
Herbicide Resistance to Metsulfuron-Methyl in Rumex dentatus L. in North-West India and Its Management Perspectives for Sustainable Wheat Production

 , , , ,
, , , ,
Abstract
:1. Introduction
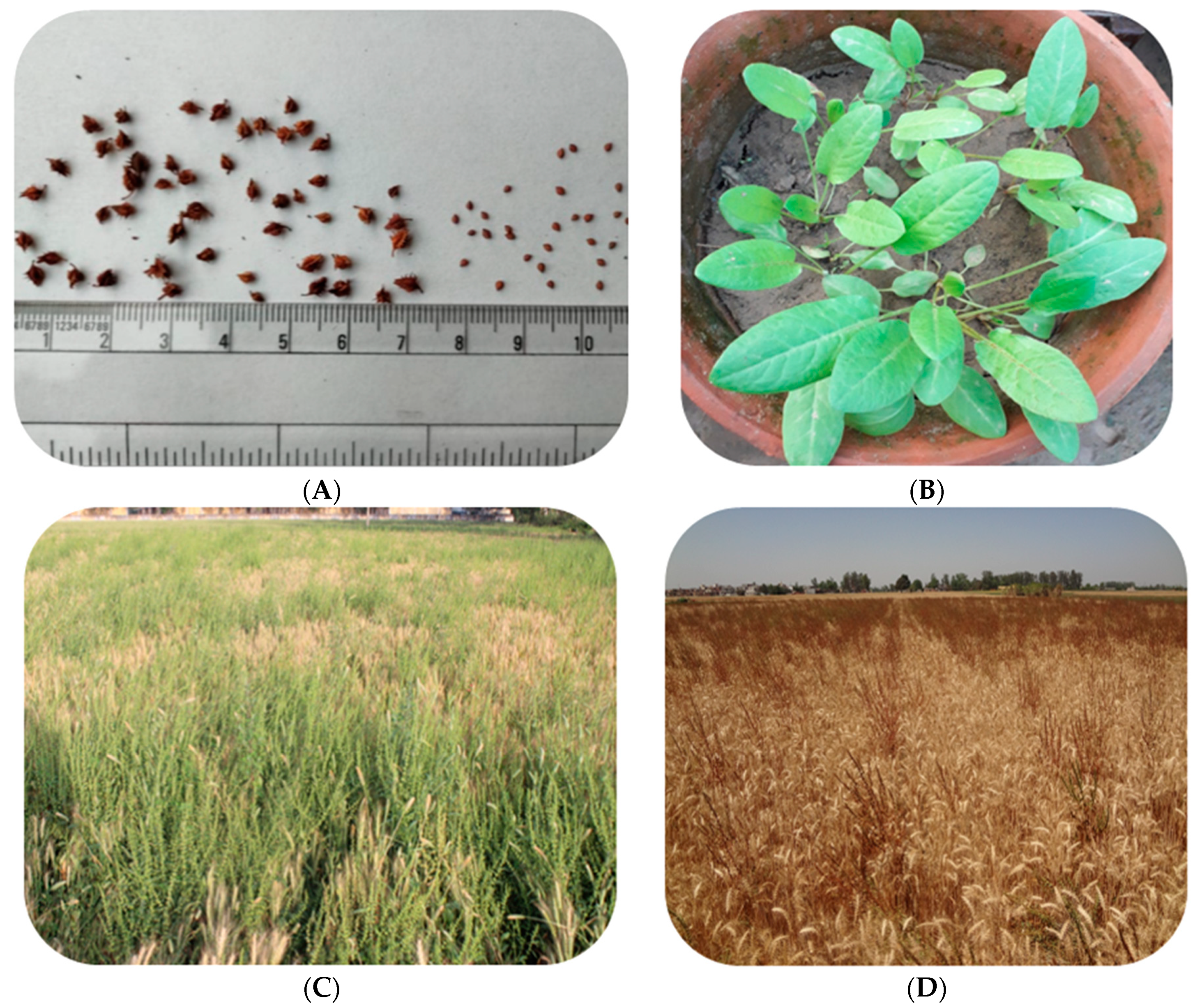
2. Economic Importance, Biology and Ecology of Rumex dentatus
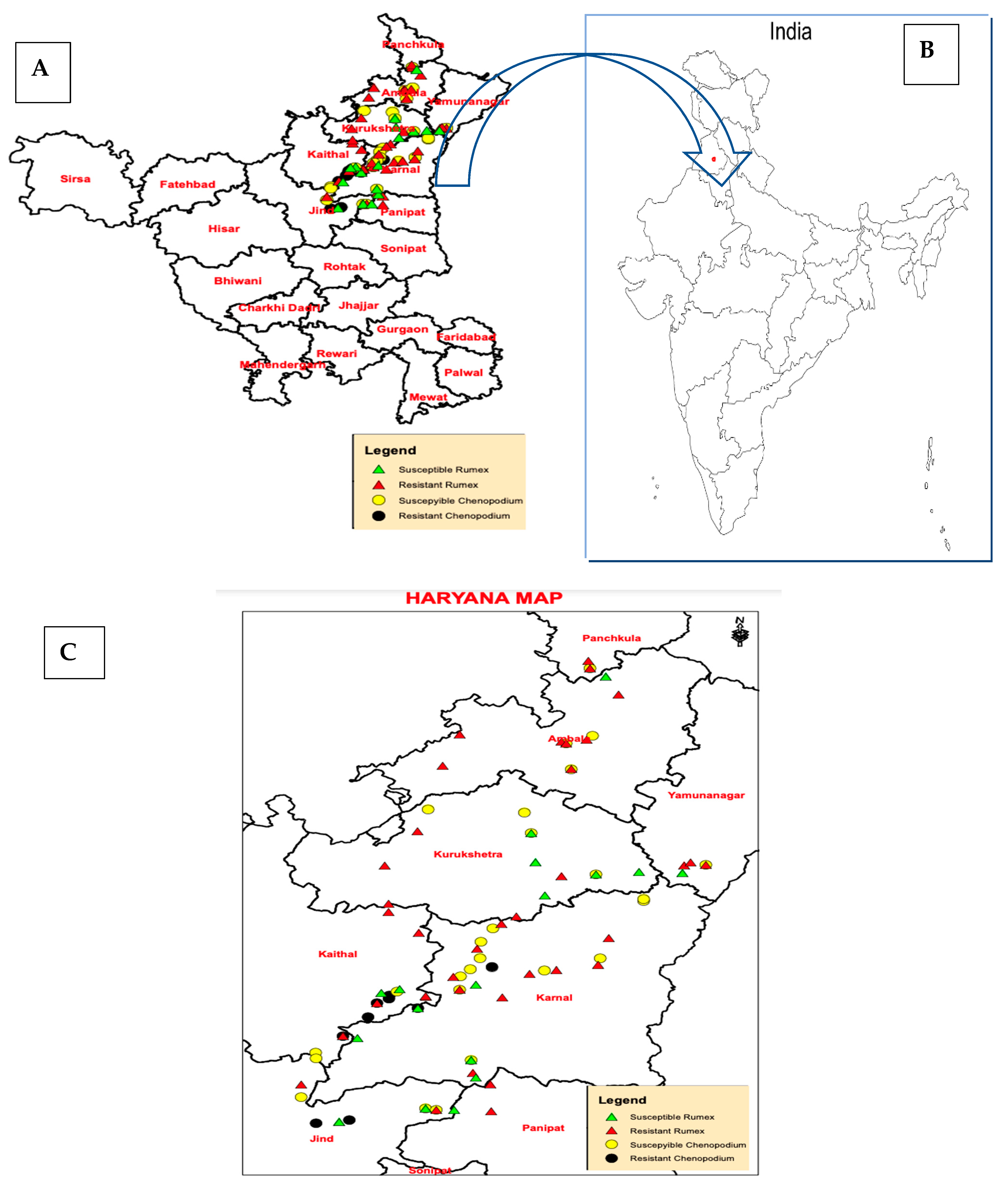
3. Herbicide Resistance in Rumex dentatus
4. Agronomic Practices Influencing Rumex dentatus Infestation in Wheat
4.1. Tillage
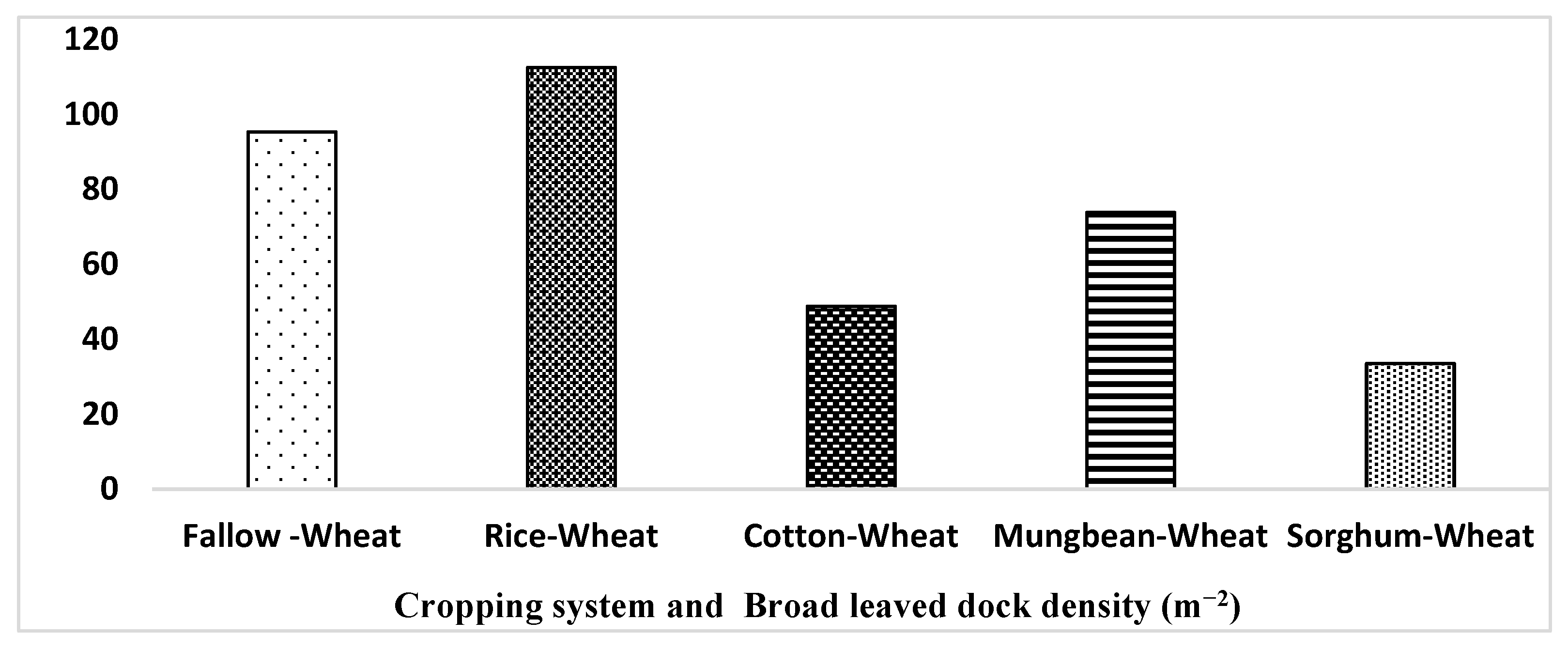
4.2. Crop Rotation
4.3. Paddy Straw Management (Burning/Retention)
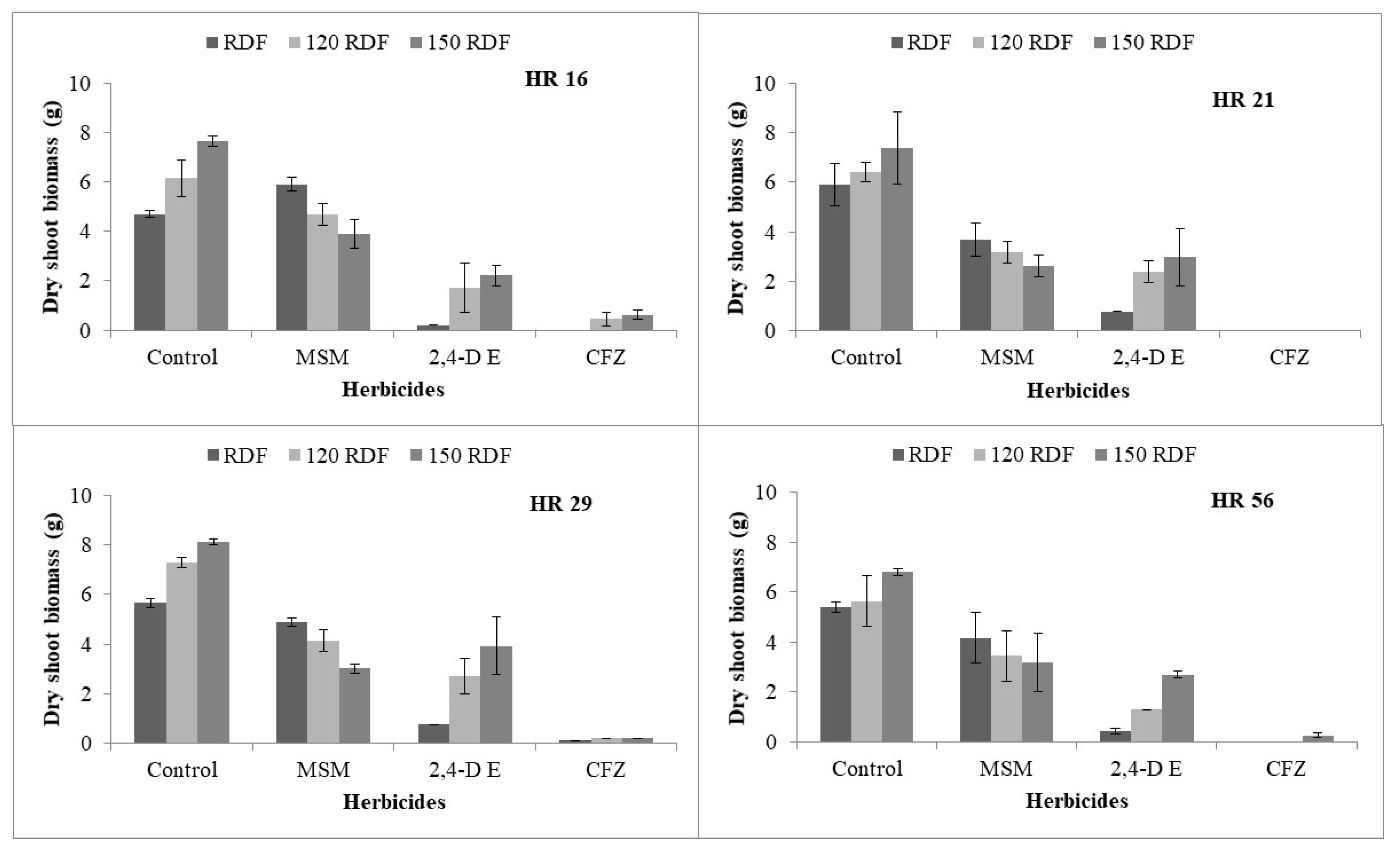
4.4. Nutrient Management
4.5. Flooding/Irrigation Management
5. Management of Metsulfuron-Methyl-Resistant Rumex dentatus
6. Climate Change and Weed Dynamics
7. Future Prospects
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gupta, R.K.; Naresh, R.K.; Hobbs, P.R.; Jiaguo, Z.; Ladha, J.K. Sustainability of Post-Green Revolution Agriculture: The Rice-Wheat Cropping Systems of the Indo-Gangetic Plains and China. In Animal Manure; Wiley: Hoboken, NJ, USA, 2015; Volume 65, pp. 1–25. [Google Scholar]
- Gupta, R.; Seth, A. A review of resource conserving technologies for sustainable management of the rice-wheat cropping systems of the Indo-Gangetic plains (IGP). Crop Prot. 2007, 26, 436–447. [Google Scholar] [CrossRef]
