Silicon Alleviates Copper Toxicity in Flax Plants by Up-Regulating Antioxidant Defense and Secondary Metabolites and Decreasing Oxidative Damage



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Procedures
- The first group was divided into two subgroups:
- Plants grown in soil irrigated with 400 mL/pot tap water to serve as the control.
- Plants grown in soil irrigated with 400 mL/pot of 1.5 mM potassium silicate (K2SiO3) only three times 25, 30, and 35 days from sowing.
- The second group was divided into two subgroups:
- Plants grown in soil irrigated with CuSO4 concentrations (100 μM).
- Plants grown in soil irrigated with CuSO4 concentrations (100 μM) + and irrigated with 400 mL/pot of potassium silicate (K2SiO3).
- The third group was divided into two subgroups:
- Plants grown in soil irrigated with CuSO4 concentrations (200 μM).
- Plants grown in soil irrigated with CuSO4 concentrations (200 μM) + and irrigated with 400 mL/pot of 1.5 mM potassium silicate (K2SiO3).
2.3. Morphological Characteristics and Plant Sample Preparation
2.4. Biochemical Studies
2.4.1. Estimation of Photosynthetic Pigments
2.4.2. Estimation of Total Nucleic Acids (DNA and RNA)
2.4.3. Determination of Total Phenols
2.4.4. Determination of Total Flavonoids
2.4.5. Determination of Tocopherol
2.4.6. Determination of the Total Soluble Protein
2.4.7. Extraction of Enzymes and ROS Content
2.4.8. Determination of Antioxidant Enzymes
2.4.9. Determination of ROS Content
2.4.10. Determination of Minerals
2.4.11. Determination of Fatty Acids by Gas–liquid chromatography (GLC)
2.5. Statistical Analysis
3. Results
3.1. Preliminary Study
3.2. Effect of Silicon and Copper on Morphological Characteristics
3.3. Effect of Silicon and Copper on Yield Attributes
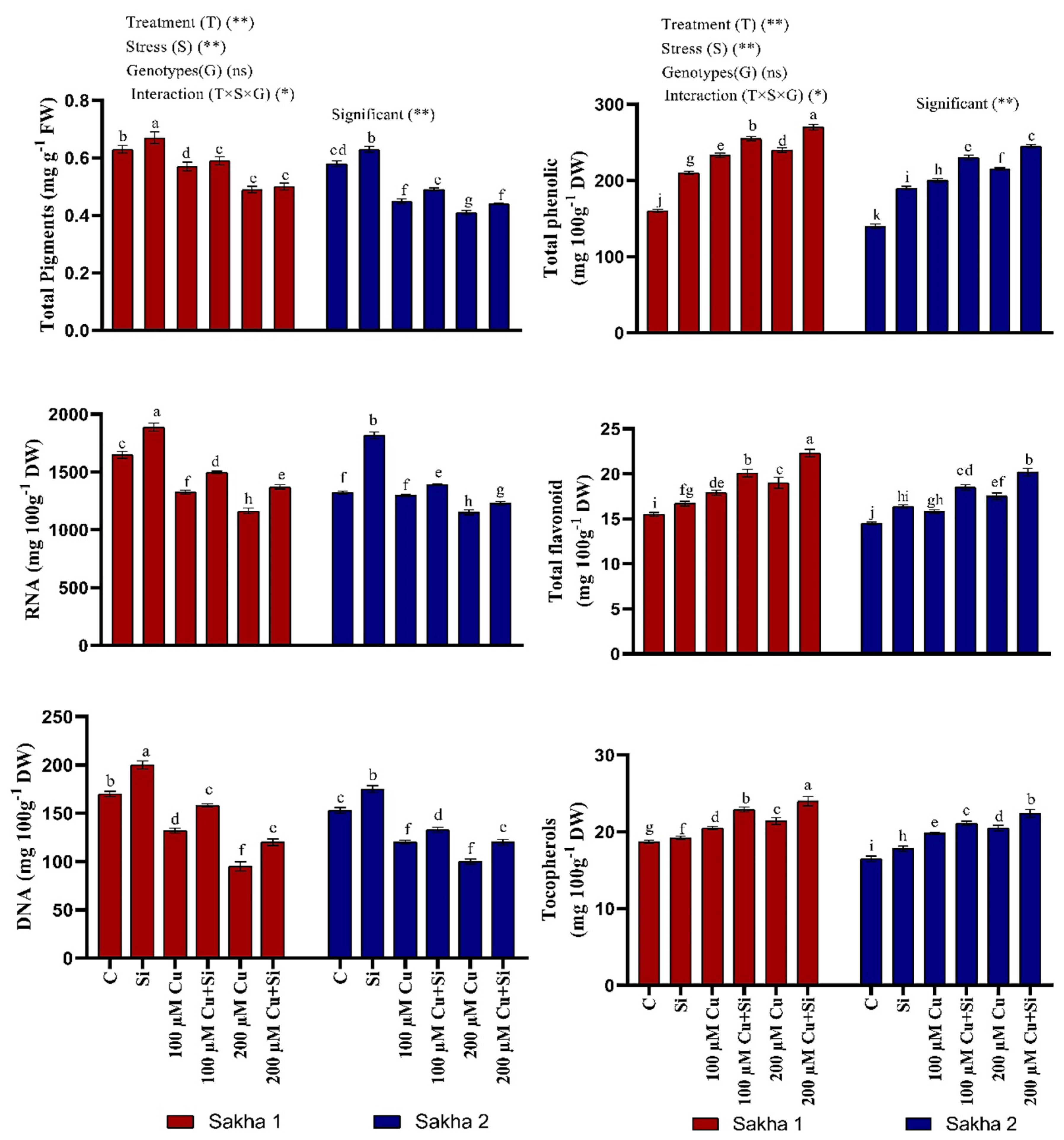
3.4. Effect of Silicon and Copper on Total Chlorophyll, Nucleic Acid, and Secondary Metabolites
3.5. Effect of Silicon and Copper on Antioxidant Enzymes
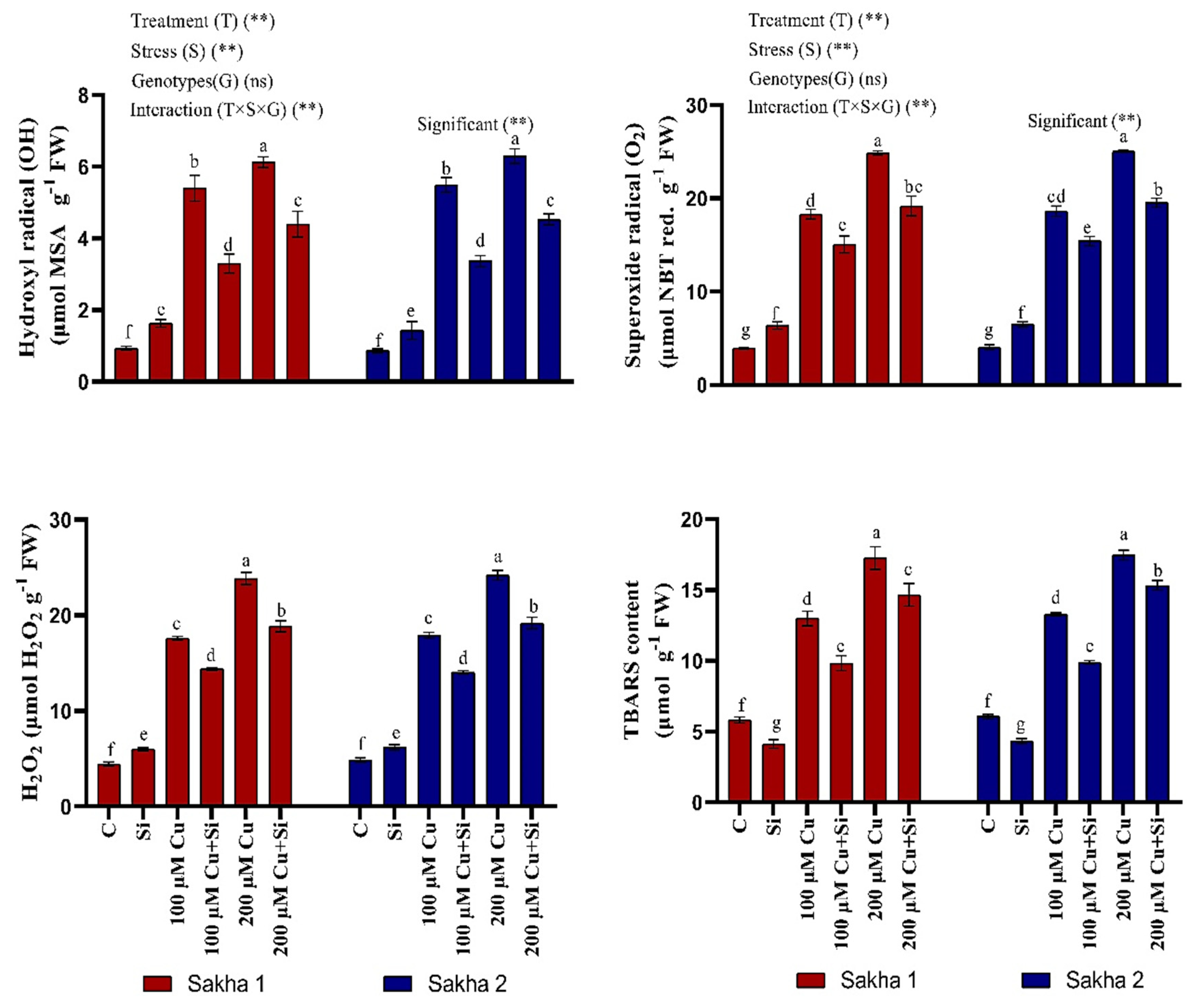
3.6. Effect of Silicon and Copper on Oxidative Damage
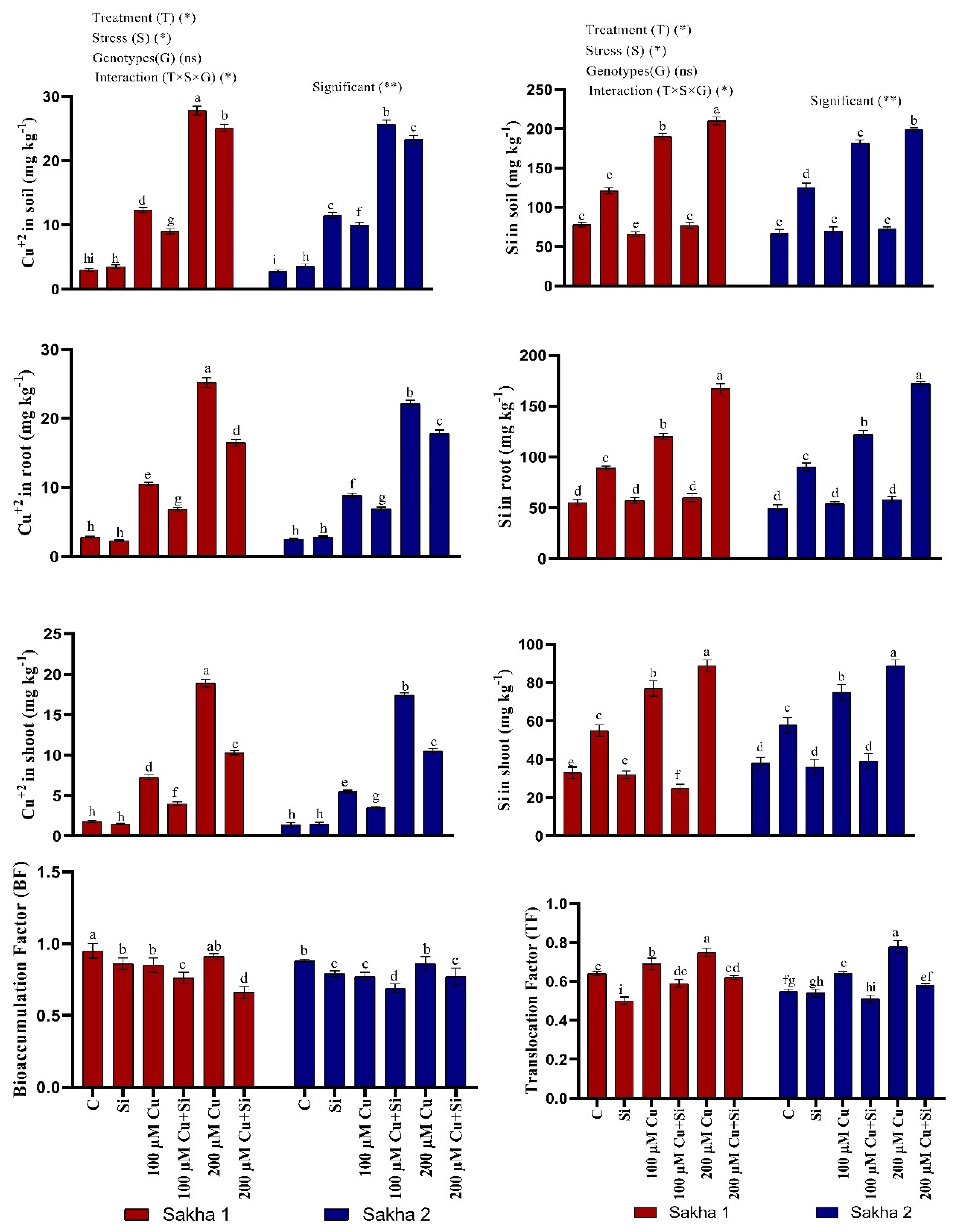
3.7. Effect of Silicon and Copper on Copper Contents
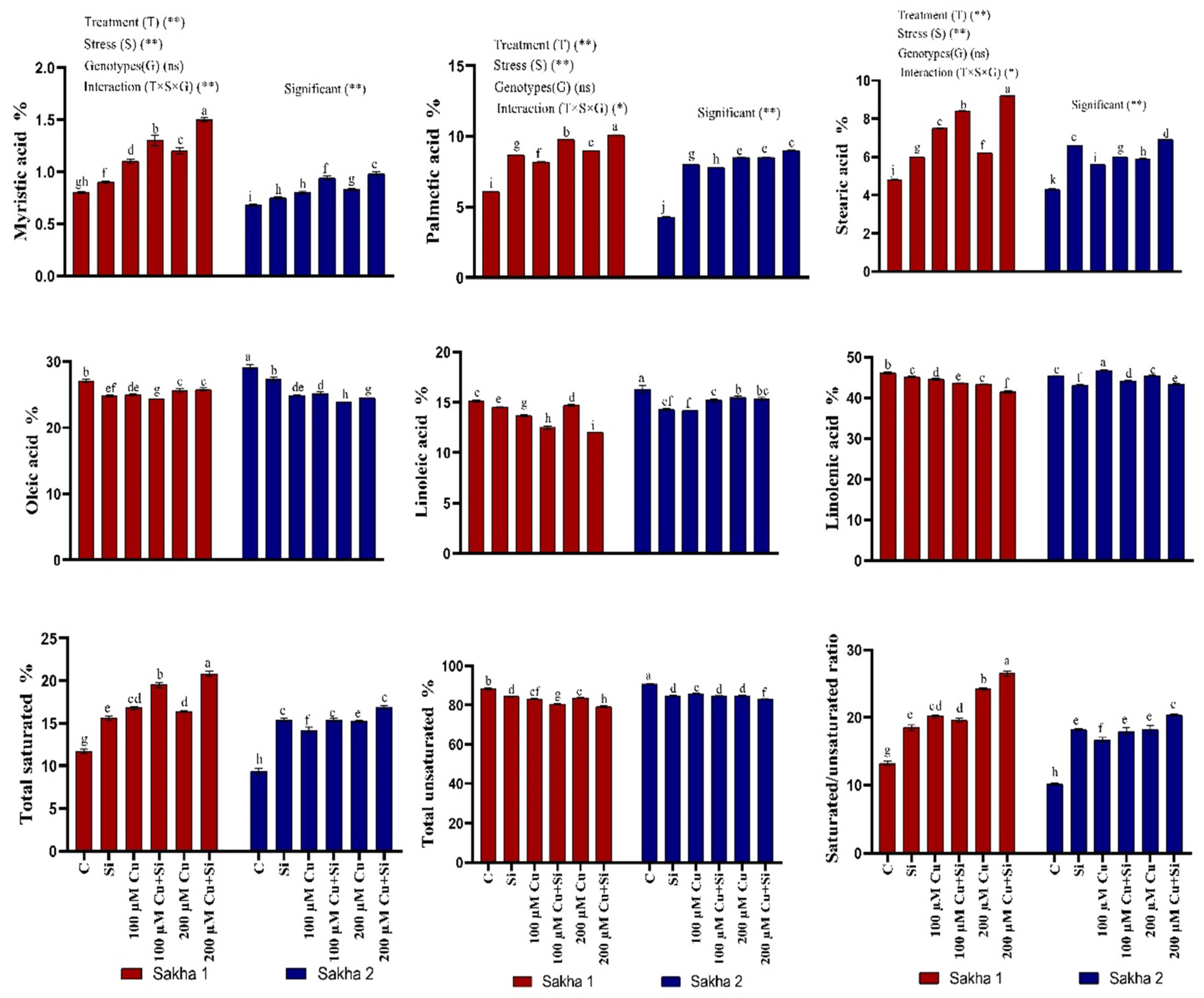
3.8. Effect of Silicon and Copper on Fatty Acid Composition of Harvested Seeds
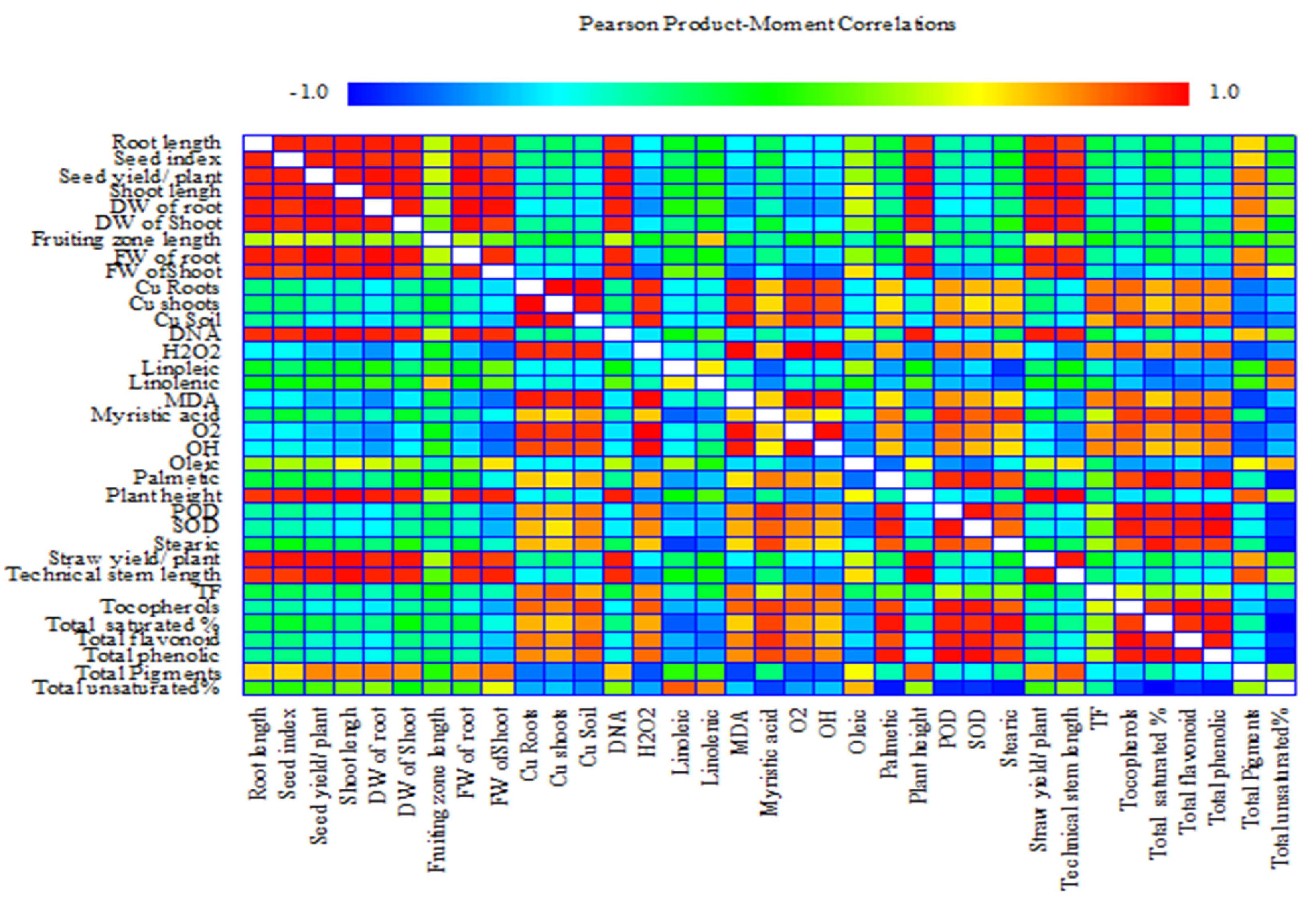
3.9. Correlation
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rehman, M.; Maqbool, Z.; Peng, D.; Liu, L. Morpho-physiological traits, antioxidant capacity and phytoextraction of copper by ramie (Boehmerianivea L.) grown as fodder in copper-contaminated soil. Environ. Sci. Pollut. Res. 2019, 26, 5851–5861. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Shafi, M.; Ma, J.; Zhong, B.; Guo, J.; Hu, X.; Xu, W.; Yang, Y.; Ruan, Z.; Wang, Y.; et al. Effect of amendments on contaminated soil of multiple heavy metals and accumulation of heavy metals in plants. Environ. Sci. Pollut. Res. 2018, 25, 28695–28704. [Google Scholar] [CrossRef] [PubMed]
- Mokhtar, S.; El Agroudy, N.; Shafiq, F.A.; Abdel Fatah, H.Y. The Effects of the Environmental Pollution in Egypt. Inter. J. Environ. 2015, 4, 21–26. [Google Scholar]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Ziaur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci Pollut. Res. 2015, 22, 8148–8162. [Google Scholar] [CrossRef]
