
Bioactive Peptides: An Understanding from Current Screening Methodology



1
Department of Tropical Agriculture and International Cooperation, National Pingtung University of Science and Technology, Pingtung 91201, Taiwan
2
Department of Basic Science, Thainguyen University of Agriculture and Forestry, Quyetthang Ward, Thai Nguyen 250000, Vietnam
3
Department of Biological Science and Technology, National Pingtung University of Science and Technology, Pingtung 91201, Taiwan
4
International Master’s Degree Program in Food Science, National Pingtung University of Science and Technology, Pingtung 91201, Taiwan
5
Research Center for Animal Biologics, National Pingtung University of Science and Technology, Pingtung 91201, Taiwan
*
Author to whom correspondence should be addressed.
Processes 2022, 10(6), 1114; https://doi.org/10.3390/pr10061114
Submission received: 28 April 2022
/
Revised: 27 May 2022
/
Accepted: 31 May 2022
/
Published: 2 June 2022
(This article belongs to the Section Biological Processes and Systems)
Abstract
:Bioactive peptides with high potency against numerous human disorders have been regarded as a promising therapy in disease control. These peptides could be released from various dietary protein sources through hydrolysis processing using physical conditions, chemical agents, microbial fermentation, or enzymatic digestions. Considering the diversity of the original proteins and the complexity of the multiple structural peptides that existed in the hydrolysis mixture, the screening of bioactive peptides will be a challenge task. Well-organized and well-designed methods are necessarily required to enhance the efficiency of studying the potential peptides. This article, hence, provides an overview of bioactive peptides with an emphasis on the current strategy used for screening and characterization methods. Moreover, the understanding of the biological activities of peptides, mechanism inhibitions, and the interaction of the complex of peptide–enzyme is commonly evaluated using specific in vitro assays and molecular docking analysis.
1. Introduction
Bioactive peptides (BP) have been identified as specific peptides that possess beneficial pharmacological properties regarding human health [1,2]. BP displays a positive impact on living organisms, particularly in both tendencies of providing nutrients and enhancing immunology defense [2]. Via in vitro, in silico, and in vivo examinations, the BP’s high-potency effects on antioxidant, anti-diabetes, antihypertensive, or multiple functional activities have been discovered [2,3]. According to innovation technology, pharmaceutical drugs have been developed rapidly and shown high efficiency in disease therapies, parallel with concerns about their undesirable side effects [4]. Therefore, natural peptide therapies in human disease prevention and treatment have always been attractive to displace synthetic compounds. The BIOPEP website, known as a bioactive peptide data bank, has contained information of more than 4000 BP with various biology activities [5]. The peptides’ potency in the pharmaceutical industry is additionally expressed through a rather impressive number, with approximately 7% of peptides or peptide-containing molecules in a total of 208 new drugs approved by the US Food Drug Administration (FDA) from 2015 to 2019 [6]. Moreover, hundreds of preclinical and clinical trials have been conducted on a variety of peptides, indicating a significant interest in bioactive peptides [7].
Food proteins have long been considered a potential BP source, and popular protein origins include animal, plant, and marine species [1,3]. Commonly, BP could be found in mixture peptides that are products of food protein hydrolysate processed under chemical and physical mechanisms, microbial fermentation, and enzyme digestion [1,8]. An optimally efficient method of discovering potent peptides from their mixture is a major challenge in terms of time, cost, and labor. Based on the differences in physicochemical properties of peptides, such as molecular weight, hydrophobic, hydrophilic, electric charges, etc., bioassay-guided fractionation methods could be selected to use individual or a combination of several techniques for carrying out this work [9,10,11]. This review article aims to provide a methodological basis for the current strategies applied in the study of BP and analyze their strengths and weaknesses. Furthermore, the processes of determining the characteristics of peptides are also illustrated through modern bioassays.
2. Production of Protein Hydrolysates
2.1. Protein Sources
Bioactive peptides are known as short fragments that are derived from dietary proteins by hydrolysis processing. Commonly, the length of bioactive peptides is approximately two to twenty residues, which can be readily absorbed by the intestinal track or can feasibly access the active sites of certain enzymes due to their small molecular weights [12,13]. Owing to their biological effects and application ability, more and more potential BPs have been identified from food proteins under special conditions, such as chemical agents, fermentation, or enzymatic digestions [1]. Multiple functional peptides could be obtained from various natural bioactive protein sources, popularly animal, plant, marine species, or their by-product hydrolysates (Figure 1).
By far, milk and dairy products are well-known as the largest sources of BP derived from food [14]. For example, peptides were isolated from camel whey hydrolysates relating to cholesterol esterase (CE) inhibitory and pancreatic lipase (PL) inhibitory activities [15]; camel and bovine casein hydrolysates have been associated with α-amylase, α-glucosidase, and dipeptidyl peptidase IV (DPP-IV) inhibitory effects (glucose regulation) [16]; traditional yogurt or sprouted quinoa yogurt beverages have been reported to have anti-oxidative activity, α-glucosidase inhibition, and angiotensin-converting enzyme (ACE) inhibition (antihypertensive activity) [17,18]. Moreover, other animal proteins are considered as potential BP sources, such as egg hydrolysate [9,19,20], cooked beef, pork, chicken, and turkey meat hydrolysate [21], deer velvet antler [22], and even some less popular edible insects [23].
As another rich source of BP, products that originate from plants, such as fruits and vegetables, have been used extensively [24]. Consuming soybean has shown evidence for controlling poor health conditions, such as hypertension, diabetes, and oxidative stress [25,26,27]. Alternatively, amaranth leaf protein hydrolysate showed the presence of multiple functional peptides relating to antioxidant, ACE inhibitory, α-amylase inhibitory, and α-glucosidase inhibitory activities [28]. In addition, peptides derived from marine species, such as pearl oyster meat [29], salmon skin [30,31], silver carp muscle [12], and Trachinus Draco (greater weever) myofibrillar hydrolysates [32] have been demonstrated to have certain health effects.
Food and beverage processing products that have been used in the human diet for a long history are proven to possess a large number of BP. Peptides with the inhibitory activities of α-glucosidase and ACE were identified from the dry-cured ham hydrolysate [33,34] and wine lees hydrolysate [34], respectively. Furthermore, the significant mass of byproducts produced during food processing should be of concern as it may cause high stress to the environment and the economic loss of raw materials. As waste from animal products, bovine bones contain a source of collagen, while the chicken feather protein is a source of keratin, providing high levels of amino acids, such as glycine, alanine, serine, cysteine, and valine. The discovery of antioxidant, DPP-IV inhibitory, and ACE inhibitory peptides has been carried out from bone gelatin and chicken feather hydrolysate [35,36]. Research evidence has shown that chicken foot is a new source of highly effective peptide-calcium chelate, a possible element in functional foods [37]. For other examples of BP derived from byproducts, pearl oyster shells [38], orange seed [39], and almond oil manufacture residue [40] have been reported to be rich sources of BP with antioxidative, antihypertensive, and antidiabetic activity, respectively.
2.2. Production and Processing of Bioactive Peptides
2.2.1. Chemical and Physical Methods
The characteristics of chemical hydrolysis are the presentation of an acidic or basic environment and high temperatures to break the stable protein structure [41,42]. The method commonly uses alkaline treatments that are simple and quick operations to cleavage amide bonds for nonspecific protein hydrolysis. The biggest concern of their drawbacks is losing the nutritional content of the peptides in the extreme pH range controlled at high temperatures [41,43]. Missing amino acids, such as cysteine, lysine, arginine, serine, threonine, and isoleucine, are typical examples that occur during alkaline hydrolysis [44]. Another disadvantage is the lack of specificity in the cleavage of peptide chains [43]. Generally, alkaline hydrolysis has been applied more suitably in the industry than in the laboratory [44]. Application of alkaline hydrolysis was reported in soybean [26], germinated soybean [27], pearl oyster meat [29], and chicken egg white hydrolysate [45] to induce peptides concerning the antioxidant, ACE inhibitory, α-glucosidase inhibitory, DPP-IV inhibitory, and α-amylase inhibitory activities.
The benefits of the acid hydrolysis method are economic, simple, quick, and applicable on a large scale [42,43]. However, the damage of amino acids, such as tryptophan, methionine, and cysteine, appeared. Furthermore, the asparagine and glutamine will be converted to aspartic acid and glutamic acid, respectively, because the deamidation occurs during the acidic hydrolysis. Moreover, the final product is accompanied by a high concentration of salts during the neutralization process, which will interfere with the biological activities of investigated peptides [44]. Therefore, an additional desalting step prior to bioassay is required. For example, α-glucosidase inhibitory peptides were extracted from the Iberian dry-cured ham protein using 0.01 N HCl and then filtered through 0.45 μm nylon filters, and the remained salt and unhydrolyzed proteins were removed from supernatant using ethanol precipitation [33].
Physical methods such as ultrasonic and microwave techniques are widely used to release BP from inactive original protein forms [10,41]. These approaches are considered more cost-effective and simple applications at the industrial level [10]. Principally, cell disruption can be carried out under the direct force of physical techniques. Ultrasonication systems with high pressure, high temperatures, and shock waves can break down molecular bonds and bring about cavitational bubble collapse, finally leading to cell membrane damage and releasing small molecules inside them [41,42]. However, the drawbacks of the physical methods are non-specific and will consequently generate unpredictable products [42]. Therefore, to date, the enzymatic digestion method is usually employed after using the physical process to enhance hydrolysis efficiency. For example, chia seed protein was applied via microwave to assist hydrolysis for sequential proteases with alcalase and flavourzyme to obtain potent antimicrobial peptides [46]. Similarly, an ultra-sonicator was used to enhance the hydrolysis process of SSTY (soft-shelled turtle yolk) protein with proteases to achieve promising DPP-IV inhibitory peptides [9].
According to the benefits and limitations of chemical/physical processes, both approaches are cost-effective for BP’s industrial production. However, their nonspecific nature, unpredictable reaction, and harmfulness to sensitive residues may limit their applications. To circumvent these limitations, enzymatic hydrolysis and fermentation are the most popular methods to release the BP from food proteins.
