
Using Drones to Determine Chimpanzee Absences at the Edge of Their Distribution in Western Tanzania

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
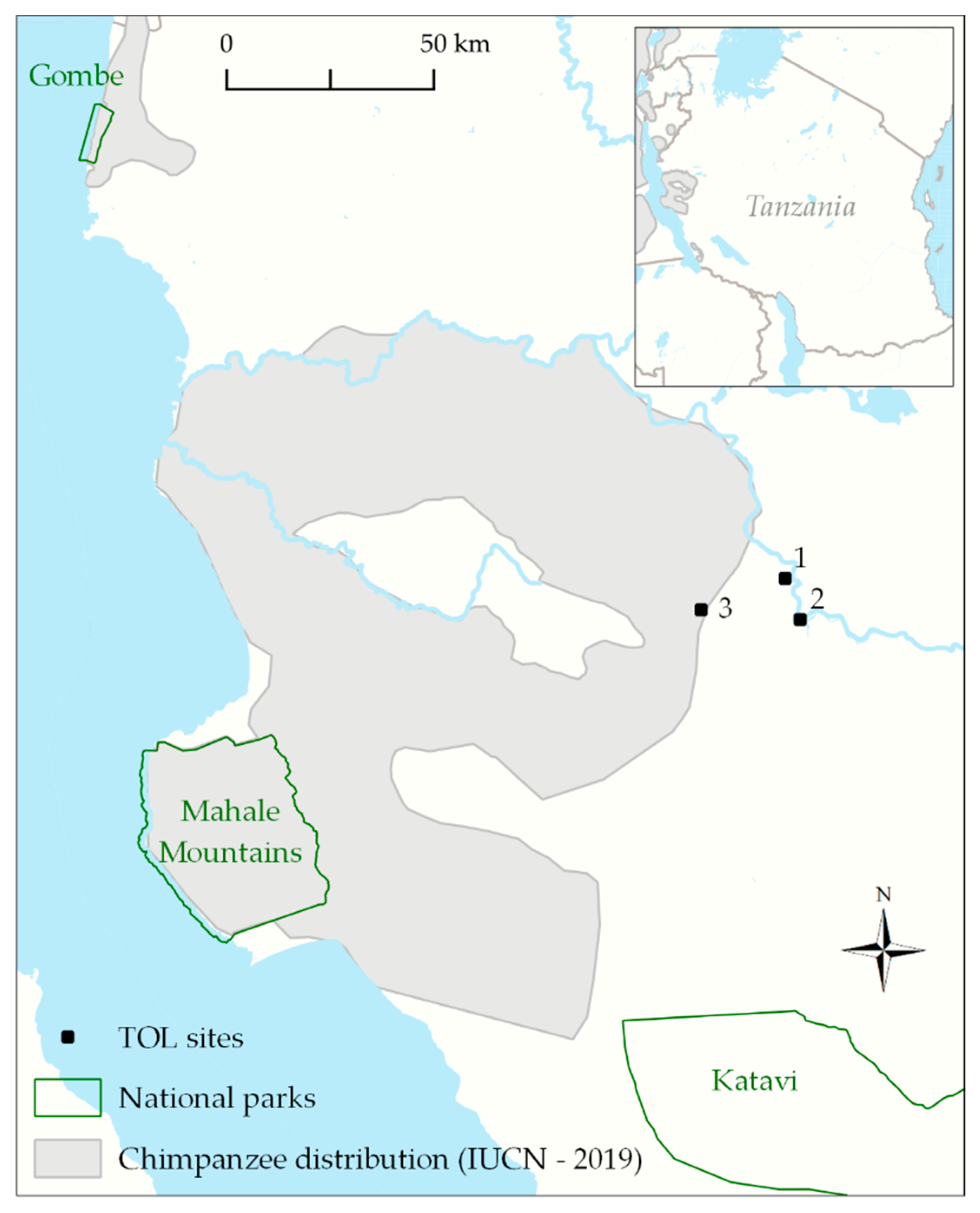
2. Methods
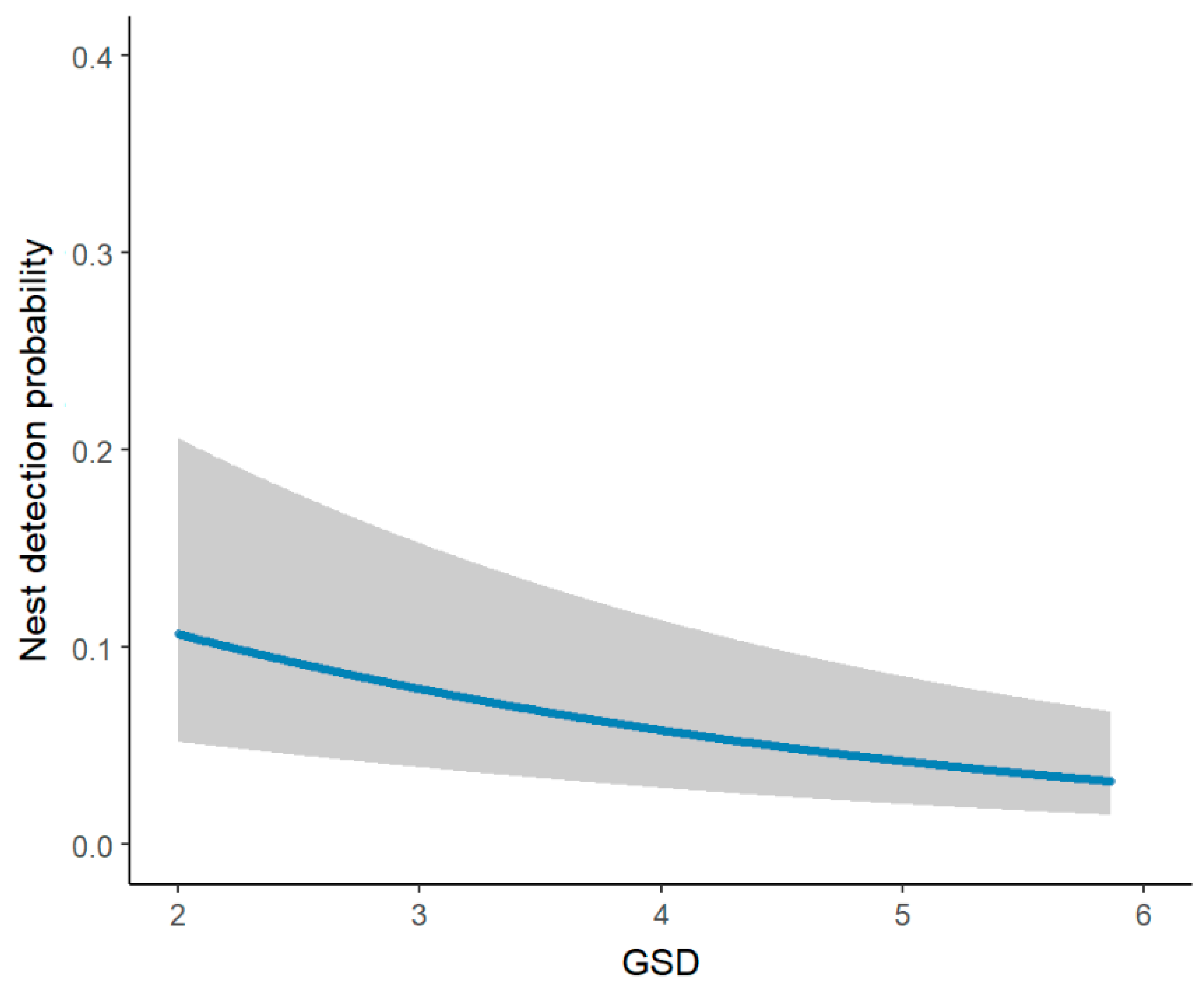
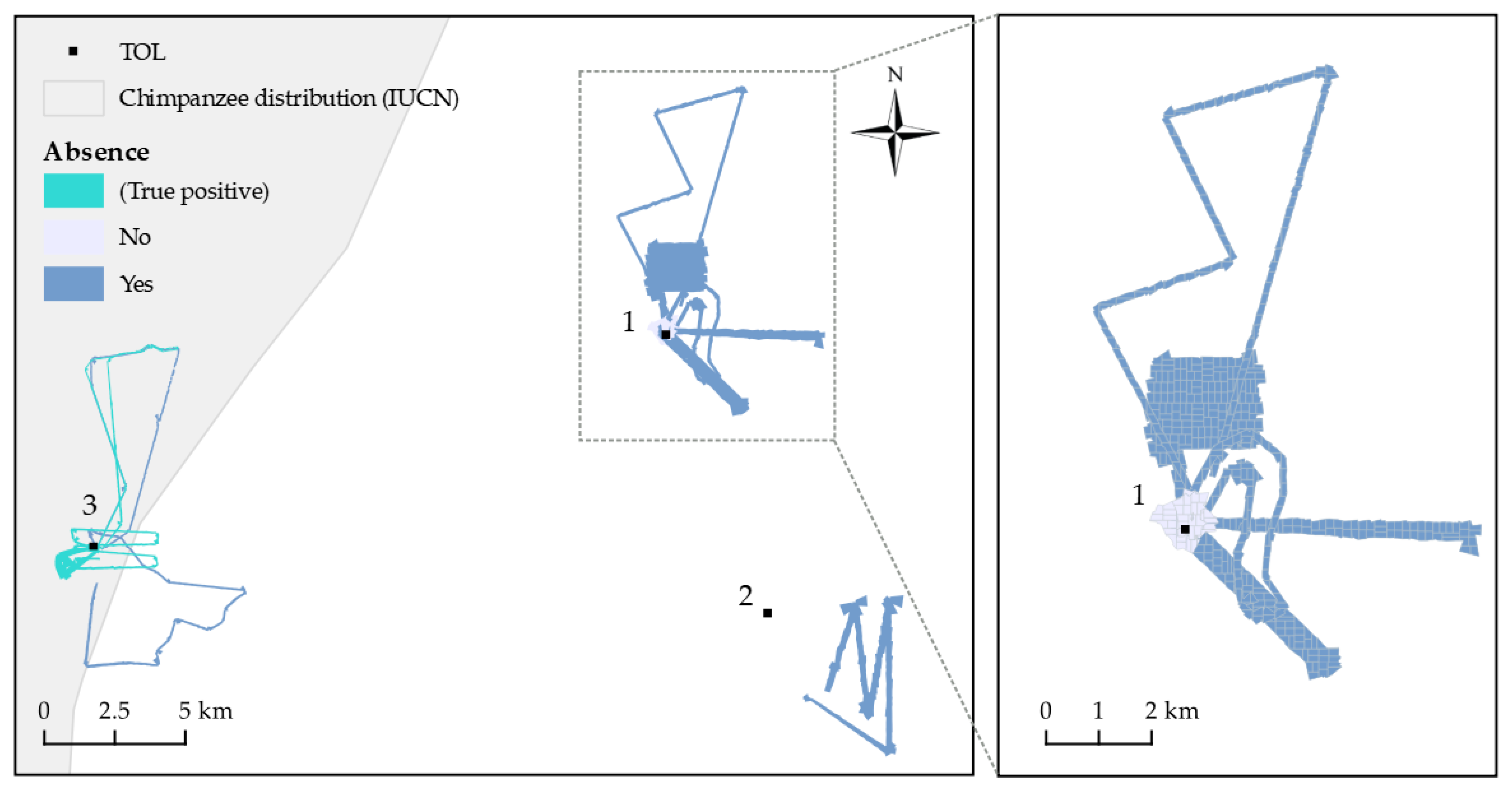
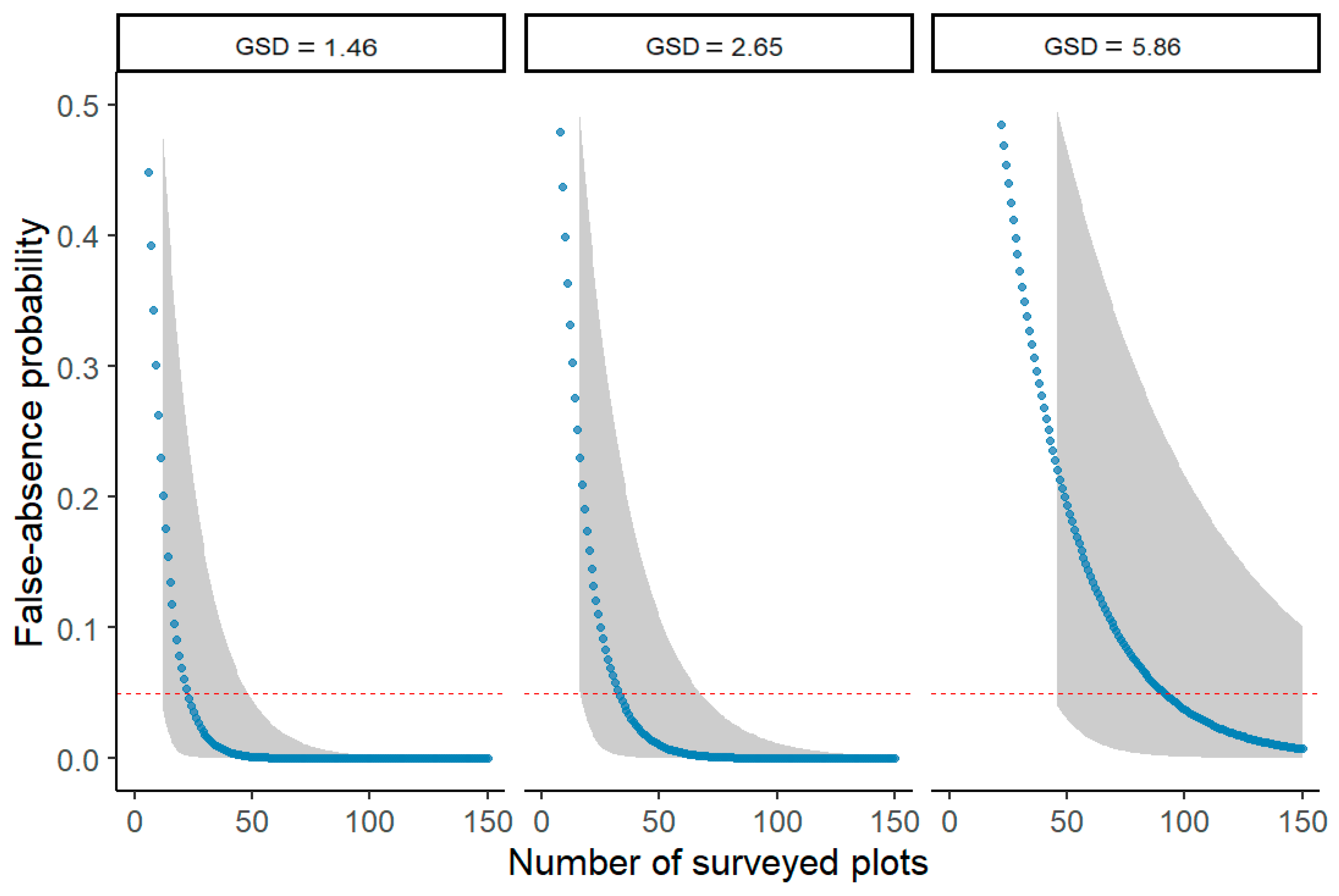
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ancrenaz, M.; Gimenez, O.; Ambu, L.; Ancrenaz, K.; Andau, P.; Goossens, B.; Payne, J.; Sawang, A.; Tuuga, A.; Lackman-Ancrenaz, I. Aerial Surveys Give New Estimates for Orangutans in Sabah, Malaysia. PLOS Biol. 2005, 3, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Boesch, C.; Hohmann, G.; Marchant, L.F. (Eds.) Behavioural Diversity in Chimpanzees and Bonobos; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Bonnin, N.; Stewart, F.A.; Wich, S.A.; Pintea, L.; Jantz, S.M.; Dickson, R.; Bellis, J.; Chitayat, A.; Ingram, R.; Moore, R.J.; et al. Modelling landscape connectivity change for chimpanzee conservation in Tanzania. Biol. Conserv. 2020, 252, 108816. [Google Scholar] [CrossRef]
- Bonnin, N.; Van Andel, A.C.; Kerby, J.T.; Piel, A.K.; Pintea, L.; Wich, S.A. Assessment of Chimpanzee Nest Detectability in Drone-Acquired Images. Drones 2018, 2, 17. [Google Scholar] [CrossRef] [Green Version]
- Brack, I.V.; Kindel, A.; Oliveira, L.F.B. Detection errors in wildlife abundance estimates from Unmanned Aerial Systems (UAS) surveys: Synthesis, solutions, and challenges. Methods Ecol. Evol. 2018, 9, 1864–1873. [Google Scholar] [CrossRef]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Advanced Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Buckland, S.; Plumptre, A.J.; Thomas, L.; Rexstad, E.A. Design and Analysis of Line Transect Surveys for Primates. Int. J. Primatol. 2010, 31, 833–847. [Google Scholar] [CrossRef] [Green Version]
