Functional Near-Infrared Spectroscopy for the Classification of Motor-Related Brain Activity on the Sensor-Level

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
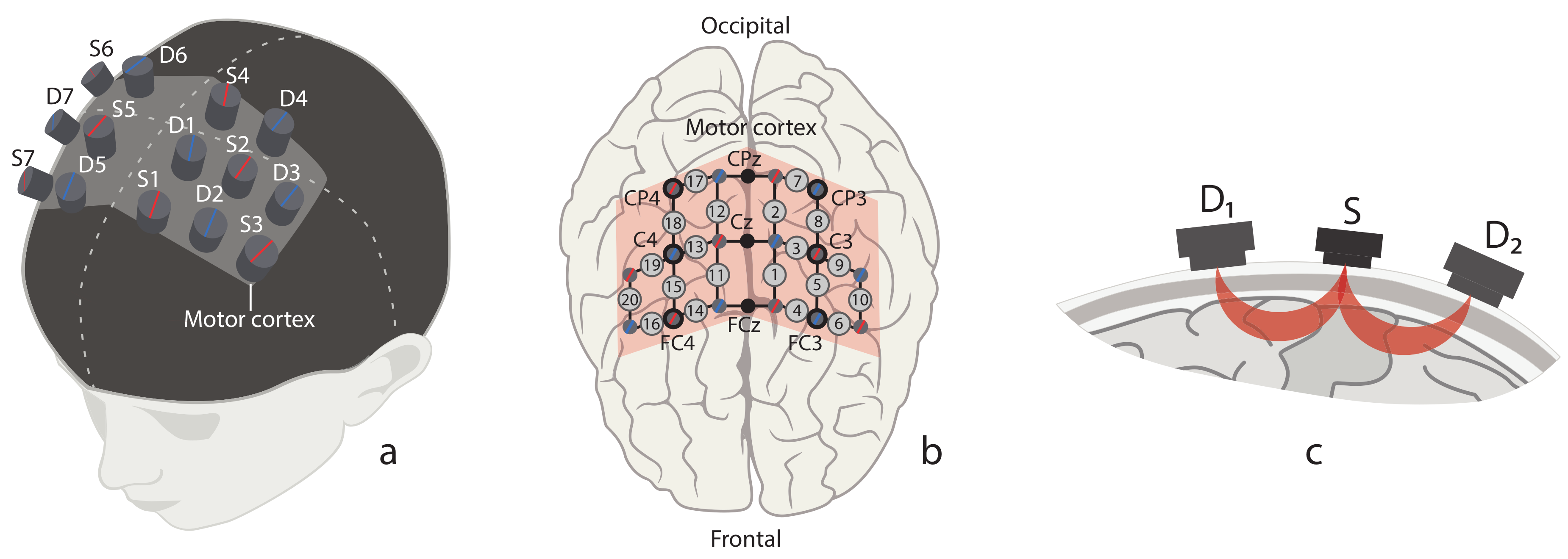
2.2. Experimental Equipment
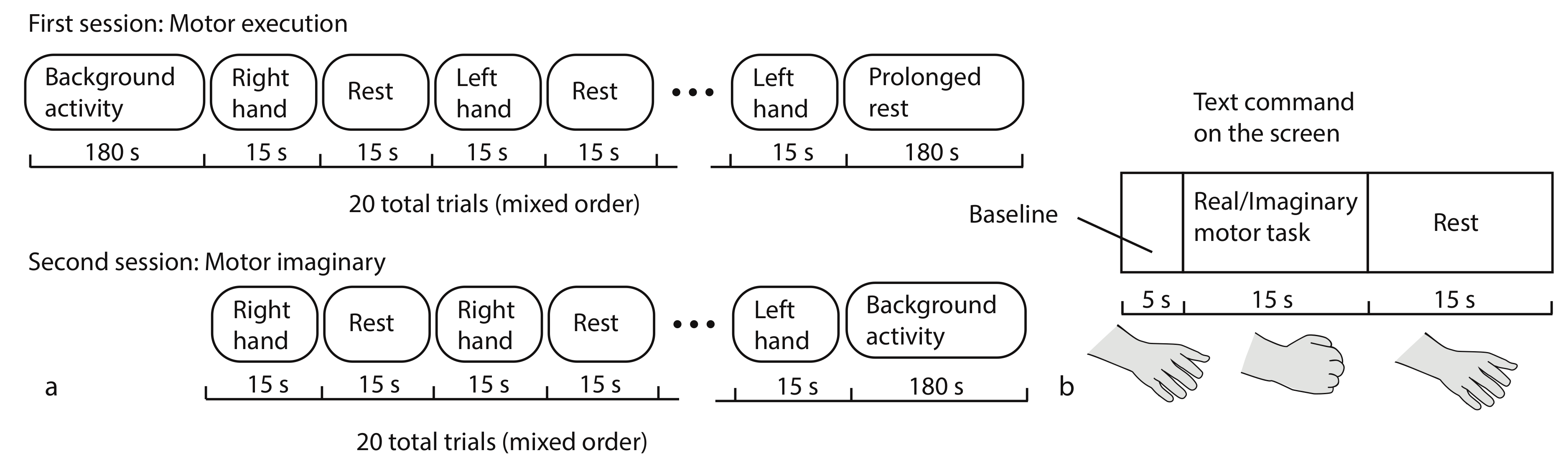
2.3. Experimental Procedure
2.4. Data Acquisition and Pre-Processing
2.5. Data Analysis
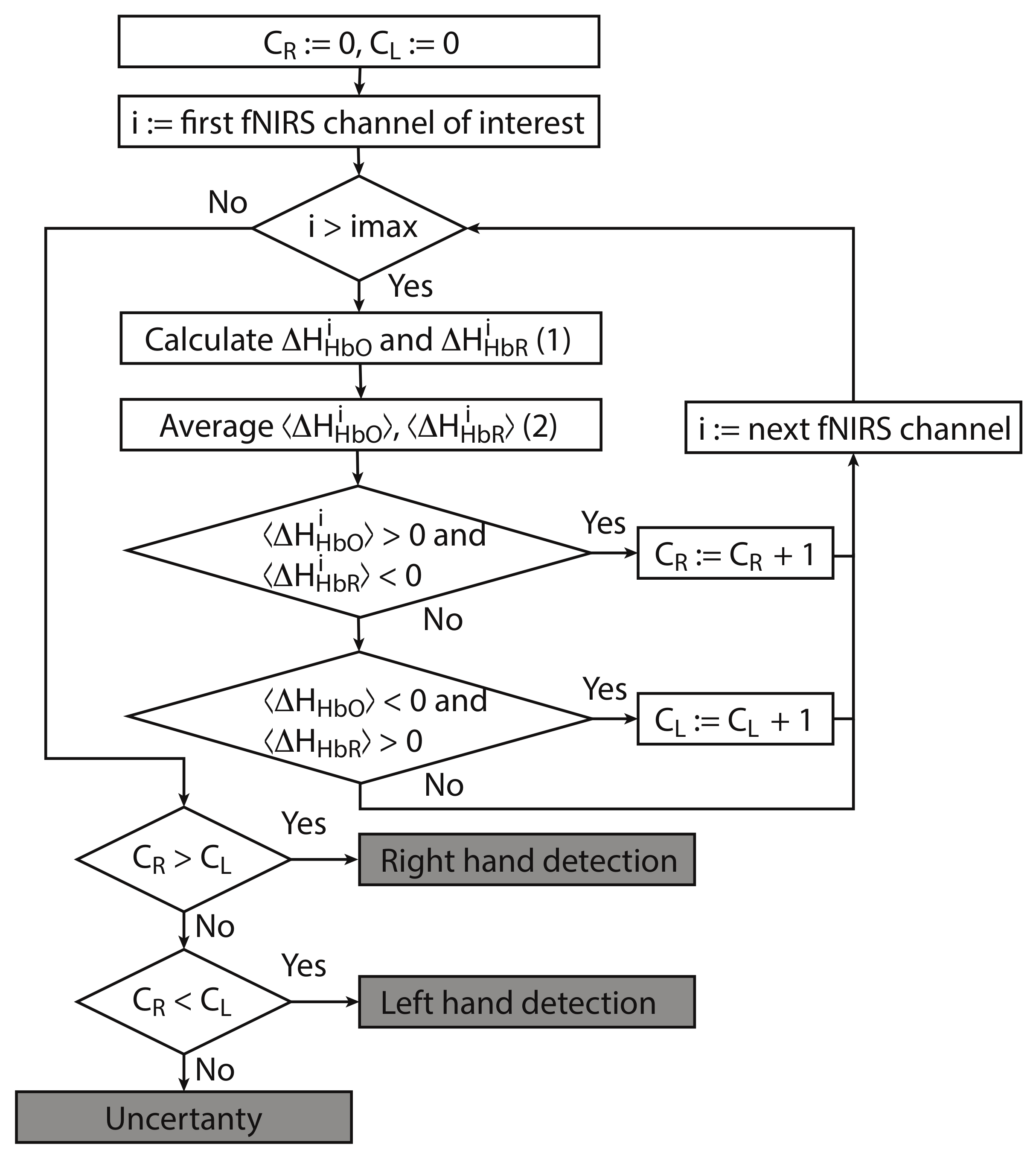
2.6. Algorithm for Classification of Brain Activity During Real and Imagery Motor Executions
- For each considered channel i and type of motor activity (right/left hand, execution/imagery) we subtract spatial oxyhemoglobin (HbO) () and deoxyhemoglobin (HbR) () distributions for the right hemisphere (, fNIRS channels j of interest from right hemisphere) from the corresponding distribution for the left hemisphere (, symmetrical channels i from left hemisphere). Similar to our approach (2) which uses average values, we calculate differences for individual symmetrical channels in the left and right hemispheres as
- Then, we average and over the time interval corresponding to motor activity s to find and as
- For each separate fNIRS signal trial, we calculate characteristics and taking into account the following criteria for each considered symmetric fNIRS channels in the left and right hemispheres.
- (i)
- If and is true for one of the channels i, then (value takes discrete values, minimal value is and peak value is equal to the number of considered fNIRS channels of interest in one of the hemispheres).
- (ii)
- If and is true for one of the channels i, then (value takes discrete values, minimal value is and peak value is the same as peak value of ).
- Finally, we make a decision according to the following criteria.
- (i)
- If , then right-hand (real or imaginary) motor activity takes place.
- (ii)
- If , then left-hand (real or imaginary) activity takes place.
- (iii)
- If , then the type of activity is uncertain.
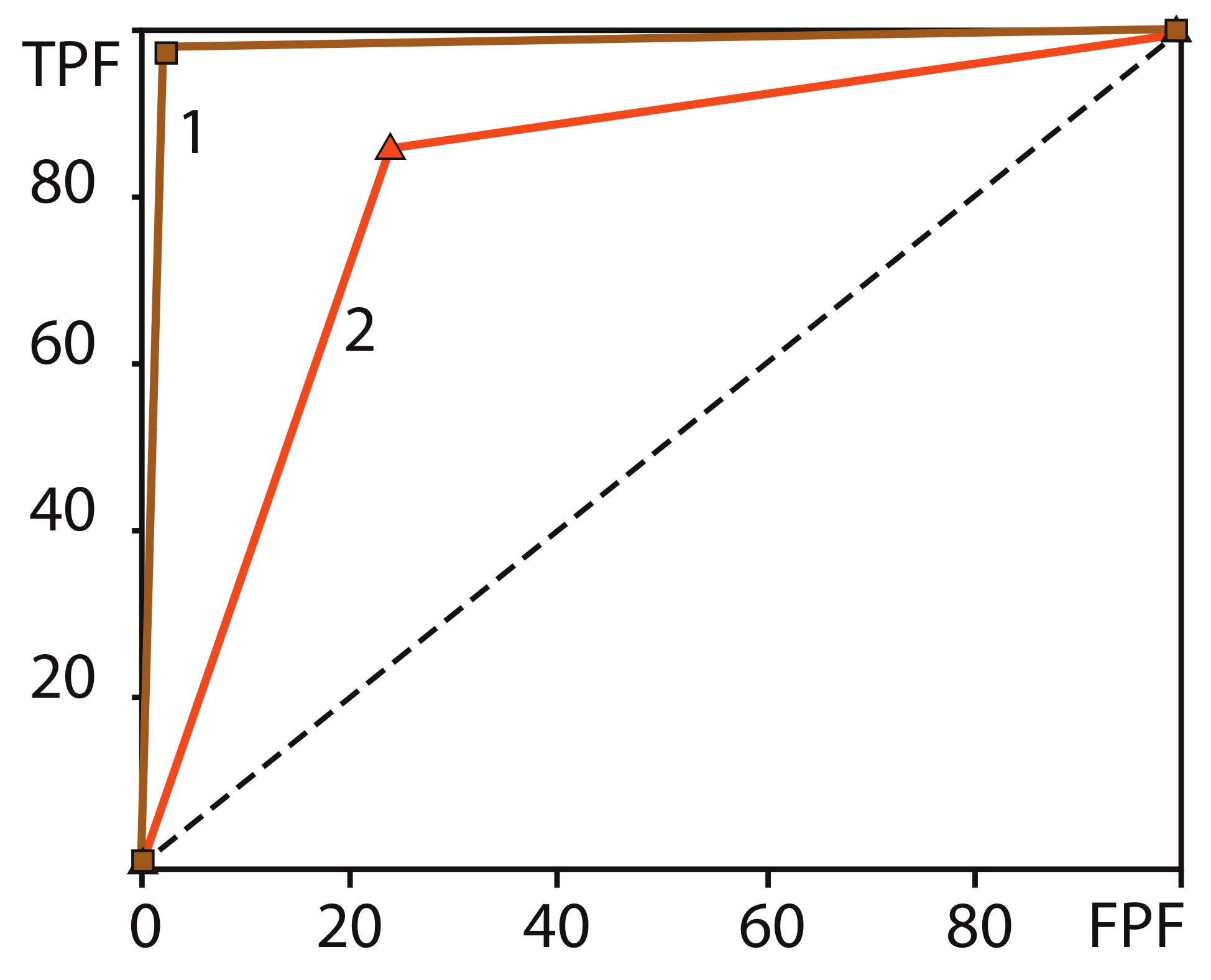
2.7. Estimation of Classification Accuracy of Brain Activity During Motor Execution and Motor Imagery
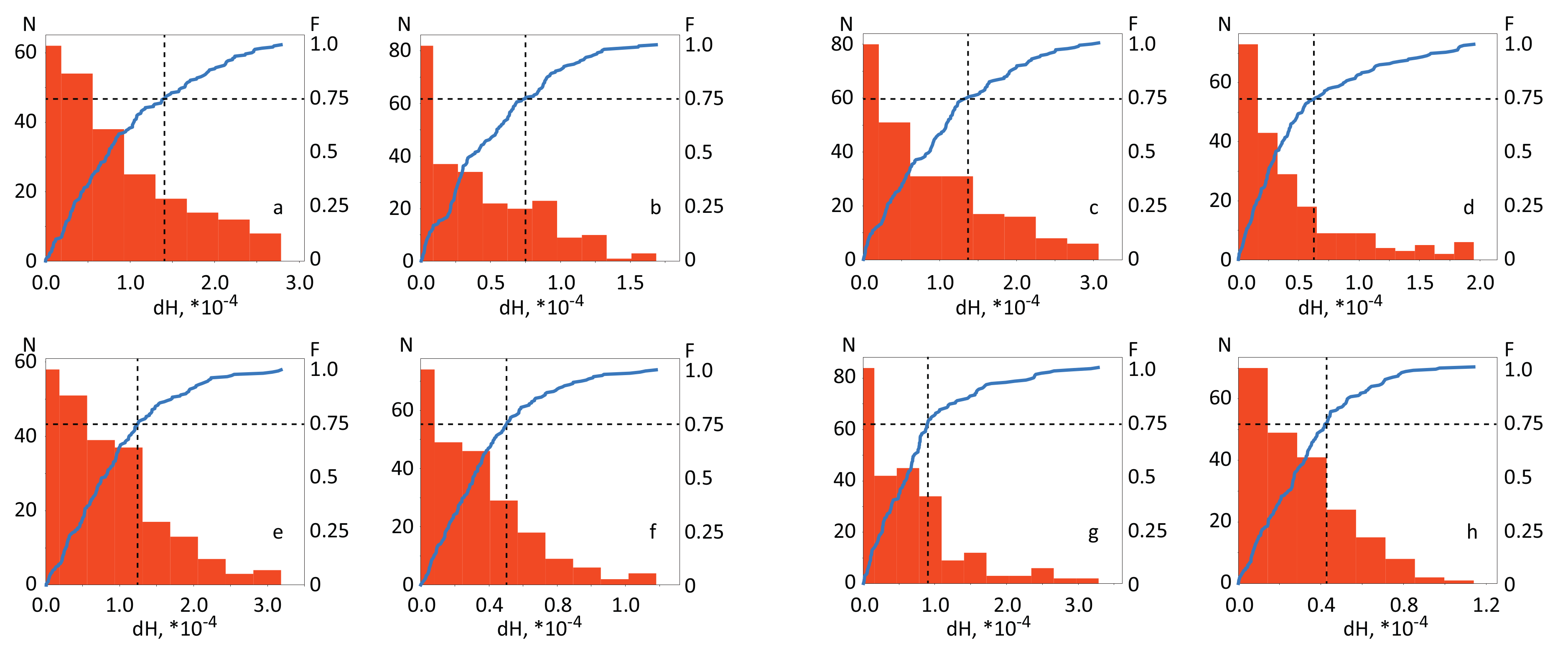
3. Results
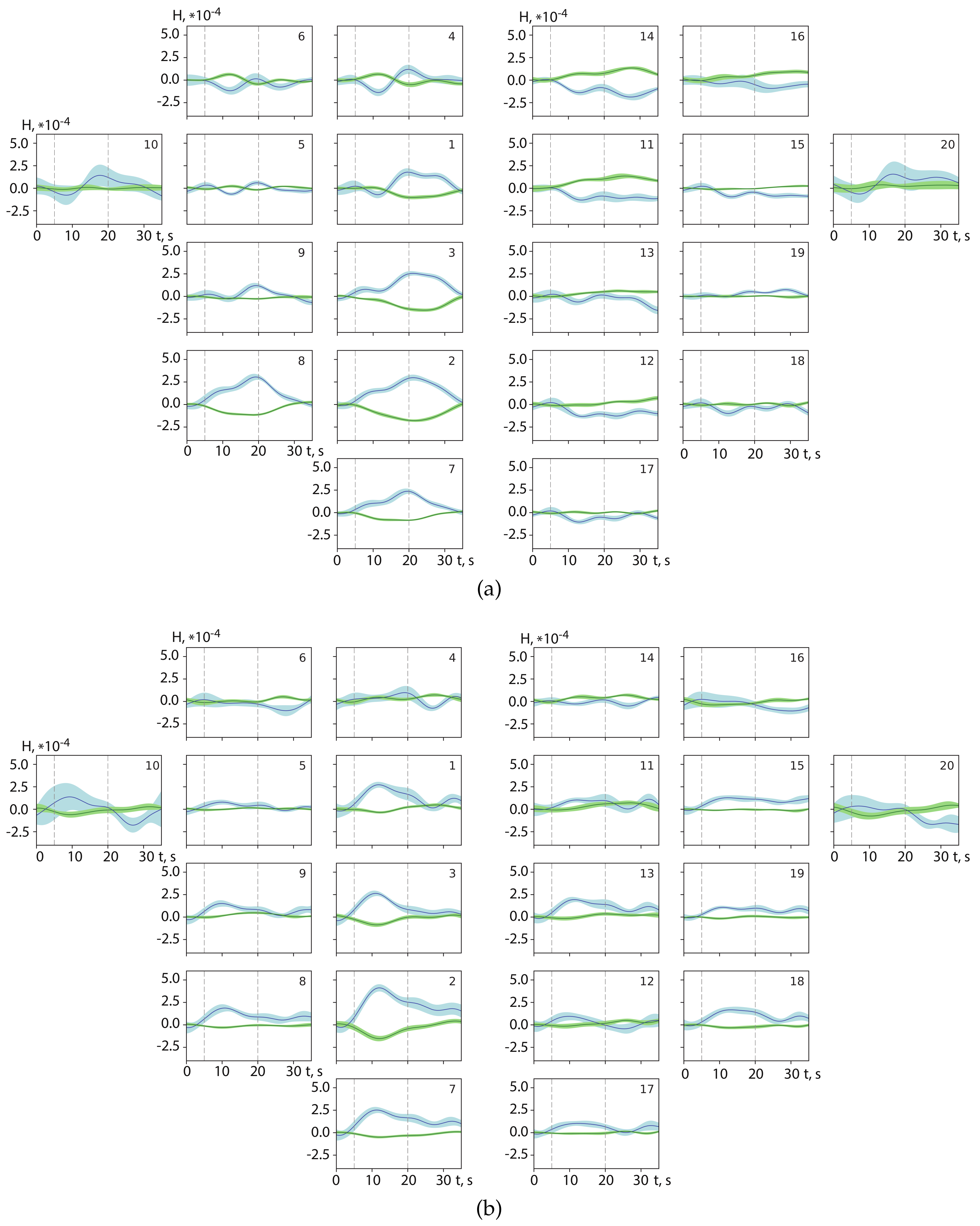
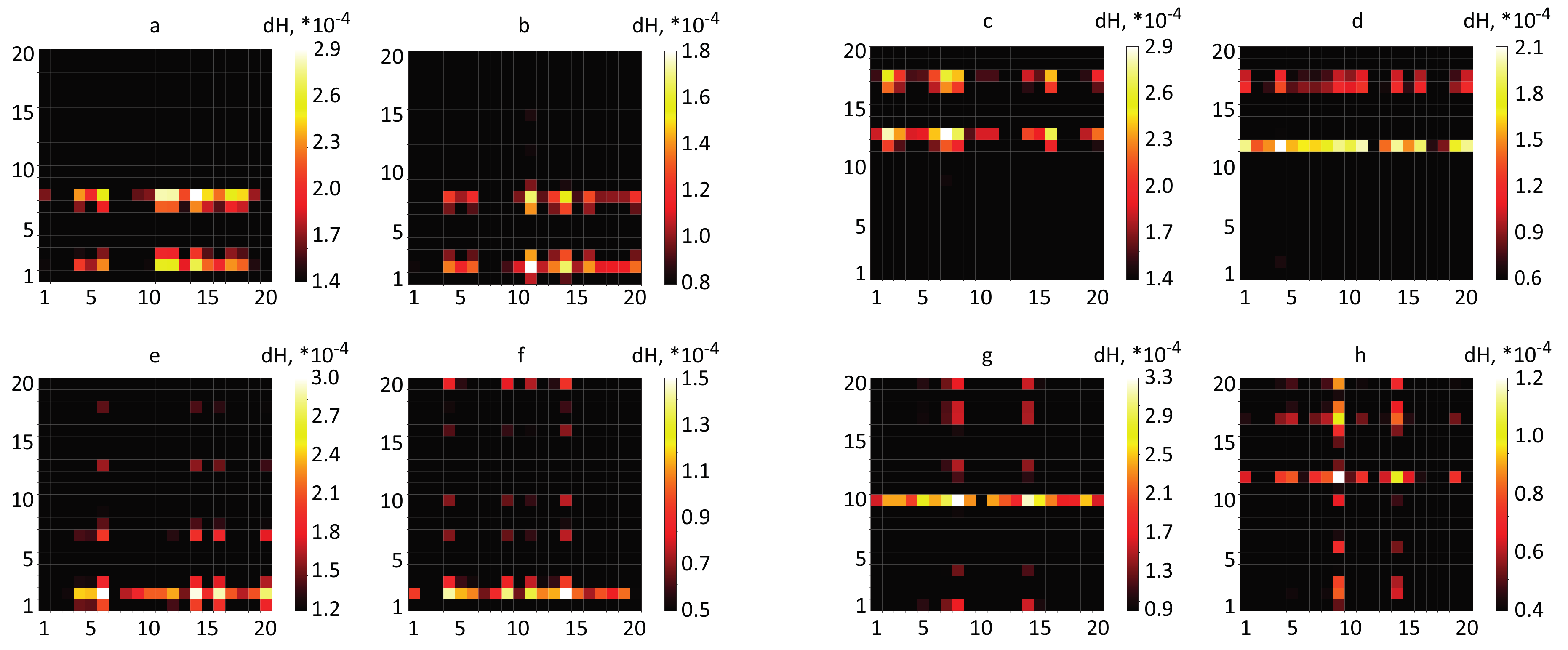
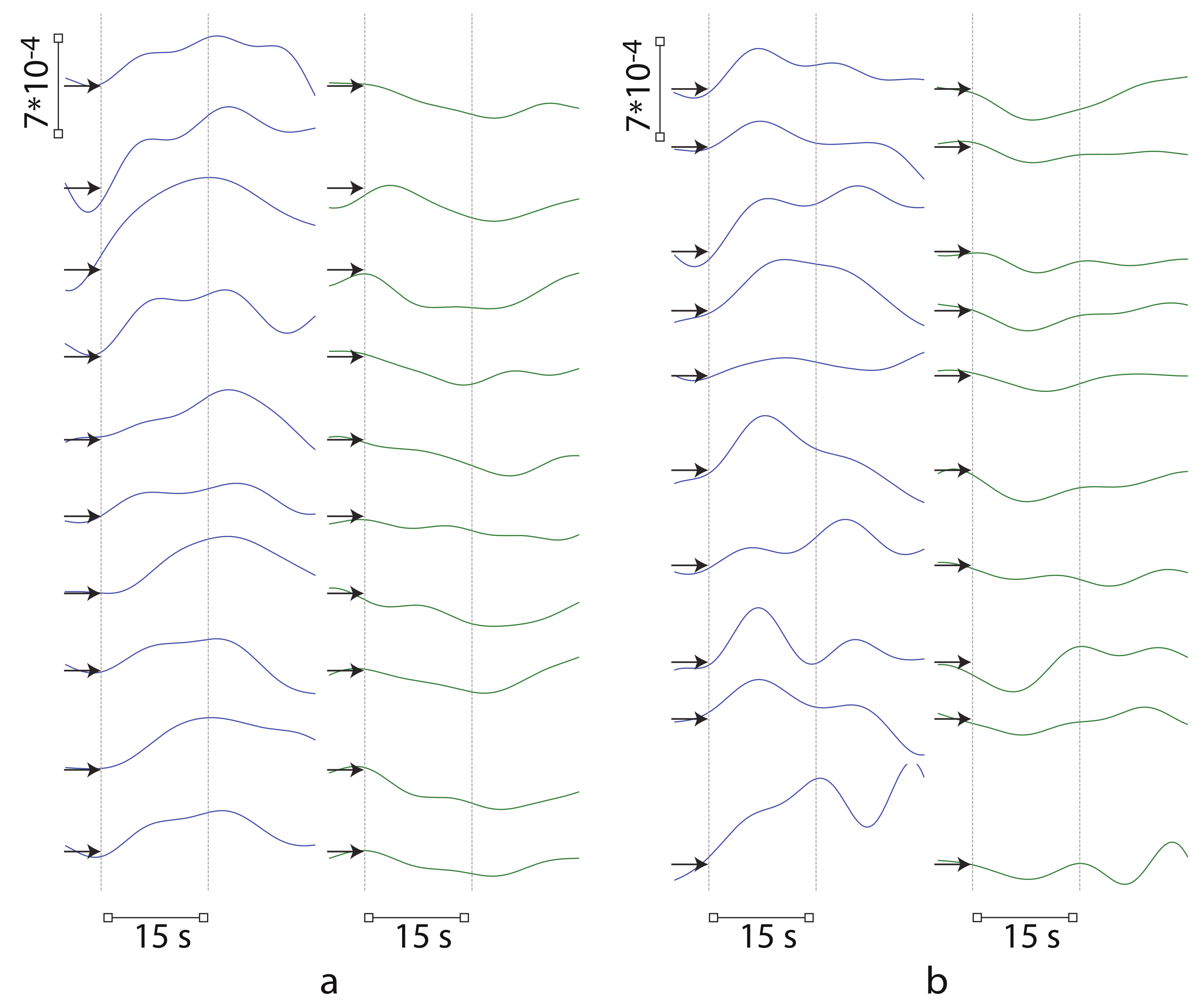
3.1. Spatial Brain Activity During Motor Execution and Motor Imagery
3.2. Results of Real-Time Classification of Brain Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| fNIRS | Functional near-infrared spectroscopy |
| BCI | Brain-computer interface |
| EEG | Electroencephalography |
| MRI | Magnetic resonance imaging |
| MEG | Magnetoencephalography |