- Burkhead, J.L.; Reynolds, K.A.G.; Abdel-Ghany, S.E.; Cohu, C.M.; Pilon, M. Copper homeostasis. New Phytol. 2009, 182, 799–816. [Google Scholar] [CrossRef]
- Saleem, M.H.; Kamran, M.; Zhou, Y.; Parveen, A.; Rehman, M.; Ahmar, S.; Malik, Z.; Mustafa, A.; Anjum, R.M.A.; Wang, B.; et al. Appraising growth, oxidative stress and copper phytoextraction potential of flax (Linum usitatissimum L.) grown in soil differentially spiked with copper. J. Environ. Manag. 2020, 257, 109994. [Google Scholar] [CrossRef]
- Cuypers, A.; Smeets, K.; Ruytinx, J.; Opdenakker, K.; Keunen, E.; Remans, T.; Horemans, N.; Vanhoudt, N.; Van Sanden, S.; Van Belleghem, F.; et al. The cellular redox state as a modulator in cadmium and copper responses in Arabidopsis thaliana seedlings. J. Plant. Physiol. 2011, 168, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Bouazizi, H.; Jouili, H.; Geitmann, A.; Ferjani, E.E. Copper toxicity in expanding leaves of Phaseolus vulgaris L.; antioxidant enzyme response and nutrient element uptake. Ecotoxicol. Environ. Saf. 2010, 73, 1304–1308. [Google Scholar] [CrossRef]
- Murakami, M.; Ae, N. Potential for phytoextraction of copper, lead, and zinc by rice (Oryza sativa L.), soybean (Glycine max [L.] Merr.), and maize (Zea mays L.). J. Hazard. Mater. 2009, 162, 1185–1192. [Google Scholar] [CrossRef]
- Bertini, L.; Focaracci, F.; Proietti, S.; Papetti, P.; Caruso, C. Physiological response of Posidonia oceanica to heavy metal pollution along the Tyrrhenian coast. Funct. Plant. Biol. 2019, 46, 933–941. [Google Scholar] [CrossRef]
- Feigl, G.; Kumar, D.; Lehotai, N.; Tugyi, N.; Molnár, Á.; Ördög, A.; Szepesi, Á.; Gémes, K.; Laskay, G.; Erdei, L.; et al. Physiological and morphological responses of the root system of Indian mustard (Brassica juncea L. Czern.) and rapeseed (Brassica napus L.) to copper stress. Ecotoxicol. Environ. Saf. 2013, 94, 179–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Beltagi, H.S.; Mohamed, H.I. Alleviation of cadmium toxicity in Pisum sativum L. seedlings by calcium chloride. Not. Bot. Horti Agrobot. 2013, 411, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Akladious, S.A.; Mohamed, H.I. Ameliorative effects of calcium nitrate and humic acid on the growth, yield component and biochemical attribute of pepper (Capsicum annuum) plants grown under salt stress. Sci. Hortic. 2018, 236, 244–250. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Akmar, A.S.N.; Rafii, M.Y.; Azizi, P.; Tengoua, F.F.; Azwa, J.N.M.; Shabanimofrad, M. Importance of silicon and mechanisms of biosilica formation in plants. BioMed Res. Int. 2015. [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 7, 1072. [Google Scholar] [CrossRef] [Green Version]
- Cuong, T.X.; Ullah, H.; Datta, A.; Hanh, T.C. Effects of silicon-based fertilizer on growth, yield and nutrient uptake of rice in tropical zone of Vietnam. Rice Sci. 2017, 24, 283–290. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes, suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon-mediated tolerance to metal toxicity. In Silicon in Agriculture, from Theory to Practice; Springer: Dordrecht, The Netherlands, 2015; pp. 83–122. [Google Scholar]
