are natural substances with low toxicity and environmental safety, and 2e-EO1 has the potential to become a green pesticide.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su142416348/s1, Figure S1. ESI-MS analysis of ferulate esters; Figure S2. 1H NMR spectrum of ferulate esters; Figure S3. Antimicrobial activity of 2e-EO1 against R. solanacearum.
Author Contributions
Conceptualization, F.-A.W.; data curation, Q.-B.T.; formal analysis, Q.-B.T. and S.S.; funding acquisition, Q.-B.T. and F.-A.W.; investigation, H.-C.S.; methodology, Q.-B.T. and P.L.; project administration, F.-A.W.; resources, F.-A.W.; supervision, F.-A.W.; validation, H.-C.S.; writing—original draft preparation, Q.-B.T.; writing—review and editing, Q.-B.T. and F.-A.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Science and Technology Plan Project of Taizhou City (TN202124), the Key Research and Development Program (Modern Agriculture) of Zhenjiang City (NY2020005), the Key Research and Development Program (Modern Agriculture) of Jiangsu Province (BE2021344), the Jiangsu Graduate Research Innovation Program (KYCX18_2347), the Innovation and entrepreneurship training program for college students of Jiangsu Province (201913981010Y), and the Special Fund for China Agriculture Research System (CARS-18).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data and contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyska, M.; Cunniffe, N.; Gilligan, C. Trade-off between disease resistance and crop yield: A landscape-scale mathematical modelling perspective. J. R. Soc. Interface 2016, 13, 20160451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurabachew, H.; Ayana, G. Bacterial Wilt caused by Ralstonia solanacearum in Ethiopia: StatusaAnd Management Approaches: A Review. Int. J. Phytopathol. 2017, 5, 107–119. [Google Scholar] [CrossRef]
- Mansfeld, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicker, E.; Grassart, L.; Coransonbeaudu, R.; Mian, D.; Guilbaud, C.; Fegan, M.; Prior, P. Ralstonia solanacearum strains from Martinique (French West Indies) exhibiting a new pathogenic potential. Appl. Environ. Microb. 2007, 73, 6790–6801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuliar; Nion, Y.A.; Toyota, K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Eeviron. 2015, 30, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Genin, S.; Denny, T.P. Pathogenomics of the Ralstonia solanacearum species complex. Annu. Rev. Phytopathol. 2012, 50, 67–89. [Google Scholar] [CrossRef]
- Schreinemachers, P.; Tipraqsa, P. Agricultural pesticides and land use intensification in high, middle and low income countries. Food Policy 2012, 37, 616–626. [Google Scholar] [CrossRef]
- Fu, L.; Penton, C.R.; Ruan, Y.Z.; Shen, Z.Z.; Xue, C. Inducing the rhizosphere microbiome by biofertilizer application to suppress banana Fusarium wilt disease. Soil. Biol. Biochem. 2017, 104, 39–48. [Google Scholar] [CrossRef]
- Chellemi, D.O.; Rosskopf, E.N.; Kokalis-Burelle, N. The Effect of Transitional Organic Production Practices on Soilborne Pests of Tomato in a Simulated Microplot Study. Phytopathology 2013, 103, 792–801. [Google Scholar] [CrossRef]
