
Abnormal Brain Circuits Characterize Borderline Personality and Mediate the Relationship between Childhood Traumas and Symptoms: A mCCA+jICA and Random Forest Approach

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. The Present Study
3. Materials and Methods
3.1. Participants
3.2. Questionnaires
3.3. Preprocessing
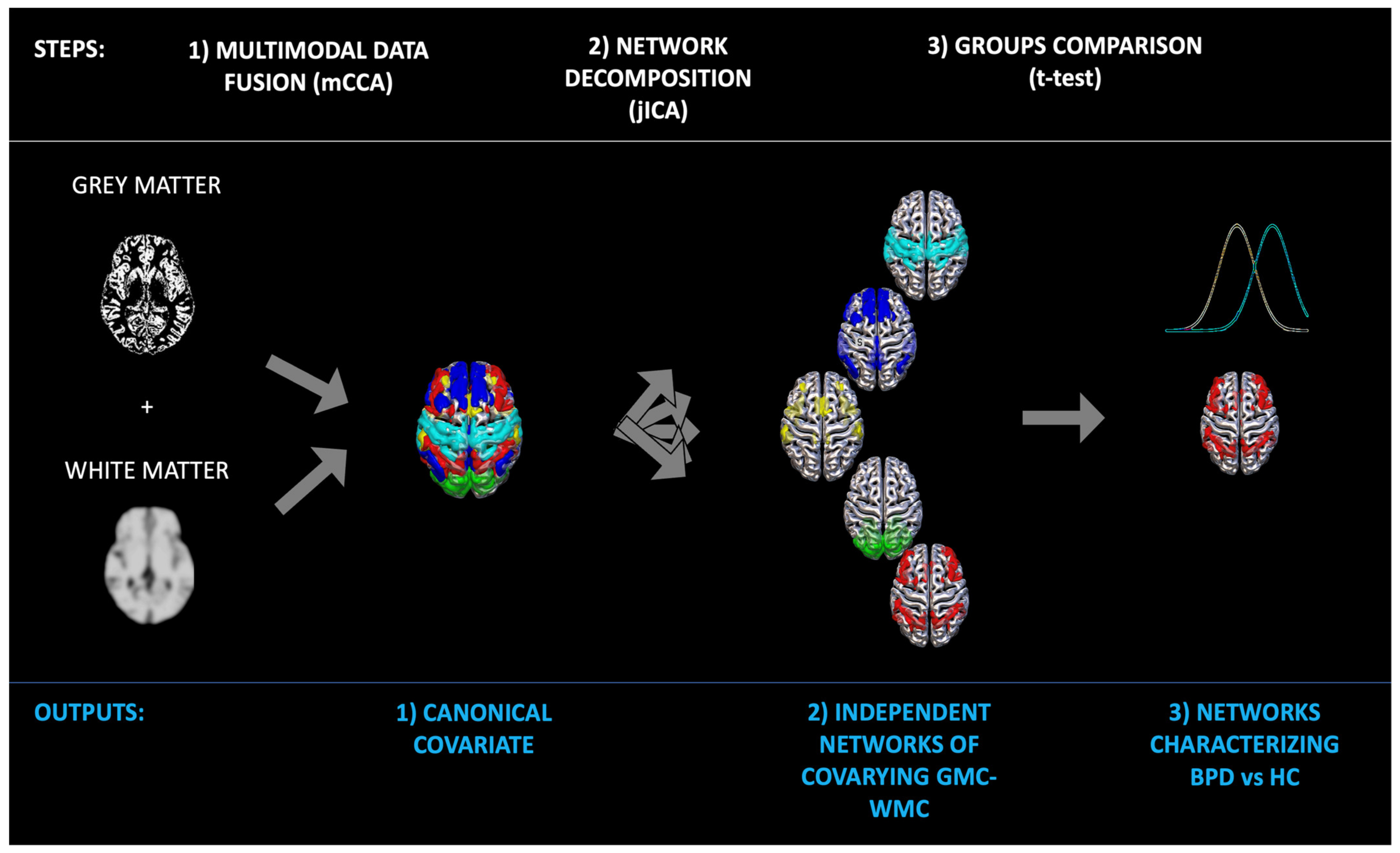
3.4. Data Fusion Unsupervised Machine Learning
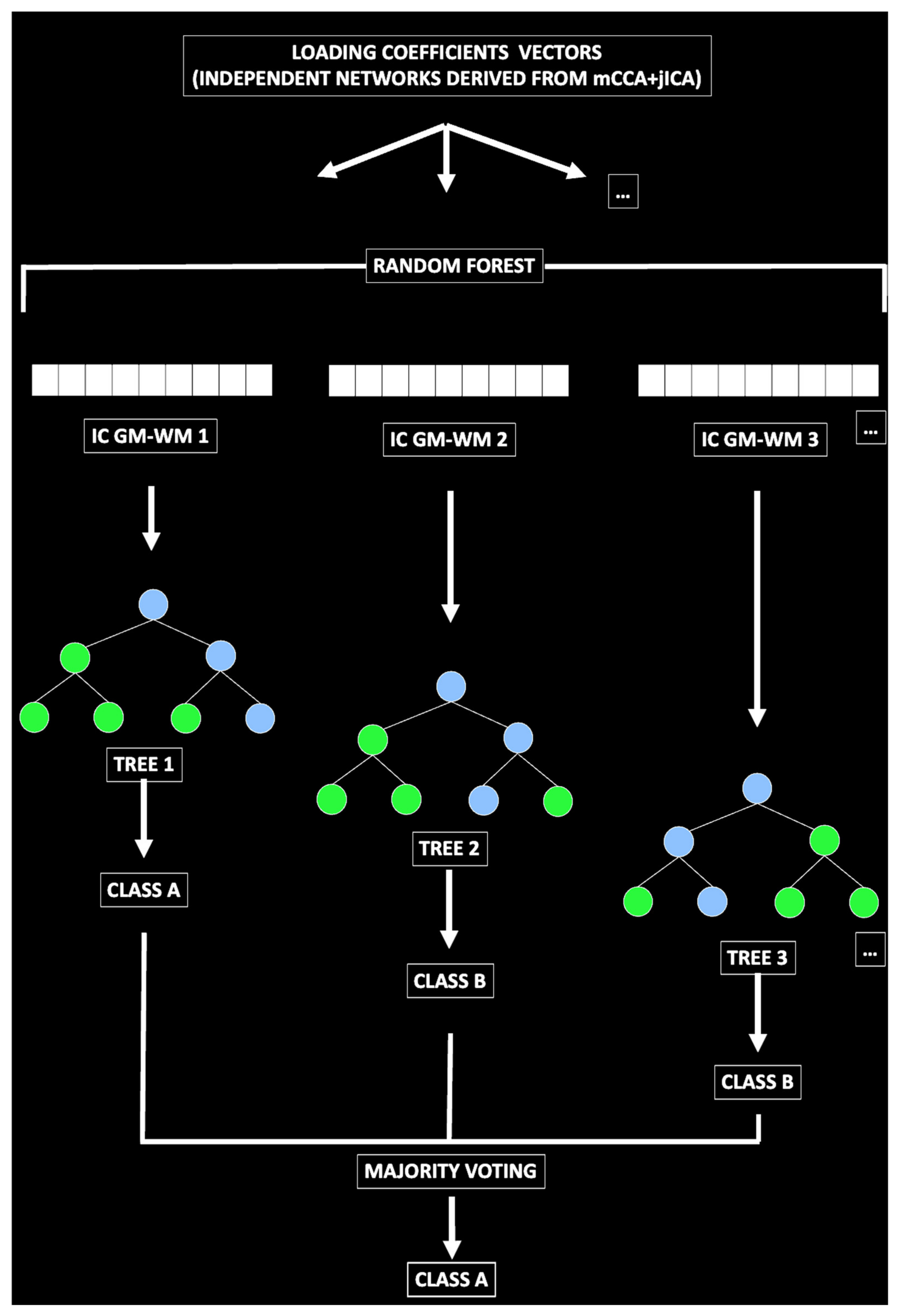
3.5. Predictive Model
4. Results
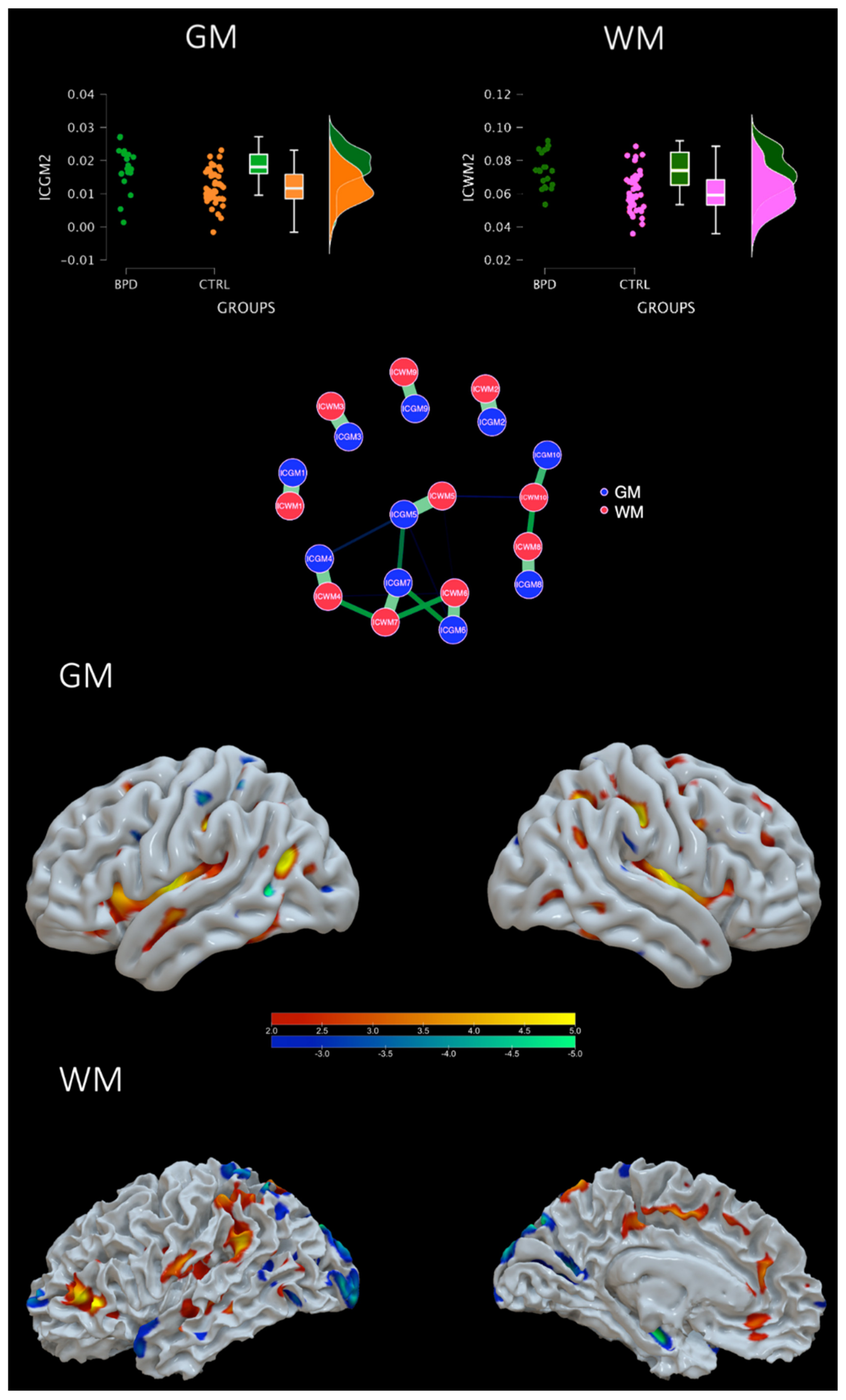
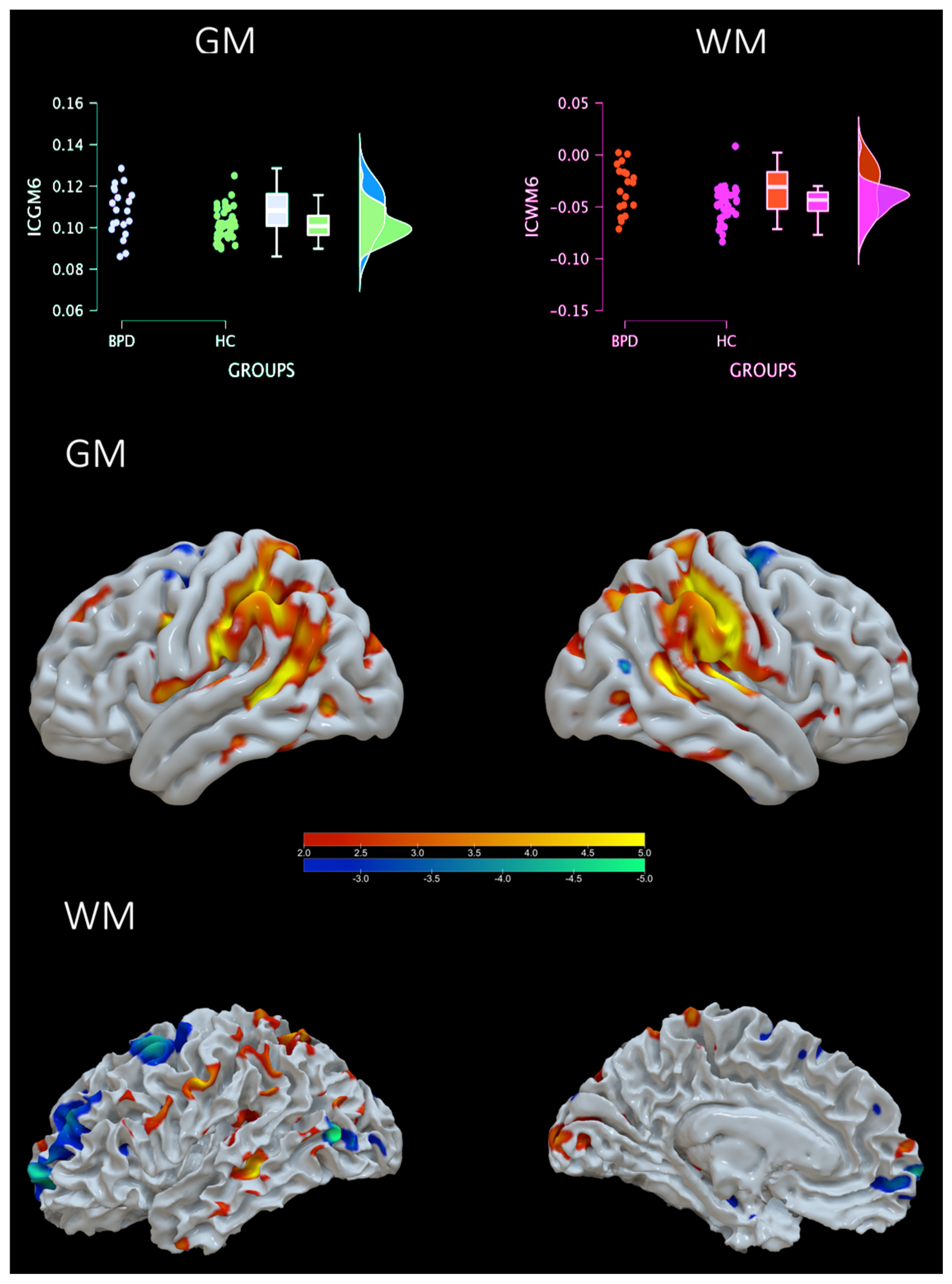
4.1. Groups Comparison
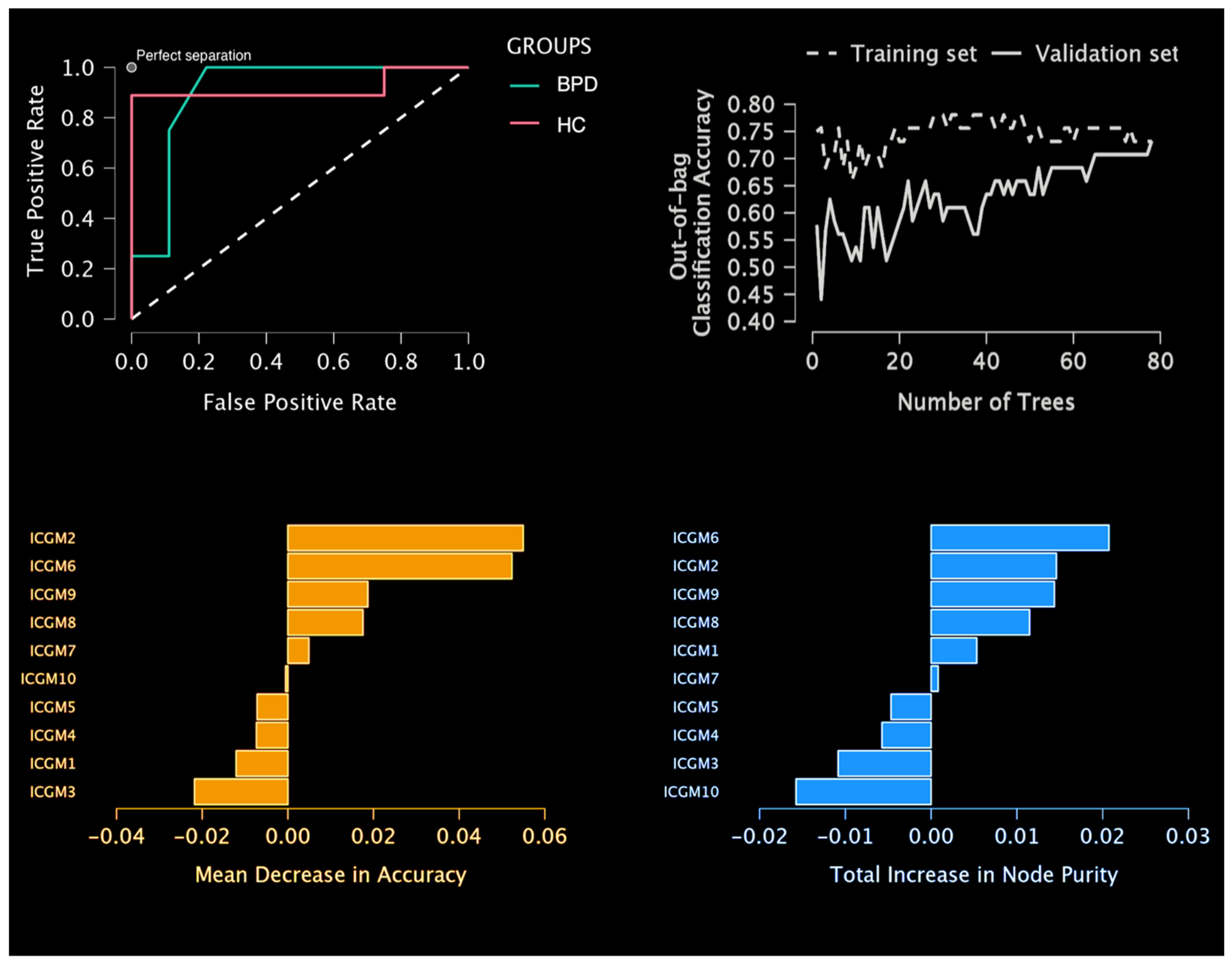
4.2. Predictive Model Results
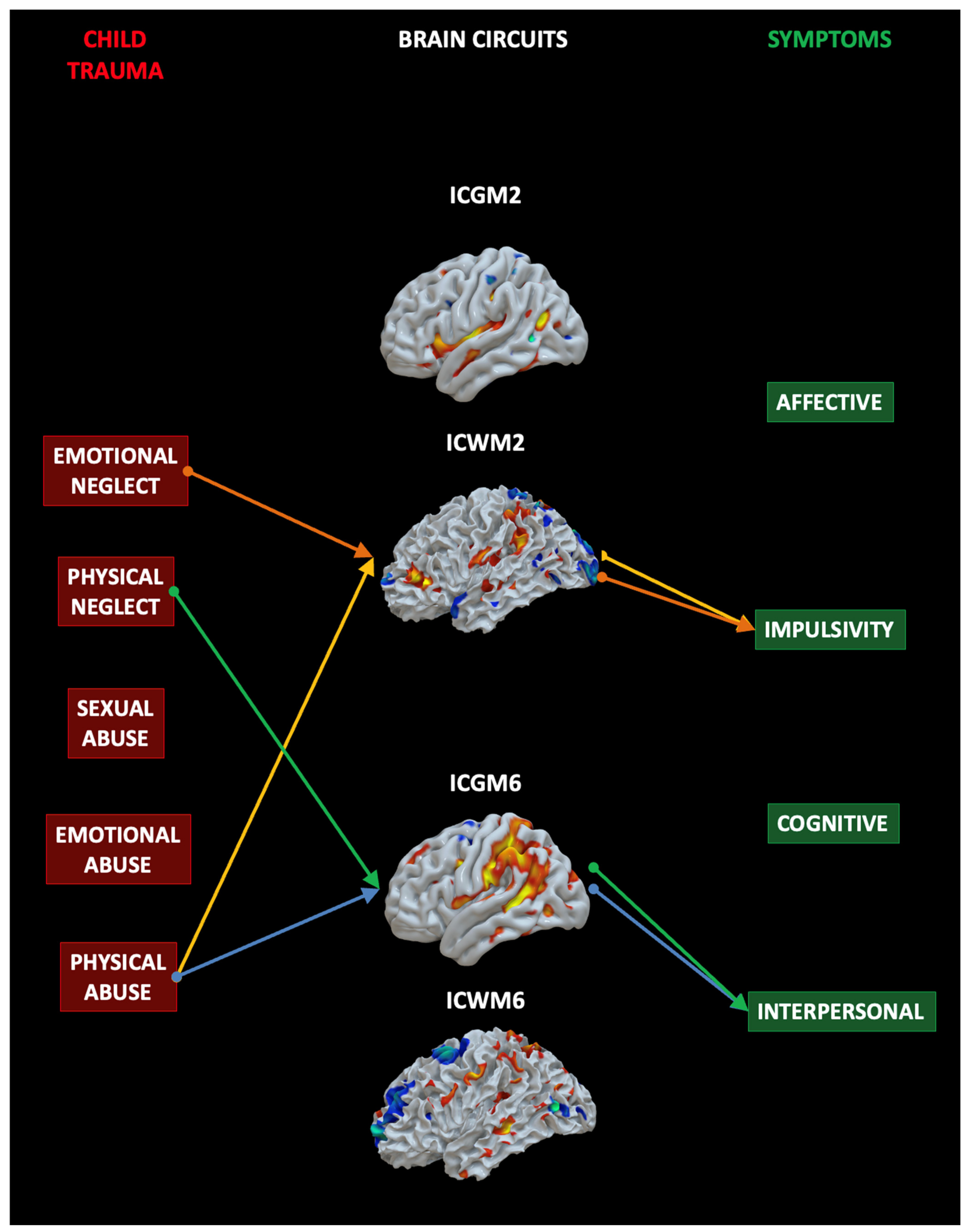
4.3. Mediation Analysis
5. Discussion
5.1. Impulsivity Network
5.2. Interpersonal Network
5.3. The Impact of Specific Childhood Traumas on the Brain and Symptoms
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koenigsberg, H.W.; Harvey, P.D.; Mitropoulou, V.; Schmeidler, J.; New, A.S.; Goodman, M.; Silverman, J.M.; Serby, M.; Schopick, F.; Siever, L.J. Characterizing affective instability in borderline personality disorder. Am. J. Psychiatry 2002, 159, 784–788. [Google Scholar] [CrossRef]
- Zanarini, M.C. Psychotherapy of borderline personality disorder. Acta Psychiatr. Scand. 2009, 120, 373–377. [Google Scholar] [CrossRef]
- Lenzenweger, M.F.; Lane, M.C.; Loranger, A.W.; Kessler, R.C. DSM-IV Personality disorders in the national comorbidity survey replication. Biol. Psychiatry 2007, 62, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Trull, T.J.; Jahng, S.; Tomko, R.L.; Wood, P.K.; Sher, K.J. Revised NESARC personality disorder diagnoses: Gender, prevalence, and comorbidity with substance dependence disorders. J. Pers. Disord. 2010, 24, 412–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomko, R.; Trull, T.J.; Wood, P.K.; Sher, K.J. Characteristics of borderline personality disorder in a community sample: Comorbidity, treatment utilization, and general functioning. J. Pers. Disord. 2014, 28, 734–750. [Google Scholar] [CrossRef]
- De Panfilis, C.; Schito, G.; Generali, I.; Gozzi, L.A.; Ossola, P.; Marchesi, C.; Grecucci, A. Emotions at the border: Increased punishment behavior during fair interpersonal exchanges in borderline personality disorder. J. Abnorm. Psychol. 2019, 128, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Meng, Y.-J.; Li, X.-J.; Zhang, C.; Liang, S.; Li, M.-L.; Li, Z.; Guo, W.; Wang, Q.; Deng, W.; et al. Common and distinct patterns of grey matter alterations in borderline personality disorder and bipolar disorder: Voxel-based meta-analysis. Br. J. Psychiatry 2019, 215, 395–403. [Google Scholar] [CrossRef]
- Wolf, R.C.; Thomann, P.A.; Sambataro, F.; Vasic, N.; Schmid, M.; Wolf, N.D. Orbitofrontal cortex and impulsivity in borderline personality disorder: An MRI study of baseline brain perfusion. Eur. Arch. Psychiatry Clin. Neurosci. 2012, 262, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Ducasse, D.; Courtet, P.; Olié, E. Physical and social pains in borderline disorder and neuroanatomical correlates: A systematic review. Curr. Psychiatry Rep. 2014, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wrege, J.S.; Ruocco, A.C.; Euler, S.; Preller, K.H.; Busmann, M.; Meya, L.; Schmidt, A.; Lang, U.E.; Borgwardt, S.; Walter, M. Negative affect moderates the effect of social rejection on frontal and anterior cingulate cortex activation in borderline personality disorder. Cogn. Affect. Behav. Neurosci. 2019, 19, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Lou, J.; Sun, Y.; Cui, Z.; Gong, L. Common and distinct patterns of gray matter alterations in borderline personality disorder and posttraumatic stress disorder: A dual meta-analysis. Neurosci. Lett. 2020, 741, 135376. [Google Scholar] [CrossRef] [PubMed]