- Shedeed, S.I.; Ahmed, H.K.; Medhat, K.A. Silicon-enhanced reduction of some heavy elements accumulation in Cowpea Plants (Vigna unguiculata L.). J. Innov. Pharm. Biol. Sci. 2018, 5, 1–10. [Google Scholar]
- Khandekar, S.; Leisner, S. Soluble silicon modulates expression of Arabidopsis thaliana genes involved in copper stress. J. Plant Physiol. 2011, 168, 699–705. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012; pp. 178–189. [Google Scholar]
- Tarabih, M.E.; El-Eryan, E.E.; El-Metwally, M.A. Physiological and pathological impacts of potassium silicate on storability of Anna apple. Am. J. Plant Physiol. 2014, 9, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Saleem, M.H.; Fahad, S.; Khan, S.U.; Din, M.; Ullah, A.; EL Sabagh, A.; Hossain, A.; Lanes, A.; Liu, L. Copper-induced oxidative stress, initiation of antioxidants and phytoremediation potential of flax (Linum usitatissimum L.) seedlings grown under the mixing of two different soils of China. Environ. Sci. Pollut. Res. 2020, 27, 5211–5221. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.A.; Mohamed, H.I.; Mansour, M.T.M.; Omar, M.R. Suppression of powdery mildew on flax by foliar application of essential oils. J. Phytopathol. 2013, 161, 376–381. [Google Scholar] [CrossRef]
- Rashwan, E.; Mousa, A.; EL-Sabagh, A.; Barutçula, C. Yield and quality traits of some flax cultivars as influenced by different irrigation intervals. J. Agric. Sci. 2016, 8, 226–240. [Google Scholar] [CrossRef] [Green Version]
- Aly, A.A.; Mansour, M.T.M.; Mohamed, H.I.; Abd-Elsalam, K.A. Examination of correlations between several biochemical components and powdery mildew resistance of flax cultivars. Plant Pathol. J. 2012, 28, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Arnon, I.D. Copper enzymes in isolated chloroplasts. Poly phenoloxidase in Beta vulgaris. Plant. Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Morse, M.L.; Carter, C.F. The synthesis of nucleic acid in cultures of Escherchia coli strains B and B/R. Bacteriology 1949, 58, 317. [Google Scholar] [CrossRef] [Green Version]
- Burton, K. Citation Classics—Study of conditions and mechanism of diphenylamine reaction for colorimetric estimation of deoxyribonucleic-acid. Curr. Contents 1977, 26, 315–323. [Google Scholar]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo-enzymology; Kalyani Publishers: New Delhi, India, 1980. [Google Scholar]
- Dihazi, A.; Jaiti, F.; Zouine, J.; Hasni, M.E.; Hadrami, I.E. Effect of salicylic acid on phenolic compounds related to date palm resistance to Fusarium oxysporum. Phytopathol. Mediterr. 2003, 42, 9–16. [Google Scholar]
- Bushra, S.; Farooq, A.; Muhammad, A. Effect of extraction solvent/technique on the antioxidant activity of selected medicinal plant extracts. Molecules 2009, 14, 2167–2180. [Google Scholar]
- Backer, H.; Frank, O.; De Angells, B.; Feingold, S. Plasma tocopherol in man at various times after ingesting free or ocetylaned tocopherol. Nutr. Rep. Int. 1980, 21, 531–536. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase, improved assays and assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase, polyphenol oxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Aebi, H.; Bergmeyer, H.U. Methods of Enzymatic Analysis. In Catalase; Verlag Chemie: Weinheim, Germany, 1983; Volume 3, pp. 273–277. [Google Scholar]