2.2.2. Microbial Fermentation
The protein substrates have been subjected to the environment culture of bacteria or yeast in the fermentation method [47]. The growth of microorganisms will release proteases that allow the hydrolysis of proteins to produce bioactive peptides from the original material [48]. Generally, the main factors directly affecting to quality and yield of hydrolysis are the strain used, the type of protein, temperature condition, and the fermentation time [47]. The advantages of microbial fermentation to release BP are being cost-effective and eco-friendly, leading to a high recommendation as a suitable method at the industrial level [43,47]. Moreover, through fermentation processing, an improvement in the organoleptic and physicochemical product quality also can be achieved [49]. Multiple species of lactic acid bacteria (LAB), for example, were evaluated for their impact to induce BP from yogurts [17]. The fermented conditions have remained with an additional 1% (v/v) of each strain at pH 4.5 for 4 h. The third yogurt type (Streptococcus thermophilus ST447, Lactobacillus acidophilus NCFM, Lactobacillus rhamnosus GG, and Bifidobacterium lactis HN019) was identified with the biggest number of peptides as compared with the first one (Lactobacillus delbrueckii ssp. bulgaricus LB4 and Streptococcus thermophilus ST447), the second (Lactobacillus delbrueckii ssp. bulgaricus LB4, Streptococcus thermophilus ST447, and Lactobacillus paracasei IMC502). However, the disadvantage of this method is a time-consuming process accompanied with unpredictable products. The fermentation time was 8 h in the study of α-glucosidase and ACE inhibitory peptides generated from sprouted quinoa yogurt beverages or up to 48 h in the case of antioxidant peptides derived from chicken feather hydrolysates [18,35].
2.2.3. Enzymatic Methods
In this process, the protein (substrate) is added to single, double, or multiple proteases to release bioactive peptides [43]. The commonly used proteases in the enzymatic hydrolysis methods have been sourced from microorganisms (e.g., alcalase, flavourzyme, and protamex), plants (e.g., papain), and animal sources (e.g., pepsin and trypsin) [3,41]. The reactions are maintained under individually controlled optimal conditions for each specific protease, such as for pH, temperature, enzyme/substrate ratio, and hydrolysis time [41]. For example, bioactive peptides obtained from edible insects concerning ACE, pancreatic lipase, and α-glucosidase inhibitory properties were hydrolyzed with pepsin (pH 2.5) for 2 h and pancreatin (pH around 7) for 1 h at 37 °C in darkness [23]. In the study of ACE inhibitory peptide derived from Cassia obtusifolia seed protein, hydrolysates were prepared using various proteases corresponding to particular conditions for trypsin (37 °C), α-chymotrypsin (37 °C), and thermolysin (60 °C) at pH 8.5, while pepsin (37 °C) is required at pH 1.3 for 16 h [11].
Interestingly, applying native proteases such as a crude melon extract (CME) has demonstrated the efficiency in hydrolyzing kilka fish (Clupeonella cultriventris caspia) proteins [50]. The CME hydrolysate showed the highest antioxidant activity with a 5.0% degree of hydrolysis (DH) level compared to the fish hydrolysis at 2.5, 5.0, 10.0, and 15.0% DH. Fish hydrolysate achieved by CME displayed better overall emulsion properties than those released from Alcalase (a commercial protease). Therefore, CME may be considered a promising proteolytic enzyme to induce fish hydrolysate. Moreover, raw bovine milk has been detected with endogenous proteases such as plasmin, cathepsins B, and D that can release numerous native peptides from the original protein [51]. After analysis, 175 modified peptides that were identified mainly originated from αS-caseins. In another milk study, plasmin and aminopeptidase had been shown to play a role in the hydrolysis process of proteins or peptides [52]. A total of 1317 peptides were detected in brown unfermented and fermented goat milk.
Additionally, protein hydrolysate can be produced using gastrointestinal digestion enzymes (pepsin, α-chymotrypsin, and trypsin/pancreatin). For instance, the application of simulated gastrointestinal digestion was performed to obtain SSTY-hydrolysate-derived DPP-IV inhibitory peptides [9], soy flour hydrolysate released antioxidant peptides [53], and tuna fish hydrolysate achieved antimicrobial peptides [54]. The containing of medium-sized and short peptide sequences in hydrolysis productions is one of the main advantages of this application (discussed further in Section 3.3).
The advantages of this method are short reaction time, specific, controllability, and predictability, leading to its being more widely used than microbial fermentation [1,48]. The hydrolysis of walnut protein using the protease Alcalase®® 2.4 L to release anti-diabetic peptides takes around 2 h [55], while chicken feather hydrolysates related to antioxidant activity were fermented with the Bacillus licheniformis, taking longer by 24 times up to 48 h [35].
Typically, enzymes have been selected based on the bioactive peptides of interest to generate promising hydrolysate products. For example, trypsin was a suitable enzyme for hydrolyzing casein protein to produce peptides with potential thrombin inhibitory activity [56]. Based on the studies of BP derived from SSTY hydrolysate, thermolysin has been described as a perfect protease to release ACE inhibitory peptides, while DPP-IV inhibitory peptides have obtained better activity under the treatment with a combination of gastrointestinal enzymes, namely trypsin, α-chymotrypsin, and pepsin [9,57]. Among different proteases, such as alcalase, trypsin, pepsin, and chymotrypsin, the amaranth leaf protein hydrolyzed using pepsin protease showed the highest antioxidant activity [28]. Alternatively, the diversity of released bioactive peptides is also caused by the different characteristics of protein sources.
3. Separation and Purification of Bioactive Peptides
3.1. Ultrafiltration
Membrane-based techniques are also widely applied to separate peptides with different molecular weights using permeable membranes. These methods are commonly classified into microfiltration (MF) and ultrafiltration (UF) [41]. The advantages of ultrafiltration techniques are being feasible to scale up the application, unnecessary for extra chemical agents, or easy to be combined with the other processes [10]. Germinated soybean peptides connecting to DPP-IV, α-amylase, and α-glucosidase inhibitory activity were separated using different ultrafiltration membranes (<5, 5–10, and >10 kDa) [27]. Higher inhibitory effects of α-amylase and α-glucosidases were observed on peptides with molecular weights distributed in 5–10 and <5 kDa, while DPP-IV inhibitory peptides were more potent with 5–10 and >10 kDa of molecular weight. Moreover, peptides derived from chia seed hydrolysate with high-potency antimicrobial properties were observed in molecular weights smaller than 3 kDa [46]. Similarly, small peptides (<3 kDa) associated with high biological activities were reported in ACE inhibitory peptides from wheat gluten hydrolysate [58] and DPP-IV inhibitory peptides from Atlantic salmon hydrolysate [31].
3.2. Fractionation and Purification
Bioassay-guided fractionation is evaluated as one of the suitable and efficient strategies in screening the specific peptides associated with a given bioactivity. Moreover, fractionation and purification of potential peptides from their mixture should be carried out to eliminate undesired products, leading to enhancing the accuracy of peptide sequence identification in the next step. The separation of peptides in protein hydrolysate can be achieved by using multiple chromatographic techniques. These methods contain ultra-filtration, reversed-phase (RP) chromatography, and size exclusion chromatography (SEC) (Table 1) [41,42]. They are high-resolution separations that allow fractionation of peptides based on their different physicochemical features, for example, by the hydrophobic properties (RP), size (SEC), and charge (ion exchange, IEX) [10].
Size exclusion chromatography (SEC), also named gel filtration chromatography (GFC), allows capably to fractionate mixtures based on their sizes [10,43]. No sample loss, strong separation, and being easy to use are the remarkable advantages of this method [41]. However, this method has also raised concerns about a limited peak capacity, low resolution, and consuming large volumes of the eluent [10]. Moreover, gel permeation chromatography (GPC) is known as a type of size-exclusion chromatography (SEC); therefore, the principle of GPC is similar to GFC, only different on the use of organic mobile phase instead of an aqueous solution. GFC was applied to separate a peptide mixture to give small peptides in the study of DPP-IV inhibitory peptides from whey hydrolysate [61] and of ACE inhibitory peptides from deer velvet antler hydrolysate [22].
Ion exchange chromatography (IEX) is an efficient technique that permits the separation of compounds depending on their charge [10]. The IEX technique achieved good results even if used independently or combined with the other chromatographic method. However, the disadvantage of this method is the presenting of a high salt concentration in the final product. Therefore, the desalting step is necessary to avoid the interference in further studies, such as the peptide identification and the bioassay of peptides [10].
Reversed-phase high-performance liquid chromatography (RP-HPLC) is a powerful chromatographic technique for separating and purification of peptides from their complex [41]. The principle of the RP-HPLC technique is built on the different hydrophobic properties of amino acids, which can be separated into the chromatographic column under the monitoring of the UV system [2,43]. This technique’s advantages are high resolution, high precision, rapid separation time, and high applicability at both the laboratory and industrial levels [66]. Compounds that are highly hydrophilic may not be suitable to apply the RP method because of raising the risk of losing extremely hydrophilic peptides. The RP-HPLC technique was used to separate the antihypertensive peptides from wine lees hydrolysate into fractions, and then the identified peptides, such as FKTTDQQTRTTVA, NPKLVTIV, TVTNPARIA, LDSPSEGRAPG, and LDSPSEGRAPGAD, showed an ACE inhibitory effect via in vitro and in vivo evaluations [34]. Moreover, the peptides with α-glucosidase and ACE inhibitory effects from quinoa yogurt beverage fractionation [18] and the germinated soybean peptides with antidiabetic activity [27] are also several examples isolated using RP-HPLC.
Typically, the peptides with high biological potency have been recognized as having a close relationship to hydrophobic properties, for example, ACE inhibitory peptides and DPP-IV inhibitory peptides. Therefore, almost all chromatography techniques are considered suitable for application to bioactive peptides discovery (Table 1). However, high potential biological activities may also be found in very hydrophilic peptides that can be lost if the application uses inappropriate isolation techniques. Therefore, the employment of multidimensional column chromatography is highly recommended to enhance the efficiency of BP separation.
