Predicting Microhabitat Suitability for an Endangered Small Mammal Using Sentinel-2 Data

 , , , ,
, , , ,
Abstract
:
1. Introduction
- i.
- Quantify the importance of Sentinel-2 derived predictors relative to more conventional predictors (e.g., topographical and distance to landscape elements) in predicting vole microhabitat suitability;
- ii.
- Identify which Sentinel-2 derived predictors best explain vole distribution at fine spatial scales.
2. Materials and Methods
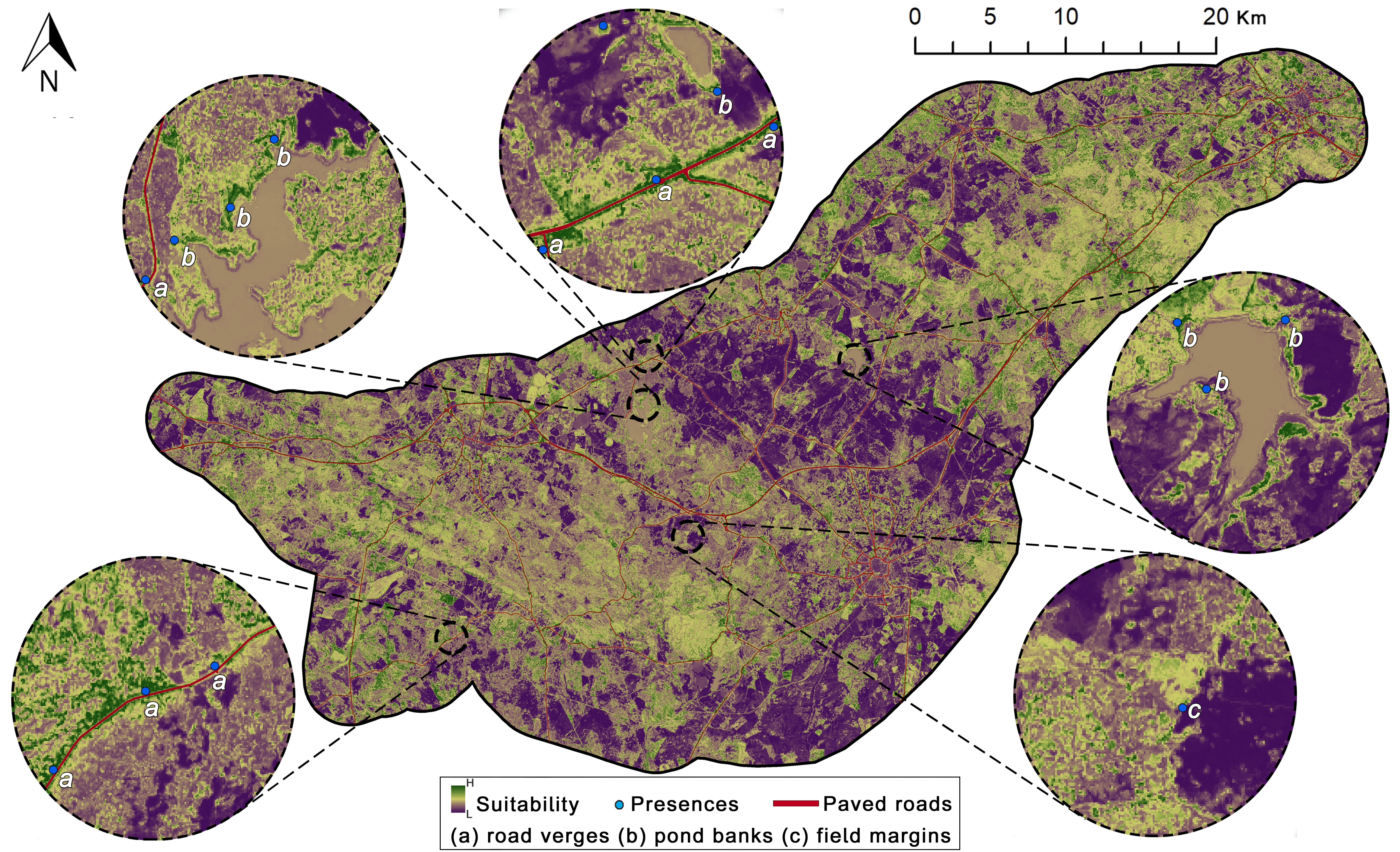
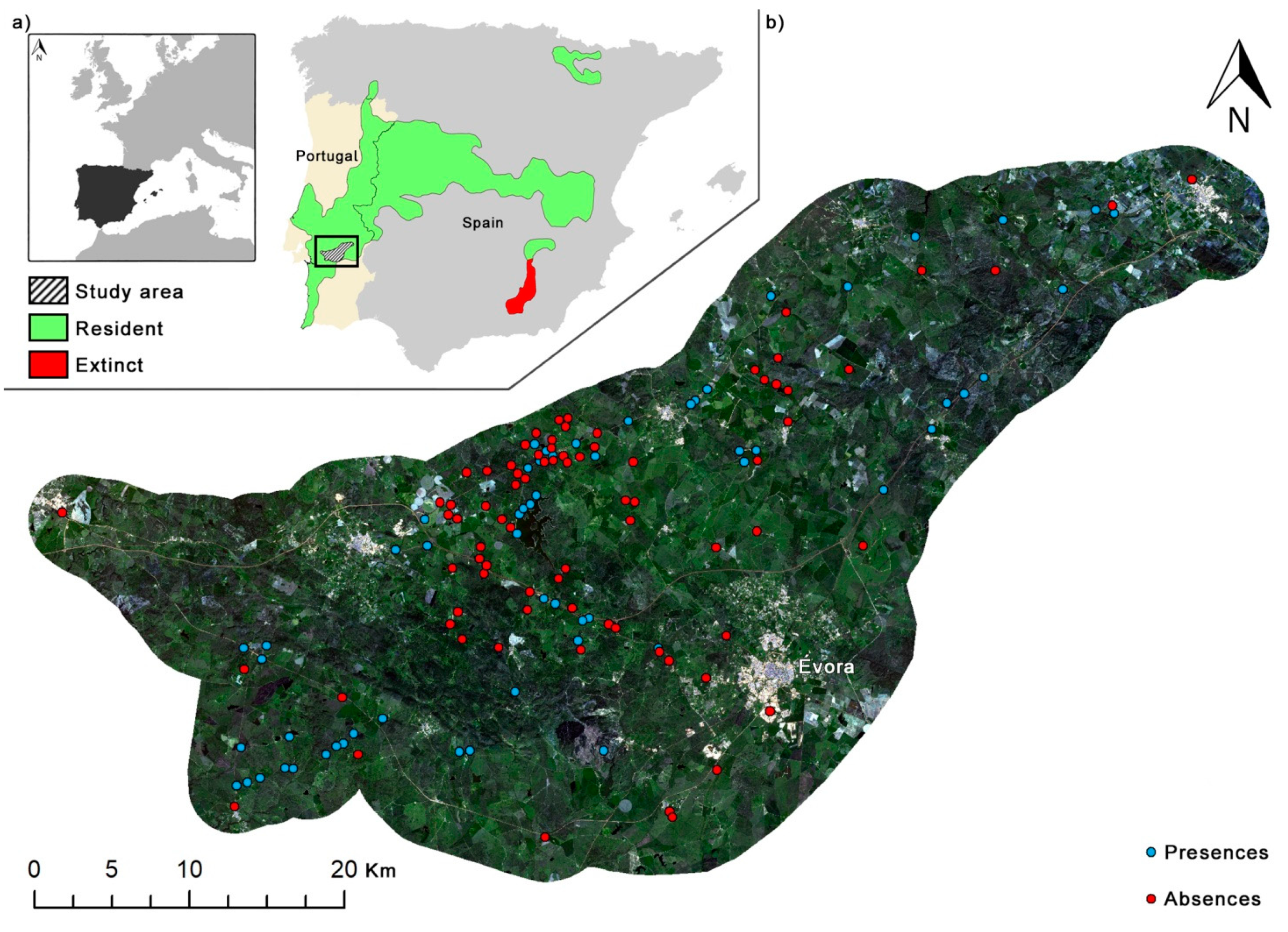
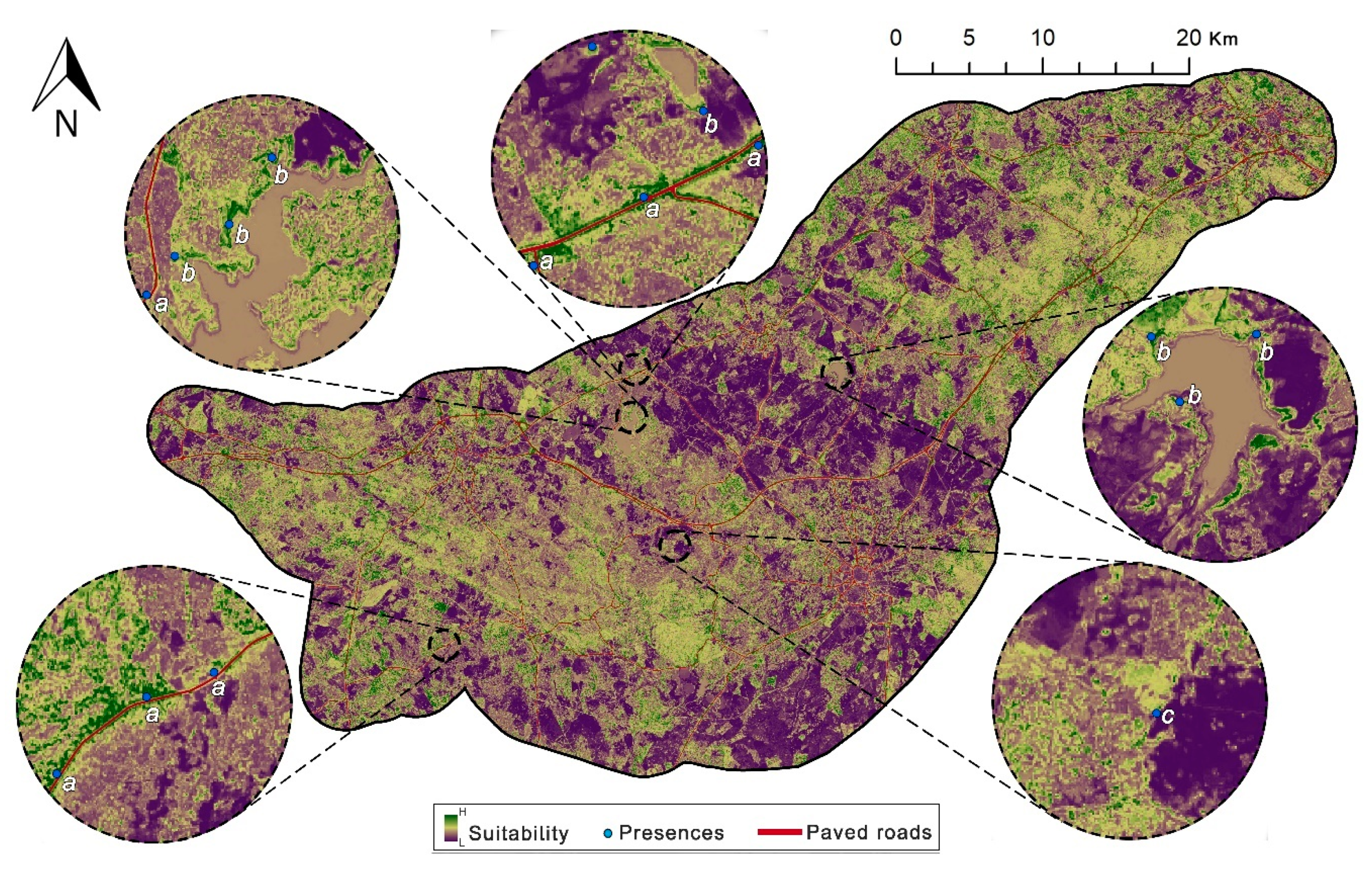
2.1. Study Area
2.2. Cabrera Vole Field Surveys
2.3. Predictor Variables
2.3.1. Sentinel-2 Derived Predictor Variables
2.3.2. Topographical Predictor Variables
2.3.3. Distance to Landscape Elements
2.4. Habitat Suitability Model
3. Results
3.1. Model Performance
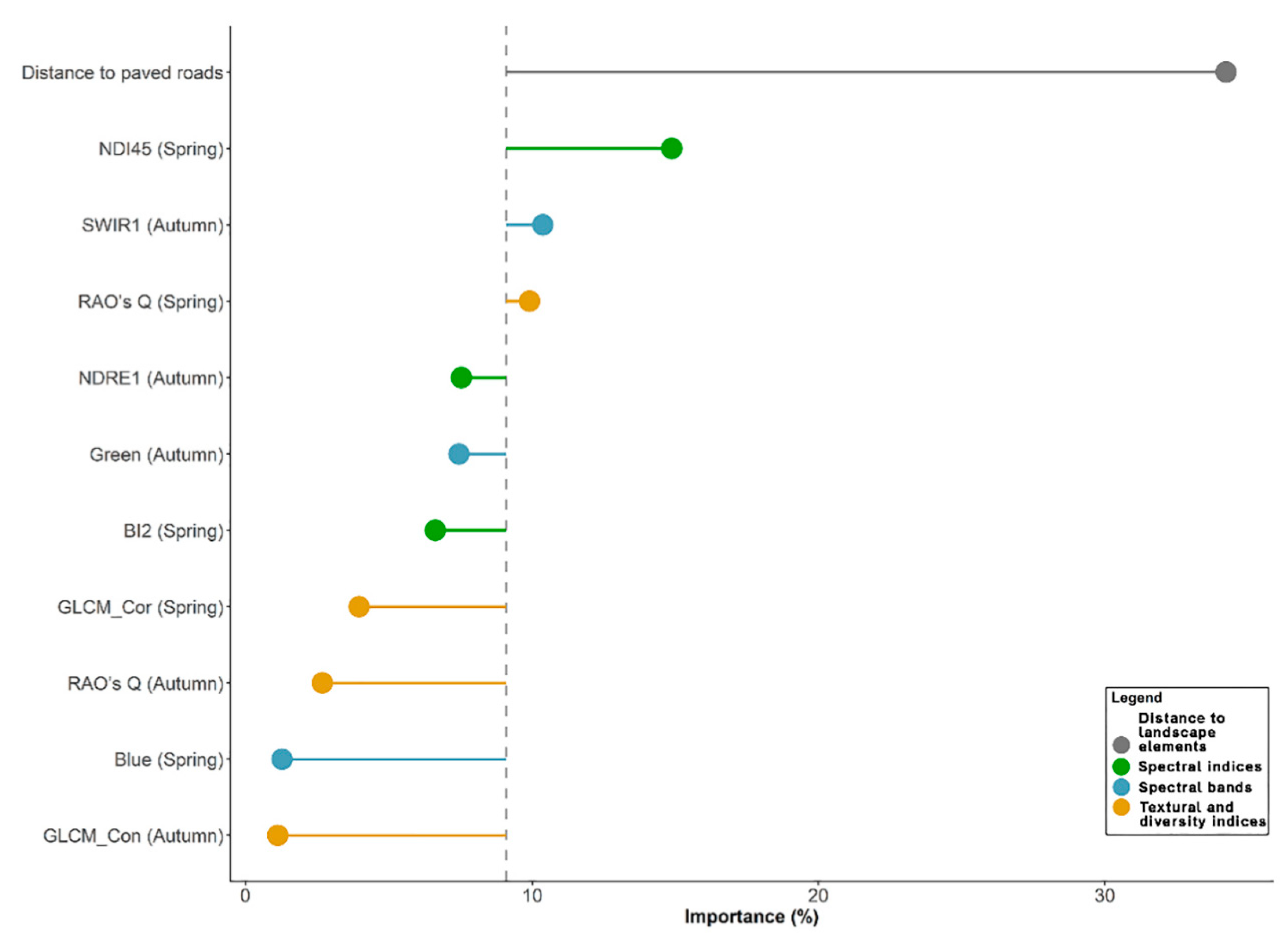
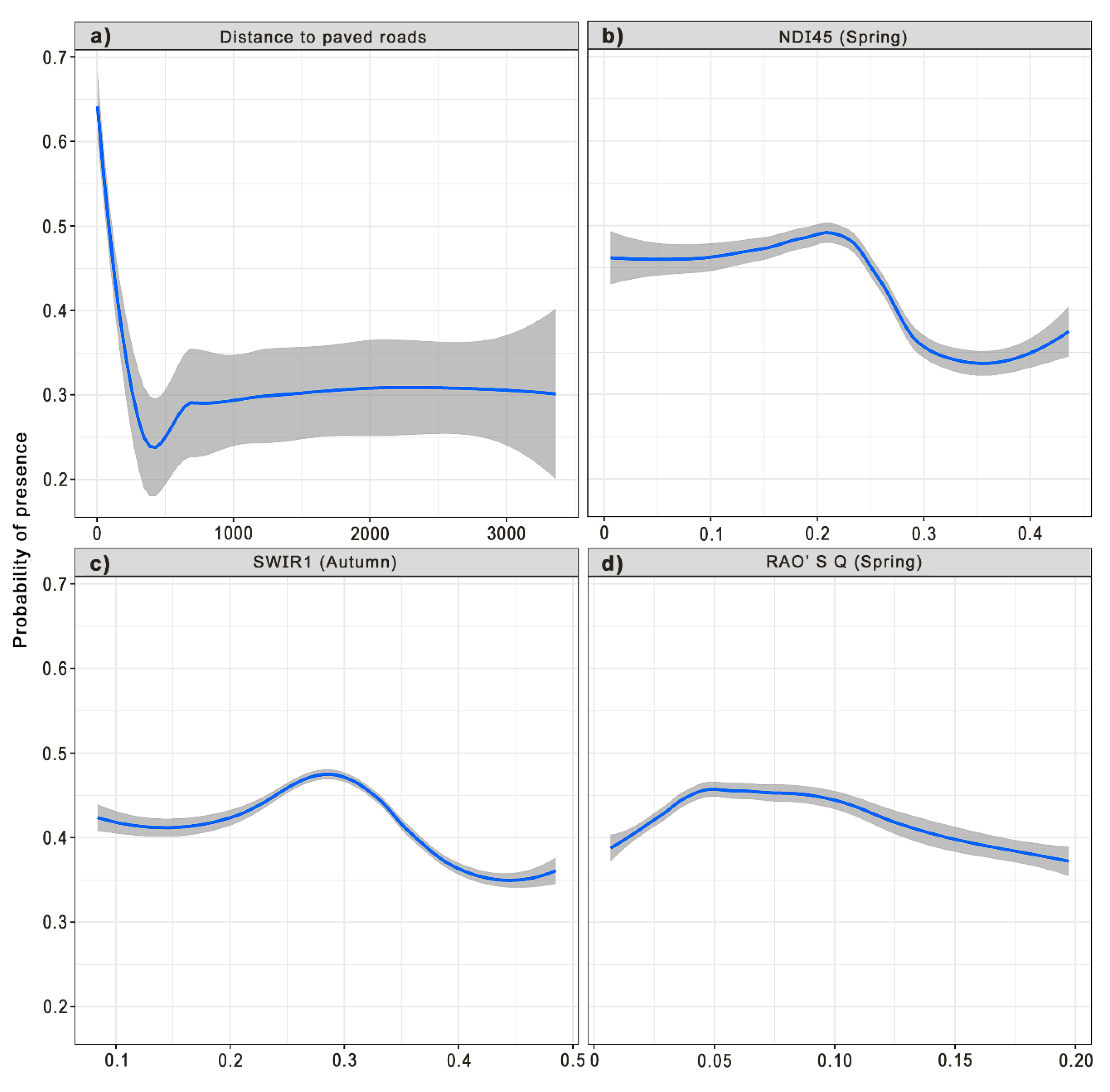
3.2. Variable Importance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human–induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehtonen, A.; Salonen, A.; Cantell, H.; Riuttanen, L.; MacMillan, P. IPBES: Nature’s Dangerous Decline ‘Unprecedented’; Species Extinction Rates ‘Accelerating’, Paris, France. Drama and Theatre Education. 26 May 2019. Available online: https://www.ipbes.net/news/Media-Release-Global-Assessment#_Indigenous_Peoples,_Local (accessed on 8 May 2019).
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otto, S.P. Adaptation, speciation and extinction in the Anthropocene. Proc. Biol. Sci. 2018, 285, 20182047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Hoffmann, M.; Wirsing, A.J.; McCauley, D.J. Extinction risk is most acute for the world’s largest and smallest vertebrates. Proc. Nat. Acad. Sci. USA 2017, 114, 10678–10683. [Google Scholar] [CrossRef] [Green Version]
- Brook, B.W.; Sodhi, N.S.; Bradshaw, C.J. Synergies among extinction drivers under global change. Trends Ecol. Evol. 2008, 23, 453–460. [Google Scholar] [CrossRef]