- Timsina, J.; Connor, D.J. Productivity and management of rice–wheat cropping systems: Issues and challenges. Field Crops Res. 2001, 69, 93–132. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Mahajan, G.; Sardana, V.; Timsina, J.; Jat, M.L. Productivity and sustainability of the rice-wheat cropping system in the Indo-Gangetic Plains of the Indian subcontinent: Problems, opportunities, and strategies. Adv. Agron. 2012, 117, 315–369. [Google Scholar]
- Ladha, J.K.; Dawe, D.; Pathak, H.; Padre, A.T.; Yadav, R.L.; Singh, B.; Sakal, R. How extensive are yield declines in long-term rice–wheat experiments in Asia? Field Crops Res. 2003, 81, 159–180. [Google Scholar] [CrossRef]
- Dey, A.; Dinesh; Rashmi. Rice and wheat production in India: An overtime study on growth and instability. J. Pharmacogn. Phytochem. 2020, 9, 158–161. [Google Scholar]
- Humphreys, E.; Kukal, S.S.; Christen, E.W.; Hira, G.S.; Sharma, R.K. Halting the groundwater decline in north-west India-which crop technologies will be winners? Adv. Agron. 2010, 109, 155–217. [Google Scholar]
- Chaudhary, A.; Chhokar, R.S.; Yadav, D.B.; Sindhu, V.K.; Ram, H.; Rawal, S.; Khedwal, R.S.; Sharma, R.K.; Gill, S.C. In-situ paddy straw management practices for higher resource use efficiency and crop productivity in Indo-Gangetic Plains (IGP) of India. J. Cereal Res. 2019, 11, 172–198. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.K.; Sahai, S.; Singh, N.; Dixit, C.K.; Singh, D.P.; Sharma, C.; Garg, S.C. Residue burning in rice–wheat cropping system: Causes and implications. Curr. Sci. 2004, 87, 1713–1717. [Google Scholar]
- Kumar, V.; Ladha, J.K. Direct seeding of rice: Recent developments and future research needs. Adv. Agron. 2011, 111, 297–413. [Google Scholar]
