
Bacterial Resistance against Heavy Metals in Pseudomonas aeruginosa RW9 Involving Hexavalent Chromium Removal

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Culture Conditions
2.2. Determination of Cr(VI) Tolerance by Cell Viability Measurement
2.3. Chromium (Cr(VI)) Detection and Removal Efficiency
2.4. Quantification of Biosurfactant
2.5. Biosurfactant Extraction and Characterization
3. Results and Discussion
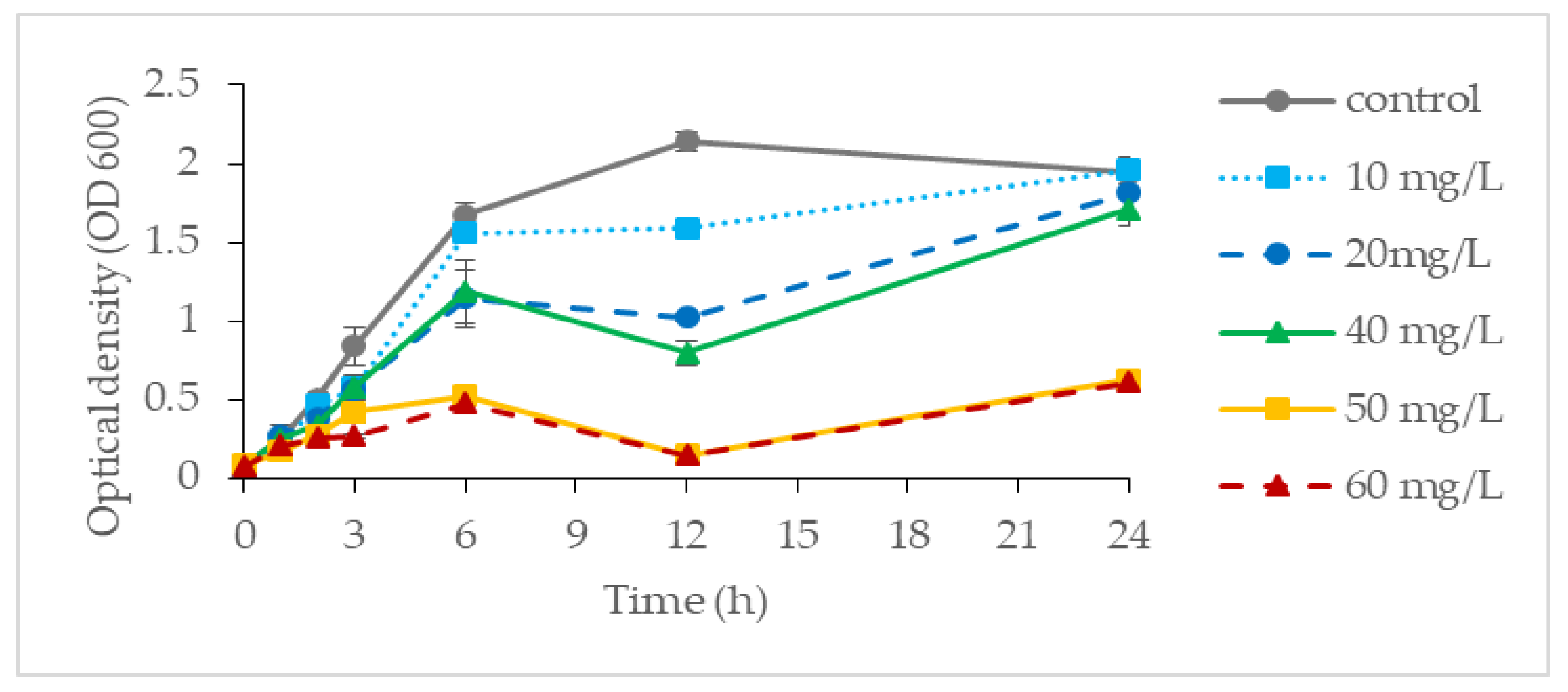
3.1. Cr(VI) Tolerance of Pseudomonas aeruginosa RW 9
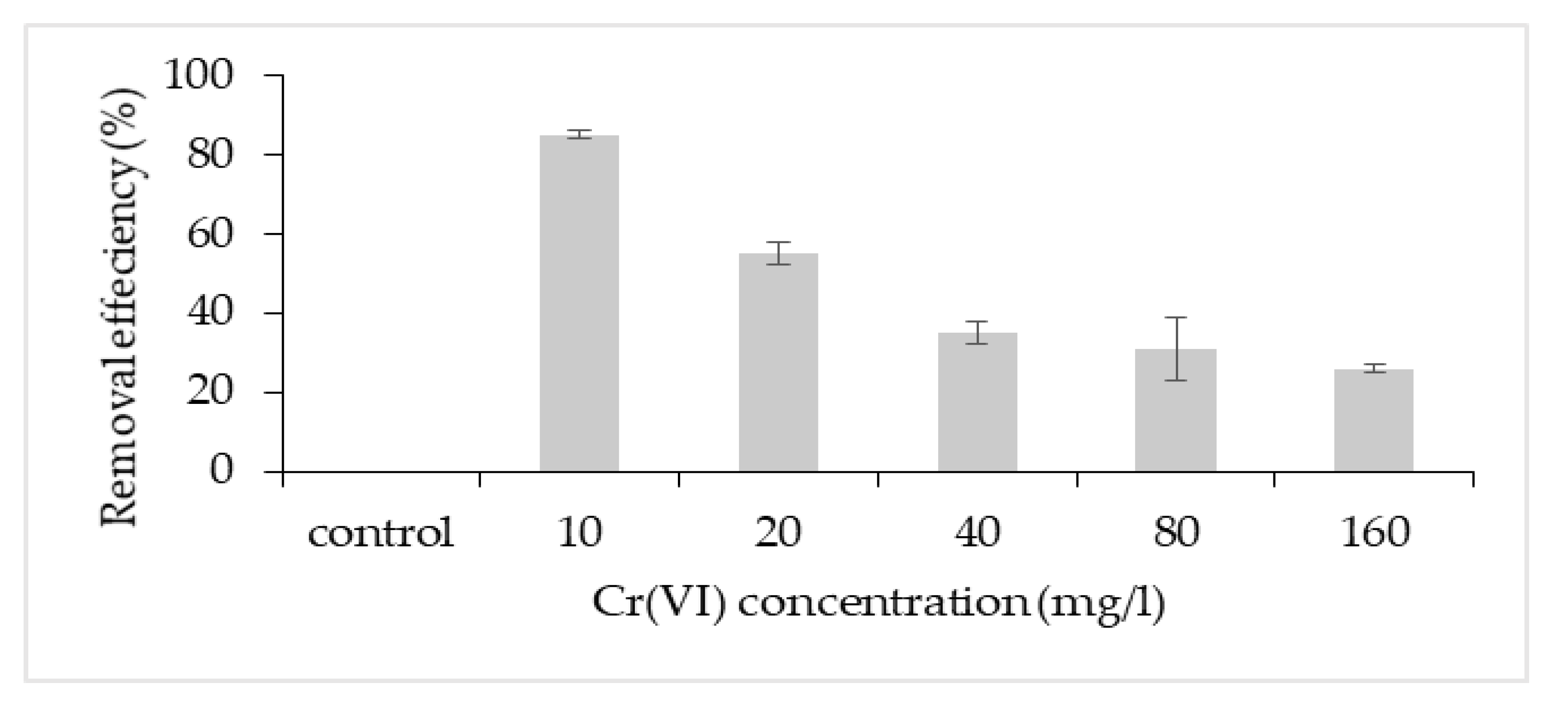
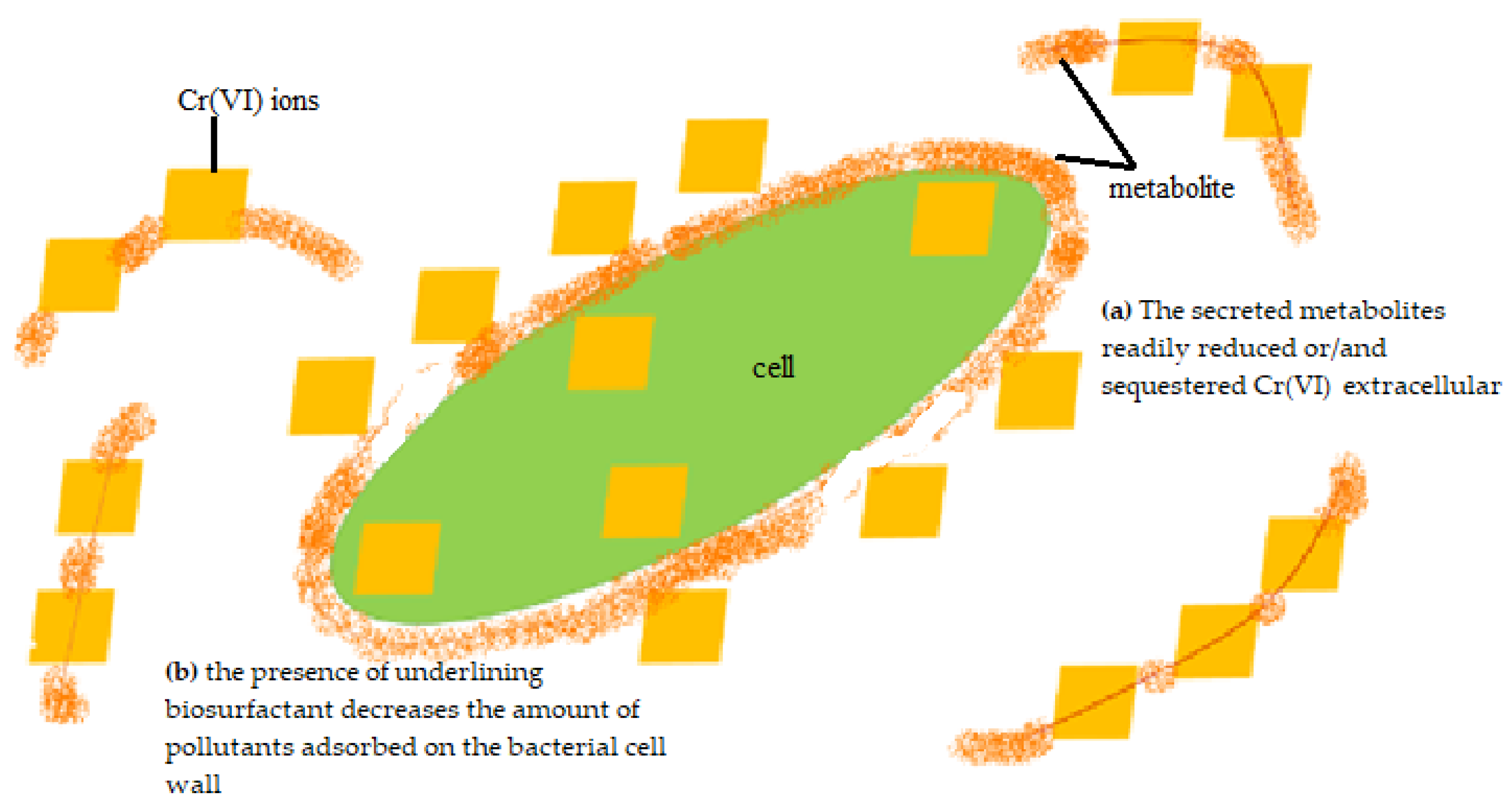
3.2. Chromium Removal Mechanisms
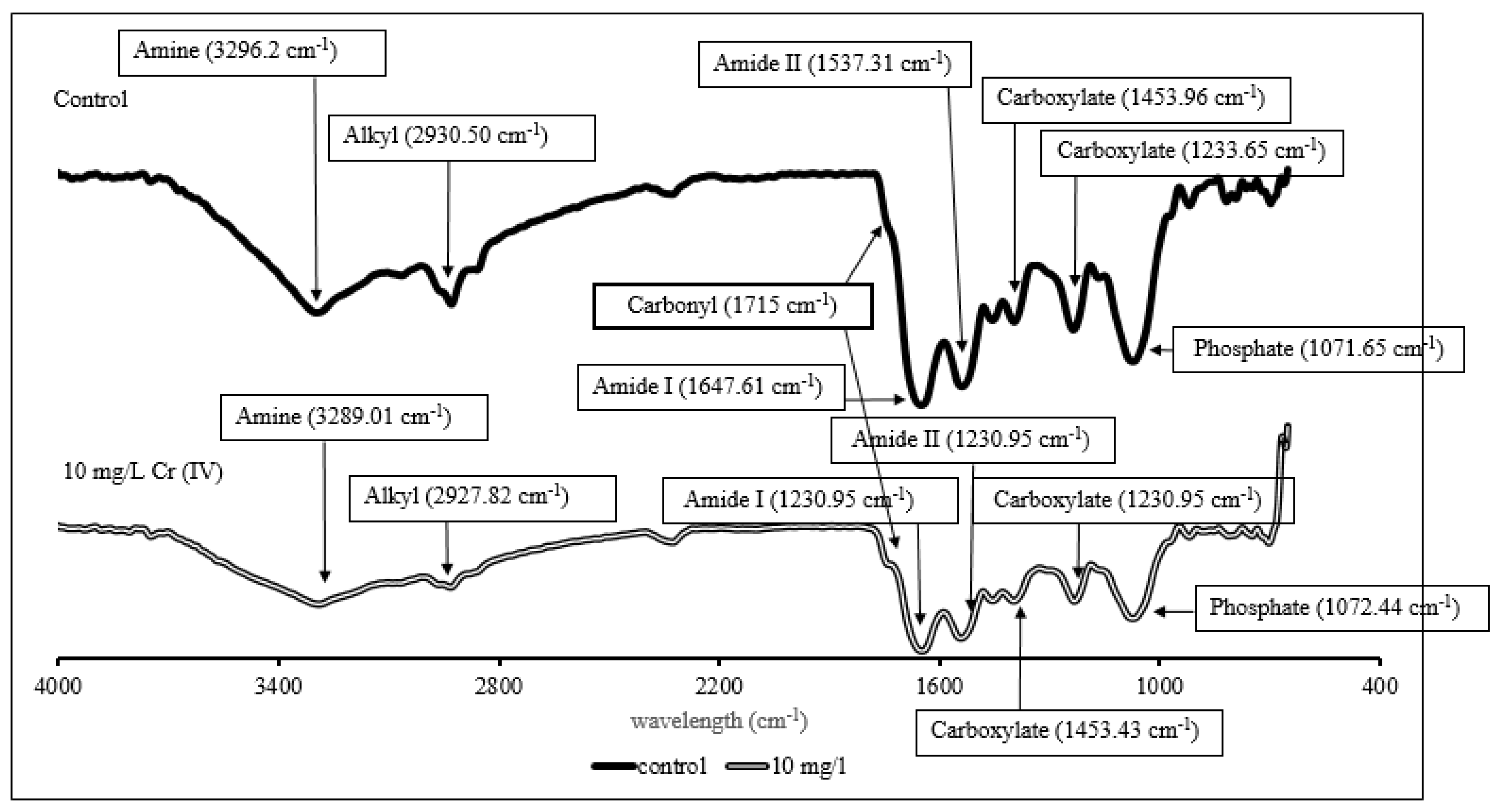
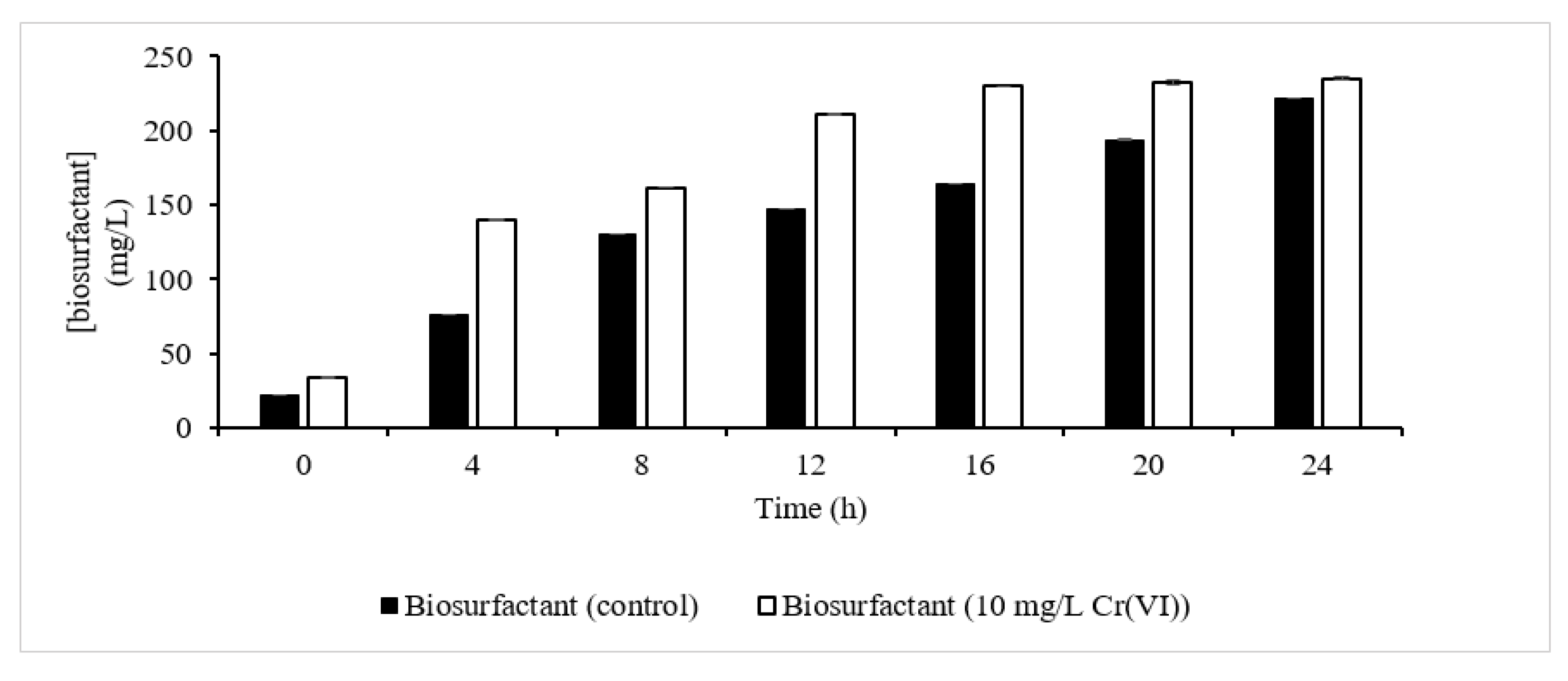
3.3. Biosurfactant Quantification and Characterization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Huang, H.; Wu, K.; Khan, A.; Jiang, Y.; Ling, Z.; Liu, P.; Chen, Y.; Tao, X.; Li, X. A novel Pseudomonas gessardii strain LZ-E simultaneously degrades naphthalene and reduces hexavalent chromium. Bioresour. Technol. 2016, 207, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Dula, T.; Siraj, K.; Kitte, S.A. Adsorption of Hexavalent Chromium from Aqueous Solution Using Chemically Activated Carbon Prepared from Locally Available Waste of Bamboo (Oxytenanthera abyssinica). ISRN Environ. Chem. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Hargreaves, A.J.; Vale, P.; Whelan, J.; Alibardi, L.; Constantino, C.; Dotro, G.; Cartmell, E.; Campo, P. Coagulation—Flocculation process with metal salts, synthetic polymers and biopolymers for the removal of trace metals (Cu, Pb, Ni, Zn) from municipal wastewater. Clean Technol. Environ. Policy 2018, 20, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Khulbe, K.C.; Matsuura, T. Removal of heavy metals and pollutants by membrane adsorption techniques. Appl. Water Sci. 2018, 8, 19. [Google Scholar] [CrossRef] [Green Version]
- Jobby, R.; Jha, P.; Yadav, A.K.; Desai, N. Chemosphere Biosorption and biotransformation of hexavalent chromium [Cr(VI)]: A comprehensive review. Chemosphere 2018, 207, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Barrera-díaz, C.E.; Lugo-lugo, V.; Bilyeu, B. A review of chemical, electrochemical and biological methods for aqueous Cr(VI) reduction. J. Hazard. Mater. 2012, 223–224, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, D.; Sukla, L.B.; Sawyer, M.; Rahman, P.K.S.M. Recent bioreduction of hexavalent chromium in wastewater treatment: A review. J. Ind. Eng. Chem. 2017, 55, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Patra, R.C.; Malik, S.; Beer, M.; Megharaj, M.; Naidu, R. Molecular characterization of chromium (VI) reducing potential in Gram positive bacteria isolated from contaminated sites. Soil Biol. Biochem. 2010, 42, 1857–1863. [Google Scholar] [CrossRef]
- Shukla, V.Y.; Tipre, D.R.; Dave, S.R. Optimization of Chromium(VI) Detoxification by Pseudomonas aeruginosa and Its Application for Treatment of Industrial Waste and Contaminated Soil. Bioremediat. J. 2014, 18, 128–135. [Google Scholar] [CrossRef]
- Chong, T.M.; Yin, W.F.; Mondy, S.; Grandclément, C.; Dessaux, Y.; Chan, K.G. Heavy-metal resistance of a France Vineyard soil bacterium, pseudomonas mendocina strain S5.2, revealed by whole-genome sequencing. J. Bacteriol. 2012, 194, 6366. [Google Scholar] [CrossRef] [Green Version]
- Suthersan, S.S. In situ bioremediation. In Remediation Engineering: Desing Concepts; CRC Press LLC: Boca Raton, FL, USA, 1999; pp. 123–158. ISBN 1566701376. [Google Scholar]