Chromatographic techniques are known as powerful tools for the isolation and separation of a complex peptide mixture; however, the number of peptides that coexist in the most active fraction is still too much. The validation of peptide activities is estimated as a costly, time-consuming process, meaning a higher number of required investigated peptides is required. Therefore, orthogonal bioassay-guided fractionation has been applied to circumvent this issue. The principle of this strategy is to use two independent fractionations coupled with an in vitro desired bioassay and then select the identified peptides simultaneously presenting in the most active fractions derived from each method [3,57]. The purpose is to reduce the number of peptide candidates needed for validation, in this way decreasing the cost of the downstream processes to rule out the inactive peptides, especially in the case of complex hydrolysate. For example, Shih et al. used two independent fractionation techniques, RP-HPLC and strong cation-exchange (SCX) liquid chromatography, coupled with an ACE inhibitory assay to identify antihypertensive peptides from Cassia obtusifolia seed hydrolysate [11]. In their study, the de novo sequencing gave only two peptides, FHAPWK and L(I)YL(I)PH (L and I were not differentiated in that study), simultaneously identified from the most active RP-HPLC and SCX fractions.
3.3. Isolation of Short Peptide Sequence
Typically, peptides containing two to four residues in their sequences are considered short peptides that are of significant interest based on these peptides and may present highly biological properties and be easily absorbed into the gastrointestinal tract. However, short peptide sequences analysis has faced several challenges, for example (i) the high risk of loss of very hydrophilic peptides; (ii) the need for a suitable proteomics database to determine sequences shorter than five amino acids; (iii) the requirement of mass-spectrometric (MS) investigation with high resolution to detect exact mass to charge ratios (m/z) values [67]. The application of different separation techniques can maintain short peptides, as determined for short DPP-IV inhibitory peptides from whey hydrolysate [68]. These peptides were successfully identified using HILIC and RP-associated nanofiltration (NF) and LC-MS/MS strategies. Similar to this strategy, the analysis of short peptides in cow milk was performed by HILIC followed by a UHPLC separation and detected using UHPLC-MS/MS (ultra-HPLC–high-resolution MS) investigation [69]. Hydrophilic interaction chromatography and reversed-phase have been particularly advantageous for the isolation of short sequences [67,68].
4. Identification of Peptide Sequences
Following separation and purification processes, the peptide sequence identification can be performed using two methods, Edman degradation and tandem mass spectrometry (MS/MS). The sequencing of peptides can be carried out using Edman degradation with some reagents, such as phenyl isothioeyanate [70]. The cleaved amino acid is started from the N-terminal amino acid residue and is performed one by one amino acid until a complete sequence. The limitations of this strategy can include (i) the requirement of high purity and enough peptides; (ii) being time-consuming; and (iii) not feasible for N-terminal-blocked peptides [71,72]. The combination of Edman degradation and MS has often been observed to analyze the masses of peptides. For example, ACE inhibitory peptides were identified using Edman degradation coupled with MALDI-TOF [73] or combined with fast atom bombardment (FAB)–MS techniques [74,75].
Based on the improved technology of electrospray ionization mass spectrometry (ESI-MS), mass spectrometry (MS) became an effective technique for peptide identification. The foremost advantage of the MS/MS approach is its high reliability, sensitivity, reduced time-consuming analysis, and applicability to peptide mixtures, leading to the replacement of the Edman degradation procedure [76,77]. The sequencing of peptides from MS/MS spectra has been classified into database-assisted and de novo sequencing (Figure 2). In the database matching method, the sequence databases have been provided under ubiquitous Fasta format downloaded from the National Center for Biotechnology Information in the USA or the European Bioinformatics Institute [78]. The available software tool using MS/MS analysis coupled with the support of database sequence matching can be mentioned as PEAKS DB [79], MASCOT [80], SEQUEST [81], OMSSA [82], and X! TANDEM [83]. The advantage of this method is providing a high-accuracy distinction for amino acid residues even in a confusing case, for example, with isomers such as Isoleucine and Leucine. However, peptide sequencing coupled with a database search has been considered difficult when the species’ genome database is unavailable or incomplete.
The de novo sequencing is a useful technique to identify peptide sequences based on their MS/MS spectra without the assistance of a sequence database [76]. The application of de novo peptide sequencing prefers materials with protein databases that have not yet been available or are incomplete [76,84,85]. In addition, the advantage of de novo peptide sequencing is being able to be performed for identifying peptides that have modified amino acids or non-essential amino acids in their sequences, for example, BP derived from fungi [86]. The principle of this approach is based on the match between the residue’s mass and the mass difference of two adjacent fragments (Figure 2) [85,87]. The de novo peptide sequencing requires high mass accuracy; therefore, the application of high-resolution tandem mass spectrometry is a necessary condition to achieve highly reliable results. Fortunately, manual calculation is only one of the options to carry out the de novo sequencing approach [88]. Until now, de novo sequencing algorithms have been developed with various available software tools based on the same basic algorithm, including PEAK, Lutefisk, PepNovo, Unnamed, NovoHMM, SeqMS, EigenMS, AuDeNs, MSNovo, MAARIAN, PFIA, and Vonode [84]. The main algorithmic features, scoring function, and remarks of these software tools were also discussed in Allmer’s study [84]. The commercial tool PEAKS has received a high recommendation as a powerful software of reliability and accuracy for peptide de novo sequencing [85,89]. According to the success rate of the application of the de novo sequencing method, the identities of short peptide sequences have been obtained more suitably due to the increasing length quickly posing more complexity in terms of handling [84].
The biggest drawback of de novo sequencing of peptides is how to distinguish accurately between two residues possessing the same molecular weight, such as Leucine (113.17 Da) and Isoleucine (113.17 Da). The strategies to handle this problem have been mentioned in the literature [90,91,92]. The distinction of Ile/Leu could also be carried out via the presentation of unique w-ions employing MS3 (ETD-HCD) [90], or combining reductive stable isotope-dimethyl-labeled and MS3 fragmentation [92]. Alternatively, the application of charge transfer dissociation mass spectrometry (CTD-MS) is also one of the other selections to overcome this problem [93]. The benefit of the CTD method has been mentioned as no demand for the precursor peptides undergoing any chemical modifications and no need for supplemental levels of isolation and fragmentation.
As mentioned in Section 3.3, the LC-MS/MS investigation of the short peptide sequences is still challenging [94]. The UHPLC-MS/MS technique connecting to Compound Discoverer software is commonly considered a high-efficiency method for the identification of short peptides, for example, antioxidant peptides from soy flour hydrolysate [53] or antimicrobial peptides derived from yellowfin tuna (Thunnus albacares) hydrolysate [54]. Compound Discoverer software is specially designed for the analysis of small peptides based on a high-resolution MS result [94].
5. Characteristics of Bioactive Peptides
5.1. Bioassays for BPs
Once the sequences of BP candidates are identified, the potential peptides are isolated or synthesized for the evaluation of their biological activities using various bioassays. To compare the biological efficacy of hydrolysates or peptides against the target enzyme, their IC50 value, the concentration of which could inhibit 50% of the enzyme’s activity, should be determined. However, the efficacy comparisons are relative due to the use of the same or different conditions, such as the enzyme activity, substrate, buffer, means of detection, etc. The bioassays are regularly carried out under the monitoring of different types of equipment, such as RP-HPLC, spectrophotometer, or spectrofluorometer [95].
To date, the potential functions of hydrolysates and peptides acquired from natural sources have been mainly divided into several biological activities, such as antioxidant, antihypertensive, antidiabetes, and others [1]. The measurement of ACE activity has been popularly performed using the substrate N-α-hippuryl-L-histidyl-L-leucine (HHL), developed on the analogy of angiotensin I, a natural substrate. Cushman and Cheung used it as the ACE’s substrate for the first time and monitored the formation of its hydrolysis products hippuric acid (HA) and histidyl-leucine (HL) at 228 nm [96]. Alternatively, until now, the other synthetic substrates have been widely used for in vitro ACE assays, such as APGG (N-[3-(2-furyl) acryloyl]-L-phenylalanyl-glycyl-glycine) and Abz (Abz-Gly-Phe-(NO2)-Pro; o-aminobenzoylglycyl-p-nitrophenylalanylproline) [97]. The different substrates are adaptively selected according to detectors used to identify products after ACE hydrolysis. Compared to FAPGG or Abz, HHL has been the most common substrate for measuring ACE activity in terms of cost and detector availability [97,98].
In the Henda et al. study, the ACE inhibitory activity of marine cryptides was measured using different substrates, including HHL, angiotensin-I, and FAPGG, that correspond with means of detection, with the former two using RP-HPLC and the last one using a spectrophotometer [97]. Commonly, using a spectrophotometer seems useful, simple, and convenient to quickly acquire results for a large number of samples [97,99]. On the other hand, the application of HPLC allows specific reaction products to be distinguished by the separated peaks on the chromatogram, which can avoid the false positive results comprehensively caused by the combination of multiple products when using a spectrophotometer. Therefore, the HPLC separation coupled with UV–Vis detection received a high recommendation of accuracy. However, this method is time-consuming; for example, one measurement takes at least 15 min, leading to the analysis time for the completion of 96 samples being approximately a day, which is much slower than that performed on a 96-well microplate using a spectrophotometer (e.g., ELISA reader) [97]. To date, both methods are commonly used in the ACE inhibitory assay [99,100].