- Chhokar, R.S.; Chaudhary, A.; Sharma, R.K. Herbicide resistant weeds in India and their management. In Fifty Years of Weed Science Research in India; Kumar, S., Mishra, J.S., Eds.; Indian Society of Weeds Science: Jabalpur, India, 2018; pp. 288–308. [Google Scholar]
- Bhullar, M.S.; Punia, S.S.; Tomar, S.S.; Singh, V.P.; Sharma, J.D. Little seed canary grass resistance to clodinafop in Punjab: Farmers’ perspective. Indian J. Weed Sci. 2014, 46, 237–240. [Google Scholar]
- Malik, R.K.; Singh, S. Littleseed canarygrass (Phalaris minor) resistance to isoproturon in India. Weed Technol. 1995, 9, 419–425. [Google Scholar] [CrossRef]
- Yaduraju, N.T. Control of herbicide resistant Phalaris minor: Need for a sound weed management system. Pestology 1999, 264–270. [Google Scholar]
- Yaduraju, N.T. Weed management perspectives for India in changing agriculture scenario in country. Pak. J. Weed Sci. Res. 2012, 18, 703–710. [Google Scholar]
- Duke, S.O. Why have no new herbicide mode of action appeared in recent years? Pest Manag. Sci. 2012, 68, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Chhokar, R.S.; Sharma, R.K.; Sharma, I. Weed management strategies in wheat—A review. J. Wheat Res. 2012, 4, 1–21. [Google Scholar]
- Kaur, S.; Kaur, R.; Chauhan, B.S. Understanding crop-weed-fertilizer-water interactions and their implications for weed management in agricultural systems. Crop Prot. 2018, 103, 65–72. [Google Scholar] [CrossRef]
- Mongia, A.D.; Sharma, R.K.; Kharub, A.S.; Tripathi, S.C.; Chhokar, R.S.; Shoran, J. Coordinated Research on Wheat Production Technology in India; Directorate of Wheat Research Bulletin: Karnal, India, 2005. [Google Scholar]
