A New Group II Phospholipase A2 from Walterinnesia aegyptia Venom with Antimicrobial, Antifungal, and Cytotoxic Potential



1
Botany and Microbiology Department, Science College, King Saud University, P.O. Box 22452, Riyadh 11495, Saudi Arabia
2
Laboratory of Plant Biotechnology Applied to Crop Improvement, Faculty of Science of Sfax, University of Sfax, Sfax 3038, Tunisia
3
Biochemistry Department, Science College, King Saud University, P.O. Box 22452, Riyadh 11495, Saudi Arabia
*
Author to whom correspondence should be addressed.
Processes 2020, 8(12), 1560; https://doi.org/10.3390/pr8121560
Submission received: 15 October 2020
/
Revised: 22 November 2020
/
Accepted: 23 November 2020
/
Published: 27 November 2020
(This article belongs to the Special Issue Applied Enzymology & Environmental Biotechnology)
Abstract
:Many venomous species, especially snakes, contain a variety of secreted phospholipases A2 that contribute to venom toxicity and prey digestion. We characterized a novel highly toxic phospholipase A2 of group II, WaPLA2-II, from the snake venom of Saudi Walterinnesia aegyptia (W. aegyptia). The enzyme was purified using a reverse phase C18 column. It is a monomeric protein with a molecular weight of approximately 14 kDa and an NH2-terminal amino acid sequence exhibiting similarity to the PLA2 group II enzymes. WaPLA2-II, which contains 2.5% (w/w) glycosylation, reached a maximal specific activity of 1250 U/mg at pH 9.5 and 55 °C in the presence of Ca2+ and bile salts. WaPLA2-II was also highly stable over a large pH and temperature range. A strong correlation between antimicrobial and indirect hemolytic activities of WaPLA2 was observed. Additionally, WaPLA2-II was found to be significantly cytotoxic only on cancerous cells. However, chemical modification with para-Bromophenacyl bromide (p-BPB) inhibited WaPLA2-II enzymatic activity without affecting its antitumor effect, suggesting the presence of a separate ‘pharmacological site’ in snake venom phospholipase A2 via its receptor binding affinity. This enzyme is a candidate for applications including the treatment of phospholipid-rich industrial effluents and for the food production industry. Furthermore, it may represent a new therapeutic lead molecule for treating cancer and microbial infections.
1. Introduction
The phospholipase A2 (PLA2) superfamily consists of many enzymes that catalyze the hydrolysis of the sn-2 ester bond in a variety of phospholipids. Among the six groups of enzymes, the secreted PLA2 group (sPLA2) is composed of small proteins (14–18 kDa) containing five to eight disulfide bonds, which are found in pancreatic and intestine juices, dromedary tears, and venoms [1,2,3,4,5,6,7,8]. The mammalian secreted PLA2 enzymes consist of 12 members, (IB, IIA, IIC, IID, IIE, IIF, III, V, X, IX, XIIA, XIIB) and otoconin-95, classified by amino acid composition, catalytic function, and sequence similarity [9,10,11,12].
Snake venom sPLA2 (svPLA2), plays a significant role in early morbidity and mortality from snakebite, causing destruction of tissues, derangement of homeostatic mechanisms critical for coagulation and oxygen transport, and death by paralysis [13,14,15]. Based on their primary structure, svPLA2s have been classified into four groups [16,17]. Snake venom sPLA2 belongs to either group (G) IA (Elapidae and Hydrophidae snakes) or G IIA/IIB (Crotalidae and Viperidae snakes), based on sequence similarity, position of disulfide bonds, and loop insertions [18,19,20]. They exhibit a variety of pharmacologic and antitumor activities [21,22,23,24]. The svPLA2 G IIA enzyme is the most studied among the sPLA2s with respect to infection, inflammation, and its bactericidal effects [25,26,27,28,29]. SvPLA2s belonging to G II are subdivided into two major types: isoforms Asp49 and Lys49. They exhibit unique interactions and different mechanisms of action. Asp49 represents toxins with catalytic activity, whereas Lys49 are catalytically inactive [19,30]. Interestingly, these isoforms show a significant functional diversity regardless of their ability to hydrolyze phospholipids, which explains why they can be used for a variety of therapeutic purposes [31,32,33]. These activities are not limited to svPLA2s as they may also be found in mammalian sPLA2s. However, svPLA2 is of considerable interest because of its wide variety of isoforms and pharmacological properties. These depend not only on the particular species, but also on the breeding conditions of the snake [34,35,36]. The therapeutic values of the remarkable panoply of biomolecules in snakes can be exploited, which provide researchers with a foundation to development mimetic compounds for therapeutic applications.
In Saudi Arabia, vertebrate fauna is an active field of study. Snakes from the family Elapidae are large in size, consisting of a large head, a long tail, and contain a neurotoxic venom [37,38]. There are three species belonging to this family in the Kingdom: The Arabian cobra (Naja haje arabica) in the Southwestern region, the black cobra (W. aegyptia) in the Central, Northern, and Eastern regions, and Walterinnesia morgani which was identified for the first time in the Turaif region [39].
Despite the medical relevance of accidents caused by W. aegyptia in Saudi Arabia, only in recent years have studies to elucidate the venom composition and its pharmacological properties received significant attention. In our laboratory, a previous study by Ben Bacha et al. [40] resulted in the discovery and purification of a novel PLA2 G I enzyme from the venom of W. aegyptia. Its antimicrobial, antitumoral, and hemolytic properties were evaluated and it was found to exhibit potent, indirect hemolytic and antimicrobial activities. The present work extends these findings and reports the characterization of a new purified enzyme, WaPLA2 G II. Comparative studies were performed using the WaPLA2 G I previously isolated from the same venom [40]. Our results provide important information on the structure/function relationship of svPLA2 and focus on the importance of the spreading effect of the enzyme on snakebite envenomation. This may assist our understanding of the pathophysiological processes involved in sPLA2 and improve anti-venom therapy for bites caused by W. aegyptia and other species.
2. Materials and Methods
2.1. Venom Collection
Walterinnesia aegyptia specimens were captured from the southwest region of Saudi Arabia and were kept in large tanks in the serpentarium in the Zoology Department of the College of Science, King Saud University. Venom was milked from the adult snakes by applying mechanical pressure to the venom glands. The venom was collected in sterile tubes, dried, and stored in the dark at 44 °C. They were resuspended in saline prior to use.
2.2. Protein Analysis
Protein concentration was measured using the Bradford technique [41]. Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was performed using 15% polyacrylamide gels containing a reducing agent (β-mercaptoethanol). For N-terminal sequencing, Edman’s degradation technique was used according to [42]. For the detection of glycan chains in purified sPLA2, the Spiro method [43] was followed using glucose as a standard, whereas the pure sPLA2 was chemically modified using p-BPB following the protocol of [44].
2.3. Phospholipase Assay
PLA2 activity was measured titrimetrically using a pH-stat (842 Titrando, Metrohm) at pH and temperatures below optimal conditions, and phosphatidylcholine (PC) as a substrate in the presence of Ca2+ and bile salts [45]. One μmole of fatty acid liberated under optimum conditions was equivalent to one unit of phospholipase activity.
2.4. Purification of WaPLA2-II
W. aegyptia venom was dried, suspended in saline solution (0.15 M), and centrifuged for 30 min at 16,000× g at 4 °C. Following precipitation with 65% ammonium sulfate, the protein fraction was centrifuged (16,000× g, 30 min, 4 °C). The resulting pellet was dissolved in water, incubated at 70 °C for 30 min, and finally centrifuged for 20 min at 16,000× g at 4 °C. The dissolved pellet was applied to a reverse phase C18 column (250 × 4.6 mm, 5 mm; Beckman, Fullerton, CA, USA) pre-equilibrated with 0.1% TFA in water. Protein elution was performed at a flow rate of 0.5 mL/min over 50 min with a linear acetonitrile gradient (0–80%). The active fractions were pooled and lyophilized. The molecular weight of the isolated enzyme was estimated by size exclusion chromatography. The lyophilized WaPLA2-II (50 µg) was dissolved in 20 µL of 100 mM phosphate buffer, pH 7, containing 0.15 M NaCl and loaded onto a size exclusion high performance liquid chromatography (HPLC) column (Bio-sil SEC-125, 300 mm × 7.8 mm) pre-equilibrated with the same buffer. Elution was performed with phosphate buffer at 1 mL/min over 20 min.
2.5. Enzymes
2.6. In Vitro Hemolytic Activities
Direct and indirect hemolytic activities were determined according to [46]. To measure direct hemolytic activity, suspensions of washed red blood cells (RBCs) were prepared in phosphate-buffered saline (PBS) and used as a substrate. Different concentrations of the enzyme (0 to 3 μg) were incubated with 1 mL of the substrate suspension at 37 °C for 30 min. To determine indirect hemolytic activity, 1 mL of fresh egg phosphatidylcholine (PC) was mixed with 8 mL of the substrate. To terminate the reaction, 9 mL of ice-cold PBS was added and the mixture was centrifuged for 20 min at 1500× g at 4 °C. The degree of hemolysis by PLA2 was calculated as the percentage of hemoglobin released at 530 nm compared with 100% hemolysis obtained by adding 1% Triton X-100.
2.7. Antimicrobial Activity
We tested the snake PLA2 against 4 human pathogenic fungi strains and 9 bacterial species, all of them associated with the human intestinal and oral tract or skin. The fungi used in our study were: Aspergillus niger, Botrytis cinerea, Fusarium solani, and Penicillium digitatum. The five gram-positive (Bacillus subtilis ATCC 6633, Bacillus cereus ATCC 14579, Enterococcus faecalis ATCC 29122, Staphylococcus epidermidis ATCC 14990, and Staphylococcus aureus ATCC 25923) and four gram-negative (Escherichia coli ATCC 25966, Klebsiella pneumonia ATCC 700603, Pseudomonas aeruginosa ATCC 27853, and Salmonella enteric ATCC 43972) bacteria species were also used.
- –
- Antibacterial activity
The bactericidal activity of the purified sPLA2 was evaluated by the well diffusion technique on nutrient agar medium following the method of [47]. The inhibitory effect on bacterial growth was measured by the inhibition zone and compared with that of standard antibiotics such as ampicillin at 10 μg/well. Petri dishes containing nutrient agar medium were uniformly smeared with pure test cultures of the human pathogenic bacteria. After the plates were incubated at 37 °C for 24 h, 10 μL of purified sPLA2 (1 mg/mL) was loaded into the pores. The zone of inhibition was measured in triplicate by comparing the control with the standard antibiotic. We also determined bacterial viability by calculating the colony-forming ability (CFU) of bacteria incubated in the presence or absence of the enzyme. For this assay, the enzyme was mixed with 2 × 107 CFU/mL bacteria in sterile Brain Heart Infusion (BHI) medium and incubated at 37 °C for 60 min in a shaker. The bactericidal activity of sPLA2 was calculated as the residual number of colony-forming units compared with the initial inoculums. The IC50 value corresponded to the enzyme concentration required to kill 50% of the initial inoculum. The results represent the means of three independent experiments.
- –
- Antifungal activity
We measured the antifungal activity of pure sPLA2 against 4 fungal strains by the disc diffusion technique using Sabouraud dextrose agar [48]. The pure enzyme (10 μL) was placed on sterile paper discs, deposited onto the center of inoculated Petri dishes, and incubated at 30 °C for 24 h. Cycloheximide (1 mg/mL) was used as positive control.