- Green, M.J.; Hill, M.A.O. Chemistry of dioxygen. Meth. Enzymol. 1984, 105, 3–22. [Google Scholar]
- Babbs, C.F.; Pham, J.A.; Coolbaugh, R.C. Lethal hydroxyl radical production in paraquat-treated plants. Plant Physiol. 1989, 90, 1267–1270. [Google Scholar] [CrossRef] [Green Version]
- Pick, E. Micro assays for superoxide and hydrogen peroxide production and nitroblue tetrazolium reduction using an enzyme immunoassay microplate reader. Meth. Enzymol. 1986, 132, 407–421. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Abdul, H.B.O.; Thomas, B.V. Translocation and bioaccumulation of trace metals in desert plants of Kuwait Governorates. Res. J. Environ. Sci. 2009, 3, 581–587. [Google Scholar]
- Malik, R.N.; Husain, S.Z.; Nazir, I. Heavy metal contamination and accumulation in soil and wild plant species from industrial area of Islamabad, Pakistan. Pak. J. Bot. 2010, 42, 123–127. [Google Scholar]
- Van der Vorm, P.D.J. Dry ashing of plant material and dissolution of the ash in HF for the colorimetric determination of silicon. Commun. Soil Sci. Plant 1987, 18, 1181–1189. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of The Association of Official Analytical Chemistis, 15th ed.; Association of Official Analytical Chemistis Arlington: Virginia, VA, USA, 1990. [Google Scholar]
- Stahl, E. Thin Layer Chromatography; Academic Press. Inc. Pulishers: New York, NY, USA; London, UK, 1965. [Google Scholar]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons Inc.: Singapore, 1984; p. 680. [Google Scholar]
- Ben Massoud, M.; Karmous, I.; El Ferjani, E.; Chaoui, A. Alleviation of copper toxicity in germinating pea seeds by IAA, GA3, Ca and citric acid. J. Plant Interact. 2018, 13, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Noreen, S.; Akhter, M.S.; Yaamin, T.; Arfan, M. The ameliorative effects of exogenously applied proline on physiological and biochemical parameters of wheat Triticum aestivum L. crop under copper stress condition. J. Plant Interact. 2018, 13, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Sheo, M.P. Heavy metal tolerance in plants, role of transcriptomics, proteomics, metabolomics and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing adverse effects of pb on maize plants by combined treatment with jasmonic, salicylic acids and proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Akladious, S.A.; Mohamed, H.I. Physiological role of exogenous nitric oxide in improving performance, yield and some biochemical aspects of sunflower plant under zinc stress. Acta Biol. Hung. 2017, 68, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.I.; Elsherbiny, E.A.; Abdelhamid, M.T. Physiological and biochemical responses of Vicia faba plants to foliar application with zinc and iron. Gesunde Pflanz. 2016, 68, 201–212. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Latif, H.H.; Hanafy, R.S. Influence of nitric oxide application on some biochemical aspects, endogenous hormones, minerals and phenolic compounds of Vicia faba plant grown under arsenic stress. Gesunde Pflanz. 2016, 68, 99–107. [Google Scholar] [CrossRef]