- Burton, A.C.; Neilson, E.; Moreira, D.; Ladle, A.; Steenweg, R.; Fisher, J.T.; Bayne, E.; Boutin, S. REVIEW: Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes. J. Appl. Ecol. 2015, 52, 675–685. [Google Scholar] [CrossRef]
- Campos Cerqueira, M.; Aide, M.T. Improving distribution data of threatened species by combining acoustic monitoring and occupancy modeling. Methods Ecol. Evol. 2016, 7, 1340–1348. [Google Scholar] [CrossRef]
- Carvalho, J.S.; Stewart, F.A.; Marques, T.A.; Bonnin, N.; Pintea, L.; Chitayat, A.; Ingram, R.; Moore, R.J.; Piel, A.K. Spatio-temporal changes in chimpanzee density and abundance in the Greater Mahale Ecosystem, Tanzania. Ecol. Appl. 2022, 32, e2715. [Google Scholar] [CrossRef]
- Chabot, D.; Bird, D.M. Wildlife research and management methods in the 21st century: Where do unmanned aircraft fit in? J. Unmanned Veh. Syst. 2015, 3, 137–155. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Shioi, H.; Montesinos, C.F.; Koh, L.P.; Wich, S.; Krause, A. Active detection via adaptive submodularity. In Proceedings of the 31st International Conference on Machine Learning, Beijing, China, 21–26 June 2014. [Google Scholar]
- Chitayat, A.B.; Wich, S.A.; Lewis, M.; Stewart, F.A.; Piel, A.K. Ecological correlates of chimpanzee (Pan troglodytes schweinfurthii) density in Mahale Mountains National Park, Tanzania. PLoS ONE 2021, 16, e0246628. [Google Scholar]
- Conn, P.B.; Hoef, J.M.V.; McClintock, B.T.; Moreland, E.E.; London, J.M.; Cameron, M.F.; Dahle, S.P.; Boveng, P.L. Estimating multispecies abundance using automated detection systems: Ice-associated seals in the Bering Sea. Methods Ecol. Evol. 2014, 5, 1280–1293. [Google Scholar] [CrossRef]