- Teixeira, F.R.; Lima, M.C.O.P.; Almeida, H.O.; Romeiro, R.S.; Silva, D.J.H.; Pereira, P.R.G.; Fontes, E.P.B.; Baracat-Pereira, M.C. Bioprospection of cationic and anionic antimicrobial peptides from bell pepper leaves for inhibition of Ralstonia solanacearum and Clavibacter michiganensis ssp. michiganensis growth. J. Phytopathol. 2006, 154, 418–421. [Google Scholar] [CrossRef]
- Yuan, G.Q.; Li, Q.Q.; Qin, J.; Ye, Y.F. Isolation of methyl gallate from Toxicodendron sylvestre and its effect on tomato bacterial wilt. Plant Dis. 2012, 96, 1143–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim, E.P.D.R.; De-Andrade, F.W.R.; Moraes, E.M.D.S.; Silva, J.C.D.S.; Lima, R.D.S.; Lemos, E.E.P.D. Antibacterial activity of essential oils and extracts on the development of Ralstonia solanacearum in banana seedlings/Atividade antibacteriana de oleos essenciais e extratos vegetais sobre o desenvolvimento de Ralstonia solanacearum em mudas de bananeira. Rev. Bras. Frutic. 2011, 33, 392–399. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, S.C.; Soares, A.C.F.; Brito, A.S.; Laranjeira, F.F.; Ledo, C.A.S.; Santos, A.P. Control of tomato bacterial wilt through the incorporation of aerial part of pigeon pea and crotalaria to soil. Summa Phytopathol. 2006, 32, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Mishra, J.; Dutta, V.; Arora, N.K. Biopesticides in India: Technology and sustainability linkages. 3 Biotech 2020, 10, 210. [Google Scholar] [CrossRef]
- Ashmawy, N.A.; Behiry, S.I.; Al-Huqail, A.A.; Ali, H.M.; Salem, M.Z.M. Bioactivity of selected phenolic acids and hexane extracts from Bougainvilla spectabilis and Citharexylum spinosum on the growth of Pectobacterium carotovorum and Dickeya solani Bacteria: An opportunity to save the environment. Processes 2020, 8, 482. [Google Scholar] [CrossRef]
- Ikram, M.; Ali, N.; Jan, G.; Hamayun, M.; Iqbal, A. Novel antimicrobial and antioxidative activity by endophytic Penicillium roqueforti and Trichoderma reesei isolated from Solanum surattense. Acta Physiol. Plant. 2019, 41, 164. [Google Scholar] [CrossRef]
- Li, L.; Feng, X.; Tang, M.; Hao, W.; Han, Y.; Zhang, G.; Wan, S. Antibacterial activity of Lansiumamide B to tobacco bacterial wilt (Ralstonia solanacearum). Microbiol. Res. 2014, 169, 522–526. [Google Scholar] [CrossRef]
- Vu, T.T.; Choi, G.J.; Kim, J.C. Plant-derived Antibacterial Metabolites Suppressing Tomato Bacterial Wilt Caused by Ralstonia solanacearum. Res. Plant Dis. 2017, 23, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Abo-Elyousr, K.A.M.; Seleim, M.A.A.; Abd-El-Moneem, K.M.H.; Saead, F.A. Integrated effect of glomus mosseae and selected plant oils on the control of bacterial wilt disease of tomato. Crop Prot. 2014, 66, 67–71. [Google Scholar] [CrossRef]
- Alves, A.O.; Santos, M.M.B.; Santos, T.C.G.; Souza, E.B.; Mariano, R.L.R. Biofumigation with essential oils for managing bacterial wilt of sweet peppers. J. Plant Pathol. 2014, 96, 363–367. [Google Scholar]
- Ferraz, C.A.; Pastorinho, M.R.; Palmeira-de-Oliveira, A.; Sousa, A.C. Ecotoxicity of plant extracts and essential oils: A review. Environ. Pollut. 2022, 292, 118319. [Google Scholar] [CrossRef] [PubMed]
- Ebadi, M.T.; Mollaei, S.; Khurizadeh, S. Evaluation of volatile and phenolic compounds, and antioxidant activity of different parts of Ferulago angulata (schlecht.) Boiss. Ind. Crop. Prod. 2019, 140, 111589. [Google Scholar]