- De Conti, L.; Cesco, S.; Mimmo, T.; Pii, Y.; Valentinuzzi, F.; Melo, G.W.B.; Ceretta, C.A.; Trentin, E.; Marques, A.C.R.; Brunetto, G. Iron fertilization to enhance tolerance mechanisms to copper toxicity of ryegrass plants used as cover crop in vineyards. Chemosphere 2020, 243, 125298. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.H.; Qiu, H.; Tian, T.; Zhan, S.S.; Chaney, R.L.; Wang, S.Z.; Tang, Y.T.; Morel, J.L.; Qiu, R.L. Mitigation effects of silicon rich amendments on heavy metal accumulation in rice (Oryza sativa L.) planted on multi-metal contaminated acidic soil. Chemosphere 2011, 83, 1234–1240. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Hasnain, A.; Akhtar, S.; Hussain, A.; Yasin, G.; Wahid, A.; Mahmood, S. Antioxidant enzymes as bio-markers for copper tolerance in safflower Carthamus tinctorius L. Afr. J. Biotechnol. 2010, 9, 5441–5444. [Google Scholar]
- Drazkeiwice, M. Chlorophyll-occurrence, function and mechanism of action. Effects of external and internal factors. Photosynthetica 1994, 50, 321–331. [Google Scholar]
- Da Cunha, K.P.V.; Do Nascimento, C.W.A.; Da Silva, A.J. Silicon alleviates the toxicity of cadmium and zinc for maize Zea mays L. grown on a contaminated soil. J. Plant. Nutr. Soil Sci. 2008, 171, 849–853. [Google Scholar] [CrossRef]
- Fu, Y.Q.; Shen, H.; Wu, D.M.; Cai, K.Z. Silicon-mediated amelioration of Fe2+ toxicity in rice Oryza sativa L. roots. Pedosphere 2012, 22, 795–802. [Google Scholar] [CrossRef]
- Wu, J.W.; Shi, Y.; Zhu, Y.X.; Wang, Y.C.; Gong, H.J. Mechanisms of enhanced heavy metal tolerance in plants by silicon, a review. Pedosphere 2013, 23, 815–825. [Google Scholar] [CrossRef]
- Flora, C.; Khandekar, S.; Boldt, J.; Leisner, S. Silicon alleviates long-term copper toxicity and influences gene expression in Nicotiana tabacum. J. Plant Nutr. 2019, 42, 864–878. [Google Scholar] [CrossRef]
- El-Rahman, S.S.A.; Mazen, M.M.; Mohamed, H.I.; Mahmoud, N.M. Induction of defence related enzymes and phenolic compounds in lupin (Lupinus albus L.) and their effects on host resistance against Fusarium wilt. Eur. J. Plant Pathol. 2012, 134, 105–116. [Google Scholar] [CrossRef]
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of Silicon in Mitigation of Heavy Metal Stresses in Crop Plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [Green Version]
- Vega, I.; Nikolic, M.; Sofía, P.; Karina, G.; Mora, M.; Cartes, P. Silicon improves the production of high antioxidant or structural phenolic compounds in barley cultivars under aluminum stress. Agronomy 2019, 9, 388. [Google Scholar] [CrossRef] [Green Version]
- Kidd, P.S.; Llugany, M.; Poschenrieder, C.; Gunse, B.; Barcelo, J. The role of root exudates in aluminium resistance and silicon-induced amelioration of aluminium toxicity in three varieties of maize Zea mays L. J. Exp. Bot. 2001, 52, 1339–1352. [Google Scholar] [PubMed]
- Khorasaninejad, S.; Zaire, F.; Hemmati, K. Effects of silicon on some phytochemical traits of purple coneflower (Echinacea purpurea L.) under salinity. Sci. Hortic. 2020, 264, 108954. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Bashri, G.; Shweta, S.; Ahmad, P.; Singh, V.P. Efficacy of silicon against aluminum toxicity in plants: An overview. In Silicon in Plants, Advances and Future Prospects; CRC Press: Boca Raton, FL, USA, 2017; Volume 1, pp. 355–366. [Google Scholar]