| ERS | Event-related synchronization |
| ERD | Event-related desynchronization |
| HbO | oxygenated |
| HbR | deoxygenated |
| SVM | Support vector machine |
| ICA | Independent component analysis |
| ROC | Receiver operating characteristic |
References
- Rabinovich, M.I.; Varona, P.; Selverston, A.I.; Abarbanel, H.D.I. Dynamical principles in neuroscience. Rev. Mod. Phys. 2006, 78, 1213–1265. [Google Scholar] [CrossRef] [Green Version]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 2011, 12, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Hramov, A.E.; Frolov, N.S.; Maksimenko, V.A.; Makarov, V.V.; Koronovskii, A.A.; Garcia-Prieto, J.; Antón-Toro, L.F.; Maestú, F.; Pisarchik, A.N. Artificial neural network detects human uncertainty. Chaos 2018, 28, 033607. [Google Scholar] [CrossRef] [PubMed]
- Maksimenko, V.A.; Kurkin, S.A.; Pitsik, E.N.; Musatov, V.Y.; Runnova, A.E.; Efremova, T.Y.; Hramov, A.E.; Pisarchik, A.N. Artificial neural network classification of motor-related EEG: An increase in classification accuracy by reducing signal complexity. Complexity 2018, 2018, 9385947. [Google Scholar] [CrossRef] [Green Version]
- Edge, L. Working across disciplines in neuroscience. Cell 2019, 179, 283–284. [Google Scholar]
- Herculano-Houzel, S. The human brain in numbers: A linearly scaled-up primate brain. Front. Hum. Neurosci. 2009, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Niedermeyer, E.; Fernando, L.S. Electroencephalography: Basic Principles, Clinical Applications, and Related Fields; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2004. [Google Scholar]
- Breakspear, M. Dynamic models of large-scale brain activity. Nat. Neurosci. 2017, 20, 340. [Google Scholar] [CrossRef]
- Maksimenko, V.A.; Runnova, A.E.; Frolov, N.S.; Makarov, V.V.; Nedaivozov, V.; Koronovskii, A.A.; Pisarchik, A.; Hramov, A.E. Multiscale neural connectivity during human sensory processing in the brain. Phys. Rev. E 2018, 97, 052405. [Google Scholar] [CrossRef] [Green Version]
- Maksimenko, V.A.; Frolov, N.S.; Hramov, A.E.; Runnova, A.E.; Grubov, V.V.; Kurths, J.; Pisarchik, A.N. Neural interactions in a spatially-distributed cortical network during perceptual decision-making. Front. Behav. Neurosci. 2019, 13, 220. [Google Scholar] [CrossRef] [Green Version]
- Buch, E.; Weber, C.; Cohen, L.G.; Braun, C.; Dimyan, M.A.; Ard, T.; Mellinger, J.; Caria, A.; Soekadar, S.; Fourkas, A.; et al. Think to move: A neuromagnetic brain-computer interface (BCI) system for chronic stroke. Stroke 2008, 39, 910–917. [Google Scholar] [CrossRef] [Green Version]