- Wallis, C.M.; Chen, J. Grapevine phenolic compounds in xylem sap and tissues are significantly altered during infection by Xylella fastidiosa. Phytopathology. 2012, 102, 816–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.M.; Yu, J.P. Chemical Composition, Antimicrobial Activity and Mechanism of Action of Essential Oil from the Leaves of Macleaya Cordata (Willd.) R. B r. J. Food Safety 2015, 35, 227–236. [Google Scholar] [CrossRef]
- Miyague, L.; Macedo, R.E.F.; Meca, G.; Holley, R.A.; Luciano, F.B. Combination of phenolic acids and essential oils against Listeria monocytogenes. LWT Food Sci. Technol. 2015, 64, 333–336. [Google Scholar] [CrossRef]
- Mutlu-Ingok, A.; Tasir, S.; Seven, A.; Akgun, N.; Karbancioglu-Guler, F. Evaluation of the single and combined antibacterial efficiency of essential oils for controlling Campylobacter coli, Campylobacter jejuni, Escherichia coli, Staphylococcus aureus, and mixed cultures. Flavour Fragr. J. 2019, 34, 280–287. [Google Scholar] [CrossRef]
- Kim, J.H.; Seo, C.S.; Shin, H.K. Simultaneous Determination of (-)-Menthone and (-)-Menthol in Menthae Herba by Gas Chromatography and Principal Component Analysis. Nat. Prod. Sci. 2010, 16, 180–184. [Google Scholar]
- Nadal, J.; Pedroso, F.B.; Minozzo, B.R.; Brito, P.S.D.; Farago, P.V.; Vellosa, J.C.R.; Miyoshi, E. A simple and high-yield synthesis of hexadecyl ferulate and its in vitro antioxidant potential. Braz. Arch. Biol. Technol. 2018, 61, 1–10. [Google Scholar] [CrossRef]
- Xu, Y.; Sheng, S.; Liu, X.; Wang, C.; Xiao, W.; Wang, J.; Wu, F.A. Cooperative reinforcement of ionic liquid and reactive solvent on enzymatic synthesis of cafeic acid phenethyl ester as an in vitro inhibitor of plant pathogenic bacteria. Molecules 2017, 22, 72. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Ding, W.; Xu, Y.Q.; Wu, D.S.; Li, S.L.; Chen, J.N.; Guo, B. New insights into the antibacterial activity of hydroxycoumarins against Ralstonia solanacearum. Molecules 2016, 21, 468. [Google Scholar] [CrossRef] [PubMed]
- Ayari, S.; Shankar, S.; Follett, P.; Hossain, F.; Lacroix, M. Potential synergistic antimicrobial efficiency of binary combinations of essential oils against Bacillus cereus and Paenibacillus amylolyticus—Part A. Microb. Pathog. 2020, 141, 104008. [Google Scholar] [CrossRef] [PubMed]
- Klančnik, A.; Piskernik, S.; Jeršek, B.; Možina, S.S. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J. Microbiol. Meth. 2010, 81, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Sowndarya, J.; Rubini, D.; Sinsinwar, S.; Senthilkumar, M.; Nithyanand, P.; Vadivel, V. Gallic Acid an Agricultural Byproduct Modulates the Biofilm Matrix Exopolysaccharides of the Phytopathogen Ralstonia solanacearum. Curr. Microbiol. 2020, 77, 3339–3354. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Koberto, R. Genetic approaches to study of biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar] [PubMed]
- Wang, J.Z.; Yan, C.H.; Zhang, X.R.; Tu, Q.B.; Xu, Y.; Sheng, S.; Wu, F.A.; Wang, J. A novel nanoparticle loaded with methyl caffeate and caffeic acid phenethyl ester against Ralstonia solanacearum—A plant pathogenic bacteria. RSC Adv. 2020, 10, 3978–3990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatkar, A.; Nanda, A.; Kumar, P.; Narasimhan, B. Synthesis and antimicrobial evaluation of ferulic acid derivatives. Res. Chem. Intermed. 2015, 41, 299–309. [Google Scholar] [CrossRef]