The inhibitory activities of alpha-glucosidase, dipeptidyl peptidase IV (DPP-IV), or protein-tyrosine phosphatase (PTP-1B) enzymes are known as promising approaches in the management of diabetes. The screening method for these diabetic inhibitors is traditionally measured under controlling of fluorescence and chromogenic detections [101]. The principle of the DPP-IV inhibitor screening is based on the inhibition of DPP-IV’s ability to cleave GP-pNA (Gly-Pro p-nitroanilidee) substrate to release pNA as the reaction product, corresponding to switching the color of the reaction solution from yellow to violet, and can be detected at a wavelength of 405 nm. This method has been the most popular used to carry out DPP-IV screening assays until now. Alternatively, the DPP-IV inhibitory activity can be analyzed using LC-MS introduced by Liu et al. [102]. In the LC-MS method, the detection of Gly-Pro, one of the reaction products of DPP-IV hydrolysis, has been the focus instead of detecting pNA as the traditional method. Therefore, this strategy allows detecting any substrate containing Gly-Pro (m/z value of 173) in the N-terminal of their sequence. Substrate Gly-Pro-pNA and synthetic tripeptide substrates containing Gly-Pro at the N-terminus were reacted with DPP-IV and the properties for the DPP-IV assay were measured by LC-MS analysis. Mass spectrometry (MS) is considered a powerful analysis technique; the degree of confidence in the precision of the DPP-IV assay is also a significant improvement with high specificity and not interfered with by the screening compound.
5.2. Kinetic Study
According to the reaction mechanism, the inhibitors that can inhibit the enzyme’s catalytic activity are generally categorized into competitive, non-competitive, uncompetitive, and mix-type inhibitory modes [3,103,104]. An inhibitor is considered a competitive mode when this inhibitor has a trend competitive with the substrate to occupy the active site of an enzyme, thereby blocking the binding of the enzyme to the substrate. Typically, competitive modes increase the Km value correlating to the substrate concentration while not affecting the Vmax value [103,104]. The addition of substrate concentration leads to an infinite increase in the inhibitor’s IC50 value [104].
Generally, competitive inhibitors have a reaction mechanism that resembles the substrate, and numerous competitive inhibitors possess high-potency enzyme inhibitory activity. For instance, diprotin A (IPI) is an effective DPP-IV inhibitory peptide with the IC50 value of 3.5 μM acting as a competitive inhibitor [105]. The tripeptide sequence is like the substrate structure of the DPP-IV; therefore, peptide IPI is competitive with substrate Gly-Pro-pNA to associate the DPP-IV catalytic site. Alternatively, Pak et al. designed an HMG-CoA reductase inhibitory peptide with a competitive inhibition based on a bioactive conformation of statins (specifically, simvastatin and rosuvastatin) [106]. Statins are commonly known as effective drugs to inhibit HMG–CoA reductase via competitive inhibitory mechanisms. Tetrapeptide YVAE, a competitive inhibitor of HMG–CoA, showed the highest HMG-CoA inhibitory effect as compared with the three remaining peptides ((IAVE, YAVE, and IVAE). Furthermore, they also concluded that the C-terminal residue Glu (E) plays a significant role in affecting HMG-CoA reductase activity since no inhibition activity was observed when IAVE was replaced by IAVA. As the other example, YIE, designed based on peptide sequences (LPYP and IAVPGEVA) from soybeans, showed the inhibitory effect to achieve an overall 700-fold increase compared to LPYP [107]. The kinetic study indicated that this peptide is a competitive-type inhibitor against HMG-CoA reductase. Furthermore, kinetic experiments displayed that GFPTGG peptide and soybean peptide IVAP inhibit HMG-CoA reductase via competitive inhibitory mode and interact with this enzyme as a substrate [108,109].
Uncompetitive inhibitors can attach to the complex of the enzyme and substrate and then prevent the enzyme’s reaction with the substrate. In this circumstance, the Km value obtains an increasing trend, while the Vmax value keeps going in the opposite direction [3,103]. Lan et al. studied the inhibition mode of peptides with the sequence of Trp-Arg-Xaa (where Xaa represents any amino acid). Their result indicated a total of 19 peptides as uncompetitive-type inhibitors against human DPP-IV [110]. Recently, more and more peptides acting as uncompetitive inhibitors with strong biology effects have been reported, such as DPP-IV inhibitory peptides derived from SSTY hydrolysate [9] and the ACE inhibitory peptides characterized from hydrolyzed tilapia [111].
Non-competitive inhibitors can bind to an enzyme or the enzyme–substrate complex and reduce the Vmax value while maintaining the Km value [3,104]. In most cases, non-competitive inhibitors can inhibit the enzyme without being affected by substrate concentration. The ACE inhibitory peptides derived from hazelnut [59], bovine collagen [112], and Enteromorpha clathrate [113] hydrolysates are several examples of enzymes that act in a non-competitive mode. Moreover, the Lineweaver–Burk plot analysis revealed that the DPP-IV inhibitory peptides derived from α-lactalbumin-rich whey protein [61] and salmon skin [31] hydrolysate exhibited non-competitive inhibitory types. Non-competitive inhibitory peptides do not interact with the catalytically active sites of enzymes but can bind to enzymes at the secondary binding positions and inactivate the complex, leading to suppressing the enzyme activity.
Although less common as compared with the mentioned enzyme inhibitors, numerous mixed inhibitors have been found in peptides derived from dietary sources. The mechanism of mixed-type is still incompletely known, and they are generally a mixture of competitive and uncompetitive modes [103,104]. Moreover, mixed-type inhibitors are known as one kind of non-competitive inhibitor [104]. Therefore, mixed activity mode can interact with a free enzyme or the complex of enzyme–substrate [103]. For example, peptides EVSQGRP, VSRHFASYAN, SAAVGSP, and CRQNTLGHNTQTSIAQ from Stichopus horrens hydrolysate showed high ACE inhibition potency [114]. The kinetic analysis indicated that the three former peptides inhibited ACE in mixed inhibition patterns, while the latter one exhibited a non-competitive manner. As for mixed-type inhibitory peptides, the Km and Vmax values have a decreased tendency when increasing the concentration of inhibitor. LSGYGP from tilapia skin gelatin hydrolysate was another example which exhibited a mixed-type inhibitory effect on the ACE enzyme [115]. Captopril, a potent ACE inhibitor, was used as a positive control to compare the inhibitory effect to LSGYGP. According to the in vivo data, this hexapeptide significantly reduced blood pressure after 6 h of oral administration, similar to of captopril, a commercially available ACE inhibitor that was used as a positive control in their study. In addition, some soybean peptides have been demonstrated as the mixed-type inhibitors, which simultaneously showed competitive and uncompetitive inhibitory manners towards α-amylase [116].
5.3. Docking Simulation
Molecular docking is the virtual methodology to study the interactive simulation between ligands (potential peptides) and the receptor (investigating enzyme) that was introduced based on the “induced-fit” theory of Koshland [117,118,119]. Docking tools and programs are one of the most noticeable factors that significantly contributed to the success of docking simulation analysis. The amount of docking software available peaks at over sixty different programs from both academic and commercial sources [120]. According to the literature, the most common software packages that have been used are AutoDock [121], DOCK [122], FlexX [123], GOLD [124], Glide [125], ICM [126], and AutoDock Vina [127]. Regularly, molecular docking is performed according to the following process: (i) selecting the suitable receptor molecule; (ii) simulation preparation for receptor and ligand; (iii) conducting docking simulation; and (iv) analysis simulation results; an illustration appears in Figure 3.
The X-ray crystal structures of receptors are chosen and downloaded from the protein data bank (PDB) [117]. Different codes of crystal conformations from the same enzyme but obtained by different protein-expression systems are presented suitable with the various purpose experiments (Table 2) [3]. Typically, the receptor complex is selected corresponding to the investigating peptides by docking simulation. [128]. The molecular docking analysis of 3-hydroxy-3-methylglutarylcoenzyme A (HMG-CoA) reductase inhibitory peptides used several HMG-CoA PBD codes, particularly the thermodynamic and structure-guided design of statin HMG-CoA reductase inhibitors (PBD code: 3CCZ) [129]; the structure of human HMG-CoA reductase complex with HMG and CoA (PBD code: 1DB8) [130], and Simvastain (PBD code: 1HW9) [131,132]. In an in silico simulation, the structures of IAF, QDF, and QGF peptides derived from cowpea β-vignin contain an aromatic amino acid (Phenylalanine, F) that may interact with the catalytic region of HMG-CoA reductase (PBD code: 1HW9) via the same binding sites as the decalin ring of simvastatin. Therefore, these peptides can inhibit HMG-CoA reductase activity, leading to cholesterol-lowering effects [131].
The interaction bonds between receptors and ligands include hydrogen bonding, pi-pi stacking, polar interaction, hydrophobic interaction, electrostatic interaction, and van der Waals forces [138]. On the enzyme conformations, active positions known as pockets, the catalytic triads are sites binding to peptides to induce the enzyme–peptide complex. For example, the three active sites of the ACE enzyme are S1 (Ala 354, Glu 384, and Tyr 523), S2 (Gln 281, Lys 511, His 513, Tyr 520, and His 353), and S1′ (Glu 162) [36,139,140]. The key residues of the HMG-CoA (3- hydroxy-3-methylglutaryl-coenzyme A reductase) enzyme possess pocket 1 (Ser684, Asp690, Lys691, Lys692) and pocket 2 (Glu559, Lys735, His752, Asn755, and Leu853) [141] and catalytic triad (Tyr479, Asp767, and His866) [142].
Theoretically, the low binding free energy value corresponds to the high biological activity of the peptide. For the explanation, the low interaction energy score indicates that the binding of a ligand and a target receptor is stable. Consequently, this in silico strategy has been popularly used to predict the most promising peptides selected from numerous BP instead of screening individual peptides via in vitro experiments, leading to reduce the cost, time, and laboratory resources for BP screening. Until now, docking simulations have been widely applied for approaches relating to the investigation of BP with the studying enzyme, specifically (i) the identification of their best binding positions; (ii) the revelation of the interaction residues and interaction bonds; (iii) the determination of the binding energy score of the peptide–enzyme complex [118]. For example, the docking simulation was applied to study the ACE inhibitory peptides discovered from pearl oyster [29], hazelnut [59], and chicken egg white hydrolysates [45]; HMG-CoA reductase inhibitory peptides derived from soybean [129], dry-cured ham [130], and the cowpea [131,132]; cholesterol esterase (CE) inhibitory and pancreatic lipase (PL) inhibitory peptides screened from camel whey hydrolysates [15].