- Teo, W.P.; Chew, E. Is motor-imagery brain-computer interface feasible in stroke rehabilitation? PM R 2014, 6, 723–728. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, U.; Birbaumer, N.; Ramos-Murguialday, A. Brain-computer interfaces for communication and rehabilitation. Nat. Rev. Neurol. 2016, 12, 513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tucker, M.R.; Olivier, J.; Pagel, A.; Bleuler, H.; Bouri, M.; Lambercy, O.; del Millán, J.R.; Riener, R.; Vallery, H.; Gassert, R. Control strategies for active lower extremity prosthetics and orthotics: A review. J. Neuroeng. Rehabil. 2015, 12, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curran, E.A.; Stokes, M.J. Learning to control brain activity: A review of the production and control of EEG components for driving brain–computer interface (BCI) systems. Brain Cognit. 2003, 51, 326–336. [Google Scholar] [CrossRef]
- Chholak, P.; Niso, G.; Maksimenko, V.A.; Kurkin, S.A.; Frolov, N.S.; Pitsik, E.N.; Hramov, A.E.; Pisarchik, A.N. Visual and kinesthetic modes affect motor imagery classification in untrained subjects. Sci. Rep. 2019, 9, 9838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolpaw, J.; Wolpaw, E.W. Brain-Computer Interfaces: Principles and Practice; OUP USA: New York, NY, USA, 2012. [Google Scholar]
- Abdalmalak, A.; Milej, D.; Norton, L.; Debicki, D.; Gofton, T.; Diop, M.; Owen, A.M.; Lawrence, K.S. Single-session communication with a locked-in patient by functional near-infrared spectroscopy. Neurophotonics 2017, 4, 040501. [Google Scholar] [CrossRef]
- Abdalmalak, A.; Milej, D.; Diop, M.; Shokouhi, M.; Naci, L.; Owen, A.M.; Lawrence, K.S. Can time-resolved NIRS provide the sensitivity to detect brain activity during motor imagery consistently? Biomed. Opt. Express 2017, 8, 2162–2172. [Google Scholar] [CrossRef] [Green Version]
- Tomita, Y.; Vialatte, F.B.; Dreyfus, G.; Mitsukura, Y.; Bakardjian, H.; Cichocki, A. Bimodal BCI using simultaneously NIRS and EEG. IEEE Trans. Biomed. Eng. 2014, 61, 1274–1284. [Google Scholar] [CrossRef]
- Schudlo, L.C.; Chau, T. Towards a ternary NIRS-BCI: Single-trial classification of verbal fluency task, stroop task and unconstrained rest. J. Neural Eng. 2015, 12, 066008. [Google Scholar] [CrossRef]
- Power, S.D.; Kushki, A.; Chau, T. Automatic single-trial discrimination of mental arithmetic, mental singing and the no-control state from prefrontal activity: Toward a three-state NIRS-BCI. BMC Res. Notes 2012, 5, 141. [Google Scholar] [CrossRef] [Green Version]
- Nicolas-Alonso, L.F.; Gomez-Gil, J. Brain computer interfaces, a review. Sensors 2012, 12, 1211–1279. [Google Scholar] [CrossRef] [PubMed]