- Coro, G.; Magliozzi, C.; Berghe, E.V.; Bailly, N.; Ellenbroek, A.; Pagano, P. Estimating absence locations of marine species from data of scientific surveys in OBIS. Ecol. Model. 2016, 323, 61–76. [Google Scholar] [CrossRef]
- Crunchant, A.S.; Borchers, D.; Kühl, H.; Piel, A. Listening and watching: Do camera traps or acoustic sensors more efficiently detect wild chimpanzees in an open habitat? Methods Ecol. Evol. 2020, 11, 542–552. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Fiske, I.; Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Fristrup, K.M.; Clark, C.W. Acoustic Monitoring of Threatened and Endangered Species in Inaccessible Areas, DTIC Document; Cornell University: Ithaca, NY, USA, 2009. [Google Scholar]
- Griffiths, M.; Van Schaik, C.P. Camera trapping: A new tool for the study of elusive rain forest mammals. Trop. Biodivers. 1993, 1, 131–135. [Google Scholar]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Jantz, S.M.; Pintea, L.; Nackoney, J.; Hansen, M.C. Landsat ETM+ and SRTM Data Provide Near Real-Time Monitoring of Chimpanzee (Pan troglodytes) Habitats in Africa. Remote Sens. 2016, 8, 427. [Google Scholar] [CrossRef] [Green Version]
- Joseph, L.N.; Elkin, C.; Martin, T.G.; Possingham, H.P. Modeling abundance using N-mixture models: The importance of considering ecological mechanisms. Ecol. Appl. 2009, 19, 631–642. [Google Scholar] [CrossRef] [Green Version]
- Kalan, A.K.; Piel, A.K.; Mundry, R.; Wittig, R.M.; Boesch, C.; Kühl, H.S. Passive acoustic monitoring reveals group ranging and territory use: A case study of wild chimpanzees (Pan troglodytes). Front. Zoöl. 2016, 13, 34. [Google Scholar] [CrossRef] [Green Version]