- Chhokar, R.S.; Malik, R.K. Isoproturon-resistant littleseed canarygrass (Phalaris minor) and its response to alternate herbicides. Weed Technol. 2002, 16, 116–123. [Google Scholar] [CrossRef]
- Chhokar, R.S.; Sharma, R.K.; Jat, G.R.; Pundir, A.K.; Gathala, M.K. Effect of tillage and herbicides on weeds and productivity of wheat under rice–wheat growing system. Crop Prot. 2007, 26, 1689–1696. [Google Scholar] [CrossRef]
- Singh, S. Role of management practices on control of isoproturon-resistant littleseed canarygrass (Phalaris minor) in India. Weed Technol. 2007, 21, 339–346. [Google Scholar] [CrossRef]
- Singh, S.; Punia, S.S. Effect of seeding depth and flooding on emergence of Malva parviflora, Rumex dentatus and R. spinosus. Indian J. Weed Sci. 2008, 40, 178–186. [Google Scholar]
- Umm-e-Kulsoom; Gul, J.; Bakht, T.; Kanwal, S. Increasing densities of Avena fatua and Rumex dentatus reduce the yield of wheat under field conditions. Pak. J. Weed Sci. Res. 2018, 24, 155–164. [Google Scholar] [CrossRef]
- Waheed, Z.; Usman, K.; Ali, I. Response of wheat to varying densities of Rumex dentatus under irrigated condition of Dera Ismail Khan, Pakistan. Sarhad J. Agric. 2017, 33, 1–7. [Google Scholar] [CrossRef]
- Anjum, T.; Bajwa, R. Competition losses caused by Rumex dentatus L. and Chenopodium album L. in wheat (Triticum aestivum L.). Philipp. Agric. Sci. 2010, 93, 365–368. [Google Scholar]