- Lemm, S.; Schafer, C.; Curio, G. BCI competition 2003-data set III: Probabilistic modeling of sensorimotor/ spl mu/rhythms for classification of imaginary hand movements. IEEE Trans. Biomed. Eng. 2004, 51, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Jeannerod, M. Mental imagery in the motor context. Neuropsychologia 1995, 33, 1419–1432. [Google Scholar] [CrossRef]
- Munzert, J.; Lorey, B.; Zentgraf, K. Cognitive motor processes: The role of motor imagery in the study of motor representations. Brain Res. Rev. 2009, 60, 306–326. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Jones, P.S.; Carpenter, T.; Baron, J.C. Mapping the involvement of BA 4a and 4p during motor imagery. Neuroimage 2008, 41, 92–99. [Google Scholar] [CrossRef]
- Solodkin, A.; Hlustik, P.; Chen, E.E.; Small, S.L. Fine modulation in network activation during motor execution and motor imagery. Cerebral Cortex 2004, 14, 1246–1255. [Google Scholar] [CrossRef]
- Daly, D.; Pedley, T. Current Practice of Clinical Electroencephalography; Raven Press: New York, NY, USA, 1990. [Google Scholar]
- Zschocke, S.; Speckmann, E.J. Basic Mechanisms of the EEG; Birkhäuser: Boston, MA, USA, 1993. [Google Scholar]
- Buzsaki, G.; Draguhn, A. Neuronal oscillations in cortical networks. Science 2004, 304, 1926–1929. [Google Scholar] [CrossRef] [Green Version]
- Jorge, J.; Van der Zwaag, W.; Figueiredo, P. EEG-fMRI integration for the study of human brain function. Neuroimage 2014, 102, 24–34. [Google Scholar] [CrossRef]
- Wackermann, J.; Lehmann, D.; Dvorak, I.; Michel, C.M. Global dimensional complexity of multi-channel EEG indicates change of human brain functional state after a single dose of a nootropic drug. Electroencephal. Clin. Neurophysiol. 1993, 86, 193–198. [Google Scholar] [CrossRef]
- Pavlov, A.; Grishina, D.; Runnova, A.; Maksimenko, V.; Pavlova, O.; Shchukovsky, N.; Hramov, A.; Kurths, J. Recognition of electroencephalographic patterns related to human movements or mental intentions with multiresolution analysis. Chaos Solitons Fractals 2019, 126, 230–235. [Google Scholar] [CrossRef]
- Pisarchik, A.N.; Maksimenko, V.A.; Hramov, A.E. From novel technology to novel applications: Comment on “An integrated brain-machine interface platform with thousands of channels” by Elon Musk and Neuralink. J. Med. Internet Res. 2019, 21, e16356. [Google Scholar] [CrossRef] [PubMed]
- Maksimenko, V.A.; Pavlov, A.N.; Runnova, A.E.; Nedaivozov, V.; Grubov, V.V.; Koronovskii, A.A.; Pchelintseva, S.V.; Pitsik, E.; Pisarchik, A.; Hramov, A. Nonlinear analysis of brain activity, associated with motor action and motor imaginary in untrained subjects. Nonlinear Dyn. 2018, 91, 2803–2817. [Google Scholar] [CrossRef]
