Overexpression of Citrate Synthase Increases Isocitric Acid Biosynthesis in the Yeast Yarrowia lipolytica

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Growth Media and Conditions
2.3. General Genetic Techniques, Plasmid Construction and Transformation
2.4. Bioreactor Studies
2.5. Analytical Methods
2.6. Calculation
2.7. Bioinformatic Analysis
3. Results and Discussions
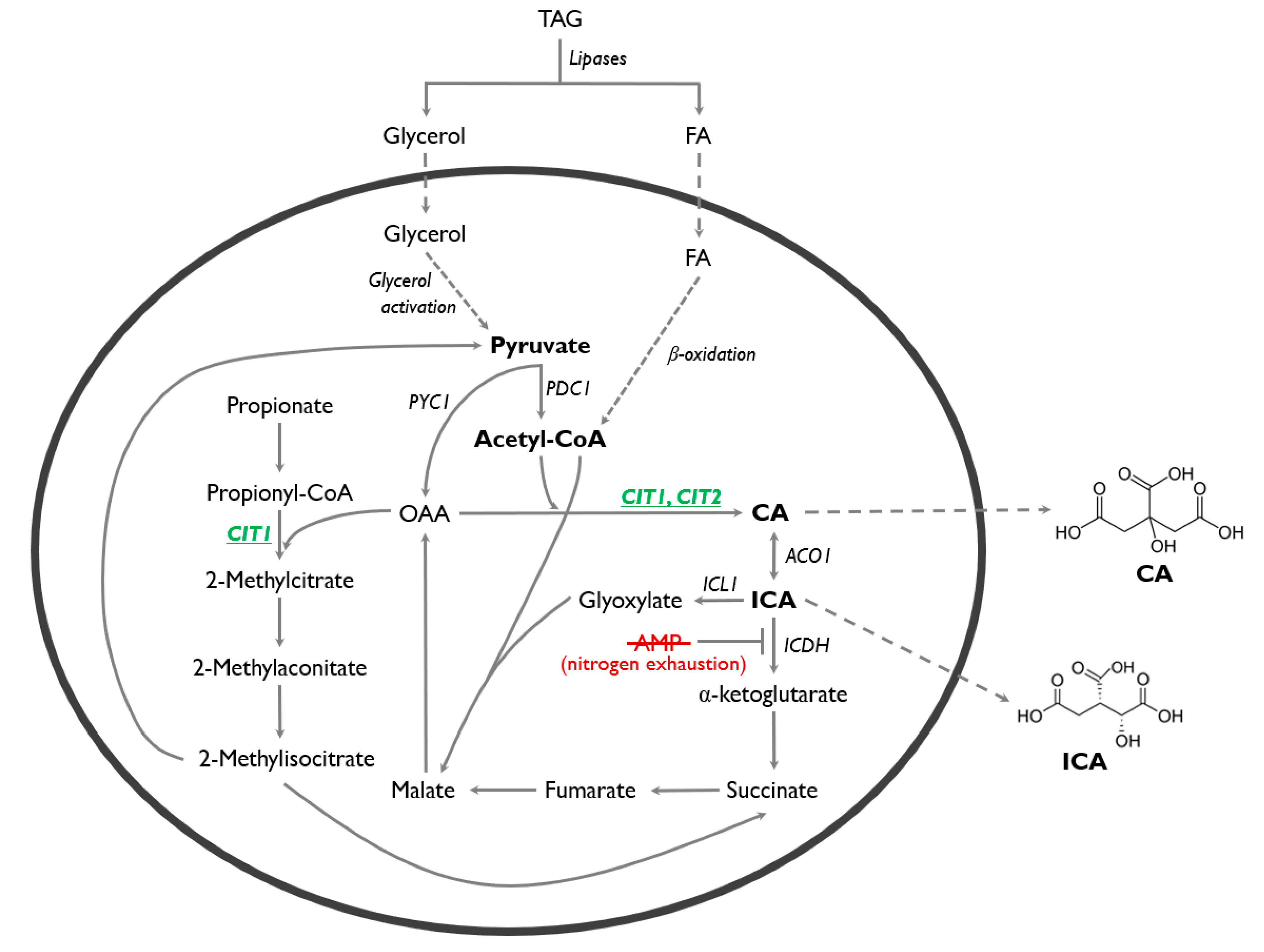
3.1. Genes Encoding Citrate Synthases in Y. lipolytica
3.2. CIT1 and CIT2 Overexpression Enhances Citrate Synthase Activity