- Waters, B.W.; Hung, Y. The Effect of pH and Chloride Concentration on the Stability and Antimicrobial Activity of Chlorine-Based Sanitizers. J. Food Sci. 2014, 79, 1750. [Google Scholar] [CrossRef]
- Langeveld, W.T.; Veldhuizen, E.J.; Burt, S.A. Synergy between essential oil components and antibiotics: A review. Crit. Rev. Microbiol. 2014, 40, 76–94. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic interactions of phytochemicals with antimicrobial agents: Potential strategy to counteract drug resistance. Chem. Interactions 2019, 308, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Yap, P.S.; Krishnan, T.; Yusoff, K.; Chan, K.; Yap, W.; Lai, K.; Lim, S.E. Mode of Action: Synergistic Interaction of Peppermint (Mentha × piperita L. Carl) Essential Oil and Meropenem Against Plasmid-Mediated Resistant E. coli. Rec. Nat. Prod. 2018, 12, 582–594. [Google Scholar] [CrossRef]
- Sajna, K.V.; Sukumaran, R.K.; Gottumukkala, L.D.; Jayamurthy, H.; Dhar, K.S.; Pandey, A. Studies on structural and physical characteristics of a novel exopolysaccharide from Pseudozyme sp. NII 08165. Int. J. Biol. Macromol. 2013, 59, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Shoda, M. Bacterial control of plant diseases. J. Biosci. Bioeng. 2000, 89, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Pan, Z.C.; Prior, P.; Xu, J.S.; Zhang, Z.; Zhang, H.; Zhang, L.Q.; He, L.Y.; Feng, J. Genetic diversity of Ralstonia solanacearum strains from China. Eur. J. Plant Pathol. 2009, 125, 641–653. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, Y.F.; Wang, Z.Y.; Fan, Y.; Yang, X.; Wang, Z.G.; Yu, S.; Pang, Q.X.; Cao, A.C. Deoxymikanolide adversely altered physiology and ultrastructure of Ralstonia solanacearum. Pestic. Biochem. Phys. 2010, 174, 104803. [Google Scholar] [CrossRef]
- Diao, W.R.; Hu, Q.P.; Zhang, H.; Xu, J.G. Chemical composition, antibacterial activity and mechanism of action of essential oil from seeds of fennel (Foeniculum vulgare Mill.). Food Control. 2014, 35, 109–116. [Google Scholar] [CrossRef]
- Cunnac, S.; Boucher, C.; Genin, S. Characterization of the cis-acting regulatory element controlling HrpB-mediated activation of the type III secretion system and effector genes in Ralstonia solanacearum. J. Bacteriol. 2004, 186, 2309–2318. [Google Scholar] [CrossRef]
Scheme 1.
Synthesis of 2a–2g.

Figure 1.
FT-IR spectrum of ferulate esters. 1: Ferulic acid, 2a: Ethyl ferulate, 2b Propyl ferulate, 2c: Isopropyl ferulate, 2d: Butyl ferulate, 2e: Chlorobutyl ferulate, 2f: Amyl ferulate, 2g: Heptyl ferulate.
Figure 1.
FT-IR spectrum of ferulate esters. 1: Ferulic acid, 2a: Ethyl ferulate, 2b Propyl ferulate, 2c: Isopropyl ferulate, 2d: Butyl ferulate, 2e: Chlorobutyl ferulate, 2f: Amyl ferulate, 2g: Heptyl ferulate.

Figure 2.
Antimicrobial activity of ferulic acid ester against R. solanacearum. 1: Ferulic acid, 2a: Ethyl ferulate, 2b Propyl ferulate, 2c: Isopropyl ferulate, 2d: Butyl ferulate, 2e: Chlorobutyl ferulate, 2f: Amyl ferulate, 2g: Heptyl ferulate. The OD600nm values of the bacterial solution was detemined after cultivation for 24 h at 30 °C. The EC50 values were fitted by Logistic evaluation.
Figure 2.