The molecular docking study was also recognized for its advantages in predicting a potential peptide possessing both ACE and DPP-IV inhibitory activities [133]. The molecular simulation was applied to investigate 20 peptide sequences derived from fat globule membrane protein hydrolysates of buffalo colostrum relating to their ACE and DPP-IV inhibitory properties, and then only one peptide with the best prediction result was selected for synthetic. For another example, two synthesized tetrapeptides SVPA and SEPA that were selected from 844 in silico simulated gastrointestinal resistant peptides via molecular docking showed their high potential to inhibit α-glucosidase and α-amylase activities [143].
Typically, the result simulated by molecular docking will be further confirmed using an in vitro or in vivo assay. However, according to some previous publications, the application of molecular docking has been considered not suitable to predict the inhibitory activity for non-competitive inhibitors [136,144,145]. In the work reported by Nongonierma et al., the interactions of 8000 possible tripeptides derived from milk protein with the active site of the DPP-IV were simulated using molecular docking [145]. Some inconsistencies occurred between the Vina score predicted by molecular docking and the DPP-IV inhibitory properties of tripeptides via experimental testing. For instance, the peptide WWW showed only a moderate inhibitory activity against DPP-IV, while this peptide obtained the highest docking score among these tripeptides. The docking prediction suggested WWW can directly interact with the pocket site of the DPP-IV; however, it did not correlate with the result of the Lineweaver–Burk plot, which revealed that it is a non-competitive inhibitor. The inconsistency may be due to the factors in the real experiment conditions, such as enzyme concentration, pH, and temperature [128].
6. Conclusions and Future Perspectives
In the current studies, the peptide effects inhibiting the aiming enzyme are usually identified based on most in silico and in vitro assays. Although not as popular as the above assays, in vivo examinations have been significantly performed for this mission. The in vivo studies provide more sufficiency of the efficacy of the peptides under controlling of metabolic interactions and physiological transformations inside the real body. The DPP-IV inhibitory activities of peptides with the in vivo studies are regularly analyzed by oral glucose tolerance tests. Pentapeptide VPLVM from broccoli hydrolysate with an IC50 value of 99.68 μM for DPP-IV inhibitory activity showed remarkable potential in reducing blood glucose after 30 min administration in healthy C57BL/6 female mice models [146]. In another study, the ACE inhibitory peptide LVLPGE (the IC50 value of 13.5 μM) is known as a product after simulated digestion from peptide LVLPGELAK (the IC50 value of 184 μM) derived from broccoli hydrolysate [147]. Hexapeptide LVLPGE had a positive antihypertensive effect in spontaneous hypertensive rat (SHRs) models at doses of 30 mg/kg. Dual functional umami peptide, IPIPATKT, exhibited DPP-IV (the IC50 value of 64 μM) and ACE inhibitory (the IC50 value of 265 μM) activities [148]. IPIPATKT displayed good management of blood glucose levels at a dosage of 150 mg/kg per body weight in C57BL/6N mice after glucose administration. Moreover, IPIPATKT (30.0 mg/kg) demonstrated a hypotensive effect in spontaneous hypertensive rat (SHRs) models.
Furthermore, the human clinical analyses usually received significant interest from peptide researchers. Focusing on ACE inhibitory properties, bovine whey protein hydrolysate possessing IW and WL was supplemented into the diet of healthy participants for ten days [149]. After oral administration of 5 and 50 g hydrolysate, the reduction in plasma ACE activity was recognized to obtain 86.4 ± 5.9 and 75.1 ± 6.9% of the baseline activity, respectively. However, no change in blood pressure was observed. Recently, Cruz-Chamorro et al. studied the effect of lupine (Lupinus angustifolius) protein hydrolysates at the dose of 1 g per day in healthy subjects [150]. After 28 days, an improvement in inflammation, oxidative stress, and cholesterol metabolism was observed in these volunteers.
Generally, the biological activities of peptides are evaluated at a lab scale; however, large-scale investigations are also required to produce bioactive peptides with purposing at industry levels. For example, DPP-IV inhibitory peptides from a boarfish (Capros aper) protein hydrolysate were performed at a semi-pilot scale using Alcalase 2.4L and Flavourzyme 500L [151,152]. Moreover, the studies on ACE inhibitory peptides were performed at the pilot plant to release these peptides from by-products, such as the ricotta-cheese-exhausted whey hydrolysates [153]. Furthermore, peptide fractions from codfish blood and sardine cooking wastewater indicated their good antimicrobial properties, inhibiting Escherichia coli growth [154]. The number of scale-up levels for peptide studies needs to be increased due to their potential replacement of synthetic drugs.
The rapid development of non-communicable diseases, such as hypertension, diabetes, high cholesterol, cardiovascular diseases, and cancers, has attracted great attention, so finding solutions to these disorders becomes an urgent issue. Food-derived bioactive peptides have been regarded as valuable functional food elements that offer various biological activities to prevent and treat diseases. However, the exploitation of BP has also faced several challenges, such as (i) the cost to generate and screen BP from raw materials; (ii) the requirement of an optimization system to obtain target peptides with high potency and yield, leading to large-scale application at the industrial level, not only the laboratory scale; and (iii) the need to overcome in vitro and in vivo models and clinical trials to validate BP’s biological effects and safety before introducing new products in the market. Therefore, the validation of the bioavailability of food-derived peptides is necessary and a requirement. In addition, the application of in silico approaches in predicting the potential BP has achieved significant agreement in terms of cost-effectiveness and efficiency compared with the conventional methods.
Author Contributions
N.T.P.N.: Writing—original draft preparation, Data curation, Investigation, Formal analysis, Conceptualization, Writing—review & editing. J.-L.H.: Conceptualization, Investigation, Methodology, Software, Writing—review & editing, Supervision, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by Ministry of Science and Technology (MOST 110-2113-M-020-001) and Animal Biologics Research of the Featured Area Research Center within the framework of Higher Education Sprout Project programmed by both the Ministry of Education and Ministry of Science and Technology (MOST 110-2634-F-020-001), Taiwan.
Acknowledgments
We gratefully acknowledge the Precision Instrument Center of NPUST and Mithra Biotechnology Inc. for their valuable information about MS/MS-based peptide sequencing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sánchez, A.; Vázquez, A. Bioactive peptides: A review. Food Qual. Saf. 2017, 1, 29–46. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Nong, N.T.P.; Hsu, J.-L. Characteristics of Food Protein-Derived Antidiabetic Bioactive Peptides: A Literature Update. Int. J. Mol. Sci. 2021, 22, 9508. [Google Scholar] [CrossRef]
- Gourgari, E.; Wilhelm, E.E.; Hassanzadeh, H.; Aroda, V.R.; Shoulson, I. A comprehensive review of the FDA-approved labels of diabetes drugs: Indications, safety, and emerging cardiovascular safety data. J. Diabetes Complicat. 2017, 31, 1719–1727. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM database of bioactive peptides: Current opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [Green Version]
- Torre, B.G.D.L.; Albericio, F. Peptide Therapeutics 2.0. Molecules 2020, 25, 2293. [Google Scholar] [CrossRef]
- Henninot, A.; Collins, J.C.; Nuss, J.M. The current state of peptide drug discovery: Back to the future? J. Med. Chem. 2018, 61, 1382–1414. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Nong, N.T.P.; Chen, Y.-K.; Shih, W.-L.; Hsu, J.-L. Characterization of Novel Dipeptidyl Peptidase-IV Inhibitory Peptides from Soft-Shelled Turtle Yolk Hydrolysate Using Orthogonal Bioassay-Guided Fractionations Coupled with In Vitro and In Silico Study. Pharmaceuticals 2020, 13, 308. [Google Scholar] [CrossRef]
- Cermeño, M.; Kleekayai, T.; Amigo-Benavent, M.; Harnedy-Rothwell, P.; FitzGerald, R.J. Current knowledge on the extraction, purification, identification, and validation of bioactive peptides from seaweed. Electrophoresis 2020, 41, 1694–1717. [Google Scholar] [CrossRef]