2.8. Cytotoxicity Test
Human breast adenocarcinoma (MDA-MB-231), human colorectal carcinoma (HCT-116 and LoVo), and human umbilical vein endothelial (HUVEC) cell lines were used in this study and were purchased from the American Type Culture Collection (ATCC, Manassas, VA, USA). They were grown in Dulbecco’s Modified Eagles Medium containing 15% fetal bovine serum at 37 °C in a 5% CO2-humidified incubator for 24 h. The MTT assay was used for assessing cell metabolic activity. Cell viability was calculated as the mean ± SD (n = 3) and was expressed as a relative percentage of the OD values measured at 550 nm for sPLA2-treated cells relative to the control. The cells (4 × 104 in each well) were incubated in a 96-well plate at 37 °C for 24 h in the absence or presence of different concentrations of WaPLA2, previously diluted in culture medium. Then, 20 µL of MTT (5 mg/mL in PBS) were added to the cells and incubated at 37 °C for 4 h. Finally, the medium was carefully removed and replaced with an equal volume of saline solution (50:50) in each well. In order to dissolve the formazan crystals, we mixed the preparations thoroughly using a shaker before measuring the absorbance at 550 nm.
3. Results
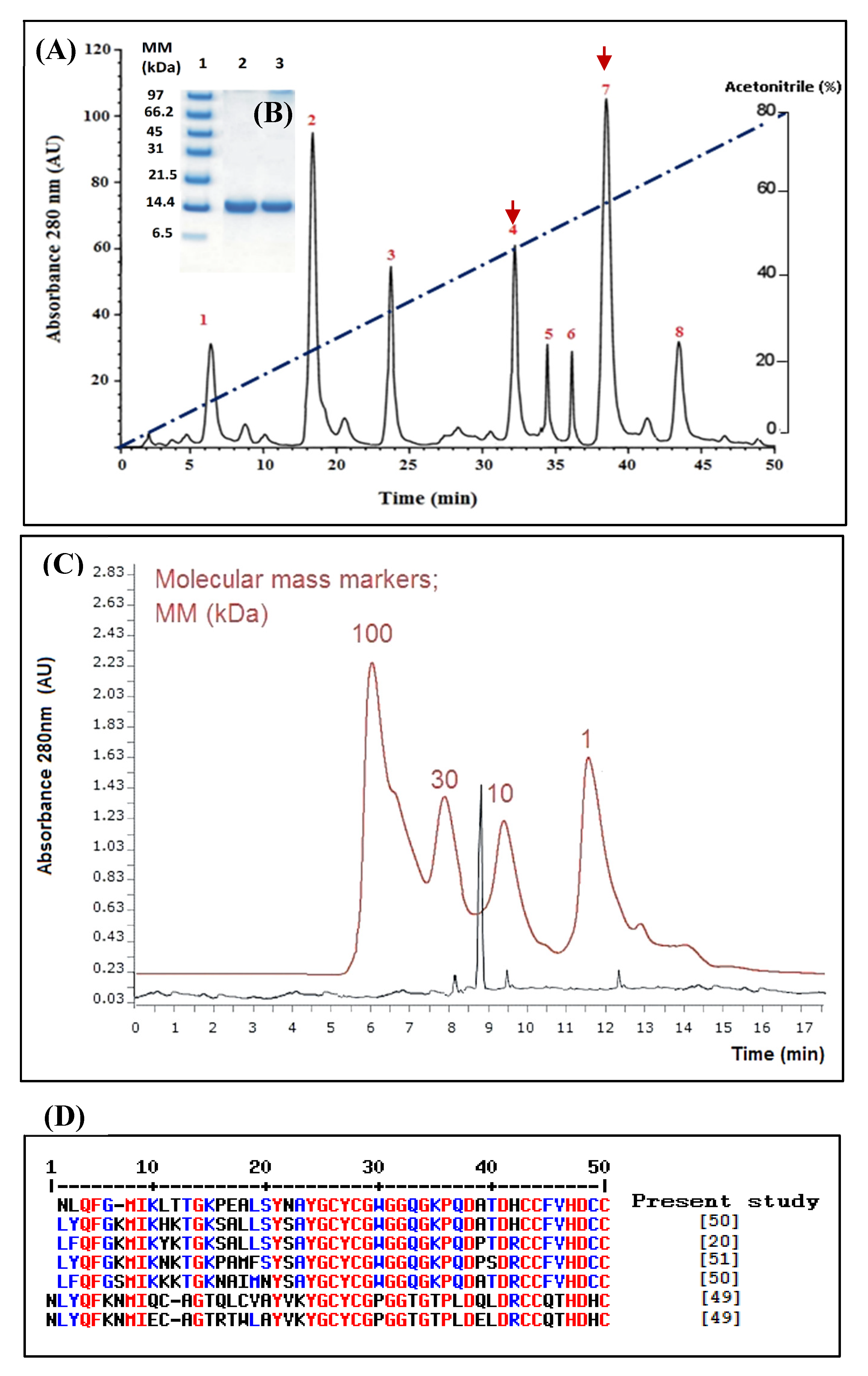
In our previous work [40], we obtained eight major peaks (1–8) upon fractionation of W. aegyptia venom by RP-HPLC on a C18 column and we, in particular, described peak 7 (WaPLA2-I) (Figure 1A). Peak 4 (named WaPLA2-II), which was eluted at 32 min, also attracted a lot our attention since it demonstrated good antibacterial and phospholipase activities. Hence, in the present study, the second active fraction (WaPLA2-II) was characterized and compared to the previously reported WaPLA2-I.
This single chromatographic step protocol demonstrated a significant degree of purity (25-fold) for WaPLA2-II with a recovery yield of 11.9% based on the original PLA2 activity (Table 1). At this stage, the purified enzyme exhibited a specific activity of 1250 U/mg at pH 9.5 and 55 °C using PC as substrate. It is of interest to note that both enzymes could be purified to homogeneity following the same rapid and high-yield protocol.
A single band corresponding to a molecular mass of approximately 14 kDa was revealed by SDS-PAGE analysis, under reducing conditions, of the active peak (Figure 1B). This was consistent with the molecular mass estimation using the HPLC size exclusion column, Bio-sil SEC-125 (300 × 7.8 mm) (Figure 1C). This indicates that the purified PLA2 is a monomeric protein as previously reported for all PLA2-IIA enzymes [8,34,36]. However, the active peak 7, WaPLA2-I, is a 30 kDa homodimer [40]. A similar result was obtained for the PLA2 G I isolated from Bungarus fasciatus venom [49].
Interestingly, SPLA2-IB used as a non-glycosylated control, WaPLA2-II was found to be glycosylated with a carbohydrate content of 2.5% compared with 2% for WaPLA2-I [40]. This finding is in accordance with that obtained by Zouari-Kessentini et al. who reported the first glycosylated PLA2 enzyme purified from venom of the Tunisian snake, Cerastes cerastes [50]. Moreover, a glycosylated form of the marine SPLA2-IIA was also identified with a carbohydrate content of 2.5% (data not shown) [8].
The NH2-terminal amino acid sequence of pure WaPLA2-II was determined (NLQFGMIKLTTGKPEALSYNAYGCYCGWGGQGKPQDATDHCCFVHDCC) and, as it is shown in Figure 1D, it exhibited a high similarity (≥83%) with the available sequences of secreted PLA2 G II isolated from the Cerastes cerastes [22], Echis carinatus, Vipera renardi, and Atheris nitschei snake venoms [22,50,51] (Figure 1D). A relatively low similarity was obtained with sPLA2 G I (≤55%) from Bungarus fasciatus venom [49]. As mentioned elsewhere, the different venom sPLA2 was classified based essentially on its primary structures, into four groups I, II, III, and IX so far. Therefore, our primary results allowed us to say that the phospholipase extracted from peak 4 could be a G II PLA2. However, further deep investigations (such as the determination of the entire sequence) are required to confirm that WaPLA2-II belongs to G II sPLA2. It is worthy to notice that G II svPLA2s have always been present in viperid and crotalid snakes, whereas G I svPLA2s are found in elapid and hydrophid snakes [50]. To our knowledge, we reported here for the first time, the purification to homogeneity of this svPLA2 G II from the snake family of Elapidae, from Saudi Arabia. This may be due to a particular composition of the venom of the Saudi snake or to change in the sequence of amino acids of the enzyme. This finding encourages us to sequence the entire enzyme and the gene coding for it to look for an eventual genetic mutation, which could generate a new subgroup.
3.1. Enzymatic Properties of W. aegyptia Venom PLA2
3.1.1. Influence of pH and Temperature on Phospholipase Activity and Stability
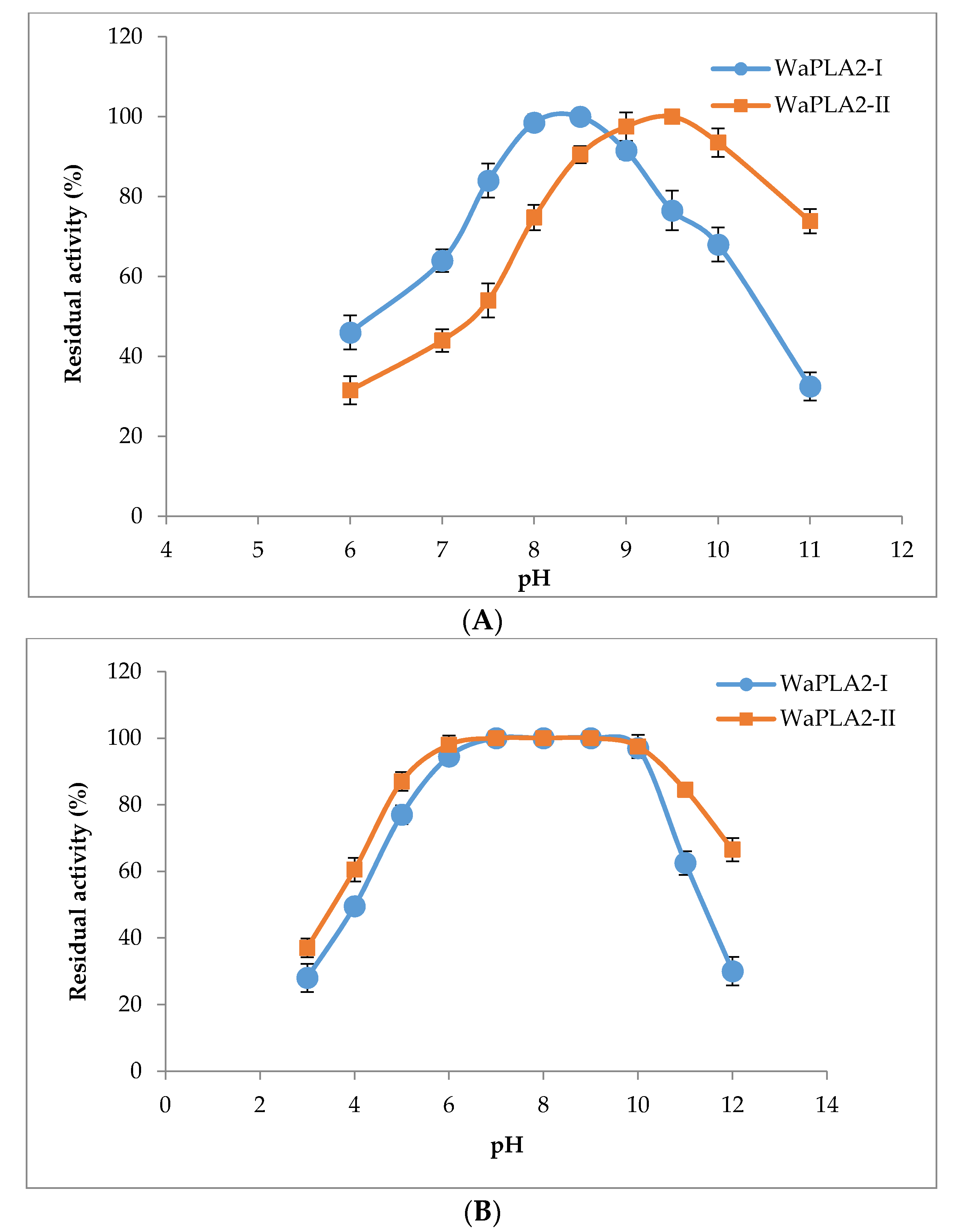
Lipolytic enzymes, including phospholipases and lipases, are versatile biocatalysts that are broadly utilized in different industrial applications, particularly for detergents, oil degumming, nutraceuticals food, and biodiesel fuels. These enzymes are also used in many other applications including cosmetics, agriculture, bioremediation, paper, and the leather industry. Hence, the importance of thermostability and alkaline phospholipases for different applications has grown rapidly [52]. Although specific properties of the biocatalysts may be required for each industrial application, there is considerable interest in finding novel PLA2 enzymes for different applications. In this regard, the pH, temperature activity, and stability profiles of purified WaPLA2-II were investigated (Figure 2A,B and Figure 3A,B). The data were compared to the results previously reported for WaPLA2-I.