- Fernández-Salvador, R. Microtus cabrerae Thomas, 1906, Topillo de Cabrera. In Atlas y Libr Rojo los Mamíferos de España; Palomo, J.L., Gisbert, J., Blanco, J.C., Eds.; Ministerio de Medio Ambiente: Madrid, Spain, 2007; pp. 429–433. [Google Scholar]
- Queiroz, A.I.; Alves, P.C.; Barroso, I.; Beja, P.; Fernandes, M.; Freitas, L.; Mathias, M.L.; Mira, A.; Palmeirim, J.M.; Prieto, R.; et al. Microtus cabrerae Rato de Cabrera. In Livro Vermelho dos Vertebrados de Portugal; Cabral, M.J., Ed.; Instituto da Conservação da Natureza: Lisboa, Portugal, 2006; pp. 481–482. [Google Scholar]
- Fernandes, M.; Pita, R.; Mira, A. Microtus cabrerae. The IUCN Red List of Threatened Species 2019: E.T13418A90931498. Available online: https://www.iucnredlist.org/species/13418/90931498 (accessed on 18 January 2020).
- Mira, A.; Marques, C.C.; Santos, S.M.; Rosário, I.T.; Mathias, M.L. Environmental determinants of the distribution of the Cabrera vole (Microtus cabrerae) in Portugal: Implications for conservation. Mamm. Biol. 2008, 2, 102–110. [Google Scholar] [CrossRef]
- Pita, R.; Mira, A.; Beja, P. Conserving the Cabrera vole, Microtus cabrerae, in intensively used Mediterranean landscapes. Agric. Ecosyst. Environ. 2006, 115, 1–5. [Google Scholar] [CrossRef]
- Pita, R.; Beja, P.; Mira, A. Spatial population structure of the Cabrera vole in Mediterranean farmland: The relative role of patch and matrix effects. Biol. Conserv. 2007, 134, 383–392. [Google Scholar] [CrossRef]
- Santos, S.M.; Simões, M.P.; da Luz Mathias, M.; Mira, A. Vegetation analysis in colonies of an endangered rodent, the Cabrera vole (Microtus cabrerae), in southern Portugal. Ecol. Res. 2006, 2, 197–207. [Google Scholar] [CrossRef]
- Pita, R.; Mira, A.; Beja, P. Microtus cabrerae (Rodentia: Cricetidae) Mammalian. Species 2014, 46, 48–70. [Google Scholar] [CrossRef] [Green Version]
- Ascensão, F.; Santos-Reis, J.F.M. Highway verges as habitat providers for small mammals in agrosilvopastoral environments. Biodivers. Conserv. 2012, 21, 3681–3697. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.M.; Mathias, M.; Mira, A.; Simoes, M.P. Vegetation structure and composition of road verge and meadow sites colonized by Cabrera vole (Microtus cabrerae Thomas). Pol. J. Ecol. 2007, 55, 481. [Google Scholar]