- Siddiqui, I.; Bajwa, R.; Javaid, A. Effect of six problematic weeds on growth and yield of wheat. Pak. J. Bot. 2010, 42, 2461–2471. [Google Scholar]
- Singh, V.; Singh, H.; Raghubanshi, A.S. Competitive interactions of wheat with Phalaris minor or Rumex dentatus: A replacement series study. Int. J. Pest Manag. 2013, 59, 245–258. [Google Scholar] [CrossRef]
- Vasas, A.; Orbán-Gyapai, O.; Hohmann, J. The Genus Rumex: Review of traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2015, 175, 198–228. [Google Scholar] [CrossRef]
- Munir, M.A.; Ahmad, M.; Ali, M.I.; Mahmood, Z.; Afzal, M.; Sharif, M.N.; Aslam, M. Correlation and regression analysis of morphological traits in Rumex dentatus. Bull. Biol. Allied Sci. Res. 2016, 1, 26–38. [Google Scholar]
- Bhargava, H.; Sawhney, R. Morphological studies in Polygonaceae. I. Contribution to the life-history of Rumex dentatus Linn. J. Univ. Saugar. 1958, 7, 21–38. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. Physiology of dormancy and germination in relation to seed bank ecology. In Ecology of Soil Seed Banks; Leck, M.A., Parker, V.T., Simpson, R.L., Eds.; Academic Press: San Diego, CA, USA, 1989; pp. 53–66. [Google Scholar]
- Dhawan, R.S. Studies on germination and emergence of Rumex maritimus. Indian J. Weed Sci. 2005, 37, 144–146. [Google Scholar]
- Al-Helal, A.A. Studies on germination of Rumex dentatus L. seeds. J. Arid Environ. 1996, 33, 39–47. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heap, I. The International Herbicide-Resistant Weed Database. Available online: www.weedscience.org (accessed on 19 April 2021).