As shown in Figure 2A, WaPLA2-II exhibited activity between pH 6.0 and 11 with an optimum pH of 9.5. Interestingly, it showed over 73% residual enzymatic activity even at pH 11 compared with only 32% for WaPLA2-I. The latter displayed its maximal activity at pH 8.5 using PC as a substrate under optimal conditions. Moreover, the pH stability profile showed that WaPLA2-II was highly stable over a large pH range, maintaining its full activity between pH 6 and 10, and more than 60% of its initial activity when incubated at pH 12 or 4 (Figure 2B). However, WaPLA2-I lost 70% of its original activity at pH 12, whereas both enzymes exhibited similar stability profiles at acidic pH.
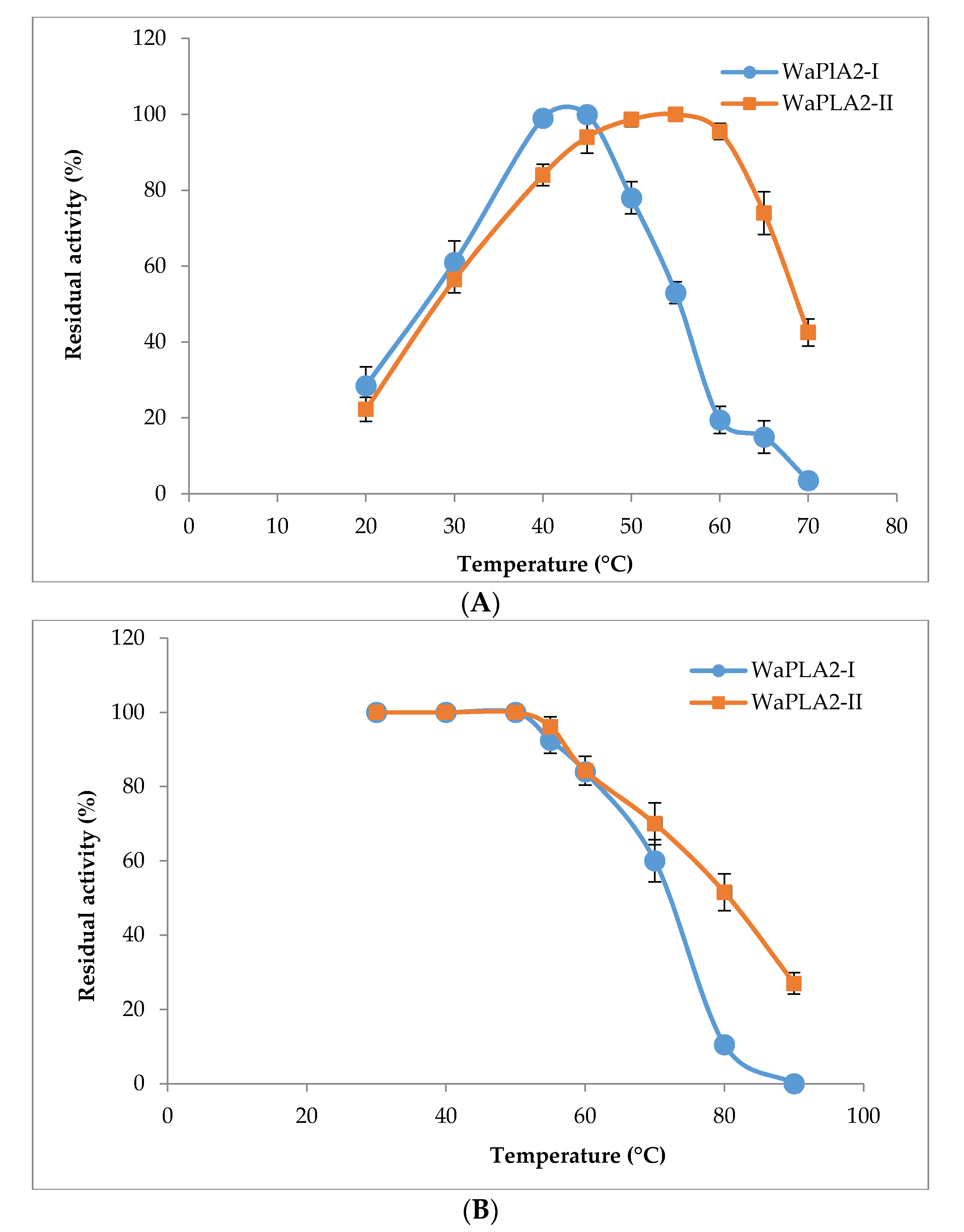
WaPLA2-I displayed maximal activity at 45 °C (Figure 3A), whereas the highest catalytic activity of WaPLA2-II was observed at 55 °C. Interestingly, it still displayed 74% and 45% of residual enzymatic activity at 65 °C and 70 °C, respectively. The results presented in Figure 3B demonstrate that WaPLA2-II is more stable against heat compared with WaPLA2-I reported previously. Even after treatment at 80 °C and 90 °C for 60 min, the protein remained active, showing 50% and 30% of its maximal activity, respectively. However, WaPLA2-I lost almost all activity when incubated under the same conditions (Figure 3B). Together, these results indicate that WaPLA2-II is an excellent enzyme for chemical and industrial processes that involve reactions at elevated temperatures. In fact, it is well established that thermophilic PLA2 displays robust activity and higher thermostability at elevated temperatures and is generally more resistant to chemical denaturation [52].
3.1.2. Calcium, Metal Ions, and Bile Salt Dependence
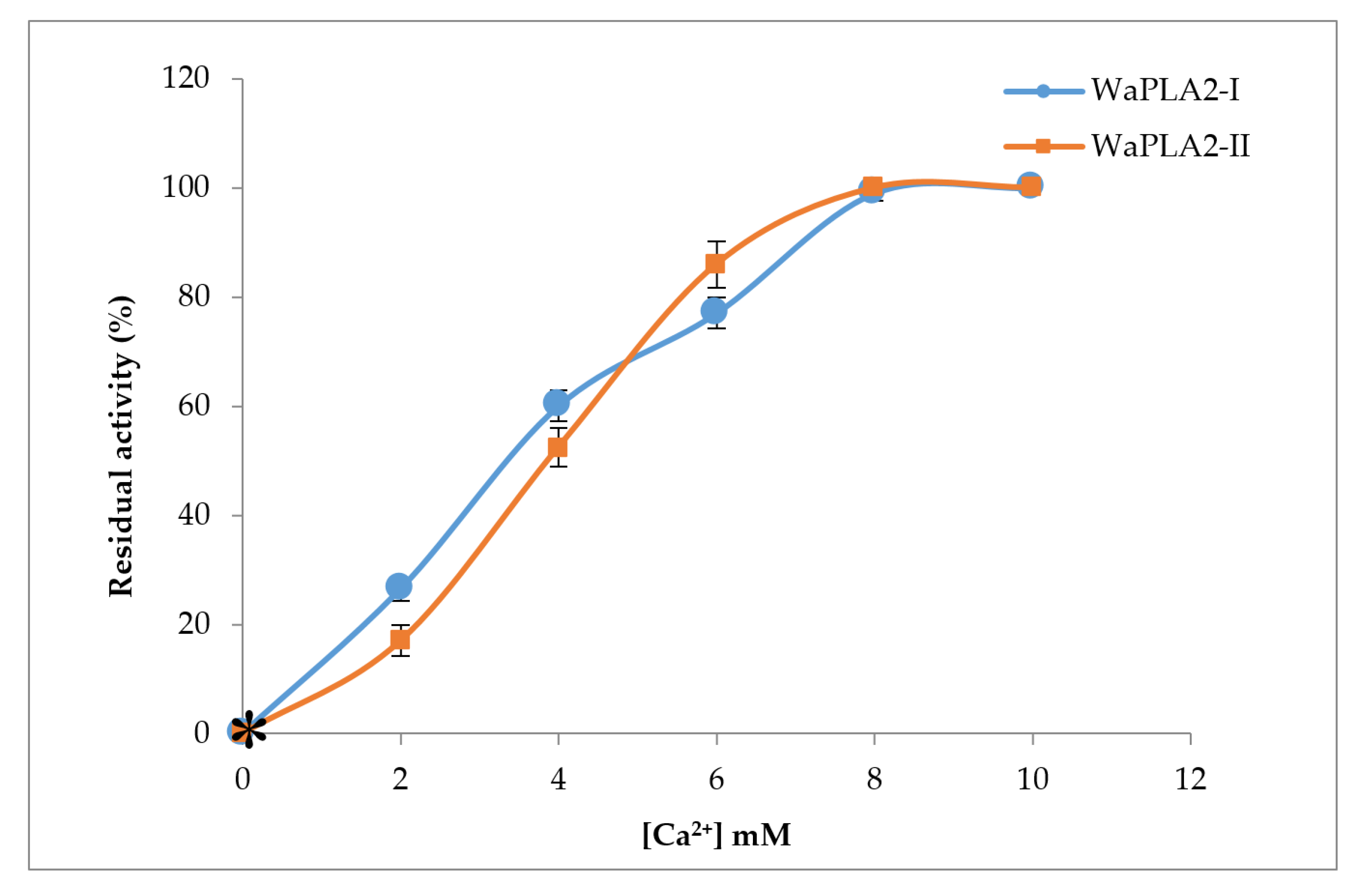
The role of Ca2+ is well known for the activity of phospholipases and their binding to substrates [53,54,55]. We investigated Ca2+ effects on WaPLA2-II activity and compared it with that obtained previously for WaPLA2-IA. We measured enzymatic activity in the presence of various concentrations of Ca2+ using PC as a substrate. As shown in Figure 4, WaPLA2-II exhibited maximal activity in the presence of 8 mM Ca2+ under optimal conditions (pH 9.5 and 55 °C). A similar result was observed with WaPLA2-I indicating that both enzymes have the same affinity for Ca2+. However, both enzymes showed a strict dependence on Ca2+ as they were completely inactive without it or when incubated with a chelator such as EDTA or EGTA. Our findings are consistent with that of previous studies of phospholipases from different organisms, thus confirming that the presence of Ca2+ optimizes the interactions between an enzyme and its substrate [56,57].