- Kéry, M. Inferring the Absence of a Species: A Case Study of Snakes. J. Wildl. Manag. 2002, 66, 330. [Google Scholar] [CrossRef]
- Kéry, M.; Royle, J.A. Chapter 10—Modeling Static Occurrence and Species Distributions Using Site-occupancy Models. In Applied Hierarchical Modeling in Ecology; Kéry, M., Royle, J.A., Eds.; Academic Press: Boston, MA, USA, 2016; pp. 551–629. [Google Scholar]
- Kuhl, H.; Maisels, F.; Ancrenaz, M.; Williamson, E. Best Practice Guidelines for Surveys and Monitoring of Great Ape Populations; Linemark Printing: Large, MD, USA, 2008. [Google Scholar] [CrossRef] [Green Version]
- Lahoz-Monfort, J.J.; Guillera-Arroita, G.; Wintle, B.A. Imperfect detection impacts the performance of species distribution models. Glob. Ecol. Biogeogr. 2014, 23, 504–515. [Google Scholar] [CrossRef]
- Linchant, J.; Lhoest, S.; Quevauvillers, S.; Semeki, J.; Lejeune, P.; Vermeulen, C. Wimuas: Developing a Tool to Review Wildlife Data from Various Uas Flight Plans. ISPRS—Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2015, XL-3/W3, 379–384. [Google Scholar] [CrossRef] [Green Version]
- MacKenzie, D.I. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- McArdle, B.H. When are rare species not there? Oikos 1990, 57, 276–277. [Google Scholar] [CrossRef]
- Milne, S.; Martin, J.; Reynolds, G.; Vairappan, C.; Slade, E.; Brodie, J.; Wich, S.; Williamson, N.; Burslem, D. Drivers of Bornean Orangutan Distribution across a Multiple-Use Tropical Landscape. Remote Sens. 2021, 13, 458. [Google Scholar] [CrossRef]