- Shih, Y.-H.; Chen, F.-A.; Wang, L.-F.; Hsu, J.-L. Discovery and study of novel antihypertensive peptides derived from Cassia obtusifolia seeds. J. Agric. Food Chem. 2019, 67, 7810–7820. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, H.; Hong, H.; Luo, Y. Purification and identification of dipeptidyl peptidase IV and angiotensin-converting enzyme inhibitory peptides from silver carp (Hypophthalmichthys molitrix) muscle hydrolysate. Eur. Food Res. Technol. 2019, 245, 243–255. [Google Scholar] [CrossRef]
- Sato, K.; Miyasaka, S.; Tsuji, A.; Tachi, H. Isolation and characterization of peptides with dipeptidyl peptidase IV (DPPIV) inhibitory activity from natto using DPPIV from Aspergillus oryzae. Food Chem. 2018, 261, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Kamal, M.; Rahman, M.; Siddiqui, M.; Haque, M.; Saha, K.K. Functional dairy products as a source of bioactive peptides and probiotics: Current trends and future prospectives. J. Food Sci. Technol. 2021, 59, 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Baba, W.N.; Mudgil, P.; Baby, B.; Vijayan, R.; Gan, C.-Y.; Maqsood, S. New insights into the cholesterol esterase- and lipase-inhibiting potential of bioactive peptides from camel whey hydrolysates: Identification, characterization, and molecular interaction. J. Dairy Sci. 2021, 104, 7393–7405. [Google Scholar] [CrossRef]
- Mudgil, P.; Kamal, H.; Kilari, B.P.; Salim, M.A.S.M.; Gan, C.-Y.; Maqsood, S. Simulated gastrointestinal digestion of camel and bovine casein hydrolysates: Identification and characterization of novel anti-diabetic bioactive peptides. Food Chem. 2021, 353, 129374. [Google Scholar] [CrossRef]
- Gu, Y.; Li, X.; Liu, H.; Li, Q.; Xiao, R.; Dudu, O.E.; Yang, L.; Ma, Y. The impact of multiple-species starters on the peptide profiles of yoghurts. Int. Dairy J. 2020, 106, 104684. [Google Scholar] [CrossRef]
- Obaroakpo, J.U.; Liu, L.; Zhang, S.; Lu, J.; Pang, X.; Lv, J. α-Glucosidase and ACE dual inhibitory protein hydrolysates and peptide fractions of sprouted quinoa yoghurt beverages inoculated with Lactobacillus casei. Food Chem. 2019, 299, 124985. [Google Scholar] [CrossRef]
- Salim, M.A.S.M.; Gan, C.-Y. Dual-function peptides derived from egg white ovalbumin: Bioinformatics identification with validation using in vitro assay. J. Funct. Foods 2020, 64, 103618. [Google Scholar] [CrossRef]
- Jahandideh, F.; Liu, P.; Wu, J. Purification and identification of adipogenic-differentiating peptides from egg white hydrolysate. Food Chem. 2018, 259, 25–30. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Comparative peptidomic profile and bioactivities of cooked beef, pork, chicken and turkey meat after in vitro gastro-intestinal digestion. J. Proteom. 2019, 208, 103500. [Google Scholar] [CrossRef] [PubMed]
- Haines, S.R.; McCann, M.J.; Grosvenor, A.J.; Thomas, A.; Noble, A.; Clerens, S. ACE inhibitory peptides in standard and fermented deer velvet: An in silico and in vitro investigation. BMC Complement. Altern. Med. 2019, 19, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielińska, E.; Karaś, M.; Baraniak, B.; Jakubczyk, A. Evaluation of ACE, α-glucosidase, and lipase inhibitory activities of peptides obtained by in vitro digestion of selected species of edible insects. Eur. Food Res. Technol. 2020, 246, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Sosalagere, C.; Kehinde, B.A.; Sharma, P. Isolation and functionalities of bioactive peptides from fruits and vegetables: A reviews. Food Chem. 2022, 366, 130494. [Google Scholar] [CrossRef]
- Sitanggang, A.B.; Lesmana, M.; Budijanto, S. Membrane-based preparative methods and bioactivities mapping of tempe-based peptides. Food Chem. 2020, 329, 127193. [Google Scholar] [CrossRef]
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856. [Google Scholar] [CrossRef]
- González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive peptides from germinated soybean with anti-diabetic potential by inhibition of dipeptidyl peptidase-IV, α-amylase, and α-glucosidase enzymes. Int. J. Mol. Sci. 2018, 19, 2883. [Google Scholar] [CrossRef] [Green Version]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, O.S.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Antioxidant and enzymes inhibitory properties of Amaranth leaf protein hydrolyzates and ultrafiltration peptide fractions. J. Food Biochem. 2021, 45, e13396. [Google Scholar] [CrossRef]
- Liu, P.; Lan, X.; Yaseen, M.; Wu, S.; Feng, X.; Zhou, L.; Sun, J.; Liao, A.; Liao, D.; Sun, L. Purification, characterization and evaluation of inhibitory mechanism of ACE inhibitory peptides from pearl oyster (Pinctada fucata martensii) meat protein hydrolysate. Mar. Drugs 2019, 17, 463. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, B.; Tian, Q.; Li, B. Purification and Characterization of Novel Collagen Peptides against Platelet Aggregation and Thrombosis from Salmo salar. ACS Omega 2020, 5, 19995–20003. [Google Scholar] [CrossRef]
- Jin, R.; Teng, X.; Shang, J.; Wang, D.; Liu, N. Identification of novel DPP–IV inhibitory peptides from Atlantic salmon (Salmo salar) skin. Food Res. Int. 2020, 133, 109161. [Google Scholar] [CrossRef] [PubMed]
- Kula, E.; Kocazorbaz, E.K.; Moulahoum, H.; Alpat, S.; Zihnioglu, F. Extraction and characterization of novel multifunctional peptides from Trachinus Draco (greater weever) myofibrillar proteins with ACE/DPP4 inhibitory, antioxidant, and metal chelating activities. J. Food Biochem. 2020, 44, e13179. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; González-Rogel, D.; Heres, A.; Toldrá, F. Iberian dry-cured ham as a potential source of α-glucosidase-inhibitory peptides. J. Funct. Foods 2020, 67, 103840. [Google Scholar] [CrossRef]
- Bravo, F.I.; Mas-Capdevila, A.; López-Fernández-Sobrino, R.; Torres-Fuentes, C.; Mulero, M.; Alcaide-Hidalgo, J.M.; Muguerza, B. Identification of novel antihypertensive peptides from wine lees hydrolysate. Food Chem. 2022, 366, 130690. [Google Scholar] [CrossRef] [PubMed]
- Alahyaribeik, S.; Sharifi, S.D.; Tabandeh, F.; Honarbakhsh, S.; Ghazanfari, S. Stability and cytotoxicity of DPPH inhibitory peptides derived from biodegradation of chicken feather. Protein Expression Purif. 2021, 177, 105748. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, Y.; Hao, Y.; Zhang, W.; Zhou, G. Antihypertensive effects in vitro and in vivo of novel angiotensin-converting enzyme inhibitory peptides from bovine bone gelatin hydrolysate. J. Agric. Food Chem. 2019, 68, 759–768. [Google Scholar] [CrossRef]
- Malison, A.; Arpanutud, P.; Keeratipibul, S. Chicken foot broth byproduct: A new source for highly effective peptide-calcium chelate. Food Chem. 2021, 345, 128713. [Google Scholar] [CrossRef]
- Sasaki, C.; Tamura, S.; Tohse, R.; Fujita, S.; Kikuchi, M.; Asada, C.; Nakamura, Y. Isolation and identification of an angiotensin I-converting enzyme inhibitory peptide from pearl oyster (Pinctada fucata) shell protein hydrolysate. Process Biochem. 2019, 77, 137–142. [Google Scholar] [CrossRef]
- Mazloomi, S.N.; Mahoonak, A.S.; Mora, L.; Ghorbani, M.; Houshmand, G.; Toldrá, F. Pepsin Hydrolysis of Orange By-Products for the Production of Bioactive Peptides with Gastrointestinal Resistant Properties. Foods 2021, 10, 679. [Google Scholar] [CrossRef]
- Gu, X.; Gao, T.; Hou, Y.; Li, D.; Fu, L. Identification and characterization of two novel α-glucosidase inhibitory peptides from almond (Armeniaca sibirica) oil manufacture residue. LWT 2020, 134, 110215. [Google Scholar] [CrossRef]
- Zaky, A.A.; El-Aty, A.A.; Ma, A.; Jia, Y. An overview on antioxidant peptides from rice bran proteins: Extraction, identification, and applications. Crit. Rev. Food Sci. Nutr. 2020, 62, 1350–1362. [Google Scholar] [CrossRef] [PubMed]
- Pal, G.K.; Suresh, P. Sustainable valorisation of seafood by-products: Recovery of collagen and development of collagen-based novel functional food ingredients. Innov. Food Sci. Emerg. Technol. 2016, 37, 201–215. [Google Scholar] [CrossRef]
- Tadesse, S.A.; Emire, S.A. Production and processing of antioxidant bioactive peptides: A driving force for the functional food market. Heliyon 2020, 6, e04765. [Google Scholar] [CrossRef] [PubMed]
- Villamil, O.; Váquiro, H.; Solanilla, J.F. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017, 224, 160–171. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, J.; Liu, C.; Liu, B.; Yu, Y.; Zhang, T. Bifunctional peptides with antioxidant and angiotensin-converting enzyme inhibitory activity in vitro from egg white hydrolysates. J. Food Biochem. 2020, 44, e13347. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.; Deering, A.; Liceaga, A. New Insights into the Antimicrobial Properties of Hydrolysates and Peptide Fractions Derived from Chia Seed (Salvia hispanica L.). Probiotics Antimicrob. Proteins 2020, 12, 1571–1581. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Oh, D.H.; Lee, B.H. Bioactive peptides. Foods 2017, 6, 32. [Google Scholar] [CrossRef]
- Saleh, A.S.; Wang, P.; Wang, N.; Yang, S.; Xiao, Z. Technologies for enhancement of bioactive components and potential health benefits of cereal and cereal-based foods: Research advances and application challenges. Crit. Rev. Food Sci. Nutr. 2019, 59, 207–227. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Purification and identification of antioxidant peptides from fermented fish sauce (Budu). J. Aquat. Food Prod. Technol. 2019, 28, 14–24. [Google Scholar] [CrossRef]