- Pan, N.; Wang, S.; Qin, K.; Li, L.; Chen, Y.; Zhang, X.; Lai, H.; Suo, X.; Long, Y.; Yu, Y.; et al. Common and distinct neural patterns of attention-deficit/hyperactivity disorder and borderline personality disorder: A multimodal functional and structural meta-analysis. In Biological Psychiatry: Cognitive Neuroscience and Neuroimaging; Elsevier: Amsterdam, The Netherlands, 2022. [Google Scholar] [CrossRef]
- Irle, E.; Lange, C.; Sachsse, U. Reduced size and abnormal asymmetry of parietal cortex in women with borderline personality disorder. Biol. Psychiatry 2005, 57, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Lyoo, I.K.; Han, M.H.; Cho, D.Y. A brain MRI study in subjects with borderline personality disorder. J. Affect. Disord. 1998, 50, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Hazlett, E.A.; New, A.S.; Newmark, R.; Haznedar, M.M.; Lo, J.N.; Speiser, L.J.; Chen, A.D.; Mitropoulou, V.; Minzenberg, M.; Siever, L.J.; et al. Reduced anterior and posterior cingulate gray matter in borderline personality disorder. Biol. Psychiatry 2005, 58, 614–623. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K. Voxel-based morphometry—The methods. Neuroimage 2000, 11, 805–821. [Google Scholar] [CrossRef] [Green Version]
- Soloff, P.H.; Pruitt, P.; Sharma, M.; Radwan, J.; White, R.; Diwadkar, V.A. Structural brain abnormalities and suicidal behavior in borderline personality disorder. J. Psychiatr. Res. 2012, 46, 516–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedtfeld, I.; Schulze, L.; Krause-Utz, A.; Demirakca, T.; Bohus, M.; Schmahl, C. Voxel-based morphometry in women with borderline personality disorder with and without comorbid posttraumatic stress disorder. PLoS ONE 2013, 8, e65824. [Google Scholar] [CrossRef]
- Schulze, L.; Schmahl, C.; Niedtfeld, I. Neural correlates of disturbed emotion processing in borderline personality disorder: A multimodal meta-analysis. Biol. Psychiatry 2015, 79, 97–106. [Google Scholar] [CrossRef]
- Aguilar-Ortiz, S.; Salgado-Pineda, P.; Marco-Pallarés, J.; Pascual, J.C.; Vega, D.; Soler, J.; Brunel, C.; Martin-Blanco, A.; Soto, A.; Ribas, J.; et al. Abnormalities in gray matter volume in patients with borderline personality disorder and their relation to lifetime depression: A VBM study. PLoS ONE 2018, 13, e0191946. [Google Scholar] [CrossRef]
- Dadomo, H.; Grecucci, A.; Giardini, I.; Ugolini, E.; Carmelita, A.; Panzeri, M. Schema therapy for emotional dysregulation: Theoretical implication and clinical applications. Front. Psychol. 2016, 7, 1987. [Google Scholar] [CrossRef] [Green Version]
- Frederickson, J.J.; Messina, I.; Grecucci, A. Dysregulated anxiety and dysregulating defenses: Toward an emotion regulation informed dynamic psychotherapy. Front. Psychol. 2018, 9, 2054. [Google Scholar] [CrossRef] [Green Version]
- Ruocco, A.C.; Amirthavasagam, S.; Zakzanis, K.K. Amygdala and hippocampal volume reductions as candidate endophenotypes for borderline personality disorder: A meta-analysis of magnetic resonance imaging studies. Psychiatry Res. Neuroimaging 2012, 201, 245–252. [Google Scholar] [CrossRef]
- Sorella, S.; Lapomarda, G.; Messina, I.; Frederickson, J.J.; Siugzdaite, R.; Job, R.; Grecucci, A. Testing the expanded continuum hypothesis of schizophrenia and bipolar disorder. Neural and psychological evidence for shared and distinct mechanisms. NeuroImage Clin. 2019, 23, 101854. [Google Scholar] [CrossRef] [PubMed]
- Lapomarda, G.; Grecucci, A.; Messina, I.; Pappaianni, E.; Dadomo, H. Common and different gray and white matter alterations in bipolar and borderline personality disorder: A source-based morphometry study. Brain Res. 2021, 1762, 147401. [Google Scholar] [CrossRef]