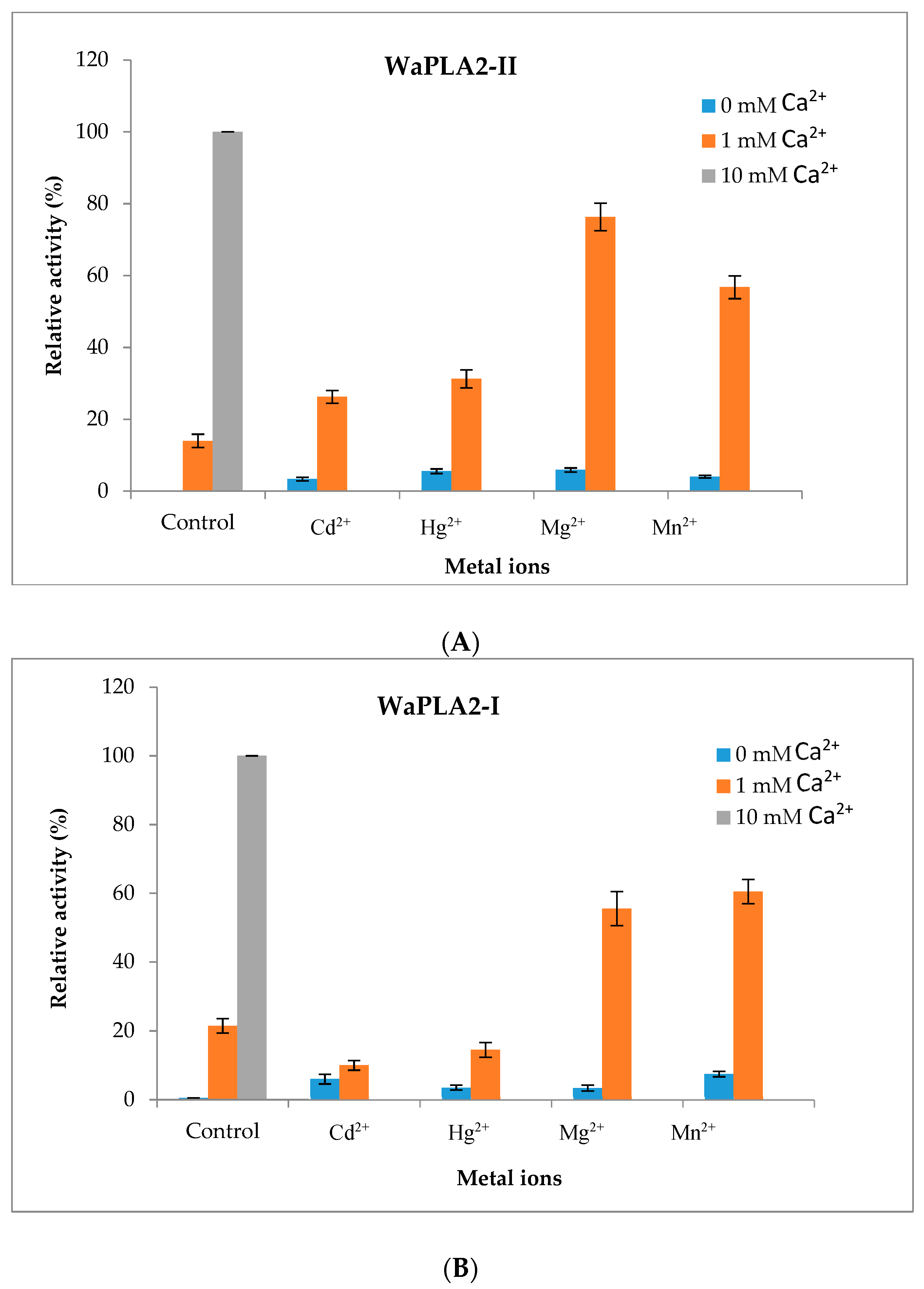
We also tested the effect of several metal ions (Mg2+, Mn2+, Cd2+ and Hg2+) at a concentration of 10 mM on the activity of WaPLA2-II using PC as substrate under optimal conditions (pH 9.5 and 55 °C) in the absence or presence of Ca2+ (1 mM). The data demonstrated that, in contrast to Ca2+, none of these ions were able to keep the substrate bound to the active enzyme (Figure 5A). However, the addition of 10 mM Mg2+ or Mn2+ significantly increased the activity of the enzyme (up to 73% and 57%, respectively), in the presence of 1 mM Ca2+, whereas Cd2+ and Hg2+ treatment resulted in only a moderate activation of WaPLA2-II (up to 26% and 31%, respectively). Using the same conditions, one can draw the same conclusion for WaPLA2-I (Figure 5B). In fact, both enzymes are significantly induced by Mg2+ and Mn2+ and less active in the presence of Cd2+ and Hg2+, which failed to competitively inhibit Ca2+-mediated activity or productively bind to the enzyme.
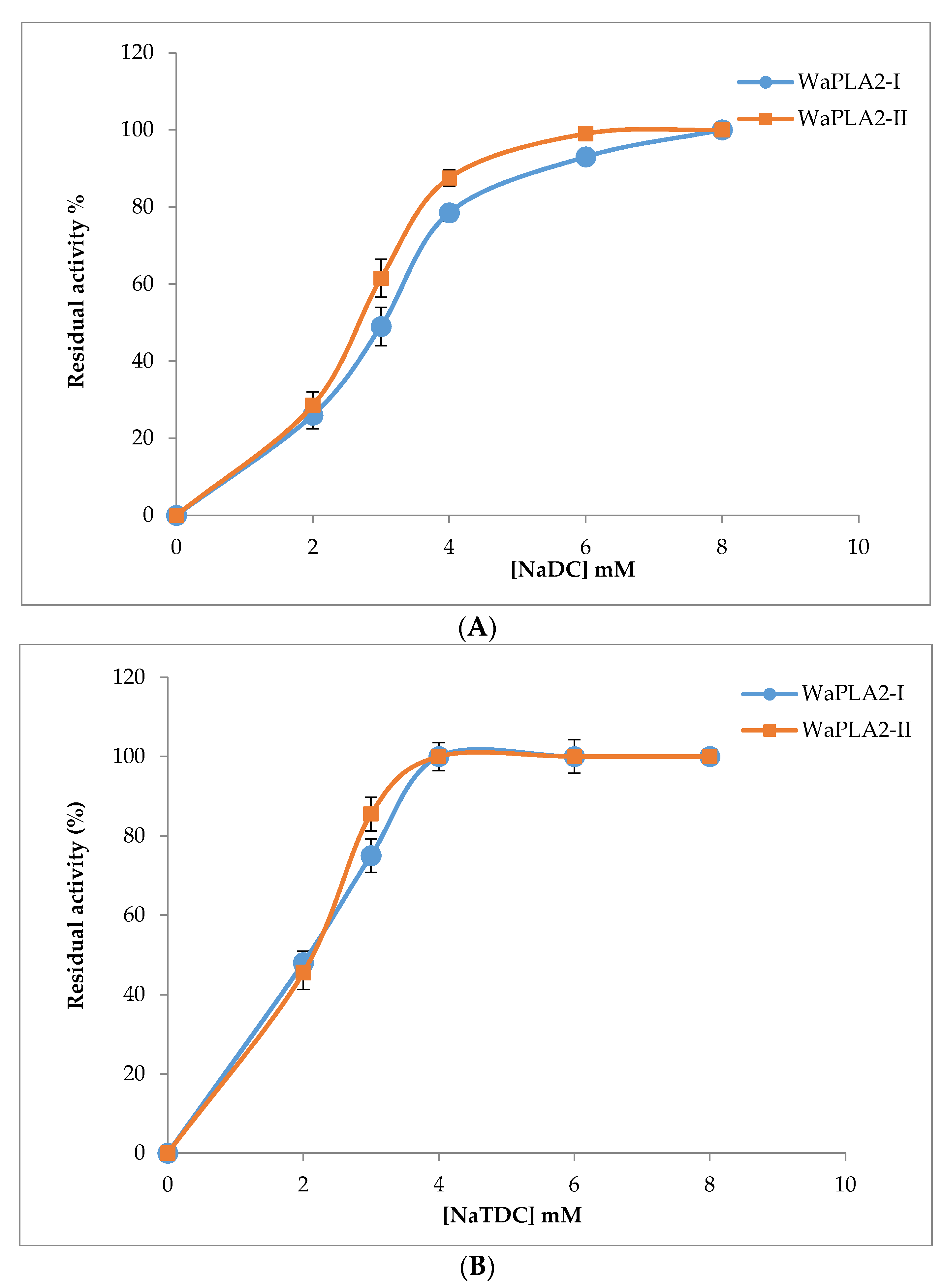
Many studies have shown that bile salts are tensioactive agents that play a key role in the dispersion of hydrolysis products [58]. Therefore, we investigated and compared the effect of bile salts on WaPLA2-II activity and compared the results with that previously reported for WaPLA2-I. As shown in Figure 6, the rate of PC hydrolysis catalyzed by the two enzymes was determined at different concentrations of bile salts (0, 2, 4, 6, and 8 mM) under optimal conditions. The data showed that NaTDC (Figure 6B) and NaDC (sodium deoxycholate) (Figure 6A) had a significant effect on the activity of both G I and G II WaPLA2s. The highest activity for WaPLA2-II was achieved in the presence of 4 mM NaTDC and 6 mM NaDC, whereas more NaDC was required for maximal WaPLA2-I activity (8 mM). These results are in agreement with that of other studies [6,8,50].
3.2. Antimicrobial and Hemolytic Activities
Since 1948, researchers have described the potent antibacterial properties of snake venoms in killing human pathogens, such as Bacillus subtilis, Escherichia coli, and Staphylococcus aureus [59]. Subsequent studies have identified and confirmed the antibiotic potential of secretions from different families of snakes. Among the numerous toxins from these natural collections, phospholipase A2, especially G IIA, are the primary biomolecules associated with antibacterial activity. They are amongst the well-studied and characterized antimicrobial venom proteins [30,60,61]. The antimicrobial effects of G II phospholipases have also been described against parasites and viruses [62,63]. Often these effects depend on the catalytic action of the enzyme. However, catalytic-independent actions or binding affinity have also been identified.
We examined the in vitro antibacterial activity of the purified G II WaPLA2 compared with G I WaPLA2 and the crude extract using an agar diffusion assay. Table 2 shows that the two enzymes were more effective against all the tested bacterial strains except P. aeruginosa compared with the crude extract. Moreover, WaPLA2-II exhibited activity against all of the Gram-positive and negative bacteria tested, producing a zone diameter of inhibition from 12.5 ± 0.7 to 30 ± 1.7 mm. The highest activity was observed against two Gram-negative bacteria [E. coli (27 ± 2.1 mm) and S. enteric (28 ± 2.4 mm)] and one Gram-positive bacteria [S. aureus (30 ± 1.7 mm)]. Significantly lower WaPLA2-II antibacterial activity was observed against P. aeruginosa, with an inhibition diameter value of 12.5 ± 0.7 mm. Notably, E. faecalis, S. epidermidis, S. aureus, E. coli, and S. Enteric microorganisms were more sensitive to WaPLA2-II than to WaPLA2-I, which produced clear zones of inhibition ranging from 25 ± 2.2 to 30 ± 1.7 mm and from 21 ± 1.5 to 27 ± 1.2 mm, respectively (Table 2). Interestingly, the anti-bacterial potential of WaPLA2-II was significantly higher than the standard, ampicillin, particularly against S. enteric (28 ± 2.4 mm and 19.5 ± 0.3 mm, respectively) and S. aureus (30 ± 1.7 mm and 21.5 ± 1.4 mm, respectively).
We also tested the antimicrobial activity of the purified WaPLA2-II by determining IC50 values using the agar dilution method. The results clearly showed that WaPLA2-II was effective against the human pathogenic bacteria with IC50 values ranging from 9 ± 0.2 to 20 ± 1 µg/mL while WaPLA2-I displayed IC50 values between 10 ± 0.3 and 17 ± 1.4 µg/mL [40] (Table 2). It is worth noting that WaPLA2-II enzyme was found to be more effective than WaPLA2-I against S. epidermidis (IC 50: 12 ± 0.7 µg/mL and 15 ± 1.1 µg/mL, respectively), E. coli (IC 50: 12 ± 0.9 µg/mL and 14 ± 1.1 µg/mL, respectively), P. aeruginosa (IC 50: 9 ± 0.2 µg/mL and 12 ± 0.7 µg/mL, respectively), and S. enteric (IC 50: 11 ± 0.5 µg/mL and 17 ± 1.4 µg/mL, respectively) (Table 2).