- Aliakbaryhosseinabadi, S.; Kamavuako, E.N.; Jiang, N.; Farina, D.; Mrachacz-Kersting, N. Classification of EEG signals to identify variations in attention during motor task execution. J. Neurosci. Methods 2017, 284, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitsik, E.; Frolov, N.; Hauke Kraemer, K.; Grubov, V.; Maksimenko, V.; Kurths, J.; Hramov, A. Motor execution reduces EEG signals complexity: Recurrence quantification analysis study. Chaos 2020, 30, 023111. [Google Scholar] [CrossRef] [PubMed]
- Pfurtscheller, G.; Da Silva, F.L. Event-related EEG/MEG synchronization and desynchronization: Basic principles. Clin. Neurophysiol. 1999, 110, 1842–1857. [Google Scholar] [CrossRef]
- Ayaz, H.; Onaral, B.; Izzetoglu, K.; Shewokis, P.A.; McKendrick, R.; Parasuraman, R. Continuous monitoring of brain dynamics with functional near infrared spectroscopy as a tool for neuroergonomic research: Empirical examples and a technological development. Front. Hum. Neurosci. 2013, 7, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villringer, A.; Chance, B. Non-invasive optical spectroscopy and imaging of human brain function. Trends Neurosci. 1997, 20, 435–442. [Google Scholar] [CrossRef]
- Cui, X.; Bray, S.; Bryant, D.M.; Glover, G.H.; Reiss, A.L. A quantitative comparison of NIRS and fMRI across multiple cognitive tasks. Neuroimage 2011, 54, 2808–2821. [Google Scholar] [CrossRef] [Green Version]
- Leff, D.R.; Orihuela-Espina, F.; Elwell, C.E.; Athanasiou, T.; Delpy, D.T.; Darzi, A.W.; Yang, G.Z. Assessment of the cerebral cortex during motor task behaviours in adults: A systematic review of functional near infrared spectroscopy (fNIRS) studies. Neuroimage 2011, 54, 2922–2936. [Google Scholar] [CrossRef]
- Yamada, T.; Meng, E. Practical Guide for Clinical Neurophysiologic Testing: EEG; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2012. [Google Scholar]
- Ayaz, H.; Shewokis, P.A.; Curtin, A.; Izzetoglu, M.; Izzetoglu, K.; Onaral, B. Using MazeSuite and functional near infrared spectroscopy to study learning in spatial navigation. J. Vis. Exp. 2011, 56, e3443. [Google Scholar] [CrossRef] [Green Version]
- Cope, M. The Development of a Near Infrared Spectroscopy System and Its Application for Non Invasive Monitoring of Cerebral Blood and Tissue Oxygenation in the Newborn Infants. Ph.D. Thesis, University of London, London, UK, 1991. [Google Scholar]