- Pappaianni, E.; De Pisapia, N.; Siugzdaite, R.; Crescentini, C.; Calcagnì, A.; Job, R.; Grecucci, A. Less is more: Psychological and morphometric differences between low vs high reappraisers. Cogn. Affect. Behav. Neurosci. 2020, 20, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Grecucci, A.; Lapomarda, G.; Messina, I.; Monachesi, B.; Sorella, S.; Siugzdaite, R. Structural features related to affective instability correctly classify patients with borderline personality disorder. A supervised machine learning approach. Front. Psychiatry 2022, 13, 804440. [Google Scholar] [CrossRef]
- Sui, J.; Pearlson, G.; Caprihan, A.; Adali, T.; Kiehl, K.A.; Liu, J.; Yamamoto, J.; Calhoun, V.D. Discriminating schizophrenia and bipolar disorder by fusing fMRI and DTI in a multimodal CCA+ joint ICA model. Neuroimage 2011, 57, 839–855. [Google Scholar] [CrossRef] [Green Version]
- Mechelli, A.; Viera, S. Machine Learning: Methods and Applications to Brain Disorders; Academic Press: Cambridge, MA, USA, 2019. [Google Scholar]
- Calhoun, V.; Adali, T.; Giuliani, N.; Pekar, J.; Kiehl, K.; Pearlson, G. Method for multimodal analysis of independent source differences in schizophrenia: Combining gray matter structural and auditory oddball functional data. Hum. Brain Mapp. 2005, 27, 47–62. [Google Scholar] [CrossRef]
- Correa, N.M.; Li, Y.-O.; Adali, T.; Calhoun, V.D. Canonical Correlation analysis for feature-based fusion of biomedical imaging modalities and its application to detection of associative networks in schizophrenia. IEEE J. Sel. Top. Signal Process. 2008, 2, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Plis, S.M.; Weisend, M.P.; Damaraju, E.; Eichele, T.; Mayer, A.; Clark, V.P.; Lane, T.; Calhoun, V.D. Effective connectivity analysis of fMRI and MEG data collected under identical paradigms. Comput. Biol. Med. 2011, 41, 1156–1165. [Google Scholar] [CrossRef] [Green Version]
- Sui, J.; He, H.; Yu, Q.; Chen, J.; Rogers, J.; Pearlson, G.D.; Mayer, A.; Bustillo, J.; Canive, J.; Calhoun, V.D. Combination of resting state fMRI, DTI, and sMRI data to discriminate schizophrenia by N-way MCCA + jICA. Front. Hum. Neurosci. 2013, 7, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grecucci, A.; Sorella, S.; Consolini, J. Decoding individual differences in expressing and suppressing anger from structural brain networks: A supervised machine learning approach. Behav. Brain Res. 2023, 439, 114245. [Google Scholar] [CrossRef]
- Ho, T.K. The random subspace method for constructing decision forests. IEEE Trans. Pattern Anal. Mach. Intell. 1998, 20, 832–844. [Google Scholar] [CrossRef] [Green Version]
- Ho, B.C.; Andreasen, N.C. Do structural brain deficits worsen following onset of first-episode schizophrenia? In Schizophrenia Research; Elsevier: Amsterdam, The Netherlands, 2002; Volume 53, p. 98. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Fossati, A.; Madeddu, F.; Maffei, C. Borderline personality disorder and childhood sexual abuse: A meta-analytic study. J. Pers. Disord. 1999, 13, 268–280. [Google Scholar] [CrossRef]
- Porter, C.; Palmier-Claus, J.; Branitsky, A.; Mansell, W.; Warwick, H.; Varese, F. Childhood adversity and borderline personality disorder: A meta-analysis. Acta Psychiatr. Scand. 2019, 141, 6–20. [Google Scholar] [CrossRef] [PubMed]
- Cicchetti, D.; Rogosch, F.A.; Hecht, K.F.; Crick, N.R.; Hetzel, S. Moderation of maltreatment effects on childhood borderline personality symptoms by gender and oxytocin receptor and FK506 binding protein 5 genes. Dev. Psychopathol. 2014, 26, 831–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hecht, K.F.; Cicchetti, D.; Rogosch, F.A.; Crick, N.R. Borderline personality features in childhood: The role of subtype, developmental timing, and chronicity of child maltreatment. Dev. Psychopathol. 2014, 26, 805–815. [Google Scholar] [CrossRef] [Green Version]