- Schwartz, M.W.; Cook, C.N.; Pressey, R.L.; Pullin, A.S.; Runge, M.C.; Salafsky, N.; Sutherland, W.J.; Williamson, M.A. Decision Support Frameworks and Tools for Conservation. Conserv. Lett. 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction (Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2010. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Razgour, O.; Rebelo, H.; di Febbraro, M.; Russo, D. Painting maps with bats: Species distribution modelling in bat research and conservation. Hystrix 2016, 27, 30–37. [Google Scholar]
- Valerio, F.; Carvalho, F.; Barbosa, A.M.; Mira, A.; Santos, S.M. Accounting for Connectivity Uncertainties in Predicting Roadkills: A Comparative Approach between Path Selection Functions and Habitat Suitability Models. Environ. Manag. 2019, 64, 329–343. [Google Scholar] [CrossRef] [Green Version]
- Tulloch, A.I.T.; Sutcliffe, P.; Naujokaitis-Lewis, I.; Tingley, R.; Brotons, L.; Ferraz, K.M.P.M.B.; Possingham, H.; Guisan, A.; Rhodes, J.R. Conservation planners tend to ignore improved accuracy of modelled species distributions to focus on multiple threats and ecological processes. Biol. Conserv. 2016, 199, 157–171. [Google Scholar] [CrossRef] [Green Version]
- Valerio, F.; Basile, M.; Balestrieri, R.; Posillico, M.; Di Donato, S.; Altea, T.; Matteucci, G. The reliability of a composite biodiversity indicator in predicting bird species richness at different spatial scales. Ecol. Indic. 2016, 71, 627–635. [Google Scholar] [CrossRef]
- Mestre, F.; Pita, R.; Paupério, J.; Martins, F.M.S.; Alves, P.C.; Mira, A.; Beja, P. Combining distribution modelling and non-invasive genetics to improve range shift forecasting. Ecol. Modell. 2015, 297, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, H.; Jones, G. Ground validation of presence-only modelling with rare species: A case study on Barbastella barbastellus (Chiroptera: Vespertilionidae). J. Appl. Ecol. 2010, 47, 410–420. [Google Scholar] [CrossRef]