Our results are consistent with that of Van Hensbergen et al. [25], who demonstrated that sPLA2-IIA is unique in its particular differentiation and potency in killing Gram-positive bacteria, and it plays an undervalued role in defense against Gram-negative pathogens. However, this was somewhat surprising since the marked bactericidal activity of phospholipases G II compared with the G I enzyme is well established [30].
Regarding the antifungal activities of our enzyme, we found that the disc diameter zones of inhibition of the fungal strains ranged from 13.5 ± 1.7 to 19 ± 1.4 mm, with IC50 values ranging from 10.2 to 32.7 µg/mL. B. cinerea was the most sensitive fungi to WaPLA2-II (19 ± 1.4 mm) followed by A. niger, P. digitatum, and F. solani (15.5 ± 0.5 mm, 15.4 ± 0.7 mm and 13.5 ± 1.7 mm, respectively). Interestingly, WaPLA2-II showed a more pronounced inhibitory effect against almost all tested fungi except A. niger compared with WaPLA2-I, which displayed a zone of inhibition between 10.5 ± 0.7 to 19 ± 0.5 mm [40]. However, both enzymes exhibited lower activity compared with the fungicide, cycloheximide, which was used as a positive control.
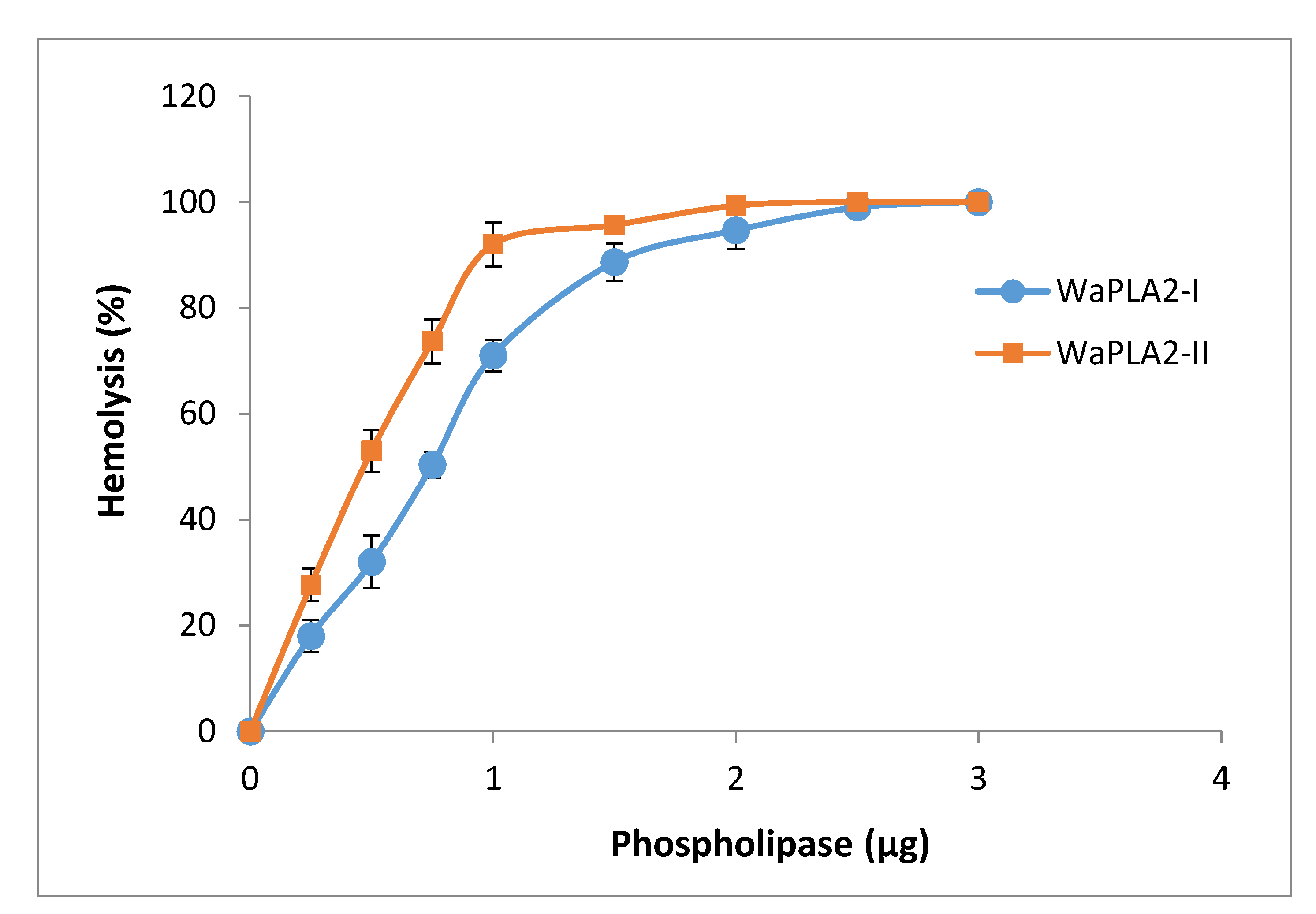
Several groups have reported that there is a strong correlation between the antibacterial and hemolytic activities of svPLA2s [30]. Indeed, PLA2 promotes phospholipids hydrolysis with liberation of lysophospholipids and free fatty acids, therefore causing indirect hemolysis. PLA2 may also block blood clotting by direct interaction with clotting factors or via degradation of phospholipids implicated in coagulation complexes, besides neurotoxicity, myotoxicity, cardiotoxicity, and antiplatelet effects. In order to support this finding, we examined the ability of WaPLA2-II to induce indirect or direct hemolysis in the presence or the absence of its substrate (PC), respectively. For the sake of comparison, the obtained data were compared to those previously reported for WaPLA2-I shown in the same figure (Figure 7). We observed that 2 µg WaPLA2-II exhibited a potent, wholly indirect hemolytic activity as evidenced by a dose-dependent 100% hemolysis of blood cells, whereas WaPLA2-I had the same effect albeit at a higher dose (3 µg) (Figure 7). Our results are in line with that of other studies in which VRV-PL-V and VRV-PL-VIII, snake venom phospholipases from Daboia russellii pulchella showed a significant correlation between their antibacterial and hemolytic activities at doses of 6 µg and 5 µg, respectively [64,65]. In fact, the correlation between these activities clearly indicates that the catalytic activity of PLA2 is primarily involved in antibacterial activities [60,61,66]. Furthermore, pre-incubation of the pure WaPLA2-II with p-BPB (40 mM in acetone), an inhibitor of svPLA2 enzymatic activity caused a significant decrease in its antimicrobial activity (Table 2). This suggests that WaPLA2-mediated antimicrobial activity is dependent upon its catalytic activity, which most likely occurs through the enzymatic membrane degradation effect [28,59].
3.3. Cytotoxicity
In addition to the antimicrobial properties of svPLA2, the toxicity of some of the peptides is an important issue that must be considered. In fact, there is a growing evidence that phospholipase A2 plays a significant role in human gastrointestinal cancer [35]. Antitumoral activity was first described for an acidic PLA2 isolated from Bothrops jararacussu venom [67]. Bazaa and co-workers observed that a G II phospholipase A2 (MVL-PLA2) from snake venom of Macrovipera lebetina transmediterranea inhibited cell adhesion and migration of various human tumor cells [68]. They purified and characterized two other phospholipase A2 enzymes from Cerastes cerastes venom, which also inhibited cancer cell adhesion and migration [50].
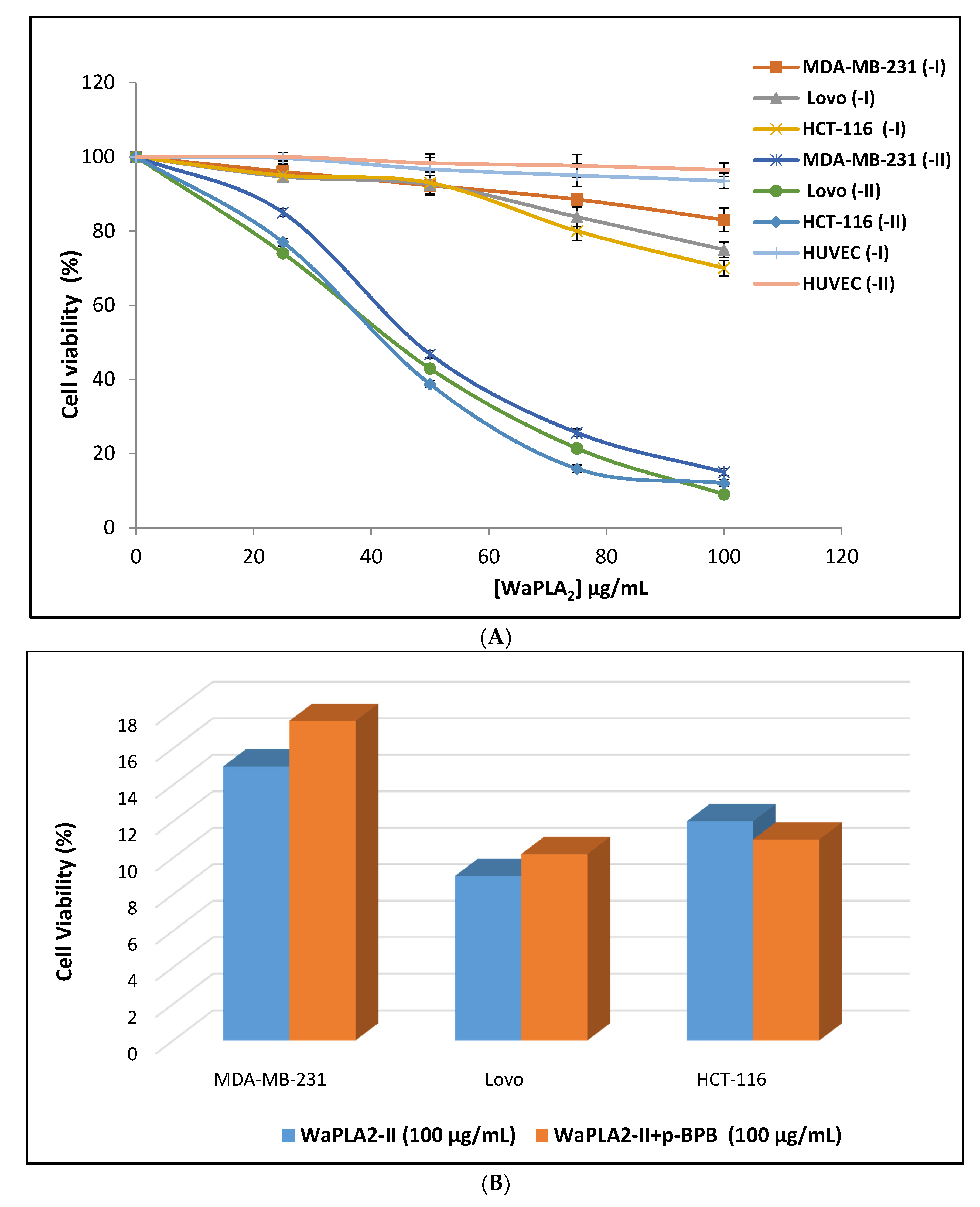
Despite these preliminary studies, little is known with respect to the cytotoxicity of svPLA2 and its mechanism of action on cancer cells. We measured the cytotoxic effects of pure WaPLA2-II with p-BPB on three human cell lines (MDA-MB-231, HCT-116, and HCT) using the MTT assay and compared the results with that observed with WaPLA2-I [40]. Cell lines were cultured with different PLA2s at concentrations ranging from 0 to 100 µg/mL. As shown in Figure 8, the percentage of viable cells in the medium containing sterile saline or WaPLA2-II after 24 h of treatment was approximately 80% at an enzyme concentration of 25 µg/mL. The rate of viable cells significantly decreased at 50 µg/mL of enzyme and reached 10% at 100 µg/mL. In contrast with WaPLA2-I, which was not cytotoxic at 50 µg/mL, the percentage of cell viability was approximately 70–80% depending on the cell line, even at 100 µg/mL. The effect exerted by WaPLA2-II on human cancer cell lines was similar to treatment with 0.1% Triton X-100 (Figure 8A). It is worth noting that both WaPLA2-I and WaPLA2-II did not show any effect on the viability of human umbilical vein endothelial cells after 24 h, at concentrations ranging from 0 to 100 µg/mL, using the MTT assay (Figure 8A). This study indicates that WaPLA2-II presents very low toxicity in normal human cell lines (HUVEC) but affects several human cancer cell lines: HCT-116, MDA-MB-231 and Lovo. The obtained results here are a very initial indication that WaPLA2-II could have anti-carcinogenic capabilities given its effects on the proliferation of cancer cell lines, but further studies are needed to confirm such a result.