- Belsky, D.W.; Caspi, A.; Arseneault, L.; Bleidorn, W.; Fonagy, P.; Goodman, M.; Houts, R.; Moffitt, T. Etiological features of borderline personality related characteristics in a birth cohort of 12-year-old children. Dev. Psychopathol. 2012, 24, 251–265. [Google Scholar] [CrossRef] [Green Version]
- Gratz, K.L.; Latzman, R.D.; Tull, M.T.; Reynolds, E.K.; Lejuez, C. Exploring the association between emotional abuse and childhood borderline personality features: The moderating role of personality traits. Behav. Ther. 2011, 42, 493–508. [Google Scholar] [CrossRef] [Green Version]
- Winsper, C.; Zanarini, M.; Wolke, D. Prospective study of family adversity and maladaptive parenting in childhood and borderline personality disorder symptoms in a non-clinical population at 11 years. Psychol. Med. 2012, 42, 2405–2420. [Google Scholar] [CrossRef] [Green Version]
- Widom, C.S.; Czaja, S.J.; Paris, J. A prospective investigation of borderline personality disorder in abused and neglected children followed up into adulthood. J. Pers. Disord. 2009, 23, 433–446. [Google Scholar] [CrossRef]
- Dadomo, H.; Salvato, G.; Lapomarda, G.; Ciftci, Z.; Messina, I.; Grecucci, A. Structural features predict sexual trauma and interpersonal problems in borderline personality disorder but not in controls: A multi-voxel pattern analysis. Front. Hum. Neurosci. 2022, 16, 773593. [Google Scholar] [CrossRef]
- Bernstein, D.P.; Fink, L.; Handelsman, L.; Foote, J. Childhood trauma questionnaire. Assessment of Family Violence: A Handbook for Researchers and Practitioners; American Psychological Association: Washington, DC, USA, 1994. [Google Scholar]
- Zanarini, M.C. Zanarini Rating Scale for borderline personality disorder (ZAN-BPD): A continuous measure of DSM-IV borderline psychopathology. J. Pers. Disord. 2003, 17, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Aron, A.R.; Robbins, T.W.; Poldrack, R.A. Inhibition and the right inferior frontal cortex. Trends Cogn. Sci. 2004, 8, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Penny, W.D.; Mattout, J.; Trujillo-Barreto, N. Bayesian model selection and averaging. Statistical Parametric Mapping: The Analysis of Functional Brain Images; Elsevier: London, UK, 2006. [Google Scholar]
- Gaser, C.; Dahnke, R.; Thompson, P.M.; Kurth, F.; Luders, E. CAT—A computational anatomy toolbox for the analysis of structural MRI data. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ashburner, J. A fast diffeomorphic image registration algorithm. Neuroimage 2007, 38, 95–113. [Google Scholar] [CrossRef]
- Acar, E.; Levin-Schwartz, Y.; Calhoun, V.D.; Adali, T. Tensor-based fusion of EEG and FMRI to understand neurological changes in schizophrenia. In Proceedings of the 2017 IEEE International Symposium on Circuits and Systems (ISCAS), Baltimore, MD, USA, 28–31 May 2017; pp. 1–4. [Google Scholar] [CrossRef] [Green Version]
- Wax, M.; Kailath, T. Detection of signals by information theoretic criteria. IEEE Trans. Acoust. Speech Signal Process. 1985, 33, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Himberg, J.; Hyvärinen, A.; Esposito, F. Validating the independent components of neuroimaging time series via clustering and visualization. Neuroimage 2004, 22, 1214–1222. [Google Scholar] [CrossRef]
- Himberg, J.; Hyvarinen, A. Icasso: Software for investigating the reliability of ICA estimates by clustering and visualization. In Proceedings of the 2003 IEEE XIII Workshop on Neural Networks for Signal Processing (IEEE Cat. No.03TH8718), Toulouse, France, 17–19 September 2003; pp. 259–268. [Google Scholar] [CrossRef] [Green Version]