- Alavi, F.; Jamshidian, M.; Rezaei, K. Applying native proteases from melon to hydrolyze kilka fish proteins (Clupeonella cultriventris caspia) compared to commercial enzyme Alcalase. Food Chem. 2019, 277, 314–322. [Google Scholar] [CrossRef]
- Wölk, M.; Milkovska-Stamenova, S.; Hoffmann, R. Comprehensive Profiling of the Native and Modified Peptidomes of Raw Bovine Milk and Processed Milk Products. Foods 2020, 9, 1841. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Du, A.; Fan, Z.; Shi, L. Novel insight into the transformation of peptides and potential benefits in brown fermented goat milk by mesoporous magnetic dispersive solid phase extraction-based peptidomics. Food Chem. 2022, 389, 133110. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, C.; Montone, A.M.I.; Aita, S.E.; Capparelli, R.; Cerrato, A.; Cuomo, P.; Laganà, A.; Montone, C.M.; Piovesana, S.; Capriotti, A.L. Production and characterization of medium-sized and short antioxidant peptides from soy flour-simulated gastrointestinal hydrolysate. Antioxidants 2021, 10, 734. [Google Scholar] [CrossRef] [PubMed]
- Cerrato, A.; Capriotti, A.L.; Capuano, F.; Cavaliere, C.; Montone, A.M.I.; Montone, C.M.; Piovesana, S.; Chiozzi, R.Z.; Laganà, A. Identification and antimicrobial activity of medium-sized and short peptides from yellowfin tuna (Thunnus albacares) simulated gastrointestinal digestion. Foods 2020, 9, 1185. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, T.; Fang, L.; Liu, C.; Liu, X.; Li, H.; Shi, J.; Li, M.; Min, W. Anti-diabetic effect by walnut (Juglans mandshurica Maxim.)-derived peptide LPLLR through inhibiting α-glucosidase and α-amylase, and alleviating insulin resistance of hepatic HepG2 cells. J. Funct. Foods 2020, 69, 103944. [Google Scholar] [CrossRef]
- Tu, M.; Feng, L.; Wang, Z.; Qiao, M.; Shahidi, F.; Lu, W.; Du, M. Sequence analysis and molecular docking of antithrombotic peptides from casein hydrolysate by trypsin digestion. J. Funct. Foods 2017, 32, 313–323. [Google Scholar] [CrossRef]
- Pujiastuti, D.Y.; Shih, Y.-H.; Chen, W.-L.; Hsu, J.-L. Screening of angiotensin-I converting enzyme inhibitory peptides derived from soft-shelled turtle yolk using two orthogonal bioassay-guided fractionations. J. Funct. Foods 2017, 28, 36–47. [Google Scholar] [CrossRef]
- Zhang, P.; Chang, C.; Liu, H.; Li, B.; Yan, Q.; Jiang, Z. Identification of novel angiotensin I-converting enzyme (ACE) inhibitory peptides from wheat gluten hydrolysate by the protease of Pseudomonas aeruginosa. J. Funct. Foods 2020, 65, 103751. [Google Scholar] [CrossRef]
- Liu, C.; Fang, L.; Min, W.; Liu, J.; Li, H. Exploration of the molecular interactions between angiotensin-I-converting enzyme (ACE) and the inhibitory peptides derived from hazelnut (Corylus heterophylla Fisch.). Food Chem. 2018, 245, 471–480. [Google Scholar] [CrossRef]
- Zheng, J.; Bu, T.; Liu, L.; He, G.; Li, S.; Wu, J. Naturally occurring low molecular peptides identified in egg white show antioxidant activity. Food Res. Int. 2020, 138, 109766. [Google Scholar] [CrossRef]
- Jia, C.-L.; Hussain, N.; Obaroakpo, J.U.; Pang, X.-Y.; Zhang, S.-W.; Lu, J.; Liu, L.; Lv, J.-P. Generation and characterization of dipeptidyl peptidase-IV inhibitory peptides from trypsin-hydrolyzed α-lactalbumin-rich whey proteins. Food Chem. 2020, 318, 126333. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Su, R.-Q.; Zhang, W.-T.; Chen, J. Purification and the secondary structure of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the alcalase hydrolysate of seahorse protein. J. Food Sci. Technol. 2020, 57, 3927–3934. [Google Scholar] [CrossRef] [PubMed]
- Kasiwut, J.; Youravong, W.; Sirinupong, N. Angiotensin I-converting enzyme inhibitory peptides produced from tuna cooking juice hydrolysate by continuous enzymatic membrane reactor. J. Food Biochem. 2019, 43, e13058. [Google Scholar] [CrossRef] [PubMed]
- Pearman, N.A.; Ronander, E.; Smith, A.M.; Morris, G.A. The identification and characterisation of novel bioactive peptides derived from porcine liver. Curr. Res. Nutr. Food Sci. 2020, 3, 314–321. [Google Scholar] [CrossRef]
- Peng, L.; Kong, X.; Wang, Z.; Ai-Lati, A.; Ji, Z.; Mao, J. Baijiu vinasse as a new source of bioactive peptides with antioxidant and anti-inflammatory activity. Food Chem. 2021, 339, 128159. [Google Scholar] [CrossRef]
- Kumar, B.R. Application of HPLC and ESI-MS techniques in the analysis of phenolic acids and flavonoids from green leafy vegetables (GLVs). J. Pharm. Anal. 2017, 7, 349–364. [Google Scholar] [CrossRef]
- Piovesana, S.; Montone, C.M.; Cavaliere, C.; Crescenzi, C.; Barbera, G.L.; Laganà, A.; Capriotti, A.L. Sensitive untargeted identification of short hydrophilic peptides by high performance liquid chromatography on porous graphitic carbon coupled to high resolution mass spectrometry. J. Chromatogr. 2019, 1590, 73–79. [Google Scholar] [CrossRef]
- Maux, S.L.; Nongonierma, A.B.; Murray, B.; Kelly, P.M.; FitzGerald, R.J. Identification of short peptide sequences in the nanofiltration permeate of a bioactive whey protein hydrolysate. Food Res. Int. 2015, 77, 534–539. [Google Scholar] [CrossRef]
- Montone, C.M.; Capriotti, A.L.; Cerrato, A.; Antonelli, M.; Barbera, G.L.; Piovesana, S.; Laganà, A.; Cavaliere, C. Identification of bioactive short peptides in cow milk by high-performance liquid chromatography on C18 and porous graphitic carbon coupled to high-resolution mass spectrometry. Anal. Bioanal. Chem. 2019, 411, 3395–3404. [Google Scholar] [CrossRef]
- Edman, P. A method for the determination of the amino acid sequence in peptides. Arch. Biochem. 1949, 22, 475–476. [Google Scholar] [CrossRef] [Green Version]
- Vecchi, M.M.; Xiao, Y.; Wen, D. Identification and sequencing of N-terminal peptides in proteins by LC-fluorescence-MS/MS: An approach to replacement of the Edman degradation. Anal. Chem. 2019, 91, 13591–13600. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.B. Peptide Sequencing by Edman Degradation. 2001. Available online: https://www.ehu.eus/biofisica/juanma/papers/EdmanDegradation.pdf (accessed on 28 April 2022).
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I converting enzyme inhibitory peptides derived from phycobiliproteins of dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetsuna, K.; Maekawa, K.; Chen, J.-R. Antihypertensive effects of Undaria pinnatifida (wakame) peptide on blood pressure in spontaneously hypertensive rats. J. Nutr. Biochem. 2004, 15, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Nakano, T. Identification of an antihypertensive peptide from peptic digest of wakame (Undaria pinnatifida). J. Nutr. Biochem. 2000, 11, 450–454. [Google Scholar] [CrossRef]
- Seidler, J.; Zinn, N.; Boehm, M.E.; Lehmann, W.D. De novo sequencing of peptides by MS/MS. Proteomics 2010, 10, 634–649. [Google Scholar] [CrossRef] [PubMed]
- Kapp, E.; Schütz, F. Overview of tandem mass spectrometry (MS/MS) database search algorithms. Curr. Protoc. Protein. Sci. 2007, 49, 25. [Google Scholar] [CrossRef]
- Cottrell, J.S. Protein identification using MS/MS data. J. Proteom. 2011, 74, 1842–1851. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. PEAKS DB: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. 2012, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophor. Int. J. 1999, 20, 3551–3567. [Google Scholar] [CrossRef]
- Yates, J.R., III; Eng, J.K.; Clauser, K.R.; Burlingame, A.L. Search of sequence databases with uninterpreted high-energy collision-induced dissociation spectra of peptides. J. Am. Soc. Mass Spectrom. 1996, 7, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Geer, L.Y.; Markey, S.P.; Kowalak, J.A.; Wagner, L.; Xu, M.; Maynard, D.M.; Yang, X.; Shi, W.; Bryant, S.H. Open mass spectrometry search algorithm. J. Proteome Res. 2004, 3, 958–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craig, R.; Beavis, R.C. TANDEM: Matching proteins with tandem mass spectra. Bioinformatics 2004, 20, 1466–1467. [Google Scholar] [CrossRef] [PubMed]
- Allmer, J. Algorithms for the de novo sequencing of peptides from tandem mass spectra. Expert Rev. Proteom. 2011, 8, 645–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medzihradszky, K.F.; Chalkley, R.J. Lessons in de novo peptide sequencing by tandem mass spectrometry. Mass Spectrom. Rev. 2015, 34, 43–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Degenkolb, T.; Brückner, H. Peptaibiomics: Towards a myriad of bioactive peptides containing Cα-dialkylamino acids? Chem. Biodivers. 2008, 5, 1817–1843. [Google Scholar] [CrossRef]
- Frank, A.M.; Savitski, M.M.; Nielsen, M.L.; Zubarev, R.A.; Pevzner, P.A. De novo peptide sequencing and identification with precision mass spectrometry. J. Proteome Res. 2007, 6, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Li, W.; Santhanam, R.K.; Wang, C.; Gao, X.; Chen, Y.; Wang, C.; Xu, L.; Chen, H. Bioactive peptide with antioxidant and anticancer activities from black soybean [Glycine max (L.) Merr.] byproduct: Isolation, identification and molecular docking study. Eur. Food Res. Technol. 2019, 245, 677–689. [Google Scholar] [CrossRef]