3.3. CIT1 is Indispensable for Propionate Utilization in Y. lipolytica
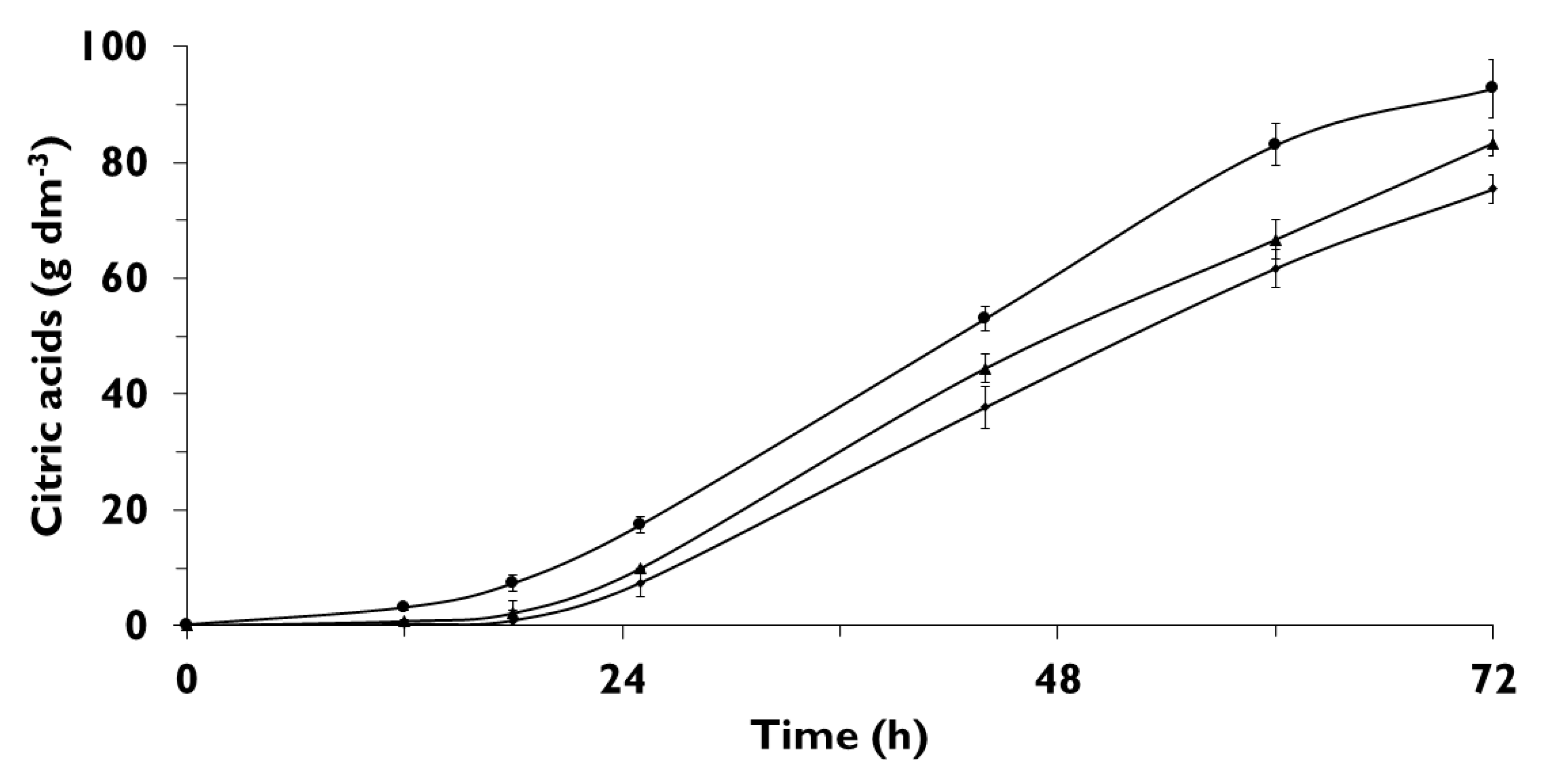
3.4. Overexpression of CIT1 or CIT2 Increases ICA Biosynthesis
3.5. ICA Production from Hydrophobic Substrate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Citric Acid Market Size, Share & Trends Analysis Report by form (Liquid, Powder), by Application (Pharmaceuticals, F&B), by Region, Competitive Landscape, and Segment Forecasts, 2018–2025; Grand View Research Inc.: San Francisco, CA, USA, 2018; Report ID: GVR-2-68038-083-5.