- Franke, A.C. Ecological and Socio-Economical Aspects of Phalaris minor Epidemic in the Rice-Wheat System in Haryana. Ph.D. Thesis, Scottish Agricultural College, GIasgow University, Glasgow, UK, 2002. [Google Scholar]
- Singh, S.; Malik, R.K.; Balyan, R.S.; Singh, S. Distribution of weed flora of wheat in Haryana. Indian J. Weed Sci. 1995, 27, 114–121. [Google Scholar]
- Walia, U.S.; Brar, L.S.; Dhaliwal, B.K. Resistance to isoproturon in Phalaris minor Retz. In Punjab. Plant Prot. Q. 1997, 12, 138–140. [Google Scholar]
- Dhawan, R.S.; Punia, S.S.; Singh, S.; Yadav, D.; Malik, R.K. Productivity of wheat (Triticum aestivum) as affected by continuous use of new low dose herbicides for management of littleseed canarygrass (Phalaris minor). Indian J. Agron. 2009, 54, 58–62. [Google Scholar]
- Chhokar, R.S.; Sharma, R.K. Multiple herbicide resistance in littleseed canarygrass (Phalaris minor): A threat to wheat production in India. Weed Biol. Manag. 2008, 8, 112–123. [Google Scholar] [CrossRef]
- Yadav, A.; Malik, R.K. Herbicide resistant Phalaris minor in wheat–A sustainability issue. In Resource Book; Department of Agronomy and Directorate of Extension Education CCSHAU: Hisar, India, 2005. [Google Scholar]
- Jena, T.; Singh, R.K.; Bisen, N. Surfactant influence on efficacy of herbicides in barley. Indian J. Weed Sci. 2018, 50, 56–58. [Google Scholar] [CrossRef] [Green Version]
- Palsaniya, D.R.; Kumar, T.K.; Prabhu, G.; Dixit, A.K.; Rai, A.K.; Kumar, S. Weed dynamics in fodder oat (Avena sativa L.) genotypes. Range Manag. Agrofor. 2015, 36, 107–108. [Google Scholar]
- Singh, R. Effect of cropping sequence, seed rate and weed management on weed growth and yield of Indian mustard in western Rajasthan. Indian J. Weed Sci. 2006, 38, 69–72. [Google Scholar]
- Singh, S. Studies on weed management in berseem (Trifolium alexandrinum L.). Haryana J. Agron. 2012, 28, 77–80. [Google Scholar]
- Chaudhary, A.; Kaur, S. ALS/AHAS (Group B/2) inhibitors resistance in Rumex dentatus L. and Chenopodium album L. halting future wheat productivity of Haryana and Punjab. In Proceedings of the ISWS Golden Jubilee International Conference on “Weeds and Society: Challenges and Opportunities”, ICAR-DWR, Jabalpur, India, 21–24 November 2018; Indian Society of Weed Science: Jabalpur, India, 2018; p. 428. [Google Scholar]
- Chhokar, R.S.; Sharma, R.K.; Garg, R.; Sharma, I. Metsulfuron resistance in Rumex dentatus. In Wheat and Barley Newsletter; Directorate of Wheat Research: Karnal, India, 2013. [Google Scholar]
- Dhanda, S.; Chaudhary, A.; Kaur, S.; Bhullar, M.S. Herbicide resistance in Rumex dentatus against metsulfuron herbicide in Punjab and Haryana, India. Indian J. Weed Sci. 2020, 52, 259–264. [Google Scholar]
- Singh, S. Herbicide resistance in Rumex dentatus L. (toothed dock) confirmed. IWSS Newsl. 2016, 13–17. Available online: https://www.iwss.info/downloads/files/n5a40c6670eb87.pdf (accessed on 15 April 2021).
- Yadav, D.B.; Punia, S.S.; Singh, N.; Garg, R. Herbicide resistance in toothed dock population from Haryana. In Proceedings of the Biennial Conference of the Indian Society of Weed Science on “Doubling Farmers’ Income by 2022: The Role of Weed Science” MPUA&T, Udaipur, India, 1–3 March 2017; p. 63. [Google Scholar]
- Chhokar, R.S.; Sharma, R.K.; Gill, S.C.; Meena, R.P. Herbicides for broad-leaved weeds management in wheat. Indian J. Weed Sci. 2015, 47, 353–361. [Google Scholar]
- Chhokar, R.S.; Sharma, R.K.; Gill, S.C.; Singh, R.; Singh, G.P. Management of herbicide resistant weeds for sustainable wheat production. In Proceedings of the Biennial Conference of the Indian Society of Weed Science on “Doubling Farmers’ Income by 2022: The Role of Weed Science” MPUA&T, Udaipur, India, 1–3 March 2017; p. 63. [Google Scholar]
- Tranel, P.J.; Wright, T.R. Resistance of weeds to ALS-inhibiting herbicides: What have we learned? Weed Sci. 2002, 50, 700–712. [Google Scholar] [CrossRef]
- Yadav, A.; Malik, R.K.; Gill, G.; Singh, S.; Chauhan, B.S.; Bellinder, R.R. Current status of weed resistance to herbicides in rice-wheat cropping system in Haryana and its management. Indian J. Weed Sci. 2006, 38, 194–206. [Google Scholar]
- Buhler, D.D. Effects of tillage and light environment on emergence of 13 annual weeds. Weed Technol. 1997, 11, 496–501. [Google Scholar] [CrossRef]
- Buhler, D.D. Influence of tillage systems on weed population dynamics and management in corn and soybean in the central USA. Crop Sci. 1995, 35, 1247–1258. [Google Scholar] [CrossRef]