Interestingly, at 100 µg/mL, catalytically inactivated WaPLA2-II was still able to cause significant cytotoxic effects on three different human cancer cell lines (MDAMB-231, HCT-116, and LoVo) at the same level as the active enzyme (Figure 8B). These findings indicate a separate antitumor activity of WaPLA2-II from its catalytic activity, which is supported by the previous study of [68]. An oncogenic role was also proposed by Ozturk et al. [24], who found that human G IIA PLA2 was overexpressed during the progression of prostate cancer. However, further studies are needed to determine a role in cancer etiology and therapy.
4. Conclusions
In conclusion, we have purified and characterized a novel PLA2 G II from desert cobra venom of W. aegyptia in Saudi Arabia. This enzyme exhibits several desirable properties such as high stability in acidic and alkaline pH media, and at high temperatures. Additionally, it exerts potent cytotoxic and antibacterial activities. WaPLA2-II also exhibits antitumor activity, which may serve as starting point for more studies leading to the design of a new generation of anticancer drugs.
Author Contributions
Conceptualization, A.B.B. and M.A.; methodology, M.A. and I.J.; formal analysis, I.A. and A.B.B.; investigation, I.A. and M.A.; writing—original draft preparation, I.A.; writing—review and editing, I.A. and I.J.; visualization, A.B.B.; supervision, A.B.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors extend their appreciation to the Deanship of Scientific Research at King Saud University for funding this work through research group no. RG-1441-471.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Casale, J.; Kacimi, S.E.O.; Varacallo, M. Biochemistry, Phospholipase A2. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 biochemistry. Cardiovasc. Drugs Ther. 2009, 23, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Burke, J.E.; Dennis, E.A. Phospholipase A2 structure/function, mechanism, and signaling. J. Lipid Res. 2009, 50, S237–S242. [Google Scholar] [CrossRef] [Green Version]
- Six, D.A.; Dennis, E.A. The expanding superfamily of phospholipase A2 enzymes: Classification and characterization. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1488, 1–19. [Google Scholar] [CrossRef]
- Lambeau, G.; Gelb, M.H. Biochemistry and physiology of mammalian secreted phospholipases A2. Annu. Rev. Biochem. 2008, 77, 495–520. [Google Scholar] [CrossRef] [Green Version]
- Bacha, A.B.; Abid, I.; Horchani, H. Antibacterial properties of intestinal phospholipase A2 from the common stingray Dasyatis pastinaca. Appl. Biochem. Biotechnol. 2012, 168, 277–1287. [Google Scholar] [CrossRef]
- Bacha, A.B.; Abid, I. Secretory phospholipase A2 in dromedary tears: A host defense against staphylococci and other gram-positive bacteria. Appl. Biochem. Biotechnol. 2013, 169, 1858–1869. [Google Scholar] [CrossRef]
- Bacha, A.B.; Daihan, S.K.; Moubayed, N.M. Purification and characterization of a phospholipase A2-IIA from common stingray (Dasyatis pastinaca) intestine. Indian J. Biochem. Biophys. 2013, 50, 186–195. [Google Scholar]
- Murakami, M.; Taketomi, Y.; Girard, C. Emerging roles of secreted phospholipase A2 enzymes: Lessons from transgenic and knockout mice. Biochimie 2010, 92, 561–582. [Google Scholar] [CrossRef]
- Murakami, M.; Taketomi, Y.; Miki, Y. Recent progress in phospholipase A2 research: From cells to animals to humans. Prog. Lipid Res. 2011, 50, 152–192. [Google Scholar] [CrossRef]
- Hariprasad, G.; Srinivasan, A.; Singh, R. Structural and phylogenetic basis for the classification of group III phospholipase A2. J. Mol. Model. 2013, 19, 3779–3791. [Google Scholar] [CrossRef]
- Etienne, D.; Boilard, E. Roles of secreted phospholipase A2 group IIA in inflammation and host defense (BBA). Mol. Cell Biol. Lipids 2019, 1864, 789–802. [Google Scholar]
- Herrera, M.; Fernández, J.; Vargas, M. Comparative proteomic analysis of the venom of the taipan snake, Oxyuranus scutellatus, from Papua New Guinea and Australia: Role of neurotoxic and procoagulant effects in venom toxicity. J. Proteom. 2012, 75, 2128–2140. [Google Scholar] [CrossRef] [PubMed]
- Gulati, A.; Isbister, G.K.; Duffull, S.B. Effect of Australian elapid venoms on blood coagulation: Australian Snakebite Project (ASP-17). Toxicon 2013, 61, 94–104. [Google Scholar] [CrossRef]
- Lewin, M.; Samuel, S.; Merkel, J. Varespladib (LY315920) appears to be a potent, broad-spectrum, inhibitor of snake venom phospholipase A2 and a possible pre-referral treatment for envenomation. Toxins 2016, 8, 248. [Google Scholar] [CrossRef] [Green Version]
- Heinrikson, R.L.; Krueger, E.T.; Keim, P.S. Amino acid sequence of phospholipase A2-alpha from the venom of Crotalus adamanteus. A new classification of phospholipases A2 based upon structural determinants. J. Biol. Chem. 1977, 252, 4913–4921. [Google Scholar]
- Valentin, E.; Lambeau, G. What can venom phospholipases A2 tell us about the functional diversity of mammalian secreted phospholipases A2. Biochimie 2000, 82, 815–831. [Google Scholar] [CrossRef]
- Schaloske, R.H.; Dennis, E.A. The phospholipase A2 superfamily and its group numbering system. Biochim. Biophys. Acta 2006, 1761, 1246–1259. [Google Scholar] [CrossRef]
- Dennis, E.A.; Cao, J.; Hsu, Y.H. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [Green Version]
- Krayem, N.; Gargouri, Y. Scorpion venom phospholipases A2: A minireview. Toxicon 2020, 184, 48–54. [Google Scholar] [CrossRef]
- Teixeira, C.F.P.; Landucci, E.C.T.; Antunes, E. Inflammatory effects of snake venom myotoxic phospholipases A2. Toxicon 2003, 42, 947–962. [Google Scholar] [CrossRef]
- Costa, S.K.P.; Camargo, E.A.; Antunes, E. Inflammatory action of secretory PLA2 from snake venoms. In Toxins and Drug Discovery; Springer: Dordrecht, The Netherlands, 2015; pp. 1–18. [Google Scholar]
- Zambelli, V.O.; Picolo, G.; Fernandes, C.A.H. Secreted phospholipases A2 from animal venoms in pain and analgesia. Toxins 2017, 9, 406. [Google Scholar] [CrossRef] [Green Version]
- Ozturk, K.; Onala, M.S.; Efiloglu, O. Association of 5′UTR polymorphism of secretory phospholipase A2 group IIA (PLA2G2A) gene with prostate cancer metastasis. Gene 2020, 742, 144586. [Google Scholar] [CrossRef]
- Van Hensbergen, V.P.; Wu, Y.; Van Sorge, N.M.; Touqui, L. Type IIA Secreted phospholipase A2 in Host Defense against Bacterial Infections. Trend Immunol. 2020, 41, 313–326. [Google Scholar] [CrossRef]
- Moreira, V.; Gutiérrez, J.M.; Lomonte, B. 12-HETE is a regulator of PGE2 production via COX-2 expression induced by a snake venom group IIA phospholipase A2 in isolated peritoneal macrophages. Chem. Biol. Interact. 2020, 317, 108903. [Google Scholar] [CrossRef]
- Buckland, A.G.; Heeley, E.L.; Wilton, D.C. Bacterial cell membrane hydrolysis by secreted phospholipases A2: A major physiological role of human group IIa sPLA2 involving both bacterial cell wall penetration and interfacial catalysis. Biochim. Biophys. Acta 2000, 1484, 195–206. [Google Scholar] [CrossRef]
- Buckland, A.G.; Wilton, D.C. The antibacterial properties of secreted phospholipases A2. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2000, 1488, 71–82. [Google Scholar] [CrossRef]
- Nevalainen, T.J.; Graham, G.G.; Scott, K.F. Antibacterial actions of secreted phospholipases A2. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2008, 1781, 1–9. [Google Scholar] [CrossRef]
- Almeida, R.; Palacios, A.L.V.; Patiño, R.S.P. Harnessing snake venom phospholipasesA2 to novel approaches for overcoming antibiotic resistance. Drug Dev. Res. 2019, 80, 68–85. [Google Scholar] [CrossRef] [Green Version]
- Cecilio, A.B.; Caldas, S.; Oliveira, R.A. Molecular characterization of Lys49 and Asp49 phospholipases A2 from snake venom and their antiviral activities against dengue virus. Toxins 2013, 5, 1780–1798. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, V.M.; Marcussi, S.; Cambraia, R.S. Bactericidal and neurotoxic activities of two myotoxic phospholipases A2 from Bothrops neuwiedi pauloensis snake venom. Toxicon 2004, 44, 305–314. [Google Scholar] [CrossRef]
- Sobrinho, J.C.; Kayano, A.M.; Simões-Silva, R. Anti-platelet aggregation activity of two novel acidic Asp49-phospholipases A2 from Bothrops brazili snake venom. Int. J. Biol. Macromol. 2018, 107, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Bustillo, S.; Fernández, J.; Chavez, S. Comparative characterization and synergism between two basic phospholipases A2 from Bothrops diporus snake venom. Toxicon 2020, 177, S44–S45. [Google Scholar] [CrossRef]
- Wang, X.; Huang, C.-J.; Yu, G.-Z.; Wang, J.-J.; Wang, R.; Li, Y.-M.; Wu, Q. Expression of group IIA phospholipase A2 is an independent predictor of favorable outcome for patients with gastric cancer. Hum. Pathol. 2013, 44, 2020–2027. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Díaz, C.; Chaves, F. Comparative characterization of Viperidae snake venoms from Perú reveals two compositional patterns of phospholipase A2 expression. Toxicon 2020, 7, 100044. [Google Scholar] [CrossRef]
- Al-Sadoon, M.K. Snake bite envenomation in Riyadh province of Saudi Arabia over the period (2005–2010). Saudi J. Biol. Sci. 2015, 22, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Ismail, M.; Memish, Z.A. Venomous snakes of Saudi Arabia and the Middle East: A keynote for travelers. Int. J. Antimicrob. Agents 2003, 21, 164–169. [Google Scholar] [CrossRef]
- Al-Sadoon, M.K.; Paray, B.A.; Al-Otaibi, H. Survey of the reptilian fauna of the Kingdom of Saudi Arabia. VI. The snake fauna of Turaif region. Saudi J. Biol. Sci. 2017, 24, 925–928. [Google Scholar] [CrossRef]
- Bacha, A.B.; Alonazi, M.A.; Elshikh, M.S.; Karray, A. A novel bactericidal homodimeric PLA2 Group-I from Walterinnesia aegyptia venom. Int. J. Biol. Macromol. 2018, 117, 1140–1146. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and sensitive method for the quantization of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hewick, R.M.; Hunkapiller, M.W.; Hood, L.E. A gas-liquid solid phase peptide and protein sequenator. J. Biol. Chem. 1981, 256, 7990–7997. [Google Scholar]
- Spiro, R.G. Analysis of sugars found in glycoproteins. Meth. Enzymol. 1966, 8, 3–26. [Google Scholar]