- Papagianni, M. Advances in citric acid fermentation by Aspergillus niger: Biochemical aspects, membrane transport and modeling. Biotechnol. Adv. 2007, 25, 244–263. [Google Scholar] [CrossRef]
- Max, B.; Salgado, J.M.; Rodríguez, N.; Cortés, S.; Converti, A.; Domínguez, J.M. Biotechnological production of citric acid. Braz. J. Microbiol. 2010, 41, 862–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenberghe, L.P.; Soccol, C.R.; Pandey, A.; Lebeault, J.M. Microbial production of citric acid. Braz. Arch. Biol. Technol. 1999, 42, 263–276. [Google Scholar] [CrossRef]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.L.; Molina-Jouve, C.; Nicaud, J.M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Förster, A.; Aurich, A.; Mauersberger, S.; Barth, G. Citric acid production from sucrose using a recombinant strain of the yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2007, 75, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Rymowicz, W.; Rywińska, A.; Żarowska, B.; Juszczyk, P. Citric acid production from raw glycerol by acetate mutants of Yarrowia lipolytica. Chem. Pap. 2006, 60, 391–394. [Google Scholar] [CrossRef]
- Rywińska, A.; Rymowicz, W.; Żarowska, B.; Wojtatowicz, M. Biosynthesis of citric acid from glycerol by acetate mutants of Yarrowia lipolytica in fed-batch fermentation. Food Technol. Biotechnol. 2009, 47, 1–6. [Google Scholar]
- Rzechonek, D.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Aseptic production of citric and isocitric acid from crude glycerol by genetically modified Yarrowia lipolytica. Bioresour. Technol. 2019, 271, 340–344. [Google Scholar] [CrossRef]
- Yuzbashevaa, E.Y.; Agrimi, G.; Yuzbashev, T.V.; Scarcia, P.; Vinogradova, E.B.; Palmieri, L.; Shutov, A.V.; Kosikhina, J.M.; Palmieri, F.; Sineoky, S.P. The mitochondrial citrate carrier in Yarrowia lipolytica: Its identification, characterization and functional significance for the production of citric acid. Metab. Eng. 2019, 54, 264–274. [Google Scholar] [CrossRef]
- Nicaud, J.M. Yarrowia lipolytica. Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Yarrowia lipolytica yeast biomass as a novel foodpursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, 5594. [Google Scholar]
- Kamzolova, S.V.; Shamin, R.V.; Stepanova, N.N.; Morgunov, G.I.; Lunina, J.N.; Allayarov, R.K.; Samoilenko, V.A.; Morgunov, I.G. Fermentation Conditions and Media Optimization for Isocitric Acid Production from Ethanol by Yarrowia lipolytica. BioMed Res. Int. 2018, 2543210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rymowicz, W.; Fatykhova, A.R.; Kamzolova, S.V.; Rywińska, A.; Morgunov, I.G. Citric acid production from glycerol-containing waste of biodiesel industry by Yarrowia lipolytica in batch, repeated batch, and cell recycle regimes. Appl. Microbiol. Biotechnol. 2010, 87, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Anastassiadis, S.; Morgunov, I.G.; Kamzolova, S.V.; Finogenova, T.V. Citric acid production patent review. Recent Pat. Biotechnol. 2008, 2, 107–123. [Google Scholar] [CrossRef]
- Fickers, P.; Benetti, P.H.; Waché, Y.; Marty, A.; Mauersberger, S.; Smit, M.S.; Nicaud, J.M. Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential applications. FEMS Yeast Res. 2005, 5, 527–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ježek, J.; Suhaj, M. Application of capillary isotachophoresis for fruit juice authentication. J. Chromatogr. A 2001, 916, 185–189. [Google Scholar] [CrossRef]