Antimicrobial activity of ferulic acid ester against R. solanacearum. 1: Ferulic acid, 2a: Ethyl ferulate, 2b Propyl ferulate, 2c: Isopropyl ferulate, 2d: Butyl ferulate, 2e: Chlorobutyl ferulate, 2f: Amyl ferulate, 2g: Heptyl ferulate. The OD600nm values of the bacterial solution was detemined after cultivation for 24 h at 30 °C. The EC50 values were fitted by Logistic evaluation.

Figure 3.
Bactericidal activity of 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, and 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). a. b, c, d, e, f and g were 3.2, 1.6, 0.8, 0.4, 0.2, 0.1, and 0.05 mg/mL, respectively. The culture condition was 30 °C for 96 h.
Figure 3.
Bactericidal activity of 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, and 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). a. b, c, d, e, f and g were 3.2, 1.6, 0.8, 0.4, 0.2, 0.1, and 0.05 mg/mL, respectively. The culture condition was 30 °C for 96 h.

Figure 4.
The growth curve of R. solanacearum treated with 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, and 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). The culture condition was 30 °C for 36 h. The density of R. solanacearum (OD600nm ≈ 1.0) was approximately 109 CFU/mL.
Figure 4.
The growth curve of R. solanacearum treated with 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, and 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). The culture condition was 30 °C for 36 h. The density of R. solanacearum (OD600nm ≈ 1.0) was approximately 109 CFU/mL.

Figure 5.
The biofilm assay of R. solanacearum treated with 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). Biofilm assay after treatment with different concentrations of 2e, EO1 and 2e-EO1 at 30 °C for 24 h in the 96-well plate. The letters (A, B, C) on the column indicate the differences between the different treatment groups; the same letters indicate that the difference is not significant (p > 0.05), and completely different letters indicate significant differences (p < 0.05).
Figure 5.
The biofilm assay of R. solanacearum treated with 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). Biofilm assay after treatment with different concentrations of 2e, EO1 and 2e-EO1 at 30 °C for 24 h in the 96-well plate. The letters (A, B, C) on the column indicate the differences between the different treatment groups; the same letters indicate that the difference is not significant (p > 0.05), and completely different letters indicate significant differences (p < 0.05).

Figure 6.
Effect of the expression of the pathogenicity-related gene of R. solanacearum treated with 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). 2e, EO1 and 2e-EO1 were added to the bacterial suspension (OD600nm ≈ 1.0) and incubated overnight at 30 °C for 24 h. The letters (A, B, C) on the column indicate the differences between the different treatment groups; the same letters indicate that the difference is not significant (p > 0.05), and completely different letters indicate significant differences (p < 0.05).
Figure 6.
Effect of the expression of the pathogenicity-related gene of R. solanacearum treated with 2e-EO1. CK: Ethanol, 2e: Chlorobutyl ferulate, EO1: Peppermint EO, 2e-EO1: Mixture of chlorobutyl ferulate and peppermint EO (1:1.5). 2e, EO1 and 2e-EO1 were added to the bacterial suspension (OD600nm ≈ 1.0) and incubated overnight at 30 °C for 24 h. The letters (A, B, C) on the column indicate the differences between the different treatment groups; the same letters indicate that the difference is not significant (p > 0.05), and completely different letters indicate significant differences (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer information of the pathogenicity-related genes of R.solanacearum.
| Primer Name | Nucleotide Sequence(5′→3′) | Size (bp) |
|---|---|---|
| hrpB | F: TTCTCGATGATGTAGCGATAGG R: GCTGGAATTTTCGACTTCCTCTA | 238 |
| pehC | F: GTTGTTCGGATTGCTGTACG R: AGTCAAACGATTGCCTGAACTA | 227 |
| pilT | F: AAGAACAAAGCGTCTGATCTGC R: CTTCCAGGTTTTCTTCGTAATGCT | 175 |
| polA | F: GGAATGTCGGAAAGTCAAGAAA R: CTTGTAGGCGGGGTACAGTTC | 238 |
| ace | F: GCCTATGTGCGTGAGTTCTTCT R: CTTCGAACTTGACGTACGGAAC | 338 |
| egl | F: CAGCGCGACCTACTACAAGA R: TCATCAGCCCGAAGATGAC | 299 |
| phcA | F: GGACATGATCTTCACGGTCAACT R: GACTCATCCTCCTTTTCTGCATC | 298 |
| 16S rRNA | F: CTAGAGTGTGTCAGAGGGAGGTAGA R: ATGTCAAGGGTAGGTAAGGTTTTTC | 349 |
Table 2.