- Condrea, E.; Fletcher, J.E.; Rapuano, B.E. Effect of modification of one histidine residue on the enzymatic and pharmacological properties of a toxic phospholipase A2 from Naja nigricollis snake venom and less toxic phospholipases A2 from Hemachatus haemachatus and Naja naja atra snake venoms. Toxicon 1981, 19, 61–71. [Google Scholar] [CrossRef]
- Abousalham, A.; Verger, R. Egg yolk lipoproteins as substrates for lipases. Biochim. Biophys. Acta 2000, 1485, 56–62. [Google Scholar] [CrossRef]
- Boman, H.G.; Kaletta, U. Chromatography of rattle snake venom A separation of three phosphodiesterase. Biochim. Biophys. Acta 1957, 24, 619–631. [Google Scholar] [CrossRef]
- Vanden Berghe, D.A.; Vlietinck, A.J. Screening methods for anti-bacterial and antiviral agents from higher plants. Meth. Plant Biochem. 1991, 6, 47–68. [Google Scholar]
- Ronald, M.A. Microbiologica; Compania Editorial Continental, S.A. de C.V.: Mexico City, Mexico, 1990; pp. 505–525. [Google Scholar]
- Xu, C.; Ma, D.; Yu, H. A bactericidal homodimeric phospholipases A2 from Bungarus fasciatus venom. Peptides 2007, 28, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Zouari-Kessentini, R.; Luis, J.; Karray, A.; Kallech-Ziri, O.; Srairi-Abid, O.; Bazaa, A.; Loret, E.; Bezzine, S.; El Ayeb, M.; Marrakchi, N. Two purified and characterized phospholipases A2 from Cerastes cerastes venom, that inhibit cancerous cell adhesion and migration. Toxicon 2009, 53, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.-H.; Wang, Y.-M.; Cheng, A.-C.; Starkov, V.; Osipov, A.; Nikitin, I.; Makarova, Y.; Ziganshin, R.; Utkin, Y. cDNA cloning, structural, and functional analyses of venom phospholipases A₂ and a Kunitz-type protease inhibitor from steppe viper Vipera ursinii renardi. Toxicon 2011, 57, 332–341. [Google Scholar] [CrossRef] [PubMed]
- Rigoldi, F.; Donini, S.; Redaelli, A.; Parisini, E.; Gautieri, A. Engineering of thermostable enzymes for industrial applications featured. APL Bioeng. 2018, 2, 011501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleer, E.A.; Puijk, W.C.; Slotboom, A.J. Modification of arginine residues in porcine pancreatic phospholipase A2. Eur. J. Biochem. 1981, 116, 277–284. [Google Scholar] [CrossRef]
- Scott, D.L.; Otwinowski, Z.; Gelb, M.H. Crystal structure of bee-venom phospholipase A2 in a complex with a transition-state analogue. Science 1990, 250, 1563–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verger, R.; Mieras, M.C.; De Haas, G.H. Action of phospholipase A at interfaces. J. Biol. Chem. 1973, 248, 4023–4034. [Google Scholar] [PubMed]
- Yu, B.Z.; Berg, O.G.; Jain, M.K. The divalent cation is obligatory for the binding of ligands to the catalytic site of secreted phospholipase A2. Biochemistry 1993, 32, 6485–6492. [Google Scholar] [CrossRef] [PubMed]
- Janssen, M.J.W.; Vermeulen, L.; Van der Helm, H.A.; Aarsman, A.J.; Slotboom, A.J.; Egmond, M.R. Enzymatic properties of rat group IIA and V phospholipases A2 compared. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1999, 1440, 59–72. [Google Scholar] [CrossRef]
- De Haas, G.H.; Slotboom, A.J.; Bonsen, P.P. Studies on phospholipase A and its zymogen from porcine pancreas. II. The assignment of the position of the six disulfide bridges. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1970, 221, 54–61. [Google Scholar] [CrossRef]
- Glaser, H.S.R. Bactericidal activity of Crotalus venom in vitro. Copeia 1948, 4, 245–247. [Google Scholar] [CrossRef]
- Perumal Samy, R.; Gopalakrishnakone, P.; Thwin, M.M.; Chow, T.K.V.; Bow, H.; Yap, E.H.; Thong, T.W.J. Antibacterial activity of snake, scorpion and bee venoms: A comparison with purified venom phospholipase A2 enzymes. J. Appl. Microbiol. 2007, 102, 650–659. [Google Scholar] [CrossRef]
- Samy, R.P.; Manikandan, J.; Sethi, G. Snake venom proteins: Development into antimicrobial and wound healing agents. Org. Chem. 2014, 11, 4–14. [Google Scholar] [CrossRef]
- Russo, R.R.; DosSantos, J.; Cintra, A.C.O. Purification and virucidal activity of two recombinant isoforms of phospholipase A2 from Crotalus durissus terrificus venom. Arch. Virol. 2019, 164, 1159–1171. [Google Scholar] [CrossRef]
- Hebleen, B.; Loría, G.D.; Lomonte, B. Potent virucidal activity against Flaviviridae of a group IIA phospholipase A2 isolated from the venom of Bothrops asper. Biologicals 2020, 63, 48–52. [Google Scholar]
- Sudarshan, S.; Dhananjaya, B.L. Antibacterial potential of a basic phospholipase A2 (VRV-PL-V) of Daboia russellii pulchella (Russell’s viper) venom. Biochem. Mosc. 2014, 79, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Sudharshan, S.; Dhananjaya, B.L. Antibacterial potential of a basic phospholipase A2 (VRV-PL-VIIIa) from Daboia russelii Pulchella (Russell’s viper) venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Oliveira Junior, N.G.; Silva Cardoso, M.H.; Franco, O.L. Snake venoms: Attractive antimicrobial proteinaceous compounds for therapeutic purposes. Cell. Mol. Life Sci. 2013, 70, 4645–4658. [Google Scholar] [CrossRef] [PubMed]
- Roberto, P.G.; Kashima, S.; Marcussi, S. Cloning and identification of a complete cDNA coding for a bactericidal and antitumoral acidic phospholipase A2 from Bothrops jararacussu venom. Protein J. 2004, 23, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Bazaa, A.; Luis, J.; Srairi-Abid, N.; Kallech-Ziri, O.; Kessentini-Zouari, R.; Defilles, C.; Lissitzky, J.C.; ElAyeb, M.; Marrakchi, N. MVL-PLA2, a phospholipase A2 from Macrovipera lebetina transmediterranea venom, inhibits tumor cells adhesion and migration. Matrix Biol. 2009, 28, 188–193. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chromatography on RP-HPLC using a C18 column (A) and SDS-PAGE (15%) (B) of the purified WaPLA2-II. Lane 1, molecular mass markers (Enolase 100 kDa, Carbonic anhydrase 30 Kda, Cytochrome C ~ 10 kDa, and Angiotensin 1 kDa), lanes 2 and 3, 20 μg of purified WaPLA2-II eluted from the RP-HPLC column under reducing conditions. AU: arbitrary units. The arrows designate peaks showing the highest phospholipase A2 (PLA2) and antibacterial activities. The blue line indicates the linear acetonitrile gradient (C) filtration of WaPLA2-II (peak 4) on the Bio-sil SEC-125 (300 mm × 7.8 mm) high performance liquid chromatography (HPLC) column. The column was equilibrated in 0.1 M phosphate buffer, pH 7, containing 0.15 M NaCl, and elution was achieved using the same buffer at a flow rate of 1 mL/min for 20 min and at room temperature. The eluate was monitored at 280 nm. AU: arbitrary units. Elution profiles of WaPLA2-II and molecular mass markers are shown in black and red, respectively. (D) N-terminal sequence alignment of WaPLA2-II from W. aegyptia with sPLA2 G II and G I from various snake venoms (Cerastes cerastes, Echis carinatus, Vipera renardi, Atheris nitschei, and Bungarus fasciatus). Red, blue, and black denote identical, similar, and different amino acids, respectively.
Figure 1.
Chromatography on RP-HPLC using a C18 column (A) and SDS-PAGE (15%) (B) of the purified WaPLA2-II. Lane 1, molecular mass markers (Enolase 100 kDa, Carbonic anhydrase 30 Kda, Cytochrome C ~ 10 kDa, and Angiotensin 1 kDa), lanes 2 and 3, 20 μg of purified WaPLA2-II eluted from the RP-HPLC column under reducing conditions. AU: arbitrary units. The arrows designate peaks showing the highest phospholipase A2 (PLA2) and antibacterial activities. The blue line indicates the linear acetonitrile gradient (C) filtration of WaPLA2-II (peak 4) on the Bio-sil SEC-125 (300 mm × 7.8 mm) high performance liquid chromatography (HPLC) column. The column was equilibrated in 0.1 M phosphate buffer, pH 7, containing 0.15 M NaCl, and elution was achieved using the same buffer at a flow rate of 1 mL/min for 20 min and at room temperature. The eluate was monitored at 280 nm. AU: arbitrary units. Elution profiles of WaPLA2-II and molecular mass markers are shown in black and red, respectively. (D) N-terminal sequence alignment of WaPLA2-II from W. aegyptia with sPLA2 G II and G I from various snake venoms (Cerastes cerastes, Echis carinatus, Vipera renardi, Atheris nitschei, and Bungarus fasciatus). Red, blue, and black denote identical, similar, and different amino acids, respectively.

Figure 2.
The pH profile (A) and pH stability (B) of the purified WaPLA2-II and -I from the venom of W. aegyptia sPLA2 activity was assayed in a pH range from 6.0 to 11.0 at optimal temperature (60 °C). The maximum activity of WaPLA2-II obtained at pH 9.5 was designated as 100% activity. The pH stability of the two enzymes was determined by incubating the enzymes in different buffers for 30 min at room temperature and at different pH values (3–12), and the residual enzyme activity was determined under standard assay conditions. The control represents 100% of PLA2 activity at 10 mM Ca2+ under the same conditions. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).
Figure 2.
The pH profile (A) and pH stability (B) of the purified WaPLA2-II and -I from the venom of W. aegyptia sPLA2 activity was assayed in a pH range from 6.0 to 11.0 at optimal temperature (60 °C). The maximum activity of WaPLA2-II obtained at pH 9.5 was designated as 100% activity. The pH stability of the two enzymes was determined by incubating the enzymes in different buffers for 30 min at room temperature and at different pH values (3–12), and the residual enzyme activity was determined under standard assay conditions. The control represents 100% of PLA2 activity at 10 mM Ca2+ under the same conditions. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).

Figure 3.
Temperature profile (A) and thermal stability (B) of the purified WaPLA2-II and -I from the venom of W. aegyptia. Enzyme activity was measured at different temperatures ranging from 20 to 70 °C at pH 9.5, using phosphatidylcholine (PC) as a substrate in the presence of 10 mM Ca2+ and 4 mM NaTDC (sodium taurodeoxycholate). The activity of the WaPLA2-II at 55 °C was considered as 100%. For thermal stability, the enzymes were incubated at the indicated temperatures (30–90 °C) for 60 min. Residual enzyme activity was assayed under standard assay conditions. The control represents 100% of PLA2 activity at 10 mM Ca2+ under the same conditions. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).
Figure 3.