- Naseer, N.; Hong, K.S. fNIRS-based brain-computer interfaces: A review. Front. Hum. Neurosci. 2015, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frolov, N.S.; Maksimenko, V.A.; Khramova, M.V.; Pisarchik, A.N.; Hramov, A.E. Dynamics of functional connectivity in multilayer cortical brain network during sensory information processing. Eur. Phys. J. Spec. Top. 2019, 228, 2381–2389. [Google Scholar] [CrossRef]
- Maksimenko, V.A.; Lüttjohann, A.; Makarov, V.V.; Goremyko, M.V.; Koronovskii, A.A.; Nedaivozov, V.; Runnova, A.E.; van Luijtelaar, G.; Hramov, A.E.; Boccaletti, S. Macroscopic and microscopic spectral properties of brain networks during local and global synchronization. Phys. Rev. E 2017, 96, 012316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, W.; Andersen, P.M.; Georgopoulos, A.P.; Kim, S.G. Sequential activity in human motor areas during a delayed cued finger movement task studied by time-resolved fMRI. Neuroreport 1997, 8, 1257–1261. [Google Scholar] [CrossRef] [PubMed]
- Koller, D.; Friedman, N.; Džeroski, S.; Sutton, C.; McCallum, A.; Pfeffer, A.; Abbeel, P.; Wong, M.F.; Heckerman, D.; Meek, C.; et al. Introduction to Statistical Relational Learning; MIT Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Ovchinnikov, A.; Lüttjohann, A.; Hramov, A.; Van Luijtelaar, G. An algorithm for real-time detection of spike-wave discharges in rodents. J. Neurosci. Methods 2010, 194, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Grubov, V.; Sitnikova, E.; Pavlov, A.; Koronovskii, A.; Hramov, A. Recognizing of stereotypic patterns in epileptic EEG using empirical modes and wavelets. Phys. A 2017, 486, 206–217. [Google Scholar] [CrossRef]
- Hajian-Tilaki, K. Receiver operating characteristic (ROC) curve analysis for medical diagnostic test evaluation. Casp. J. Intern. Med. 2013, 4, 627. [Google Scholar]
- Yi, W.; Qiu, S.; Qi, H.; Zhang, L.; Wan, B.; Ming, D. EEG feature comparison and classification of simple and compound limb motor imagery. J. Neuroeng. Rehabil. 2013, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Pavlov, A.; Runnova, A.; Maksimenko, V.; Pavlova, O.; Grishina, D.; Hramov, A. Detrended fluctuation analysis of EEG patterns associated with real and imaginary arm movements. Phys. A 2018, 509, 777–782. [Google Scholar] [CrossRef]
- Rahma, O.; Hendradi, R.; Fadli, A. Classifying imaginary hand movement through electroencephalograph signal for neuro-rehabilitation. Walailak J. Sci. Technol. 2019, 16, 943–953. [Google Scholar]
- Rosso, O.A.; Montani, F.; Baravalle, R.; Guisande, N.; Granado, M. Characterization of visuomotor/imaginary movements in EEG: An information theory and complex network approach. Front. Phys. 2019, 7, 115. [Google Scholar]