- Pontigo, S.; Godoy, K.; Jiménez, H.; Gutiérrez-Moraga, A.; Mora, M.L.; Cartes, P. Silicon-mediated alleviation of aluminum toxicity by modulation of Al/Si uptake and antioxidant performance in ryegrass plants. Front. Plant Sci. 2017, 8, 642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, C.; Rizwan, M.; Davidian, J.-C.; Pokrovsky, O.; Bovet, N.; Chaurand, P.; Meunier, J.-D. Effect of silicon on wheat seedlings (Triticum turgidum L.) grown in hydroponics and exposed to 0 to 30 µM Cu. Planta 2015, 241, 847–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thounaojam, T.C.; Panda, P.; Mazumdar, P.; Kumar, D.; Sharma, G.D.; Sahoo, L.; Sanjib, P. Excess copper induced oxidative stress and response of antioxidants in rice. Plant Physiol. Biochem. 2012, 53, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Andrade, S.A.L.; Gratao, P.L.; Azevedo, R.A.; Silveira, A.P.D.; Schiavinato, M.A.; Mazzafera, P. Biochemical and physiological changes in jack bean under mycorrhizal symbiosis growing in soil with increasing Cu concentrations. Environ. Exp. Bot. 2010, 68, 198–207. [Google Scholar] [CrossRef]
- Sofy, A.R.; Dawoud, R.A.; Sofy, M.R.; Mohamed, H.I.; Hmed, A.A.; El-Dougdoug, N.K. Improving regulation of enzymatic and non-enzymatic antioxidants and stress-related gene stimulation in Cucumber mosaic cucumovirus-infected cucumber plants treated with glycine betaine, chitosan and combination. Molecules 2020, 25, 2341. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 1360–1385. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Fidalgo, F.; Azenha, M.; Silva, A.F.; de Sousa, A.; Santiago, A.; Ferraz, P.; Teixeira, J. Copper-induced stress in Solanum nigrum L. and antioxidant defense system responses. Food Energy Secur. 2013, 21, 70–80. [Google Scholar] [CrossRef]
- Hamid, Y.; Tang, L.; Sohail, M.I.; Cao, X.; Hussain, B.; Aziz, M.Z.; Usman, M.; He, Z.; Yang, X. An explanation of soil amendments to reduce cadmium phytoavailability and transfer to food chain. Sci. Total Environ. 2019, 660, 80–96. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H.; Jeong, B.R.; Rizwan, M. The Use of Silicon in Stressed Agriculture Management. Chapter 19. In Metalloids in Plants: Advances and Future Prospects; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Chaffai, R.; Elhammadi, M.A.; Seybou, T.N.; Tekitek, A.; Marzouk, B.; Ferjani, E.E. Altered fatty acid profile of polar lipids in maize seedlings in response to excess copper. J. Agron. Crop. Sci. 2007, 193, 207–217. [Google Scholar] [CrossRef]
- Baryla, A.; Laborde, C.; Montillet, J.L.; Triantaphylidès, C.; Chagvardieff, P. Evaluation of lipid peroxidation as a toxicity bioassay for plants exposed to copper. Environ. Pollut. 2000, 109, 131–135. [Google Scholar] [CrossRef]
- Elloumi, N.; Zouari, M.; Chaari, L.; Jomni, C.; Marzouk, B.; Abdallah, F.B. Effects of cadmium on lipids of almond seedlings Prunus dulcis. Bot. Stud. 2014, 55, 61. [Google Scholar] [CrossRef] [Green Version]
- Sharaf, A.; Farghal, I.I.; Sofy, M.R. Role of gibberellic acid in abolishing the detrimental effects of Cd and Pb on broad bean and lupin plants. Res. J. Agric. Biol. Sci. 2009, 5, 668–673. [Google Scholar]