- Gupta, P.; Diwan, B. Bacterial Exopolysaccharide mediated heavy metal removal: A Review on biosynthesis, mechanism and remediation strategies. Biotechnol. Reports 2017, 13, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Zeng, G.M.; Niu, Q.Y.; Liu, Y.; Zhou, L.; Jiang, L.H.; Tan, X.; Xu, P.; Zhang, C.; Cheng, M. Bioremediation mechanisms of combined pollution of PAHs and heavy metals by bacteria and fungi: A mini review. Bioresour. Technol. 2017, 224, 25–33. [Google Scholar] [CrossRef]
- Kenney, J.P.L.; Fein, J.B. Importance of extracellular polysaccharides on proton and Cd binding to bacterial biomass: A comparative study. Chem. Geol. 2011, 286, 109–117. [Google Scholar] [CrossRef]
- Nordin, N.; Zakaria, M.R.; Halmi, M.I.E.; Ariff, A.; Zawawi, R.M.; Wasoh, H. Isolation and screening of high efficiency biosurfactant-producing Pseudomonas sp. J. Biochem. Microbiol. Biotechnol. 2013, 1, 25–31. [Google Scholar]
- Ozturk, S.; Kaya, T.; Aslim, B.; Tan, S. Removal and reduction of chromium by Pseudomonas spp. and their correlation to rhamnolipid production. J. Hazard. Mater. 2012, 231–232, 64–69. [Google Scholar] [CrossRef]