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning; Springer: New York, NY, USA, 2001. [Google Scholar]
- Kozak, M.J.; Cuthbert, B.N. The NIMH research domain criteria initiative: Background, issues, and pragmatics. Psychophysiology 2016, 53, 286–297. [Google Scholar] [CrossRef]
- Baggio, T.; Grecucci, A.; Meconi, F.; Messina, I. Anxious brains: A combined data fusion machine learning approach to predict trait anxiety from morphometric features. Sensors 2023, 23, 610. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.R.; Potenza, M.N. Recent insights into the neurobiology of impulsivity. Curr. Addict. Rep. 2014, 1, 309–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bari, A.; Robbins, T.W. Inhibition and impulsivity: Behavioral and neural basis of response control. Prog. Neurobiol. 2013, 108, 44–79. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.-E.; Kim, S.; Seo, S.; Kang, W.; Kim, A.; Kang, Y.; Choi, K.W.; Tae, W.-S.; Ham, B.-J.; Han, K.-M. Association of prefrontal cortex thinning with high impulsivity in healthy adults. Psychiatry Investig. 2021, 18, 570–579. [Google Scholar] [CrossRef]
- Lee, T.M.; Chan, C.C.; Han, S.-H.; Leung, A.W.; Fox, P.T.; Gao, J.-H. An event-related fMRI study on risk taking by healthy individuals of high or low impulsiveness. Neurosci. Lett. 2008, 438, 138–141. [Google Scholar] [CrossRef]
- Pan, N.; Wang, S.; Zhao, Y.; Lai, H.; Qin, K.; Li, J.; Biswal, B.B.; Sweeney, J.A.; Gong, Q. Brain gray matter structures associated with trait impulsivity: A systematic review and voxel-based meta-analysis. Hum. Brain Mapp. 2021, 42, 2214–2235. [Google Scholar] [CrossRef]
- Hirjak, D.; Thomann, A.K.; Kubera, K.M.; Wolf, R.C.; Jeung, H.; Maier-Hein, K.H.; Thomann, P.A. Cortical folding patterns are associated with impulsivity in healthy young adults. Brain Imaging Behav. 2016, 11, 1592–1603. [Google Scholar] [CrossRef]
- Fineberg, N.A.; Chamberlain, S.R.; Goudriaan, A.E.; Stein, D.J.; Vanderschuren, L.J.M.J.; Gillan, C.M.; Shekar, S.; Gorwood, P.A.P.M.; Voon, V.; Morein-Zamir, S.; et al. New developments in human neurocognition: Clinical, genetic, and brain imaging correlates of impulsivity and compulsivity. CNS Spectr. 2014, 19, 69–89. [Google Scholar] [CrossRef] [Green Version]
- Dalley, J.W.; Everitt, B.J.; Robbins, T.W. Impulsivity, compulsivity, and top-down cognitive control. Neuron 2011, 69, 680–694. [Google Scholar] [CrossRef] [Green Version]
- Carrington, S.J.; Bailey, A.J. Are there theory of mind regions in the brain? A review of the neuroimaging literature. Hum. Brain Mapp. 2009, 30, 2313–2335. [Google Scholar] [CrossRef]
- Schurz, M.; Radua, J.; Tholen, M.G.; Maliske, L.; Margulies, D.S.; Mars, R.B.; Sallet, J.; Kanske, P. Toward a hierarchical model of social cognition: A neuroimaging meta-analysis and integrative review of empathy and theory of mind. Psychol. Bull. 2021, 147, 293–327. [Google Scholar] [CrossRef] [PubMed]
- Messina, I.; Sambin, M.; Beschoner, P.; Viviani, R. Changing views of emotion regulation and neurobiological models of the mechanism of action of psychotherapy. Cogn. Affect. Behav. Neurosci. 2016, 16, 571–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kernberg, O. Borderline personality organization. J. Am. Psychoanal. Assoc. 1967, 15, 641–685. [Google Scholar] [CrossRef] [PubMed]
- Roy, A. Childhood trauma and impulsivity. Possible relevance to suicidal behavior. Arch. Suicide Res. 2005, 9, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Sharkawy, A.-N. Principle of neural network and its main types: Review. Adv. Appl. Comput. Math. 2020, 7, 8–19. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BPD | HC | p-Values | |
|---|---|---|---|
| Participants | 20 | 45 | |
| Age | 35.75 (±8.61) | 36.69 (±8.46) | p = 0.401 |
| Gender | F = 17, M = 3 | F = 34, M = 11 | p = 0.647 |