- Ellis, G.; Goldberg, D.M. An improved manual and semi-automatic assay for NADP-dependent isocitrate dehydrogenase activity, with a description of some kinetic properties of human liver and serum enzyme. Clin. Biochem. 1971, 4, 175–185. [Google Scholar] [CrossRef]
- Aurich, A.; Specht, R.; Müller, R.A.; Stottmeister, U.; Yovkova, V.; Otto, C.; Holz, M.; Barth, G.; Heretsch, P.; Thomas, F.A.; et al. Microbiologically produced carboxylic acids used as building blocks in organic synthesis. In Reprogramming Microbial Metabolic Pathways; Springer: Dordrecht, The Netherlands, 2012; pp. 391–423. [Google Scholar] [CrossRef]
- Heretsch, P.; Thomas, F.; Aurich, A.; Krautscheid, H.; Sicker, D.; Giannis, A. Syntheses with a Chiral Building Block from the Citric Acid Cycle:(2R, 3S)-Isocitric Acid by Fermentation of Sunflower Oil. Angew. Chem. Int. Ed. 2008, 47, 1958–1960. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Dedyukhina, E.G.; Samoilenko, V.A.; Lunina, J.N.; Puntus, I.F.; Allayarov, R.L.; Chiglintseva, M.N.; Mironov, A.A.; Morgunov, I.G. Isocitric acid production from rapeseed oil by Yarrowia lipolytica yeast. Appl. Microbiol. Biotechnol. 2013, 97, 9133–9144. [Google Scholar] [CrossRef]
- Rånby, M.; Gojceta, T.; Gustafsson, K.; Hansson, K.M.; Lindahl, T.L. Isocitrate as calcium ion activity buffer in coagulation assays. Clin. Chem. 1999, 45, 1176–1180. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Karpukhina, O.V.; Bokieva, S.B.; Inozemtsev, A.N. Biosynthesis of isocitric acid in repeated-batch culture and testing of its stress-protective activity. Appl. Microb. Cell Physiol. 2019, 103, 3549–3558. [Google Scholar] [CrossRef] [PubMed]
- Kondrashova, M.N.; Zakharchenko, M.V.; Khunderiakova, N.V.; Fedotcheva, N.; Litvinova, E.; Romanova, O.; Gulayev, A. States of succinate dehydrogenase in the organism: Dormant vs. hyperactive (pushed out of equilibrium). Biophysics 2013, 58, 86–94. [Google Scholar] [CrossRef]
- Finogenova, T.V.; Morgunov, I.G.; Kamzolova, S.V.; Chernyavskaya, O.G. Organic acid production by the yeast Yarrowia lipolytica: A review of prospects. Appl. Biochem. Microbiol. 2005, 41, 418–425. [Google Scholar] [CrossRef]
- Karklin, R.; Peltzmane, I.; Raminya, L.; Korde, G. Overproduction of isocitric acid by wild strain of Candida lipolytica. In Metabolism of n-Alkanes and Oversynthesis of Products by Microorganisms. Scientific Investigations; USSR Academy of Sciences, Institute of Biochemistry and Physiology of Microorganisms: Pushchino, Russia, 1991; pp. 143–146. [Google Scholar]
- Kinichiro, S.; Teruhiko, B.U.S. Method of Producing Allo-isocitric Acid by Fermentation. U.S. Patent 2,949,404, 16 October 1960. [Google Scholar]
- Oogaki, M.; Inoue, M.; Kaimaktchiev, A.C.; Nakahara, T.; Tabuchi, T. Production of isocitric acid from glucose by Candida ravautii. Agric. Biol. Chem. 1984, 48, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Nakahara, T.; Kaimaktchiev, A.C.; Oogaki-Chino, M.; Uchida, Y.; Tabuchi, T. Isocitric acid production from n-alkanes by Candida catenulata. Agric. Biol. Chem. 1987, 51, 2111–2116. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Lunina, J.N.; Morgunov, I.G. Biochemistry of citric acid production from rapeseed oil by Yarrowia lipolytica yeast. J. Am. Oil Chem. Soc. 2011, 88, 1965–1976. [Google Scholar] [CrossRef]