- Moyer, D.; Plumptre, A.J.; Pintea, L.; Hernandez-Aguilar, A.; Moore, J.; Stewart, F.; Mwangoka, M. Surveys of Chimpanzees and other Biodiversity in Western Tanzania. In United States Fish and Wildlife Service; University of California: San Diego, CA, USA, 2006. [Google Scholar]
- Pellet, J.; Schmidt, B.R. Monitoring distributions using call surveys: Estimating site occupancy, detection probabilities and inferring absence. Biol. Conserv. 2005, 123, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Piel, A.K.; Cohen, S.N.; Kamenya, S.; Ndimuligo, S.A.; Pintea, L.; Stewart, F.A. Population status of chimpanzees in the Masito-Ugalla Ecosystem, Tanzania. Am. J. Primatol. 2015, 77, 1027–1035. [Google Scholar] [CrossRef]
- Piel, A.K.; Stewart, F. Census and Conservation Status of Chimpanzees (Pan troglodytes schweinfurthii) across the Greater Mahale Ecosystem; The Nature Conservancy: Arlington, VA, USA, 2014. [Google Scholar]
- Plumptre, A.J.; Reynolds, V. Censusing chimpanzees in the Budongo Forest, Uganda. Int. J. Primatol. 1996, 17, 85–99. [Google Scholar] [CrossRef]
- Seymour, A.C.; Dale, J.; Hammill, M.; Halpin, P.N.; Johnston, D.W. Automated detection and enumeration of marine wildlife using unmanned aircraft systems (UAS) and thermal imagery. Sci. Rep. 2017, 7, 45127. [Google Scholar] [CrossRef] [Green Version]
- van Andel, A.C.; Wich, S.A.; Boesch, C.; Koh, L.P.; Robbins, M.M.; Kelly, J.; Kuehl, H.S. Locating chimpanzee nests and identifying fruiting trees with an unmanned aerial vehicle. Am. J. Primatol. 2015, 77, 1122–1134. [Google Scholar] [CrossRef]
- Van Gemert, J.C.; Verschoor, C.R.; Mettes, P.; Epema, K.; Koh, L.P.; Wich, S. Nature Conservation Drones for Automatic Localization and Counting of Animals. In Computer Vision—ECCV 2014 Workshops; Agapito, L., Bronstein, M., Rother, C., Eds.; Springer: New York, NY, USA, 2014; p. 8925. [Google Scholar]