- Hanakawa, T. Organizing motor imageries. Neurosci. Res. 2016, 104, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, N.; Pomeroy, V.M.; Baron, J.C. Motor imagery: A backdoor to the motor system after stroke? Stroke 2006, 37, 1941–1952. [Google Scholar] [CrossRef] [Green Version]
- Park, C.h.; Chang, W.H.; Lee, M.; Kwon, G.H.; Kim, L.; Kim, S.T.; Kim, Y.H. Which motor cortical region best predicts imagined movement? Neuroimage 2015, 113, 101–110. [Google Scholar] [CrossRef]
- Duque, J.; Labruna, L.; Verset, S.; Olivier, E.; Ivry, R.B. Dissociating the role of prefrontal and premotor cortices in controlling inhibitory mechanisms during motor preparation. J. Neurosci. 2012, 32, 806–816. [Google Scholar] [CrossRef] [PubMed]
- Krams, M.; Rushworth, M.; Deiber, M.P.; Frackowiak, R.; Passingham, R. The preparation, execution and suppression of copied movements in the human brain. Exp. Brain Res. 1998, 120, 386–398. [Google Scholar] [CrossRef]
- Hwang, H.J.; Kwon, K.; Im, C.H. Neurofeedback-based motor imagery training for brain–computer interface (BCI). J. Neurosci. Methods 2009, 179, 150–156. [Google Scholar] [CrossRef]
- Kevric, J.; Subasi, A. Comparison of signal decomposition methods in classification of EEG signals for motor-imagery BCI system. Biomed. Signal Process. Control 2017, 31, 398–406. [Google Scholar] [CrossRef]
- Mason, S.G.; Birch, G.E. A brain-controlled switch for asynchronous control applications. IEEE Trans. Biomed. Eng. 2000, 47, 1297–1307. [Google Scholar] [CrossRef] [Green Version]
- Yong, X.; Menon, C. EEG classification of different imaginary movements within the same limb. PLoS ONE 2015, 10, e0121896. [Google Scholar] [CrossRef]
- Grozea, C.; Voinescu, C.D.; Fazli, S. Bristle-sensors—Low-cost flexible passive dry EEG electrodes for neurofeedback and BCI applications. J. Neural Eng. 2011, 8, 025008. [Google Scholar] [CrossRef] [PubMed]
- Spüler, M. A high-speed brain-computer interface (BCI) using dry EEG electrodes. PLoS ONE 2017, 12, e0172400. [Google Scholar] [CrossRef] [PubMed]
- Di Flumeri, G.; Aricò, P.; Borghini, G.; Sciaraffa, N.; Di Florio, A.; Babiloni, F. The dry revolution: Evaluation of three different EEG dry electrode types in terms of signal spectral features, mental states classification and usability. Sensors 2019, 19, 1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, D.; Coyle, D.; Wilson, S.; Callaghan, M. Games, gameplay, and BCI: The state of the art. IEEE Trans. Comput. Intell. AI Games 2013, 5, 82–99. [Google Scholar] [CrossRef]