- Kalia, V.C.; Prakash, J.; Koul, S. Biorefinery for glycerol rich biodiesel industry waste. Indian J. Microbiol. 2016, 56, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.C.; He, B.B. Characterization of crude glycerol from biodiesel production from multiple feedstocks. Appl. Eng. Agric. 2006, 22, 261–265. [Google Scholar] [CrossRef]
- Dobrowolski, A.; Mituła, P.; Rymowicz, W.; Mirończuk, A.M. Efficient conversion of crude glycerol from various industrial wastes into single cell oil by yeast Yarrowia lipolytica. Bioresour. Technol. 2016, 207, 237–243. [Google Scholar] [CrossRef]
- Gao, C.; Yang, X.; Wang, H.; Perez Rivero, C.; Li, C.; Cui, Z.; Qi, Q.; Ki Lin, C.S. Robust succinic acid production from crude glycerol using engineered Yarrowia lipolytica. Biotechnol. Biofuels 2016, 9, 179. [Google Scholar] [CrossRef]
- Zhang, S.; Jagtap, S.S.; Deewan, A.; Rao, C.V. pH selectively regulates citric acid and lipid production in Yarrowia lipolytica W29 during nitrogen-limited growth on glucose. J. Biotechnol. 2019, 290, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Wojtatowicz, M.; Rymowicz, W.; Kautola, H. Comparison of different strains of the yeast Yarrowia lipolytica for citric acid production from glucose hydrol. Appl. Biochem. Biotechnol. 1991, 31, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Holz, M.; Förster, A.; Mauersberger, S.; Barth, G. Aconitase overexpression changes the product ratio of citric acid production by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2009, 81, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual (3-Volume Set); Cold Spring Harb Lab Press: Cold Spring Harbor, NY, USA, 2001; ISBN 978-087969577-4. [Google Scholar]
- Querol, A.; Barrio, E.; Huerta, T.; Ramón, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: http://gryc.inra.fr (accessed on 4 November 2019).
- Fickers, P.; Le Dall, M.T.; Gaillardin, C.; Thonart, P.; Nicaud, J.M. New disruption cassettes for rapid gene disruption and marker rescue in the yeast Yarrowia lipolytica. J. Microbiol. Methods 2003, 55, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.C.; Beckerich, J.M.; Gaillardin, C. One-step transformation of the dimorphic yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 1997, 48, 232–235. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Available online: https://www.yeastgenome.org (accessed on 1 December 2019).
- Available online: https://www.ncbi.nlm.nih.gov (accessed on 1 December 2019).
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; López, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Graybill, E.R.; Rouhier, M.F.; Kirby, C.E.; Hawes, J.W. Functional comparison of citrate synthase isoforms from S. cerevisiae. Arch. Biochem. Biophys. 2007, 465, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Wojtatowicz, M.; Marchin, G.L.; Erickson, L.E. Attempts to improve strain A-101 of Yarrowia lipolytica for citric acid production from n-paraffins. Process Biochem. 1993, 28, 453–460. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Characterization of Yarrowia lipolytica and related species for citric acid production from glycerol. Enzym. Microb. Technol. 2007, 41, 292–295. [Google Scholar] [CrossRef]
- Ratledge, C.; Wynn, J.P. The biochemistry and molecular biology of lipid accumulation in oleaginous microorganisms. Adv. Appl. Microbiol. 2002, 51, 1–52. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, G.J.; Panneman, H.; Xu, D.B.; Visser, J. Properties of Aspergillus niger citrate synthase and effects of cit A overexpression on citric acid production. Fems Microbiol. Lett. 2000, 184, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanikolaou, S.; Beopoulos, A.; Koletti, A.; Thevenieau, F.; Koutinas, A.A.; Nicaud, J.M.; Aggelis, G. Importance of the methyl-citrate cycle on glycerol metabolism in the yeast Yarrowia lipolytica. J. Biotechnol. 2013, 168, 303–314. [Google Scholar] [CrossRef]