- van Schaik, C.P.; Priatna, A.; Priatna, D. Population estimates and habitat preferences of orang-utans based on line transects of nests. In The Neglected Ape; Nadler, R.D., Galdikas, B.M.F., Sheeran, L.K., Rosen, N., Eds.; Plenum Press: New York, NY, USA, 1995. [Google Scholar]
- Voigt, M.; Wich, S.A.; Ancrenaz, M.; Meijaard, E.; Abram, N.; Banes, G.L.; Campbell-Smith, G.; d’Arcy, L.J.; Delgado, R.A.; Erman, A.; et al. Global Demand for Natural Resources Eliminated More Than 100,000 Bornean Orangutans. Curr. Biol. 2018, 28, 761–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wearn, O.R.; Glover-Kapfer, P. Snap happy: Camera traps are an effective sampling tool when compared with alternative methods. R. Soc. Open Sci. 2019, 6, 181748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wich, S.; Dellatore, D.; Houghton, M.; Ardi, R.; Koh, L.P. A preliminary assessment of using conservation drones for Sumatran orang-utan (Pongo abelii) distribution and density. J. Unmanned Veh. Syst. 2016, 4, 45–52. [Google Scholar] [CrossRef]
- Wich, S.; Fredriksson, G.; Usher, G.; Peters, H.; Priatna, D.; Basalamah, F.; Susanto, W.; Kühl, H. Hunting of Sumatran orang-utans and its importance in determining distribution and density. Biol. Conserv. 2012, 146, 163–169. [Google Scholar] [CrossRef] [Green Version]
- Wich, S.A.; Gaveau, D.; Abram, N.; Ancrenaz, M.; Baccini, A.; Brend, S.; Curran, L.; Delgado, R.A.; Erman, A.; Fredriksson, G.M. Understanding the Impacts of Land-Use Policies on a Threatened Species: Is There a Future for the Bornean Orangutan? PLoS ONE 2012, 7, e49142. [Google Scholar] [CrossRef] [Green Version]