- Robinson, O.J.; Ruiz-Gutierrez, V.; Fink, D. Correcting for bias in distribution modelling for rare species using citizen science data. Divers. Distrib. 2018, 24, 460–472. [Google Scholar] [CrossRef] [Green Version]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.-N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Leitão, P.J.; Santos, M.J. Improving Models of Species Ecological Niches: A Remote Sensing Overview. Front. Ecol. Evol. 2019, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rivas-Martínez, S. Les étages bioclimatiques de la végétation de la Péninsule Ibérique. Anales del Jardín Botánico de Madrid 1981, 37, 251–268. [Google Scholar]
- Rosário, I.T.; Cardoso, P.E.; da Luz Mathias, M. Is habitat selection by the Cabrera vole (Microtus cabrerae) related to food preferences? Mamm. Biol. 2008, 73, 423–429. [Google Scholar] [CrossRef]
- Coops, N.C.; Wulder, M.A. Breaking the Habit(at). Trends Ecol. Evol. 2019, 34, 585–587. [Google Scholar] [CrossRef] [PubMed]
- Jones, H.G.; Vaughan, R.A. Remote Sensing of Vegetation: Principles, Techniques, and Applications; Oxford University Press: New York, NY, USA, 2011; ISBN 978-0-19-920779-4. [Google Scholar]
- Pettorelli, N.; Ryan, S.; Mueller, T.; Bunnefeld, N.; Jedrzejewska, B.; Lima, M.; Kausrud, K. The Normalized Difference Vegetation Index (NDVI): Unforeseen successes in animal ecology. Clim. Res. 2011, 46, 15–27. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 9, 503–510. [Google Scholar] [CrossRef]
- Oeser, J.; Heurich, M.; Senf, C.; Pflugmacher, D.; Belotti, E.; Kuemmerle, T. Habitat metrics based on multi-temporal Landsat imagery for mapping large mammal habitat. Remote Sens. Ecol. Conserv. 2019, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Randin, C.F.; Ashcroft, M.B.; Bolliger, J.; Cavender-Bares, J.; Coops, N.C.; Dullinger, S.; Dirnböck, T.; Eckert, S.; Ellis, E.; Fernández, N.; et al. Remote Sensing of Environment Monitoring biodiversity in the Anthropocene using remote sensing in species distribution models. Remote Sens. Environ. 2020, 239, 111626. [Google Scholar] [CrossRef]