| Education | ≥8 years of formal education | ||
| Exclusion criteria | Neurological disease, psychoactive substance, pregnancy, MRI contraindications, previous head injury | Neurological disease, psychoactive substance, mental illness (SCID-II, SCID-IV), pregnancy, MRI contraindications, previous head injury | |
| Area | Brodmann Area | Volume (cc) | Random Effects: Max Value (x, y, z) |
|---|---|---|---|
| (A) IC2 Increased Grey Matter Concentration | |||
| Postcentral gyrus | 2, 3, 40 | 1.0/1.5 | 7.4 (−45, −26, 36)/10.0 (43, −24, 39) |
| Precentral gyrus | 13 | 0.1/0.5 | 4.2 (−45, −21, 37)/8.7 (46, −21, 37) |
| Angular gyrus | * | 0.0/0.8 | 0 (0, 0, 0)/8.7 (40, −58, 33) |
| Sub-gyral | 37 | 1.3/2.0 | 6.4 (−22, 7, 47)/7.9 (40, −24, 36) |
| Middle temporal gyrus | 19, 21 | 1.9/0.1 | 7.9 (−39, −63, 22)/4.0 (58, −45, −2) |
| Insula | 13, 45 | 3.8/3.8 | 7.4 (−37, −4, 11)/6.3 (39, −11, 14) |
| Middle frontal gyrus | 6, 8, 11 | 0.4/1.8 | 5.4 (−24, 4, 44)/7.1 (25, 17, 41) |
| Precuneus | 7, 31, 39 | 0.5/2.4 | 5.9 (−16, −63, 21)/6.6 (13, −61, 38) |
| Cerebellar tonsil | * | 1.2/1.2 | 6.4 (−27, −44, −42)/6.2 (33, −46, −39) |
| Superior parietal lobule | 7 | 0.3/0.4 | 5.9 (−25, −52, 43)/6.3 (30, −55, 43) |
| Superior frontal gyrus | 6 | 0.4/0.2 | 6.0 (−22, 11, 48)/4.1 (22, 15, 43) |
| Pyramis | * | 0.4/0.6 | 4.4 (−9, −80, −23)/5.7 (3, −80, −25) |
| Inferior parietal lobule | 40 | 0.2/0.7 | 4.4 (−28, −49, 43)/5.7 (40, −59, 38) |
| Fusiform gyrus | 18, 36, 37 | 0.5/1.0 | 3.9 (−48, −42, −21)/5.7 (45, −43, −12) |
| Uncus (inc amygdala) | 20, 28, 36 | 0.4/0.0 | 5.6 (−30, −9, −29)/0 (0, 0, 0) |
| Extra-nuclear | * | 0.9/0.1 | 5.6 (−34, 6, 5)/4.0 (37, −11, 7) |
| Medial frontal gyrus | * | 0.2/0.0 | 5.6 (−24, 36, 27)/0 (0, 0, 0) |
| Culmen | * | 0.9/0.2 | 5.4 (−1, −48, −1)/4.9 (3, −48, −1) |
| Claustrum | * | 0.4/0.4 | 4.8 (−34, −10, 9)/4.4 (36, −4, 6) |
| Declive | * | 0.2/0.5 | 3.9 (−4, −81, −21)/4.7 (6, −83, −20) |
| Superior temporal gyrus | 39, 41, 42 | 0.4/0.0 | 4.6 (−48, −24, 7)/0 (0, 0, 0) |
| Inferior frontal gyrus | 47 | 0.6/0.1 | 4.0 (−37, 25, 0)/4.6 (40, 6, 33) |
| (B) IC2 Decreased Grey Matter Concentration | |||
| Posterior cingulate | 30, 31 | 2.3/1.8 | 13.4 (−22, −58, 8)/11.6 (22, −64, 10) |
| Cuneus | 17, 18, 19, 23, 30 | 4.0/5.1 | 11.7 (−16, −69, 10)/13.0 (21, −68, 10) |
| Extra-nuclear | * | 0.6/0.8 | 11.7 (−21, −53, 8)/10.7 (25, −55, 8) |
| Thalamus | * | 3.8/2.0 | 9.3 (−10, −17, 9)/5.9 (9, −13, 8) |
| Lingual gyrus | 18, 19 | 3.7/1.4 | 9.1 (−18, −52, 5)/6.8 (22, −54, 5) |
| Lateral ventricle | * | 0.3/0.5 | 6.0 (−28, −58, 8)/8.3 (28, −58, 8) |
| Middle temporal gyrus | 39 | 0.8/0.3 | 8.1 (−50, −55, 7)/5.2 (34, −72, 19) |
| Sub-gyral | * | 0.3/1.0 | 4.7 (−27, −89, 2)/6.8 (28, −54, 5) |
| Cerebellar tonsil | * | 0.0/0.6 | 0(0, 0, 0)/6.4 (12, −56, −41) |
| Anterior cingulate | 32 | 0.4/0.6 | 4.0 (−9, 26, 25)/5.8 (10, 41, 8) |
| Precuneus | 7 | 0.4/0.1 | 5.8 (−19, −62, 43)/4.2 (4, −73, 24) |
| Inferior parietal lobule | 40 | 0.3/0.5 | 5.7 (−40, −36, 57)/4.4 (53, −26, 28) |
| Middle frontal gyrus | 9 | 0.6/0.5 | 5.6 (−37, 15, 38)/5.2 (39, 18, 31) |
| Postcentral gyrus | 1, 2, 3, 40 | 1.5/0.1 | 5.6 (−48, −19, 51)/4.1 (64, −28, 21) |
| Middle occipital gyrus | 18, 19 | 0.4/0.1 | 5.4 (−28, −86, 4)/3.8 (34, −75, 16) |
| Inferior semi-lunar lobule | * | 0.0/0.6 | 0 (0, 0, 0)/4.8 (9, −59, −41) |
| (C) IC2 Increased White Matter Concentration | |||
| Middle temporal gyrus | 37, 39 | 0.2/1.2 | 4.8 (−56, −56, 10)/9.9 (59, −51, −5) |
| Inferior temporal gyrus | 37 | 0.0/0.6 | 0 (0, 0, 0)/8.4 (62, −54, −5) |
| Middle frontal gyrus | 6, 8, 9, 10, 11, 46 | 2.2/3.6 | 5.5 (−45, 30, 39)/7.6 (49, 34, 33) |
| Inferior frontal gyrus | 10, 46, 47 | 1.4/0.0 | 7.1 (−48, 33, −13)/0 (0, 0, 0) |