- Froud-Williams, R.J.; Chancellor, R.J.; Drennan, D.S.H. Influence of cultivation regime upon buried weed seeds in arable cropping systems. J. Appl. Ecol. 1983, 20, 199–208. [Google Scholar] [CrossRef]
- Franke, A.C.; Singh, S.; Mc Roberts, N.; Nehra, A.S.; Godara, S.; Malik, R.K.; Marshall, G. Phalaris minor seedbank studies: Longevity, seedling emergence and seed production as affected by tillage regime. Weed Res. 2007, 47, 73–83. [Google Scholar] [CrossRef]
- Chhokar, R.S.; Singh, S.; Sharma, R.K.; Singh, M. Influence of straw management on Phalaris minor Retz. control. Indian J. Weed Sci. 2009, 41, 150–156. [Google Scholar]
- Mishra, J.S.; Singh, V.P. Tillage and weed control effects on productivity of a dry seeded rice–wheat system on a vertisol in Central India. Soil Tillage Res. 2012, 123, 11–20. [Google Scholar] [CrossRef]
- Sharma, R.K.; Babu, K.S.; Chhokar, R.S.; Sharma, A.K. Effect of tillage on termites, weed incidence and productivity of spring wheat in rice–wheat system of North Western Indian plains. Crop Prot. 2004, 23, 1049–1054. [Google Scholar] [CrossRef]
- Brar, A.S.; Walia, U.S. Weed dynamics and wheat (Triticum aestivum L.) productivity as influenced by planting techniques and weed control practices. Indian J. Weed Sci. 2009, 41, 161–166. [Google Scholar]
- Liebman, M.; Dyck, E. Crop rotation and intercropping strategies for weed management. Ecol. Appl. 1993, 3, 92–122. [Google Scholar] [CrossRef]
- Shahzad, M.; Jabran, K.; Hussain, M.; Raza, M.A.S.; Wijaya, L.; El-Sheikh, M.A.; Alyemeni, M.N. The impact of different weed management strategies on weed flora of wheat-based cropping systems. PLoS ONE 2021, 16, e0247137. [Google Scholar] [CrossRef]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crops Res. 2015, 183, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Singh, S.; Chhokar, R.S.; Malik, R.K.; Brainard, D.C.; Ladha, J.K. Weed management strategies to reduce herbicide use in zero-till rice–wheat cropping systems of the Indo-Gangetic Plains. Weed Technol. 2013, 27, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Blackshaw, R.E.; Molnar, L.J.; Janzen, H.H. Nitrogen fertilizer timing and application method affect weed growth and competition with spring wheat. Weed Sci. 2004, 52, 614–622. [Google Scholar] [CrossRef]
- Singh, V.; Singh, H. Effect of nitrogen application on biomass and ecophysiology of Phalaris minor and Rumex dentatus in wheat field of Indo-Gangetic plains. In Proceedings of the 25th Asian-Pacific Weed Science Society Conference, Hyderabad, India, 13–16 October 2015. [Google Scholar]
- Haggar, R.J. Survey of the incidence of docks (Rumex spp.) in grassland in ten districts in the United Kingdom in 1972. A.D.A.S. Quart. Rev. 1980, 39, 256–270. [Google Scholar]
- Gebauer, G.; Melzer, A.; Rehder, H. Nitrate content and nitrate reductase activity in Rumex obtusifolius L. Oecologia 1984, 63, 136–142. [Google Scholar] [CrossRef]
- Sarabi, V. Factors that influence the level of weed seed predation: A review. Weed Biol. Manag. 2019, 19, 61–74. [Google Scholar] [CrossRef]
- Iffat, S.; Rukhsana, B.; Arshad, J. Some factors affecting the pathogenicity of Alternaria alternata against the weed Rumex dentatus. Philipp. Agric. Sci. 2009, 92, 282–289. [Google Scholar]
- Mohammed, Y.M.; Badawy, M.E. Potential of phytopathogenic fungal isolates as a biocontrol agent against some weeds. Egyp. J. Biol. Pest. 2020, 30, 1–9. [Google Scholar] [CrossRef]
- Akbar, M.; Javaid, A. Prospects of using fungal metabolites for the management of Rumex dentatus, a problematic weed of wheat. Int. J. Agric. Biol. 2013, 15, 1277–1282. [Google Scholar]
- Anjum, T.; Bajwa, R. Field appraisal of herbicide potential of sunflower leaf extract against Rumex dentatus. Field Crops Res. 2007, 100, 139–142. [Google Scholar] [CrossRef]
- Chhokar, R.S.; Sharma, R.K.; Pundir, A.K.; Singh, R.K. Evaluation of herbicides for control of Rumex dentatus, Convolvulus arvensis and Malva parviflora. Indian J. Weed Sci. 2007, 39, 214–218. [Google Scholar]
- Kaur, S.; Kaur, T.; Bhullar, M.S. Control of mixed weed flora in wheat with sequential application of pre-and post-emergence herbicides. Indian J. Weed Sci. 2017, 49, 29–32. [Google Scholar] [CrossRef]
- Naseer-ud-Din, G.M.; Shehzad, M.A.; Nasrullah, H.M. Efficacy of various pre and post-emergence herbicides to control weeds in wheat. Pak. J. Agric. Sci. 2011, 48, 185–190. [Google Scholar]