- Coyle, D.; Garcia, J.; Satti, A.R.; McGinnity, T.M. EEG-Based Continuous Control of a Game Using a 3 Channel Motor Imagery BCI: BCI Game. In Proceedings of the 2011 IEEE Symposium on Computational Intelligence, Cognitive Algorithms, Mind, and Brain (CCMB), Paris, France, 15 April 2011; pp. 1–7. [Google Scholar]
- Al-Qaysi, Z.; Zaidan, B.; Zaidan, A.; Suzani, M. A review of disability EEG based wheelchair control system: Coherent taxonomy, open challenges and recommendations. Comput. Methods Programs Biomed. 2018, 164, 221–237. [Google Scholar] [CrossRef]
- Malki, A.; Yang, C.; Wang, N.; Li, Z. Mind Guided MotionControl of Robot Manipulator Using EEG Signals. In Proceedings of the 5th International Conference on Information Science and Technology (ICIST), Changsha, China, 24–26 April 2015; pp. 553–558. [Google Scholar]
- Huang, D.; Qian, K.; Fei, D.Y.; Jia, W.; Chen, X.; Bai, O. Electroencephalography (EEG)-based brain-computer interface (BCI): A 2-D virtual wheelchair control based on event-related desynchronization/synchronization and state control. IEEE Trans. Neural Syst. Rehabil. Eng. 2012, 20, 379–388. [Google Scholar] [CrossRef]
- Putze, F.; Hesslinger, S.; Tse, C.Y.; Huang, Y.; Herff, C.; Guan, C.; Schultz, T. Hybrid fNIRS-EEG based classification of auditory and visual perception processes. Front. Neurosci. 2014, 8, 373. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.J.; Hong, K.S. Hybrid EEG–fNIRS-based eight-command decoding for BCI: Application to quadcopter control. Front. Neurorobotics 2017, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Hong, K.S.; Khan, M.J.; Hong, M.J. Feature extraction and classification methods for hybrid fNIRS-EEG brain-computer interfaces. Front. Hum. Neurosci. 2018, 12, 246. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Movement | Automatic Detection | , % | , % | , % | |||
|---|---|---|---|---|---|---|---|
| Real | |||||||
| Imaginary | |||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hramov, A.E.; Grubov, V.; Badarin, A.; Maksimenko, V.A.; Pisarchik, A.N. Functional Near-Infrared Spectroscopy for the Classification of Motor-Related Brain Activity on the Sensor-Level. Sensors 2020, 20, 2362. https://doi.org/10.3390/s20082362
Hramov AE, Grubov V, Badarin A, Maksimenko VA, Pisarchik AN. Functional Near-Infrared Spectroscopy for the Classification of Motor-Related Brain Activity on the Sensor-Level. Sensors. 2020; 20(8):2362. https://doi.org/10.3390/s20082362
Chicago/Turabian StyleHramov, Alexander E., Vadim Grubov, Artem Badarin, Vladimir A. Maksimenko, and Alexander N. Pisarchik. 2020. "Functional Near-Infrared Spectroscopy for the Classification of Motor-Related Brain Activity on the Sensor-Level" Sensors 20, no. 8: 2362. https://doi.org/10.3390/s20082362