- Domínguez, A.; Deive, F.J.; Angeles Sanromán, M.; Longo, M.A. Biodegradation and utilization of waste cooking oil by Yarrowia lipolytica CECT 1240. Eur. J. Lipid Sci. Technol. 2010, 112, 1200–1208. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lv, J.; Xu, J.; Zhang, T.; Deng, Y.; He, J. Citric acid production in Yarrowia lipolytica SWJ-1b yeast when grown on waste cooking oil. Appl. Biochem. Biotechnol. 2015, 175, 2347–2356. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Reference |
|---|---|---|
| A101 | Wild type | [36] |
| A101.1.31 | Acetate-negative UV mutant of A101 | [7] |
| A101.1.31 U- | A101.1.31 ura3-302 | This study |
| Δcit1 | A101.1.31 cit1::URA3 | |
| Δcit2 | A101.1.31. cit2::URA3 | |
| CIT1 | A101.1.31 CIT1-URA3 | |
| CIT2 | A101.1.31 CIT2-URA3 |
| X | ∑CA | CA | ICA | CA/ICA | Y∑CA/X | YCA/X | YICA/X | Y∑CA/S | Q∑CA | q∑CA | YCA/S | QCA | qCA | YICA/S | QICA | qICA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g dm−3 | g dm−3 | g dm−3 | g dm−3 | g g−1 | g g−1 | g g−1 | g g−1 | g dm−3 h−1 | g g−1 h−1 | g g−1 | g dm−3 h−1 | g g−1 h−1 | g g−1 | g dm−3 h−1 | g g−1 h−1 | ||
| A101.1.31 | 17.05 | 75.41 | 72.96 | 2.45 | 29.78 | 4.42 | 4.28 | 0.14 | 0.56 | 0.52 | 0.061 | 0.54 | 0.51 | 0.059 | 0.02 | 0.02 | 0.002 |
| CIT1 | 18.35 | 83.30 | 60.33 | 22.97 | 2.63 | 4.54 | 3.29 | 1.25 | 0.54 | 0.58 | 0.063 | 0.39 | 0.42 | 0.046 | 0.15 | 0.16 | 0.017 |
| CIT2 | 18.80 | 92.75 | 72.06 | 16.69 | 4.56 | 4.93 | 3.83 | 0.89 | 0.57 | 0.64 | 0.068 | 0.43 | 0.53 | 0.053 | 0.10 | 0.11 | 0.012 |
| Time | X | ∑CA | CA | ICA | CA:ICA | Y∑CA/X | YCA/X | YICA/X | Y∑CA/S | Q∑CAs | q∑CA | YCA/S | QCA | qCA | YICA/S | QICA | qICA | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| h | g dm−3 | g dm−3 | g dm−3 | g dm−3 | g g−1 | g g−1 | g g−1 | g g−1 | g dm−3 h−1 | g g−1 h−1 | g g−1 | g dm−3 h−1 | g g−1 h−1 | g g−1 | g dm−3 h−1 | g g−1 h−1 | ||
| A101 | 91 | 55 | 130.7 | 105.2 | 25.5 | 4.12 | 2.38 | 1.91 | 0.46 | 0.65 | 0.72 | 0.026 | 0.53 | 0.58 | 0.021 | 0.13 | 0.14 | 0.005 |
| CIT1 | 94 | 26.9 | 133.2 | 67.8 | 65.4 | 1.04 | 4.95 | 2.52 | 2.43 | 0.67 | 0.71 | 0.052 | 0.34 | 0.36 | 0.027 | 0.33 | 0.35 | 0.026 |
| CIT2 | 94 | 30.9 | 109.9 | 53.6 | 56.2 | 0.95 | 3.56 | 1.73 | 1.82 | 0.55 | 0.58 | 0.038 | 0.27 | 0.28 | 0.018 | 0.28 | 0.30 | 0.019 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hapeta, P.; Rakicka-Pustułka, M.; Juszczyk, P.; Robak, M.; Rymowicz, W.; Lazar, Z. Overexpression of Citrate Synthase Increases Isocitric Acid Biosynthesis in the Yeast Yarrowia lipolytica. Sustainability 2020, 12, 7364. https://doi.org/10.3390/su12187364
Hapeta P, Rakicka-Pustułka M, Juszczyk P, Robak M, Rymowicz W, Lazar Z. Overexpression of Citrate Synthase Increases Isocitric Acid Biosynthesis in the Yeast Yarrowia lipolytica. Sustainability. 2020; 12(18):7364. https://doi.org/10.3390/su12187364
Chicago/Turabian StyleHapeta, Piotr, Magdalena Rakicka-Pustułka, Piotr Juszczyk, Małgorzata Robak, Waldemar Rymowicz, and Zbigniew Lazar. 2020. "Overexpression of Citrate Synthase Increases Isocitric Acid Biosynthesis in the Yeast Yarrowia lipolytica" Sustainability 12, no. 18: 7364. https://doi.org/10.3390/su12187364