- Vaniscotte, A.; Pleydell, D.R.; Raoul, F.; Quéré, J.P.; Jiamind, Q.; Wang, Q.; Tiaoying, L.; Bernarda, N.; Coeurdassier, M.; Delattre, P.; et al. Modelling and spatial discrimination of small mammal assemblages: An example from western Sichuan (China). Ecol. Model. 2009, 220, 1218–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leyequien, E.; Verrelst, J.; Slot, M.; Schaepman-Strub, G.; Heitkonig, I.M.A.; Skidmore, A. Capturing the fugitive: Applying remote sensing to terrestrial animal distribution and diversity. Int. J. Appl. Earth Obs. 2007, 9, 1–20. [Google Scholar] [CrossRef]
- Kumar, S.; Simonson, S.E.; Stohlgren, T.J. Effects of spatial heterogeneity on butterfly species richness in Rocky Mountain National Park, CO, USA. Biodivers. Conserv. 2009, 18, 739. [Google Scholar] [CrossRef]
- Berger, M.; Moreno, J.; Johannessen, J.A.; Levelt, P.F.; Hanssen, R.F. ESA’s sentinel missions in support of Earth system science. Remote Sens. Environ. 2012, 120, 84–90. [Google Scholar] [CrossRef]
- Drusch, M.; Bello, U.; del Carlier, S.; Fernandez, V.; Gascon, F.; Hoersch, B.; Isola, C.; Laberinti, P.P. Sentinel-2: ESA’ s Optical High-Resolution Mission for GMES Operational Services. Remote Sens. Environ. 2012, 120, 25–36. [Google Scholar] [CrossRef]
- Andreo, V.; Belgiu, M.; Andreo, V.; Belgiu, M.; Hoyos, D.B.; Osei, F.; Provensal, C.; Stein, A. Ecological Informatics Rodents and satellites: Predicting mice abundance and distribution with Sentinel-2 data. Ecol. Inform. 2019, 51, 157–167. [Google Scholar] [CrossRef]
- Arenas-Castro, S.; Gonçalves, J.; Alves, P.; Alcaraz-Segura, D.; Honrado, J.P. Assessing the multi-scale predictive ability of ecosystem functional attributes for species distribution modelling. PLoS ONE 2018, 13, e0199292. [Google Scholar] [CrossRef] [Green Version]
- Kursa, M.B.; Rudnicki, W.R. Feature Selection with the Boruta Package. J. Stat. Softw. 2010, 36, 1–13. Available online: http://www.jstatsoft.org/v36/i11/ (accessed on 2 April 2019). [CrossRef] [Green Version]
- IPMA. Instituo Português do Mar e da Atmosfera. Normais Climatológicas 2019, 675, 1981–2010. Available online: http://www.ipma.pt/pt/index.html (accessed on 25 February 2019).