- Wich, S.A.; Koh, L.P. Conservation Drones; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Wich, S.A.; Singleton, I.; Nowak, M.G.; Atmoko, S.S.U.; Nisam, G.; Arif, S.M.; Putra, R.H.; Ardi, R.; Fredriksson, G.; Usher, G.; et al. Land-cover changes predict steep declines for the Sumatran orangutan (Pongo abelii). Sci. Adv. 2016, 2, e1500789. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.J.; Hooten, M.B.; Womble, J.N.; Bower, M.R. Estimating occupancy and abundance using aerial images with imperfect detection. Methods Ecol. Evol. 2017, 8, 1679–1689. [Google Scholar] [CrossRef] [Green Version]
- Wisz, M.S.; Guisan, A. Do pseudo-absence selection strategies influence species distribution models and their predictions? An information-theoretic approach based on simulated data. BMC Ecol. 2009, 9, 8. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Model | AICc | ΔAICc | Detection p (SE) | Abundance λ (SE) | |
|---|---|---|---|---|---|---|
| 1 | λ (·) p (GSD) | 162.98 | 0.000 | 0.196 | (0.045) | 39.6 (15.1) |
| 2 | λ (·) p (·) | 198.25 | 35.27 | 0.031 | (0.009) | 46.7 (14.2) |
| Flight Number | Number of Photos | TOL | Alt AGL (m) | GSD (cm) | Mission Type | Area Covered (km2) | # of Plots | Plots with Number of Nests | Detection Probability | Probability of a False Negative | Can Real Absence Be Assumed? (95% Probability) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 194 | 1 | 115 | 2.2 | grid | 0.87 | 19 | 0 | 0.098 | 0.141 | no |
| 2 | 333 | 1 | 177 | 3.4 | grid | 2.06 | 61 | 0 | 0.07 | 0.012 | yes |
| 3 | 183 | 1 | 190 | 3.7 | transect | 1.74 | 60 | 0 | 0.065 | 0.017 | yes |