FIC index of the combination of a mixture of 2e and EO against R. solanacearum.
| mg/mL | 2e + EO1 | 2e + EO2 | 2e + EO3 | 2e + EO4 | 2e + EO5 | 2e + EO6 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | 0.64 ± 0.03 | 2.02 ± 0.08 | 0.64 ± 0.03 | 2.42 ± 0.09 | 0.64 ± 0.03 | 1.20 ± 0.08 | 0.64 ± 0.03 | 4.51 ± 0.09 | 0.64 ± 0.03 | 4.44 ± 0.07 | 0.64 ± 0.03 | 4.38 ± 0.05 |
| MICmix | 0.16 ± 0.02 | 0.24 ± 0.03 | 0.32 ± 0.04 | 0.32 ± 0.04 | 0.32 ± 0.06 | 0.13 ± 0.04 | 0.32 ± 0.04 | 1.32 ± 0.04 | 0.32 ± 0.04 | 2.23 ± 0.07 | 0.32 ± 0.04 | 2.25 ± 0.05 |
| FIC | 0.37 ± 0.09 | 0.63 ± 0.06 | 0.61 ± 0.23 | 0.79 ± 0.18 | 1.00 ± 0.07 | 1.01 ± 0.17 | ||||||
| Effect | S | AD | AD | AD | AD | AD | ||||||
2e: Chlorobutyl ferulate, EO1: Peppermint EO, EO2: Artemisia EO, EO3: Citronella EO, EO4: Chamomile EO, EO5: Fennel EO, EO6: Patchouli EO. The FIC ≤ 0.5 indicated a synergistic effect (S). 0.5 < FIC ≤ 1 indicated additive effect (AD), 1 < FIC ≤ 4 indicated no interactive effect (NI), and FIC > 4 indicated antagonistic effect (A).
Table 3.
Relative percentages of the main constituents of EO1 determined by the GC analysis.
| Main Components | Retention Time (min) | Retention Indices | Percentages of the Main Constituents (%) | |
|---|---|---|---|---|
| SI | RSI | |||
| (+)-Dipentene | 15.97 | 874 | 877 | 3.00 |
| L-menthone | 20.13 | 942 | 958 | 8.07 |
| Menthone | 20.49 | 858 | 865 | 5.44 |
| Menthol | 20.76 | 947 | 952 | 65.38 |
| Isomenthol acetate | 24.09 | 946 | 954 | 2.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tu, Q.-B.; Shi, H.-C.; Li, P.; Sheng, S.; Wu, F.-A. Antibacterial Activity of Ferulic Acid Ester against Ralstonia solanacearum and Its Synergy with Essential Oils. Sustainability 2022, 14, 16348. https://doi.org/10.3390/su142416348
AMA Style
Tu Q-B, Shi H-C, Li P, Sheng S, Wu F-A. Antibacterial Activity of Ferulic Acid Ester against Ralstonia solanacearum and Its Synergy with Essential Oils. Sustainability. 2022; 14(24):16348. https://doi.org/10.3390/su142416348
Chicago/Turabian StyleTu, Qing-Bo, Hui-Cong Shi, Ping Li, Sheng Sheng, and Fu-An Wu. 2022. "Antibacterial Activity of Ferulic Acid Ester against Ralstonia solanacearum and Its Synergy with Essential Oils" Sustainability 14, no. 24: 16348. https://doi.org/10.3390/su142416348
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.