Temperature profile (A) and thermal stability (B) of the purified WaPLA2-II and -I from the venom of W. aegyptia. Enzyme activity was measured at different temperatures ranging from 20 to 70 °C at pH 9.5, using phosphatidylcholine (PC) as a substrate in the presence of 10 mM Ca2+ and 4 mM NaTDC (sodium taurodeoxycholate). The activity of the WaPLA2-II at 55 °C was considered as 100%. For thermal stability, the enzymes were incubated at the indicated temperatures (30–90 °C) for 60 min. Residual enzyme activity was assayed under standard assay conditions. The control represents 100% of PLA2 activity at 10 mM Ca2+ under the same conditions. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).

Figure 4.
The effect of Ca2+ concentrations on purified WaPLA2-II and -I activities. Enzyme activity was measured at various concentrations of Ca2+ (0–10 mM) using PC as the substrate at pH 9.5 and 55 °C for WaPLA2-II, and at 45 °C and pH 8.5 for WaPLA2-IA in the presence of 4 mM NaTDC. The absence of phospholipase activity checked in the presence of 10 mM EGTA/EDTA and in the absence of CaCl2 is indicated by a star symbol. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).
Figure 4.
The effect of Ca2+ concentrations on purified WaPLA2-II and -I activities. Enzyme activity was measured at various concentrations of Ca2+ (0–10 mM) using PC as the substrate at pH 9.5 and 55 °C for WaPLA2-II, and at 45 °C and pH 8.5 for WaPLA2-IA in the presence of 4 mM NaTDC. The absence of phospholipase activity checked in the presence of 10 mM EGTA/EDTA and in the absence of CaCl2 is indicated by a star symbol. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).

Figure 5.
The effects of various metal ions on the activity of purified WaPLA2-II (A) and WaPLA2-I (B). The influence of metal ions (10 mM each) on the activity of each enzyme, in the absence or presence of 1 mM Ca2+, was studied in the presence of 4 mM NaTDC using PC emulsion as a substrate at 55 °C and pH 9.5 for WaPLA2-II and 45 °C and pH 8.5 for WaPLA2-IA. The control represents 100% of sPLA2 activity at 8 mM Ca2+ under the same conditions. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).
Figure 5.
The effects of various metal ions on the activity of purified WaPLA2-II (A) and WaPLA2-I (B). The influence of metal ions (10 mM each) on the activity of each enzyme, in the absence or presence of 1 mM Ca2+, was studied in the presence of 4 mM NaTDC using PC emulsion as a substrate at 55 °C and pH 9.5 for WaPLA2-II and 45 °C and pH 8.5 for WaPLA2-IA. The control represents 100% of sPLA2 activity at 8 mM Ca2+ under the same conditions. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).

Figure 6.
The effect of increasing concentrations of NaDC (A) and NaTDC (B) on WaPLA2-II and -I activities. sPLA2 activity was measured using PC as a substrate at pH 9.5 and 55 °C for WaPLA2-II, and at 45 °C and pH 8.5 for WaPLA2-IA, in the presence of 10 mM Ca2+. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).
Figure 6.
The effect of increasing concentrations of NaDC (A) and NaTDC (B) on WaPLA2-II and -I activities. sPLA2 activity was measured using PC as a substrate at pH 9.5 and 55 °C for WaPLA2-II, and at 45 °C and pH 8.5 for WaPLA2-IA, in the presence of 10 mM Ca2+. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).

Figure 7.
Dose-dependent indirect hemolytic activity of WaPLA2-II and -I. The two purified enzymes (0–3 μg) were incubated at 37 °C for 10 min with PC, RBCs, and PBS (1:1:8 v/v) to release hemoglobin. Free hemoglobin was then determined by measuring the absorbance at 540 nm. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).
Figure 7.
Dose-dependent indirect hemolytic activity of WaPLA2-II and -I. The two purified enzymes (0–3 μg) were incubated at 37 °C for 10 min with PC, RBCs, and PBS (1:1:8 v/v) to release hemoglobin. Free hemoglobin was then determined by measuring the absorbance at 540 nm. The results represent the means of three independent experiments and expressed as the mean ± SD (n = 3).

Figure 8.
Cytotoxic effect of WaPLA2-II and -I on LoVo, HCT-116, and MDA-MB-231 cells. To determine the cell viability, cells were maintained in culture for 24 h with [0–100 µg/mL] concentrations of both enzymes using MTT solution. Cytotoxicity was assessed using the MTT assay by incubating cells for 24 h with various concentrations (25, 50, 75, and 100 μg) of each enzyme (A) or with WaPLA2-II (100 μg/mL) with or without p-BPB (B). Triton X-100 (0.1%) in the assay medium and the assay medium alone served as positive and negative controls, respectively. The results represent the means of three independent experiments. Cell viability was calculated as the mean ± SD (n = 3).
Figure 8.
Cytotoxic effect of WaPLA2-II and -I on LoVo, HCT-116, and MDA-MB-231 cells. To determine the cell viability, cells were maintained in culture for 24 h with [0–100 µg/mL] concentrations of both enzymes using MTT solution. Cytotoxicity was assessed using the MTT assay by incubating cells for 24 h with various concentrations (25, 50, 75, and 100 μg) of each enzyme (A) or with WaPLA2-II (100 μg/mL) with or without p-BPB (B). Triton X-100 (0.1%) in the assay medium and the assay medium alone served as positive and negative controls, respectively. The results represent the means of three independent experiments. Cell viability was calculated as the mean ± SD (n = 3).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characterization of WaPLA2-II at each purification step.
| Purification Step | Total Activity 1 (U) | Protein 2 (mg) | Specific Activity (U/mg) | Activity Recovery (%) | Purification Factor |
|---|---|---|---|---|---|
| Extraction | 8000 | 160 | 50 | 100 | 1 |
| Ammonium sulfate fractionation (65%) | 6400 | 61 | 104.9 | 80 | 2.1 |
| Heat treatment (70 °C, 30 min) | 5056 | 9.7 | 521.2 | 63.2 | 10.4 |
| C18 RP-HPLC | 950 | 0.76 | 1250 | 11.9 | 25 |
1 1 μmole of fatty acid liberated under assay conditions corresponds to one unit of phospholipase activity. 2 Protein concentration was measured using the Bradford method [41].
Table 2.
Antimicrobial activities of the crude extract and purified WaPLA2-II and I (1 mg/mL) with or without para-bromophenacyl bromide (p-BPB) using an agar disc diffusion assay (IC50, 50% inhibition concentration). Ampicillin and cycloheximide (1 mg/mL) were used as positive reference standards for bacteria and fungi, respectively. The results represent the means of three independent experiments and are expressed as the mean ± SD (n = 3).
Table 2.
Antimicrobial activities of the crude extract and purified WaPLA2-II and I (1 mg/mL) with or without para-bromophenacyl bromide (p-BPB) using an agar disc diffusion assay (IC50, 50% inhibition concentration). Ampicillin and cycloheximide (1 mg/mL) were used as positive reference standards for bacteria and fungi, respectively. The results represent the means of three independent experiments and are expressed as the mean ± SD (n = 3).
| Strains | Inhibition Zone (mm) | IC50 (µg/mL) | ||||||
|---|---|---|---|---|---|---|---|---|
| Gram-positive bacteria | Crude extract | WaPLA2-I | WaPLA2-I +p-BPB | WaPLA2-II | WaPLA2-II +p-BPB | Ampicillin/ Cycloheximide | WaPLA2-I | WaPLA2-II |
| B. cereus (ATCC 14579) | 11 ± 0.8 | 21 ± 1.4 | 2.3 ± 0.6 | 17 ± 1.2 | 4.2 ± 0.2 | 22 ± 1 | 15 ± 0.8 | 16 ± 0.7 |
| B. subtilis (ATCC 6633) | 13.5 ± 0.7 | 23.5 ± 0.7 | 4.3 ± 0.6 | 20 ± 0.5 | 5.1 ± 0.3 | 25 ± 1 | 14.3 ± 0.4 | 20 ± 1 |
| E. faecalis (ATCC 29122) | 10.5 ± 0.7 | 21 ± 0.5 | 1.7 ± 0.6 | 27 ± 2.5 | 2.8 ± 0.1 | 24.2 ± 0.7 | 13 ± 0.3 | 16.7 ± 0.8 |
| S. epidermidis (ATCC 14990) | 12 ± 1.4 | 23 ± 2.1 | 3.2 ± 0.3 | 25 ± 2.2 | 6.5 ± 0.5 | 26 ± 0.5 | 15 ± 1.1 | 12 ± 0.7 |
| S. aureus (ATCC 25923) | 15 ± 1.4 | 27 ± 1.2 | 2.6 ± 0.3 | 30 ± 1.7 | 3.3 ± 0.3 | 21.5 ± 1.4 | 13 ± 0.5 | 17 ± 0.4 |
| Gram-negative bacteria | ||||||||
| E. coli (ATCC 25966) | 10 ± 0.8 | 24.2 ± 1 | 3.2 ± 0.3 | 27 ± 2.1 | 4.1 ± 0.1 | 22.6 ± 1.5 | 14 ± 1.1 | 12 ± 0.9 |
| K. pneumonia (ATCC 700603) | 11.5 ± 0.7 | 23 ± 0.7 | 2 ± 0 | 18.5 ± 0.6 | 1.2 ± 0.07 | 25.4 ± 1.7 | 10 ± 0.3 | 14 ± 1.1 |
| P. aeruginosa (ATCC 27853) | 12.5 ± 0.7 | 19.5 ± 0.7 | 1.7 ± 0.3 | 12.5 ± 0.7 | 1.5 ± 0.1 | 20 ± 0.7 | 12 ± 0.7 | 9 ± 0.2 |
| S. enteric (ATCC 43972) | 11 ± 1.4 | 24 ± 1.4 | 2.7 ± 0.6 | 28 ± 2.4 | 4 ± 0.4 | 19.5 ± 0.3 | 17 ± 1.4 | 11 ± 0.5 |
| Fungi | ||||||||
| A. niger | 9 ± 0.7 | 19.5 ± 0.5 | 2.3 ± 0.5 | 15.5 ± 0.5 | 1.5 ± 0.05 | 28 ± 0.6 | 25 ± 1.3 | 20 ± 1.1 |
| B. cinerea | 12 ± 0.5 | 17.8 ± 1.4 | 1.3 ± 0.3 | 19 ± 1.4 | 2.1 ± 0.1 | 29 ± 1 | 26.2 ± 1.7 | 32.7 ± 1.2 |
| F. solani | 7 ± 0.8 | 10.5 ± 0.7 | 1 ± 0 | 13.5 ± 1.7 | 1 ± 0 | 27.5 ± 0.7 | 24.1 ± 1.4 | 19.4 ± 0.8 |
| P. digitatum | 8.5 ± 0.6 | 12 ± 0.7 | 1 ± 0 | 15.4 ± 0.7 | 1.4 ± 0.1 | 21 ± 0.5 | 15.8 ± 0.7 | 10.2 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abid, I.; Jemel, I.; Alonazi, M.; Ben Bacha, A. A New Group II Phospholipase A2 from Walterinnesia aegyptia Venom with Antimicrobial, Antifungal, and Cytotoxic Potential. Processes 2020, 8, 1560. https://doi.org/10.3390/pr8121560
AMA Style
Abid I, Jemel I, Alonazi M, Ben Bacha A. A New Group II Phospholipase A2 from Walterinnesia aegyptia Venom with Antimicrobial, Antifungal, and Cytotoxic Potential. Processes. 2020; 8(12):1560. https://doi.org/10.3390/pr8121560
Chicago/Turabian StyleAbid, Islem, Ikram Jemel, Mona Alonazi, and Abir Ben Bacha. 2020. "A New Group II Phospholipase A2 from Walterinnesia aegyptia Venom with Antimicrobial, Antifungal, and Cytotoxic Potential" Processes 8, no. 12: 1560. https://doi.org/10.3390/pr8121560
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.