| 4 | 203 | 1 | 305 | 5.9 | grid | 4.35 | 140 | 0 | 0.032 | 0.011 | yes |
| 5 | 50 | 1 | 100 | 1.9 | transect | 0.4 | 15 | 0 | 0.109 | 0.177 | no |
| 6 | 344 | 1 | 153 | 2.9 | transect | 2.62 | 102 | 0 | 0.080 | <0.001 | yes |
| 7 | 230 | 1 | 138 | 2.7 | transect | 1.71 | 66 | 0 | 0.088 | 0.002 | yes |
| 8 | 341 | 2 | 192 | 3.7 | transect | 4.81 | 192 | 0 | 0.064 | <0.001 | yes |
| 9 | 384 | 3 | 108 | 2.1 | transect | 1.64 | 62 | 0 | 0.104 | 0.001 | yes |
| 10 | 395 | 3 | 83 | 1.6 | transect | 1.23 | 29 | 0 | 0.12 | 0.024 | yes |
| 11 | 484 | 3 | 76 | 1.5 | transect | 1.17 | 46 | 1, 1 | 0.125 | 0 | (true positive) |
| 12 | 330 | 3 | 102 | 2.0 | transect | 1.29 | 50 | 1 | 0.108 | 0 | (true positive) |
| 13 | 89 | 3 | 142 | 2.7 | transect | 0.95 | 29 | 4, 4, 3, 1, 1, 2 | 0.086 | 0 | (true positive) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wich, S.A.; Bonnin, N.; Hutschenreiter, A.; Piel, A.K.; Chitayat, A.; Stewart, F.A.; Pintea, L.; Kerby, J.T. Using Drones to Determine Chimpanzee Absences at the Edge of Their Distribution in Western Tanzania. Remote Sens. 2023, 15, 2019. https://doi.org/10.3390/rs15082019
Wich SA, Bonnin N, Hutschenreiter A, Piel AK, Chitayat A, Stewart FA, Pintea L, Kerby JT. Using Drones to Determine Chimpanzee Absences at the Edge of Their Distribution in Western Tanzania. Remote Sensing. 2023; 15(8):2019. https://doi.org/10.3390/rs15082019
Chicago/Turabian StyleWich, Serge A., Noémie Bonnin, Anja Hutschenreiter, Alex K. Piel, Adrienne Chitayat, Fiona A. Stewart, Lilian Pintea, and Jeffrey T. Kerby. 2023. "Using Drones to Determine Chimpanzee Absences at the Edge of Their Distribution in Western Tanzania" Remote Sensing 15, no. 8: 2019. https://doi.org/10.3390/rs15082019