| Superior frontal gyrus | 6, 8, 9, 10 | 1.2/1.8 | 6.5 (−19, 50, 38)/7.0 (22, 0, 65) |
| Superior temporal gyrus | 13, 22, 39 | 1.0/1.2 | 4.8 (−52, −16, 9)/6.9 (61, −49, 15) |
| Claustrum | * | 0.3/0.4 | 5.7 (−34, −6, 6)/6.8 (36, −5, 7) |
| Insula | 13 | 1.7/1.1 | 6.5 (−36, −2, 8)/5.8 (36, −5, 11) |
| Postcentral gyrus | 1, 2, 3, 5 | 1.2/0.3 | 6.4 (−56, −27, 39)/5.2 (67, −14, 33) |
| Inferior parietal lobule | 40 | 2.2/1.5 | 6.0 (−46, −44, 48)/6.3 (52, −50, 43) |
| Extra-nuclear | * | 0.6/0.8 | 4.6 (−34, −3, 3)/6.2 (36, −1, 7) |
| Precuneus | 7 | 0.6/0.3 | 5.7 (−9, −54, 48)/4.8 (22, −63, 46) |
| Superior parietal lobule | 7 | 0.0/0.4 | 0 (0, 0, 0)/5.1 (25, −60, 44) |
| Lentiform nucleus | * | 0.6/0.3 | 4.5 (−15, 3, −5)/4.0 (15, 7, −4) |
| Cingulate gyrus | 23, 24 | 0.4/0.4 | 4.3 (−1, 3, 27)/4.5 (3, 1, 28) |
| Anterior cingulate | 24, 32 | 0.3/0.9 | 3.7 (−3, 35, 1)/4.3 (3, 35, 1) |
| (D) IC2 Decreased White Matter Concentration | |||
| Inferior parietal lobule | 7, 39, 40 | 0.8/2.7 | 8.1 (−28, −47, 56)/10.3 (34, −48, 56) |
| Sub-gyral | 7, 20, 40 | 1.9/2.2 | 9.6 (−28, −50, 54)/8.0 (31, −44, 51) |
| Superior parietal lobule | 7 | 1.0/0.6 | 9.5 (−30, −51, 58)/9.1 (34, −49, 61) |
| Precuneus | 7, 31 | 1.4/0.7 | 9.0 (−28, −50, 49)/6.0 (30, −47, 48) |
| Fusiform gyrus | 20, 36, 37 | 0.4/0.1 | 7.8 (−40, −17, −24)/4.1 (50, −42, −18) |
| Cuneus | 7, 17, 18, 19, 30 | 3.6/3.3 | 6.6 (−10, −76, 31)/6.7 (28, −83, 26) |
| Middle temporal gyrus | 19, 21, 39 | 0.1/1.3 | 3.7 (−62, −52, 0)/6.6 (55, −56, 8) |
| Lingual gyrus | 18, 19 | 1.5/0.8 | 5.8 (−21, −64, 1)/4.6 (22, −63, 2) |
| Postcentral gyrus | 1, 3, 5 | 0.8/0.0 | 5.6 (−43, −30, 62)/−999.0 (0, 0, 0) |
| Inferior occipital gyrus | 18 | 0.6/0.1 | 5.6 (−34, −89, −3)/4.2 (48, −80, −2) |
| Superior frontal gyrus | 9, 10, 11 | 1.2/1.0 | 5.5 (−15, 65, −10)/5.2 (16, 66, −13) |
| Inferior temporal gyrus | 20 | 0.5/0.1 | 5.5 (−43, −17, −27)/4.1 (50, −56, −12) |
| Middle occipital gyrus | 18, 19, 37 | 0.8/1.9 | 5.2 (−34, −89, 1)/5.2 (36, −79, 14) |
| Posterior cingulate | 30 | 0.5/0.7 | 5.1 (−21, −64, 6)/4.8 (19, −58, 7) |
| Area | Brodmann Area | Volume (cc) | Random Effects: Max Value (x, y, z) |
|---|---|---|---|
| (A). ICGM6 Increased Grey Matter Concentration | |||
| Postcentral gyrus | 1, 2, 3, 4, 5, 7, 40, 43 | 6.1/7.4 | 9.3 (−52, −20, 30)/9.8 (53, −17, 31) |
| Middle temporal gyrus | 21, 22, 39 | 2.9/1.0 | 9.7 (−49, −41, 5)/6.2 (50, −43, 9) |
| Precuneus | 7, 19, 39 | 1.1/2.2 | 5.0 (−21, −62, 42)/9.0 (30, −61, 35) |
| Parahippocampal gyrus | 19, 36, 37 | 3.3/2.0 | 8.7 (−28, −43, −7)/7.7 (30, −44, −5) |
| Inferior parietal lobule | 40 | 4.2/5.4 | 5.9 (−55, −23, 30)/8.7 (55, −28, 26) |
| Precentral gyrus | 4, 6, 9, 13, 43 | 0.9/2.2 | 6.0 (−36, 4, 29)/8.4 (50, −17, 34) |
| Superior temporal gyrus | 22, 39, 41, 42 | 3.4/3.4 | 8.2 (−48, −49, 13)/8.1 (53, −45, 12) |
| Superior parietal lobule | 7 | 0.4/1.0 | 5.9 (−22, −59, 44)/8.2 (27, −58, 43) |
| Sub-gyral | 43 | 1.6/3.1 | 7.0 (−46, −43, 5)/8.1 (30, −64, 32) |
| Inferior frontal gyrus | 9 | 0.5/0.0 | 7.8 (−39, 4, 32)/0 (0, 0, 0) |
| Fusiform gyrus | 19, 20, 37 | 1.2/0.6 | 7.6 (−30, −36, −12)/5.6 (28, −47, −8) |
| Insula | 13, 41 | 0.1/1.9 | 3.8 (−46, −15, 12)/7.1 (46, −22, 16) |
| Angular gyrus | 39 | 0.9/0.0 | 6.5 (−42, −58, 32)/0 (0, 0, 0) |
| Transverse temporal gyrus | 41, 42 | 0.1/0.7 | 3.9 (−53, −14, 12)/6.4 (48, −21, 12) |
| Supramarginal gyrus | 40 | 1.3/0.2 | 6.4 (−39, −53, 27)/4.8 (52, −48, 22) |
| Anterior cingulate | 24, 32 | 1.3/0.4 | 6.3 (−3, 32, −7)/4.9 (3, 29, −10) |
| Cuneus | 18, 19 | 0.2/1.3 | 4.0 (−7, −79, 14)/5.1 (10, −88, 14) |
| Culmen | * | 1.3/2.6 | 4.7 (−22, −41, −12)/4.9 (31, −39, −22) |
| Cingulate gyrus | 31 | 0.9/0.3 | 4.7 (−1, −32, 36)/4.4 (1, −32, 39) |
| Lingual gyrus | 18, 19 | 0.8/0.1 | 4.6 (−18, −55, −2)/3.6 (22, −53, −2) |