- Morsy, A.A.; Salama, K.H.A.; Kamel, H.A.; Mansour, M.M.F. Effect of heavy metals on plasma membrane lipids and antioxidant enzymes of Zygophyllum species. Eurasia J. Biosci. 2012, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bosnić, D.; Bosnić, P.; Nikolić, D.; Nikolić, M.; Samardžić, J. Silicon and iron differently alleviate copper toxicity in cucumber leaves. Plants 2019, 8, 554. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotypes | Pedigree | Type | Origin |
|---|---|---|---|
| Sakha 1 | Bombay (USA) x I. 1485 (USA) | Dual | Local variety |
| Sakha 2 | I.2348 (Hungary) x I. Hera (India) | Dual | Local variety |
| pH | E.C. dSm−1 | Cations meq/L | Anion meq/L | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 7.8 | 0.4 | Na | K | Ca | Mg | Cl | SO4 | HCO3 | CO3 |
| 2.07 | 0.27 | 1.23 | 0.88 | 4.3 | 0.82 | 0.8 | 0 | ||
| Genotypes | Treatment | Radical Length (cm) | Plumule Length (cm) | Fresh Weight of Seedling (mg) | Dry Weight of Seedling (mg) |
|---|---|---|---|---|---|
| Sakha1 | C | 11.18 ± 0.45c | 10.73 ± 0.2a | 86.93 ± 0.4bc | 13.03 ± 0.5g |
| 200 µM Cu | 5.35 ± 0.23e | 6.41 ± 0.3g | 56.33 ± 0.6e | 7.3 ± 0.3h | |
| Sakha 2 | C | 12.07 ± 0.4abc | 10.6 ± 0.26ab | 88.33 ± 0.3abc | 14.13 ± 0.4def |
| 200 µM Cu | 7.11 ± 0.25d | 7.35 ± 0.14f | 66.67 ± 0.4d | 7.4 ± 0.23h | |
| Sakha 102 | C | 12.33 ± 0.44ab | 10.21 ± 0.3abcd | 86.67 ± 0.23bc | 15.33 ± 0.54bc |
| 200 µM Cu | 11.90 ± 0.31bc | 9.85 ± 0.32cde | 84.33 ± 0.53c | 14.57 ± 0.65cde | |
| Sakha 101 | C | 11.75 ± 0.23bc | 10.39 ± 0.23abc | 93.67 ± 0.4a | 13.93 ± 0.43ef |
| 200 µM Cu | 11.37 ± 0.41bc | 10.27 ± 0.1abc | 89.33 ± 0.33abc | 13.73 ± 0.44fg | |
| Giza 7 | C | 12.87 ± 0.2a | 9.58 ± 0.2de | 88.33 ± 0.23abc | 16.43 ± 0.34a |
| 200 µM Cu | 12.30 ± 0.32ab | 9.37 ± 2.4e | 84.67 ± 0.4c | 15.93 ± 0.5ab | |
| Giza 8 | C | 11.33 ± 0.41c | 10.21 ± 0.23abcd | 93 ± 0.3a | 15.27 ± 0.34bc |
| 200 µM Cu | 11.27 ± 0.15c | 9.99 ± 0.42bcde | 92.33 ± 0.21ab | 14.85 ± 0.33cd |
| Genotypes | Treatment | Shoot Length (cm) | Root Length (cm) | Shoot Fresh Weight (g) | Shoot Dry Weight (g) | Root Fresh Weight (g) | Root Dry Weight (g) |
|---|---|---|---|---|---|---|---|
| Sakha 1 | C | 55.8 ± 0.64b | 13.4 ± 0.60b | 1.50 ± 0.02b | 0.225 ± 0.034b | 0.150 ± 0.011c | 0.051 ± 0.003c |
| Si | 59.8 ± 0.76a | 14.9 ± 0.57a | 1.56 ± 0.03a | 0.268 ± 0.038a | 0.191 ± 0.014a | 0.062 ± 0.006a | |
| 100 μM Cu | 43.6 ± 0.34g | 9.3 ± 0.15g | 0.60 ± 0.01g | 0.116 ± 0.011def | 0.118 ± 0.006de | 0.031 ± 0.001f | |
| 100 μM Cu+ Si | 49.6 ± 0.35d | 10.8 ± 0.11d | 0.83 ± 0.05d | 0.176 ± 0.020c | 0.125 ± 0.004d | 0.036 ± 0.003de | |
| 200 μM Cu | 40.2 ± 0.29i | 8.7 ± 0.11h | 0.61 ± 0.02fg | 0.103 ± 0.005ef | 0.103 ± 0.003ef | 0.028 ± 0.002f | |
| 200 μM Cu+ Si | 45.9 ± 0.38e | 10.7 ± 0.12de | 0.66 ± 0.03f | 0.137 ± 0.017de | 0.122 ± 0.002d | 0.034 ± 0.003de | |
| Sakha 2 | C | 50.03 ± 0.46d | 10.2 ± 0.19ef | 1.17 ± 0.06c | 0.143 ± 0.021cd | 0.122 ± 0.003 d | 0.040 ± 0.002d |
| Si | 53.5 ± 0.51c | 12.7 ± 0.40c | 1.49 ± 0.03b | 0.239 ± 0.030ab | 0.171 ± 0.007b | 0.055 ± 0.002b | |
| 100 μM Cu | 39.6 ± 0.22i | 8.9 ± 0.08gh | 0.58 ± 0.03g | 0.103 ± 0.007ef | 0.107 ± 0.04e | 0.030 ± 0.001f | |