- Rizvi, A.; Khan, M.S. Biotoxic impact of heavy metals on growth, oxidative stress and morphological changes in root structure of wheat (Triticum aestivum L.) and stress alleviation by Pseudomonas aeruginosa strain CPSB1. Chemosphere 2017, 185, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Frimmersdorf, E.; Horatzek, S.; Pelnikevich, A.; Wiehlmann, L.; Schomburg, D. How Pseudomonas aeruginosa adapts to various environments: A metabolomic approach. Environ. Microbiol. 2010, 12, 1734–1747. [Google Scholar] [CrossRef]
- Majumder, S.; Raghuvanshi, S.; Gupta, S. Sequestration of Zn2+ from aqueous solution using Klebsiella pneumoniae: Batch kinetics and continuous studies. J. Water Process Eng. 2016, 12, 89–104. [Google Scholar] [CrossRef]
- Posokhov, Y.O.; Kyrychenko, A. Effect of acetone accumulation on structure and dynamics of lipid membranes studied by molecular dynamics simulations. Comput. Biol. Chem. 2013, 46, 23–31. [Google Scholar] [CrossRef] [PubMed]
- APHA. Standard Methods for the Examination of Water and Wastewater, 19th ed.; APHA: Washington, DC, USA, 2005; Volume 19. [Google Scholar]
- Moreno-Garrido, I.; Ruttkay-Nedecky, B.; Nejdl, L.; Gumulec, J.; Zitka, O.; Masarik, M.; Eckschlager, T.; Stiborova, M.; Adam, V.; Kizek, R.; et al. Microbial responses to environmental arsenic. Bioresour. Technol. 2016, 12, 557–561. [Google Scholar] [CrossRef]
- Smyth, T.J.P.; Rudden, M.; Tsaousi, K.; Banat, I.M. Protocols for the Detection and Chemical Characterisation of Microbial Glycolipids; Springer-Verlag: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Saikia, R.R.; Deka, S.; Deka, M.; Sarma, H. Optimization of environmental factors for improved production of rhamnolipid biosurfactant by Pseudomonas aeruginosa RS29 on glycerol. J. Basic Microbiol. 2012, 52, 446–457. [Google Scholar] [CrossRef]
- Kiliç, N.K.; Stensballe, A.; Otzen, D.E.; Dönmez, G. Proteomic changes in response to chromium(VI) toxicity in Pseudomonas aeruginosa. Bioresour. Technol. 2010, 101, 2134–2140. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.C.; Lin, B.C.; Wu, C.H. Biofilm formation and heavy metal resistance by an environmental Pseudomonas sp. Biochem. Eng. J. 2013, 78, 132–137. [Google Scholar] [CrossRef]
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in Shape: The Impact of Cell Shape on Bacterial Survival in Diverse Environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department of Environment Malaysia. Laporan Kualiti Alam Sekeliling Environmental Quality Report; Department of Environment Malaysia: Kuala Lumpur, Malaysia, 2019.