- Pinto-Correia, T.; Ribeiro, N.; Sá-Sousa, P. Introducing the montado, the cork and holm oak agroforestry system of southern Portugal. Agrofor. Syst. 2011, 82, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Joffre, R.; Rambal, S.; Ratte, J.P. The dehesa system of southern Spain and Portugal as a natural ecosystem mimic. Agrofor. Syst. 1999, 45, 57. [Google Scholar] [CrossRef]
- Branco, O.; Bugalho, M.; Silva, L.N.; Barreira, R.; Vaz, P.; Días, F. Hotspot Areas for Biodiversity and Ecosystem Services in Montados—HABEaS. WWF Mediterr. Programme Port. 2010, 55. [Google Scholar]
- Díaz-Villa, M.D.; Marañón, T.; Arroyo, J.; Garrido, B. Soil seed bank and floristic diversity in a forest-grassland mosaic in southern Spain. J. Veg. Sci. 2003, 5, 701–709. [Google Scholar] [CrossRef]
- Oppermann, R.; Beaufoy, G.; Jones, G. High Nature Value Farming in Europe. 35 European Countries—Experiences and Perspectives; Verlag Regionalkultur: Ubstadt-Weiher, Germany, 2012. [Google Scholar]
- Pinto-Correia, T.; Godinho, S. Chapter 4 Changing Agriculture–Changing Landscapes: What is Going on in the High Valued Montado. In Agriculture in Mediterranean Europe: Between Old and New Paradigms; Emerald Group Publishing Limited: Bingley, UK, 2013; pp. 75–90. [Google Scholar]
- Proença-Ferreira, A.; Ferreira, C.; Leitão, I.; Paupério, J.; Sabino-Marques, H.; Barbosa, S.; Lambin, X.; Alves, P.C.; Beja, P.; Moreira, F.; et al. Drivers of survival in a small mammal of conservation concern: An assessment using extensive genetic non-invasive sampling in fragmented farmland. Biol. Conserv. 2019, 230, 131–140. [Google Scholar] [CrossRef] [Green Version]
- European Space Agency, 2018. SNAP—Version 6.0.0. Available online: http://step.esa.int/main/ (accessed on 14 March 2019).
- Brodu, N. Super-resolving multiresolution images with band-independent geometry of multispectral pixels. IEEE Trans. Geosci. Remote Sens. 2017, 55, 4610–4617. [Google Scholar] [CrossRef] [Green Version]
- Fragoso-Campón, L.; Quirós, E.; Mora, J. Overstory-understory land cover mapping at the watershed scale: Accuracy enhancement by multitemporal remote sensing analysis and LiDAR. Environ. Sci. Pollut. Res. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Godinho, S.; Guiomar, N.; Machado, R.; Santos, P.; Sá-Sousa, P.; Fernandes, J.P.; Neves, N.; Pinto-Correia, T. Assessment of environment, land management, and spatial variables on recent changes in montado land cover in southern Portugal. Agrofor. Syst. 2016, 1, 177–192. [Google Scholar] [CrossRef] [Green Version]
- Godinho, S.; Guiomar, N.; Gil, A. Estimating tree canopy cover percentage in a mediterranean silvopastoral systems using Sentinel-2A imagery and the stochastic gradient boosting algorithm. Int. J. Remote Sens. 2018, 39, 4640–4662. [Google Scholar] [CrossRef]
- Hill, T.C.; Williams, M.; Bloom, A.A.; Mitchard, E.T.A.; Ryan, C.M. Are inventory based and remotely sensed above-ground biomass estimates consistent? PLoS ONE 2013, 8, e74170. [Google Scholar] [CrossRef] [Green Version]
- Serrano, J.; Shahidian, S.; Marques da Silva, J.; Carvalho, M. Proximal and remote sensing to evaluate the impact of climatic inter-annual irregularity in montado Mediterranean ecosystems-a case study. In Proceedings of the 3rd Agriculture and Climate Change Conference, Budapest, Hungary, 24–27 March 2019. [Google Scholar]
- Delegido, J.; Verrelst, J.; Alonso, L.; Moreno, J. Evaluation of Sentinel-2 Red-Edge Bands for Empirical Estimation of Green LAI and Chlorophyll Content. Sensor 2011, 11, 7063–7081. [Google Scholar] [CrossRef] [Green Version]
- Navarro, G.; Caballero, I.; Silva, G.; Parra, P. Evaluation of forest fire on Madeira Island using Sentinel-2A MSI imagery. Int. J. Appl. Earth Obs. Geoinf. 2017, 58, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J., Jr.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the Great Plains with ERTS. NASA Special Publ. 1974, 351, 309. [Google Scholar]
- Marsett, R.C.; Qi, J.; Heilman, P.; Biedenbender, S.H.; Watson, M.C.; Amer, S.; Weltz, M.; Goodrich, D.; Marsett, R. Remote sensing for grassland management in the arid southwest. Rangel. Ecol. Manag. 2006, 59, 530–540. [Google Scholar] [CrossRef]
- Hardisky, M.A.; Klemas, V.; Smart, R.M. The influence of soil salinity, growth form, and leaf moisture on the spectral radiance of Spartina alterniflora canopies. Photogram Eng. Remote Sens. 1983, 49, 77–83. [Google Scholar]
- Gao, B. NDWI A Normalized Difference Water Index for Remote Sensing of Vegetation Liquid Water from Space. Remote Sens. Environ. 1996, 266, 257–266. [Google Scholar] [CrossRef]
- Merzlyak, M.N.; Gitelson, A.A.; Chivkunova, O.B.; Rakitin, V.Y. Non-destructive optical detection of pigment changes during leaf senescence and fruit ripening. Physiol. Plant. 1999, 106, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Escadafal, R. Remote sensing of arid soil surface color with Landsat thematic mapper. Adv. Space Res. 1989, 9, 159–163. [Google Scholar] [CrossRef]
- Guerschman, J.P.; Hill, M.J.; Renzullo, L.J.; Barrett, D.J.; Marks, A.S.; Botha, E.J. Estimating fractional cover of photosynthetic vegetation, non-photosynthetic vegetation and bare soil in the Australian tropical savanna region upscaling the EO-1 Hyperion and MODIS sensors. Remote Sens. Environ. 2009, 113, 928–945. [Google Scholar] [CrossRef]
- Haralick, R.M.; Shanmugam, K.; Dinstein, I. Textural Features for Image Classification. IEEE Trans. Syst. Man. Cybern. 1973, 6, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Rocchini, D.; Marcantonio, M.; Ricotta, C. Measuring Rao’s Q diversity index from remote sensing: An open source solution. Ecol. Indic. 2017, 72, 234–238. [Google Scholar] [CrossRef]
- Schulte to Bühne, H.; Pettorelli, N. Better together: Integrating and fusing multispectral and radar satellite imagery to inform biodiversity monitoring, ecological research and conservation science. Methods Ecol. Evol. 2018, 9, 849–865. [Google Scholar] [CrossRef]
- Zvoleff, A. Glcm: Calculate Textures from Grey-Level Co-Occurrence Matrices (GLCMs) R Package Version 3.5.0. 2016. Available online: https://CRAN.R-project.org/package=glcm (accessed on 12 December 2018).