- Sindhu, V.K.; Singh, S.; Punia, S.S.; Singh, S. Pre–emergence herbicides can tactically fit into conservation agriculture systems with various benefits. In Proceedings of the “Doubling Farmers’ Income by 2022: The Role of Weed Science” Biennial Conference of the Indian Society of Weed Science on MPUA&T, Udaipur, India, 1–3 March 2017. [Google Scholar]
- Das, T.K.; Ahlawat, I.P.; Yaduraju, N.T. Littleseed canarygrass (Phalaris minor) resistance to clodinafop-propargyl in wheat fields in north-western India: Appraisal and management. Weed Biol. Manag. 2014, 14, 11–20. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Ziska, L.H.; Teasdale, J.R.; Bunce, J.A. Future atmospheric carbon dioxide may increase tolerance to glyphosate. Weed Sci. 1999, 47, 608–615. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef]
- Valerio, M.; Tomecek, M.B.; Lovelli, S.; Ziska, L.H. Quantifying the effect of drought on carbon dioxide-induced changes in competition between a C3 crop (tomato) and a C4 weed (Amaranthus retroflexus). Weed Res. 2011, 51, 591–600. [Google Scholar] [CrossRef]
- Zhu, C.; Zeng, Q.; Ziska, L.H.; Zhu, J.; Xie, Z.; Liu, G. Effect of nitrogen supply on carbon dioxide-induced changes in Competition between Rice and Barnyard grass (Echinochloa crusgalli). Weed Sci. 2008, 56, 66–71. [Google Scholar] [CrossRef]
- Ziska, L.H.; Faulkner, S.; Lydon, J. Changes in biomass and root: Shoot ratio of field-grown Canada thistle (Cirsium arvense), a noxious, invasive weed, with elevated CO2: Implications for control with glyphosate. Weed Sci. 2004, 52, 584–588. [Google Scholar] [CrossRef]
- Varanasi, A.; Prasad, P.V.; Jugulam, M. Impact of climate change factors on weeds and herbicide efficacy. Adv. Agron. 2016, 135, 107–146. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Weeds | Resistance | Susceptible |
|---|---|---|
| Phalaris minor (Littleseed canarygrass) | Phenyl urea (Isoproturon), Sulfonylurea (sulfosulfuron, mesosulfuron), Aryloxyphenoxypropionic (Clodinafop), Cyclohexene oxime (Tralkoxydim), Phenylpyrazole (pinoxaden)and Triazolopyrimidine sulfonamide (pyroxsulam) | Flumioxazin, Pendimethalin, Metribuzin, Terbutryn, Flufenacet, and pyroxasulfone |
| Polypogon monspeliensis (Rabbitfoot grass) | Sulfonylurea (sulfosulfuron, mesosulfuron),Triazolopyrimidine sulfonamide (pyroxsulam) | Pendimethalin, Metribuzin Clodinafop, Fenoxaprop, Pinoxaden, Flufenacet and Pyroxasulfone |
| Rumex dentatus (Toothed dock) | Triazolopyrimidine sulfonamide (pyroxsulam, florasulam), Sulfonylurea (metsulfuron, triasulfuron, iodosulfuron) | 2,4-D, Carfentrazone, Pendimethalin, Flumioxazin Metribuzin & Terbutryn |
| Chenopodium album (Common lambsquarters) | Sulfonylurea (sulfosulfuron, metsulfuron) | 2,4-D, Carfentrazone, Flumioxazin |
| Avena ludoviciana (Wild oat) | Sulfonylurea (sulfosulfuron, mesosulfuron), Aryloxyphenoxypropionic (Clodinafop) | Pyroxasulfone, Flufenacet |
| Herbicide Name | Dose (g ha−1) | Mode of Action |
|---|---|---|
| 2, 4-D sodium salt | 500 | Synthetic auxin |
| 2, 4-D ethyl ester | 500 | Synthetic auxin |
| 2,4-D amine | 750 | Synthetic auxin |
| Carfentrazone ethyl | 20 | Protoporphyrinogen oxidase inhibitor |
| Metsulfuron methyl + Carfentrazone ethyl (premix) | 25 | ALS inhibitor + Protoporphyrinogen oxidase inhibitor |
| Bromoxynil + MCPA | 490 | Photosystem II inhibitor + synthetic auxin |
| Isoproturon | 1000–1250 | Photosystem II inhibitor |
| Metribuzin | 300 | Photosystem II inhibitor |
| Terbutryn | 1000 | Photosystem II inhibitor |
| Dicamba | 240–360 | Synthetic auxin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhary, A.; Chhokar, R.S.; Dhanda, S.; Kaushik, P.; Kaur, S.; Poonia, T.M.; Khedwal, R.S.; Kumar, S.; Punia, S.S. Herbicide Resistance to Metsulfuron-Methyl in Rumex dentatus L. in North-West India and Its Management Perspectives for Sustainable Wheat Production. Sustainability 2021, 13, 6947. https://doi.org/10.3390/su13126947
Chaudhary A, Chhokar RS, Dhanda S, Kaushik P, Kaur S, Poonia TM, Khedwal RS, Kumar S, Punia SS. Herbicide Resistance to Metsulfuron-Methyl in Rumex dentatus L. in North-West India and Its Management Perspectives for Sustainable Wheat Production. Sustainability. 2021; 13(12):6947. https://doi.org/10.3390/su13126947
Chicago/Turabian StyleChaudhary, Ankur, Rajender Singh Chhokar, Sachin Dhanda, Prashant Kaushik, Simerjeet Kaur, Todar Mal Poonia, Rajbir Singh Khedwal, Surender Kumar, and Satbir Singh Punia. 2021. "Herbicide Resistance to Metsulfuron-Methyl in Rumex dentatus L. in North-West India and Its Management Perspectives for Sustainable Wheat Production" Sustainability 13, no. 12: 6947. https://doi.org/10.3390/su13126947