- Ahemad, M. Bacterial mechanisms for Cr(VI) resistance and reduction: An overview and recent advances. Folia Microbiol. 2014, 59, 321–332. [Google Scholar] [CrossRef]
- González, A.G.; Shirokova, L.S.; Pokrovsky, O.S.; Emnova, E.E.; Martínez, R.E.; Santana-Casiano, J.M. Adsorption of copper on Pseudomonas aureofaciens: Protective role of surface exopolysaccharides. J. Colloid Interface Sci. 2010, 350, 305–314. [Google Scholar] [CrossRef]
- Chug, R.; Gour, V.S.; Mathur, S.; Kothari, S.L. Optimization of Extracellular Polymeric Substances production using Azotobacter beijreinckii and Bacillus subtilis and its application in chromium (VI) removal. Bioresour. Technol. 2016, 214, 604–608. [Google Scholar] [CrossRef]
- Bezza, F.A.; Nkhalambayausi Chirwa, E.M. Biosurfactant-enhanced bioremediation of aged polycyclic aromatic hydrocarbons (PAHs) in creosote contaminated soil. Chemosphere 2016, 144, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Won, S.W.; Choi, S.B.; Chung, B.W.; Park, D.; Park, J.M.; Yun, Y. Biosorptive Decolorization of Reactive Orange 16 Using the Waste Biomass of Corynebacterium glutamicum. Ind. Eng. Chem. Res 2004, 43, 7865–7869. [Google Scholar] [CrossRef]
- Anjos, O.; Campos, M.G.; Ruiz, P.C.; Antunes, P. Application of FTIR-ATR spectroscopy to the quantification of sugar in honey. Food Chem. 2015, 169, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Kang, Y.; Feng, Y. Bioassessment of heavy metal toxicity and enhancement of heavy metal removal by sulfate-reducing bacteria in the presence of zero valent iron. J. Environ. Manag. 2017, 203, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Durve, A.; Chandra, N. FT-IR analysis of bacterial biomass in response to heavy metal stress. Int. J. Biotechnol. 2014, 112, 386–391. [Google Scholar]
- Neilson, J.W.; Zhang, L.; Veres-Schalnat, T.A.; Chandler, K.B.; Neilson, C.H.; Crispin, J.D.; Pemberton, J.E.; Maier, R.M. Cadmium effects on transcriptional expression of rhlB/rhlC genes and congener distribution of monorhamnolipid and dirhamnolipid in Pseudomonas aeruginosa IGB83. Appl. Microbiol. Biotechnol. 2010, 88, 953–963. [Google Scholar] [CrossRef]
- Varjani, S.J.; Upasani, V.N. Critical review on biosurfactant analysis, purification and characterization using rhamnolipid as a model biosurfactant. Bioresour. Technol. 2017, 232, 389–397. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mat Arisah, F.; Amir, A.F.; Ramli, N.; Ariffin, H.; Maeda, T.; Hassan, M.A.; Mohd Yusoff, M.Z. Bacterial Resistance against Heavy Metals in Pseudomonas aeruginosa RW9 Involving Hexavalent Chromium Removal. Sustainability 2021, 13, 9797. https://doi.org/10.3390/su13179797
Mat Arisah F, Amir AF, Ramli N, Ariffin H, Maeda T, Hassan MA, Mohd Yusoff MZ. Bacterial Resistance against Heavy Metals in Pseudomonas aeruginosa RW9 Involving Hexavalent Chromium Removal. Sustainability. 2021; 13(17):9797. https://doi.org/10.3390/su13179797
Chicago/Turabian StyleMat Arisah, Fatini, Amirah Farhana Amir, Norhayati Ramli, Hidayah Ariffin, Toshinari Maeda, Mohd Ali Hassan, and Mohd Zulkhairi Mohd Yusoff. 2021. "Bacterial Resistance against Heavy Metals in Pseudomonas aeruginosa RW9 Involving Hexavalent Chromium Removal" Sustainability 13, no. 17: 9797. https://doi.org/10.3390/su13179797