| 100 μM Cu+ Si | 44.4 ± 0.21f | 11.1 ± 0.34d | 0.76 ± 0.04e | 0.138 ± 0.015de | 0.120 ± 0.03de | 0.033 ± 0.002f | |
| 200 μM Cu | 36.8 ± 0.29j | 8.0 ± 0.17i | 0.56 ± 0.01g | 0.091 ± 0.008f | 0.099 ± 0.001f | 0.027 ± 0.002fg | |
| 200 μM Cu+ Si | 41.0 ± 0.19h | 10.0 ± 0.09f | 0.66 ± 0.02f | 0.118 ± 0.011def | 0.110 ± 0.004e | 0.031 ± 0.001f | |
| Significant | ** | ** | ** | ** | ** | ** | |
| Treatment (T) | ** | ** | ** | ** | ** | ** | |
| Stress (S) | ** | ** | ** | ** | ** | ** | |
| Genotypes (G) | * | ns | ns | ns | ns | ns | |
| Interaction (T*S*G) | * | ** | ** | ** | ** | ** | |
| Genotypes | Treatment | Plant Height (cm) | Technical Stem Length (cm) | Fruiting Zone Length (cm) | Straw Yield/Plant (g) | Seed Yield/Plant (g) | Seed Index (g) |
|---|---|---|---|---|---|---|---|
| Sakha 1 | C | 76.5 ± 0.95b | 62.2 ± 0.61b | 14.3 ± 0.29d | 0.24 ± 0.015b | 0.222 ± 0.021c | 4.00 ± 0.18bc |
| Si | 83.1 ± 1.26a | 66.1 ± 0.77a | 17. 0 ± 0.37a | 0.29 ± 0.017a | 0.340 ± 0.025a | 5.22 ± 0.31a | |
| 100 μM Cu | 67.1 ± 0.88e | 53.1 ± 0.50e | 14.00 ± 0.18de | 0.18 ± 0.010d | 0.166 ± 0.013ef | 3.02 ± 0.006fg | |
| 100 μM Cu + Si | 69.0 ± 0.64d | 55.0 ± 0.39d | 14.00 ± 0.26de | 0.21 ± 0.014c | 0.199 ± 0.018d | 3.75 ± 0.08cd | |
| 200 μM Cu | 63.8 ± 0.73g | 50.4 ± 0.46f | 13.4 ± 0.13g | 0.14 ± 0.012e | 0.125 ± 0.008i | 2.77 ± 0.011g | |
| 200 μM Cu + Si | 65.5 ± 0.60f | 52.7 ± 0.31e | 12.8 ± 0.15h | 0.19 ± 0.01cd | 0.156 ± 0.012fg | 3.77 ± 0.11cd | |
| Sakha 2 | C | 72.6 ± 0.51c | 59.2 ± 0.51c | 13.4 ± 0.25g | 0.20 ± 0.011cd | 0.188 ± 0.014de | 3.70 ± 0.12d |
| Si | 75.5 ± 0.43b | 62.5 ± 0.65b | 13.0 ± 0.31e | 0.24 ± 0.015b | 0.267 ± 0.024b | 4.11 ± 0.28b | |
| 100 μM Cu | 62.3 ± 0.38h | 46.3 ± 0.40h | 16.0 ± 0.11b | 0.15 ± 0.010e | 0.130 ± 0.009h | 3.22 ± 0.10ef | |
| 100 μM Cu + Si | 65.0 ± 0.29f | 50.0 ± 0.27f | 15.0 ± 0.19h | 0.18 ± 0.009d | 0.152 ± 0.011fg | 3.33 ± 0.15e | |
| 200 μM Cu | 58.1 ± 0.59g | 44.3 ± 0.35i | 13.8 ± 0.08ef | 0.10 ± 0.008f | 0.105 ± 0.006j | 2.25 ± 0.07h | |
| 200 μM Cu + Si | 60.0 ± 0.44i | 47.5 ± 0.23g | 13.5 ± 0.14fg | 0.13 ± 0.007e | 0.132 ± 0.010h | 3.00 ± 0.08fg | |
| Significant | ** | ** | ** | ** | ** | ** | |
| Treatment (T) | ** | ** | * | ** | ** | ** | |
| Stress (S) | * | ** | * | * | ** | * | |
| Genotypes (G) | * | * | ns | * | ns | ns | |
| Interaction (T*S*G) | ns | ns | ns | ns | * | ns | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Beltagi, H.S.; Sofy, M.R.; Aldaej, M.I.; Mohamed, H.I. Silicon Alleviates Copper Toxicity in Flax Plants by Up-Regulating Antioxidant Defense and Secondary Metabolites and Decreasing Oxidative Damage. Sustainability 2020, 12, 4732. https://doi.org/10.3390/su12114732
El-Beltagi HS, Sofy MR, Aldaej MI, Mohamed HI. Silicon Alleviates Copper Toxicity in Flax Plants by Up-Regulating Antioxidant Defense and Secondary Metabolites and Decreasing Oxidative Damage. Sustainability. 2020; 12(11):4732. https://doi.org/10.3390/su12114732
Chicago/Turabian StyleEl-Beltagi, Hossam S., Mahmoud R. Sofy, Mohammed I. Aldaej, and Heba I. Mohamed. 2020. "Silicon Alleviates Copper Toxicity in Flax Plants by Up-Regulating Antioxidant Defense and Secondary Metabolites and Decreasing Oxidative Damage" Sustainability 12, no. 11: 4732. https://doi.org/10.3390/su12114732