- R Development Core Team, R. A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; R Development Core Team R: Vienna, Austria, 2018; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 10 November 2017).
- ASTER GDEM. 2018. Available online: https://asterweb.jpl.nasa.gov/gdem.asp (accessed on 11 May 2018).
- Brenning, A.; Bangs, D.; Becker, M. RSAGA: SAGA Geoprocessing and Terrain Analysis. R package Version 1.0.0. 2018. Available online: https://CRAN.R-project.org/package=RSAGASdfsdf (accessed on 11 May 2018).
- OpenStreetMap Contributors. 2018. Europe OpenStreetMap Data. Available online: https://download.geofabrik.de/europe/portugal.html (accessed on 5 February 2018).
- European Union, Copernicus Land Monitoring Service 2018, European Environment Agency (EEA). Available online: https://land.copernicus.eu/ (accessed on 19 May 2018).
- Gichenje, H.; Pinto-correia, T.; Godinho, S. Remote Sensing Applications: Society and Environment An analysis of the drivers that affect greening and browning trends in the context of pursuing land degradation-neutrality. Remote Sens. Appl. Soc. Environ. 2019, 15, 100251. [Google Scholar] [CrossRef]
- Kursa, M.B.; Rudnicki, W.R. The all relevant feature selection using random forest. arXiv 2011, arXiv:1106.5112Sdfsdf. [Google Scholar]
- Breiman, L.E.O. Random Forests. Machin. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.S.; Murphy, M.A.; Holden, Z.A.; Cushman, S.A. Modeling Species Distribution and Change Using Random Forest; Springer: New York, NY, USA, 2011; pp. 139–159. [Google Scholar] [CrossRef]
- Ehrlinger, J. ggRandomForests: Visually Exploring Random Forests. R Package Version 2.0.1. 2016. Available online: https://CRAN.R-project.org/package=ggRandomForests (accessed on 18 April 2019).
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Carrié, R.; Lopes, M.; Ouin, A.; Andrieu, E. Bee diversity in crop fields is influenced by remotely-sensed nesting resources in surrounding permanent grasslands. Ecol. Indic. 2018, 90, 606–614. [Google Scholar] [CrossRef]
- Dube, T.; Mutanga, O.; Adam, E.; Ismail, R. Intra-and-Inter Species Biomass Prediction in a Plantation Forest: Testing the Utility of High Spatial Resolution Spaceborne Multispectral RapidEye Sensor and Advanced Machine Learning Algorithms. Sensors 2014, 14, 15348–15370. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Azeda, C.; Guiomar, N.; Pinto-correia, T. The effects of grazing management in montado fragmentation and heterogeneity. Agrofor. Syst. 2016, 90, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Serafini, V.N.; Priotto, J.W.; Gomez, M.D. Effects of agroecosystem landscape complexity on small mammals: A multi-species approach at different spatial scales. Landsc. Ecol. 2019, 8, 1–13. [Google Scholar] [CrossRef]
- Coffin, A.W. From roadkill to road ecology: A review of the ecological effects of roads. J. Transp. Geogr. 2007, 15, 396–406. [Google Scholar] [CrossRef]
- Jumeau, J.; Boucharel, P.; Handrich, Y. Road-related landscape elements as a habitat: A main asset for small mammals in an intensive farming landscape. Basic Appl. Ecol. 2017, 25, 15–27. [Google Scholar] [CrossRef]
- de Redon, L.; Le Viol, I.; Jiguet, F.; Machon, N.; Scher, O.; Kerbiriou, C. Road network in an agrarian landscape: Potential habitat, corridor or barrier for small mammals? Acta Oecol. 2015, 62, 58–65. [Google Scholar] [CrossRef]
- Silva, C.; Simões, M.P.; Mira, A.; Santos, S.M. Factors influencing predator roadkills: The availability of prey in road verges. J. Environ. Manag. 2019, 247, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Pinto-Correia, T. Exploring the use of landscape as the basis for the identification of High Nature Value farmland: A case-study in the Portuguese Montado. In Proceedings of the 10th European IFSA Symposium on Producing and Reproducing Farming Systems: New Modes of Organisation for Sustainable Food Systems of Tomorrow, Aarhus, Denmark, 1–4 July 2012; p. 7. [Google Scholar]
- Merrick, M.J.; Koprowski, J.L. Circuit theory to estimate natal dispersal routes and functional landscape connectivity for an endangered small mammal. Landsc. Ecol. 2017, 32, 1163–1179. [Google Scholar] [CrossRef]
- Donald, P.F.; Evans, A.D. Habitat connectivity and matrix restoration: The wider implications of agri-environment schemes. J. Appl. Ecol. 2006, 43, 209–218. [Google Scholar] [CrossRef]
- Rapacciuolo, G. Strengthening the contribution of macroecological models to conservation practice. Global. Ecol. Biogeogr. 2019, 28, 54–60. [Google Scholar] [CrossRef]
- Gottschalk, T.K.; Aue, B.; Hotes, S.; Ekschmitt, K. Influence of grain size on species-habitat models. Ecol. Model. 2011, 222, 3403–3412. [Google Scholar] [CrossRef]
- Guisan, A.; Broennimann, O.; Engler, R.; Vust, M.; Yoccoz, N.G.; Lehmann, A.; Zimmermann, N.E. Using Niche-Based Models to Improve the Sampling of Rare Species. Conserv. Biol. 2006, 20, 501–511. [Google Scholar] [CrossRef]