| Declive | * | 0.8/0.0 | 4.1 (−18, −59, −13)/0 (0, 0, 0) |
| (B). ICGM6 Decreased Grey Matter Concentration | |||
| Sub-gyral | 6 | 0.8/0.8 | 6.4 (−21, 4, 51)/5.2 (25, −6, 56) |
| Middle temporal gyrus | * | 0.1/0.4 | 3.8 (−48, −63, 2)/5.8 (42, −66, 16) |
| Middle frontal gyrus | 6, 8 | 0.9/1.9 | 4.7 (−33, −2, 43)/5.6 (28, −6, 53) |
| Fusiform gyrus | 20 | 0.1/0.5 | 4.0 (−42, −33, −16)/5.0 (43, −27, −18) |
| Medial frontal gyrus | 6 | 0.6/0.2 | 4.9 (−18, 7, 51)/4.5 (10, −10, 61) |
| Superior frontal gyrus | 6 | 0.5/0.3 | 4.4 (−13, −11, 63)/4.1 (18, −7, 63) |
| (C). ICWM6 Increased White Matter Concentration | |||
| Middle frontal gyrus | 9, 46 | 1.4/0.1 | 10.2 (−46, 31, 33)/4.1 (48, 39, 26) |
| Precuneus | 7, 19 | 2.8/2.0 | 5.8 (−22, −74, 41)/9.9 (24, −70, 49) |
| Superior parietal lobule | 7 | 0.2/0.8 | 4.8 (−39, −57, 51)/8.8 (25, −69, 45) |
| Inferior parietal lobule | 39, 40 | 1.2/4.4 | 6.2 (−62, −40, 24)/8.4 (49, −42, 56) |
| Middle occipital gyrus | 19 | 0.9/0.0 | 7.3 (−39, −85, 17)/0 (0, 0, 0) |
| Precentral gyrus | 4, 6, 44 | 0.8/4.1 | 5.8 (−61, −6, 21)/7.3 (55, −17, 35) |
| Supramarginal gyrus | 40 | 0.7/0.2 | 7.1 (−62, −42, 27)/4.5 (49, −36, 34) |
| Postcentral gyrus | 1, 2, 3, 7, 40, 43 | 2.0/3.9 | 5.6 (−43, −29, 53)/7.1 (53, −16, 31) |
| Superior temporal gyrus | 21, 22, 41, 42 | 0.4/2.2 | 5.0 (−65, −40, 21)/7.0 (67, −27, 15) |
| Cuneus | 17, 19 | 0.4/0.6 | 4.2 (−7, −92, 3)/6.1 (31, −82, 30) |
| Sub-gyral | * | 0.7/0.3 | 6.1 (−46, −36, −13)/5.0 (18, −45, 62) |
| Transverse temporal gyrus | 41 | 0.0/0.4 | 0 (0, 0, 0)/5.9 (48, −29, 12) |
| Insula | 13 | 0.0/0.6 | 0 (0, 0, 0)/5.8 (52, −30, 18) |
| Middle temporal gyrus | 19, 21, 39 | 0.8/0.3 | 5.7 (−50, −75, 19)/5.3 (49, −63, 21) |
| Lingual gyrus | 17, 19 | 0.8/0.0 | 5.1 (−22, −64, 1)/0 (0, 0, 0) |
| Superior frontal gyrus | 6, 8, 9, 10 | 0.4/0.1 | 4.6 (−22, 46, 39)/3.7 (12, 14, 50) |
| (D). ICWM6 Decreased White Matter Concentration | |||
| Precentral gyrus | 6, 9 | 1.3/0.1 | 8.1 (−37, −6, 62)/3.7 (39, 16, 35) |
| Superior frontal gyrus | 6, 8, 9, 10, 11 | 5.1/3.0 | 7.3 (−10, −4, 67)/8.0 (25, −1, 66) |
| Middle frontal gyrus | 6, 8, 9, 10, 46 | 7.7/2.6 | 7.6 (−34, −3, 62)/7.0 (31, −3, 61) |
| Middle occipital gyrus | 19, 37 | 0.4/0.1 | 6.5 (−49, −67, 5)/3.8 (27, −70, 6) |
| Superior parietal lobule | 7 | 0.4/0.0 | 6.4 (−28, −54, 61)/0 (0, 0, 0) |
| Medial frontal gyrus | 6, 11 | 1.3/0.7 | 5.2 (−13, −4, 61)/6.4 (15, 1, 58) |
| Middle temporal gyrus | 21, 37, 39 | 0.2/0.6 | 5.5 (−46, −64, 6)/5.2 (68, −32, −7) |
| Sub-gyral | 6 | 1.6/0.3 | 4.6 (−25, 15, 41)/5.3 (18, −2, 57) |
| Insula | 13 | 0.4/0.4 | 4.5 (−36, −5, 10)/4.6 (37, 0, 8) |
| Cingulate gyrus | 32 | 0.3/0.5 | 3.8 (−18, 8, 44)/4.4 (15, 16, 36) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grecucci, A.; Dadomo, H.; Salvato, G.; Lapomarda, G.; Sorella, S.; Messina, I. Abnormal Brain Circuits Characterize Borderline Personality and Mediate the Relationship between Childhood Traumas and Symptoms: A mCCA+jICA and Random Forest Approach. Sensors 2023, 23, 2862. https://doi.org/10.3390/s23052862
Grecucci A, Dadomo H, Salvato G, Lapomarda G, Sorella S, Messina I. Abnormal Brain Circuits Characterize Borderline Personality and Mediate the Relationship between Childhood Traumas and Symptoms: A mCCA+jICA and Random Forest Approach. Sensors. 2023; 23(5):2862. https://doi.org/10.3390/s23052862
Chicago/Turabian StyleGrecucci, Alessandro, Harold Dadomo, Gerardo Salvato, Gaia Lapomarda, Sara Sorella, and Irene Messina. 2023. "Abnormal Brain Circuits Characterize Borderline Personality and Mediate the Relationship between Childhood Traumas and Symptoms: A mCCA+jICA and Random Forest Approach" Sensors 23, no. 5: 2862. https://doi.org/10.3390/s23052862