- Mi, C.; Huettmann, F.; Guo, Y.; Han, X.; Wen, L. Why choose Random Forest to predict rare species distribution with few samples in large undersampled areas? Three Asian crane species models provide supporting evidence. PeerJ 2017, 5, e2849. [Google Scholar] [CrossRef] [Green Version]
- Thornton, M.W.; Atkinson, P.M.; Holland, D.A. A linearised pixel-swapping method for mapping rural linear land cover features from fine spatial resolution remotely sensed imagery. Comput. Geosci. 2007, 33, 1261–1272. [Google Scholar] [CrossRef]
- Rapinel, S.; Mony, C.; Lecoq, L.; Lecoq, L.; Clément, B.; Thomas, A.; Hubert-Moy, L. Evaluation of Sentinel-2 time-series for mapping floodplain grassland plant communities. Remote Sens. Environ. 2019, 223, 115–129. [Google Scholar] [CrossRef]
- Pereira, O.J.R.; Melfi, A.J.; Montes, C.R. Image fusion of Sentinel-2 and CBERS-4 satellites for mapping soil cover in the Wetlands of Pantanal. Int. J. Image Data Fusion 2017, 8, 148–172. [Google Scholar] [CrossRef]
- Speiser, J.L.; Miller, M.E.; Tooze, J.; Ip, E. A comparison of random forest variable selection methods for classification prediction modeling. Expert Syst. Appl. 2019, 134, 93–101. [Google Scholar] [CrossRef]
- Engler, J.O.; Stiels, D.; Schidelko, K.; Strubbe, D.; Quillfeldt, P.; Brambilla, M. Avian SDMs: Current state, challenges, and opportunities. J. Avian. Biol. 2017, 48, 1483–1504. [Google Scholar] [CrossRef] [Green Version]
- Bishop-taylor, R.; Broich, M. Evaluating static and dynamic landscape connectivity modelling using a 25-year remote sensing time series. Landsc. Ecol. 2018, 33, 625–640. [Google Scholar] [CrossRef]
- Milanesi, P.; Della Rocca, F.; Robinson, R.A. Integrating dynamic environmental predictors and species occurrences: Toward true dynamic species distribution models. Ecol. Evol. 2019, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Arenas-Castro, S.; Regos, A.; Gonçalves, J.F.; Alcaraz-Segura, D.; Honrado, J. Remotely Sensed Variables of Ecosystem Functioning Support Robust Predictions of Abundance Patterns for Rare Species. Remote Sens. 2019, 11, 2086. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Type | Code | Description | Spectral Band or Equation | Reference |
|---|---|---|---|---|---|
| Spectral Bands | - | Blue | B2 | [40] | |
| Green | B3 | ||||
| Red | B4 | ||||
| Red Edge 1 | (SR)B5 | ||||
| Red Edge 2 | (SR)B6 | ||||
| Red Edge 3 | (SR)B7 | ||||
| NIR 1 | B8 | ||||
| NIR 2 | (SR)B8a | ||||
| SWIR1 | (SR)B11 | ||||
| SWIR2 | (SR)B12 | ||||
| Spectral Indices | Vegetation Biomass Indices | NDI45 | Normalized Difference Index 45 | [60] | |
| NDRE1 | Normalized difference red edge index 1 | [61] | |||
| NDRE2 | Normalized difference red edge index 2 | ||||
| NDRE3 | Normalized difference red edge index 3 | ||||
| NDVI | Normalized Difference Vegetation Index | [62] | |||
| SATVI | Soil-adjusted Total Vegetation Index | [63] | |||
| Vegetation and landscape Water content | NDII | Normalized Difference Infrared Index | [64] | ||
| NDWI | Normalized difference water index | [65] | |||
| Senescent vegetation and soil surfaces indices | PSRI | Plant Senescence Reflectance Index | [66] | ||
| BI2 | Second Brightness Index | [67] | |||
| SWIR32 | Shortwave infrared Reflectance 3/2 ratio | [68] | |||
| Textural and Diversity Indices | Co-occurrence matrix | GLCM_M | Mean | Calculated using the first principal component (PC1) with a 3 × 3 pixels spatial moving window in all directions (0°, 45°, 90°, and 135°) | [69] |
| GLCM_Cor | Correlation | ||||
| GLCM_Con | Contrast | ||||
| GLCM_D | Dissimilarity | ||||
| GLCM_E | Entropy | ||||
| GLCM H | Homogeneity | ||||
| GLCM S | Second Moment | ||||
| GLCM_V | Variance | ||||
| Diversity index | Rao’s Q | Rao’s quadratic entropy | Calculated using the NDVI with a 3 × 3 pixels spatial moving window | [70] |
| Group | Type | Denomination | Methodology | Data Source/Reference |
|---|---|---|---|---|
| Topographic | - | Altitude | - | [74] |
| Slope | Calculated from the Aster (2018) digital elevation model | [75] | ||
| Roughness | ||||
| Topographic wetness index | ||||
| Inference | - | Distance to paved roads | Calculated by applying Euclidean distance to a specific landscape class | [76] |
| Distance to urban | [77] | |||
| Distance to water bodies |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valerio, F.; Ferreira, E.; Godinho, S.; Pita, R.; Mira, A.; Fernandes, N.; Santos, S.M. Predicting Microhabitat Suitability for an Endangered Small Mammal Using Sentinel-2 Data. Remote Sens. 2020, 12, 562. https://doi.org/10.3390/rs12030562
Valerio F, Ferreira E, Godinho S, Pita R, Mira A, Fernandes N, Santos SM. Predicting Microhabitat Suitability for an Endangered Small Mammal Using Sentinel-2 Data. Remote Sensing. 2020; 12(3):562. https://doi.org/10.3390/rs12030562
Chicago/Turabian StyleValerio, Francesco, Eduardo Ferreira, Sérgio Godinho, Ricardo Pita, António Mira, Nelson Fernandes, and Sara M. Santos. 2020. "Predicting Microhabitat Suitability for an Endangered Small Mammal Using Sentinel-2 Data" Remote Sensing 12, no. 3: 562. https://doi.org/10.3390/rs12030562