- Ma, B.; Zhang, K.; Hendrie, C.; Liang, C.; Li, M.; Doherty-Kirby, A.; Lajoie, G. PEAKS: Powerful software for peptide de novo sequencing by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2337–2342. [Google Scholar] [CrossRef]
- Bagal, D.; Kast, E.; Cao, P. Rapid distinction of leucine and isoleucine in monoclonal antibodies using nanoflow LCMSn. Anal. Chem. 2017, 89, 720–727. [Google Scholar] [CrossRef]
- Armirotti, A.; Millo, E.; Damonte, G. How to discriminate between leucine and isoleucine by low energy ESI-TRAP MS n. J. Am. Soc. Mass Spectrom. 2007, 18, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Maibom-Thomsen, S.; Heissel, S.; Mørtz, E.; Højrup, P.; Bunkenborg, J. Discrimination of isoleucine and leucine by Dimethylation-assisted MS3. Anal. Chem. 2018, 90, 9055–9059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, H.M.; Wu, H.T.; Julian, R.R.; Jackson, G.P. Differentiation of leucine and isoleucine residues in peptides using charge transfer dissociation mass spectrometry (CTD-MS). Rapid Commun. Mass Spectrom. 2022, 36, e9246. [Google Scholar] [CrossRef] [PubMed]
- Cerrato, A.; Aita, S.E.; Capriotti, A.L.; Cavaliere, C.; Montone, C.M.; Laganà, A.; Piovesana, S. A new opening for the tricky untargeted investigation of natural and modified short peptides. Talanta 2020, 219, 121262. [Google Scholar] [CrossRef]
- Lahogue, V.; Réhel, K.; Taupin, L.; Haras, D.; Allaume, P. A HPLC-UV method for the determination of angiotensin I-converting enzyme (ACE) inhibitory activity. Food Chem. 2010, 118, 870–875. [Google Scholar] [CrossRef]
- Cushman, D.; Cheung, H. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Henda, Y.B.; Labidi, A.; Arnaudin, I.; Bridiau, N.; Delatouche, R.G.; Maugard, T.; Piot, J.-M.; Sannier, F.D.R.; Thiéry, V.R.; Bordenave-Juchereau, S.P. Measuring angiotensin-I converting enzyme inhibitory activity by micro plate assays: Comparison using marine cryptides and tentative threshold determinations with captopril and losartan. J. Agric. Food Chem. 2013, 61, 10685–10690. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Lv, X.; Xu, X.; Yu, H.; Sun, X.; Xu, N. Purification and identification of a novel ACE inhibitory peptide from marine alga Gracilariopsis lemaneiformis protein hydrolysate. Eur. Food Res. Technol. 2017, 243, 1829–1837. [Google Scholar] [CrossRef]
- Bonazzi, D.; Gotti, R.; Andrisano, V.; Cavrini, V. Analysis of ACE inhibitors in pharmaceutical dosage forms by derivative UV spectroscopy and liquid chromatography (HPLC). J. Pharm. Biomed. Anal. 1997, 16, 431–438. [Google Scholar] [CrossRef]
- Wang, W.; Wang, N.; Zhang, Y.; Cai, Z.; Chen, Q.; He, G. A convenient rp-hplc method for assay bioactivities of angiotensin i-converting enzyme inhibitory peptides. Int. Sch. Res. Not. 2013, 2013, 453910. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zou, J.; Song, Y.; Peng, J.; Wang, Y.; Bi, Y. A novel label-free fluorescence assay for dipeptidyl peptidase 4 activity detection based on supramolecular self-assembly. Chem. Commun. 2020, 56, 1629–1632. [Google Scholar] [CrossRef]
- Liu, J.; Cheng, X.; Fu, L. LC-MS based assay method for DPP-IV inhibitor screening and substrate discovery. Anal. Methods 2012, 4, 1797–1805. [Google Scholar] [CrossRef]
- Ouertani, A.; Neifar, M.; Ouertani, R. Effectiveness of enzyme inhibitors in biomedicine and pharmacotherapy. Adv. Tissue Eng. Regen Med. Open Access 2019, 5, 85–90. [Google Scholar] [CrossRef]
- Blat, Y. Non-competitive inhibition by active site binders. Chem. Biol. Drug Des. 2010, 75, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Rahfeld, J.; Schierborn, M.; Hartrodt, B.; Neubert, K.; Heins, J. Are diprotin A (Ile-Pro-Ile) and diprotin B (Val-Pro-Leu) inhibitors or substrates of dipeptidyl peptidase IV? Biochim. Et Biophys. Acta BBA Protein Struct. Mol. Enzymol. 1991, 1076, 314–316. [Google Scholar] [CrossRef]
- Pak, V.V.; Kim, S.H.; Koo, M.; Lee, N.; Shakhidoyatov, K.; Kwon, D.Y. Peptide design of a competitive inhibitor for HMG-CoA reductase based on statin structure. Pept. Sci. Orig. Res. Biomol. 2006, 84, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Pak, V.V.; Kwon, D.Y.; Khojimatov, O.K.; Pak, A.V.; Sagdullaev, S.S. Design of Tripeptides as a Competitive Inhibitor for HMG-CoA Reductase. Int. J. Pept. Res. Ther. 2021, 27, 1923–1931. [Google Scholar] [CrossRef]
- Pak, V.V.; Koo, M.; Yun, L.; Kwon, D.Y. Recognized sequence and conformation in design of linear peptides as a competitive inhibitor for HMG-CoA reductase. J. Mol. Recognit. Interdiscip. J. 2007, 20, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Pak, V.V.; Koo, M.-S.; Lee, N.-R.; Oh, S.-K.; Kim, M.-S.; Lee, J.-S.; Kwon, D.-Y. Hypocholesterolemic soybean peptide (IAVP) inhibits HMG-CoA reductase in a competitive manner. Food Sci. Biotechnol. 2005, 14, 727–731. [Google Scholar]
- Lan, V.T.T.; Ito, K.; Ito, S.; Kawarasaki, Y. Trp-Arg-Xaa tripeptides act as uncompetitive-type inhibitors of human dipeptidyl peptidase IV. Peptides 2014, 54, 166–170. [Google Scholar] [CrossRef]
- Toopcham, T.; Roytrakul, S.; Yongsawatdigul, J. Characterization and identification of angiotensin I-converting enzyme (ACE) inhibitory peptides derived from tilapia using Virgibacillus halodenitrificans SK1-3-7 proteinases. J. Funct. Foods 2015, 14, 435–444. [Google Scholar] [CrossRef]
- Fu, Y.; Young, J.F.; Rasmussen, M.K.; Dalsgaard, T.K.; Lametsch, R.; Aluko, R.E.; Therkildsen, M. Angiotensin I–converting enzyme–inhibitory peptides from bovine collagen: Insights into inhibitory mechanism and transepithelial transport. Food Res. Int. 2016, 89, 373–381. [Google Scholar] [CrossRef]
- Pan, S.; Wang, S.; Jing, L.; Yao, D. Purification and characterisation of a novel angiotensin-I converting enzyme (ACE)-inhibitory peptide derived from the enzymatic hydrolysate of Enteromorpha clathrata protein. Food Chem. 2016, 211, 423–430. [Google Scholar] [CrossRef]
- Forghani, B.; Zarei, M.; Ebrahimpour, A.; Philip, R.; Bakar, J.; Hamid, A.A.; Saari, N. Purification and characterization of angiotensin converting enzyme-inhibitory peptides derived from Stichopus horrens: Stability study against the ACE and inhibition kinetics. J. Funct. Foods 2016, 20, 276–290. [Google Scholar] [CrossRef]
- Chen, J.; Ryu, B.; Zhang, Y.; Liang, P.; Li, C.; Zhou, C.; Yang, P.; Hong, P.; Qian, Z.J. Comparison of an angiotensin-I-converting enzyme inhibitory peptide from tilapia (Oreochromis niloticus) with captopril: Inhibition kinetics, in vivo effect, simulated gastrointestinal digestion and a molecular docking study. J. Sci. Food Agric. 2020, 100, 315–324. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, X.; Luo, J.; Liu, Y.; Wang, B.; Liang, Z.; Li, L. Insight into the binding modes and mechanisms of inhibition between soybean-peptides and α-amylase based on spectrofluorimetry and kinetic analysis. LWT 2021, 142, 110977. [Google Scholar] [CrossRef]
- Salmaso, V.; Moro, S. Bridging molecular docking to molecular dynamics in exploring ligand-protein recognition process: An overview. Front. Pharmacol. 2018, 9, 923. [Google Scholar] [CrossRef] [Green Version]
- Kitchen, D.B.; Decornez, H.; Furr, J.R.; Bajorath, J. Docking and scoring in virtual screening for drug discovery: Methods and applications. Nat. Rev. Drug Discov. 2004, 3, 935–949. [Google Scholar] [CrossRef]
- Koshland, D., Jr. Application of a theory of enzyme specificity to protein synthesis. Proc. Natl. Acad. Sci. USA 1958, 44, 98. [Google Scholar] [CrossRef] [Green Version]
- Pagadala, N.S.; Syed, K.; Tuszynski, J. Software for molecular docking: A review. Biophys. Rev. 2017, 9, 91–102. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Olson, A.J. Using autodock for ligand-receptor docking. Curr. Protoc. Bioinform. 2008, 24, 8.14.1–8.14.40. [Google Scholar] [CrossRef]
- Allen, W.J.; Balius, T.E.; Mukherjee, S.; Brozell, S.R.; Moustakas, D.T.; Lang, P.T.; Case, D.A.; Kuntz, I.D.; Rizzo, R.C. DOCK 6: Impact of new features and current docking performance. J. Comput. Chem. 2015, 36, 1132–1156. [Google Scholar] [CrossRef] [Green Version]
- Rarey, M.; Kramer, B.; Lengauer, T.; Klebe, G. A fast flexible docking method using an incremental construction algorithm. J. Mol. Biol. 1996, 261, 470–489. [Google Scholar] [CrossRef] [Green Version]
- Verdonk, M.L.; Cole, J.C.; Hartshorn, M.J.; Murray, C.W.; Taylor, R.D. Improved protein–ligand docking using GOLD. Proteins Struct. Funct. Bioinform. 2003, 52, 609–623. [Google Scholar] [CrossRef]
- Friesner, R.A.; Banks, J.L.; Murphy, R.B.; Halgren, T.A.; Klicic, J.J.; Mainz, D.T.; Repasky, M.P.; Knoll, E.H.; Shelley, M.; Perry, J.K. Glide: A new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. J. Med. Chem. 2004, 47, 1739–1749. [Google Scholar] [CrossRef]
- Schapira, M.; Abagyan, R.; Totrov, M. Nuclear hormone receptor targeted virtual screening. J. Med. Chem. 2003, 46, 3045–3059. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- FitzGerald, R.J.; Cermeño, M.; Khalesi, M.; Kleekayai, T.; Amigo-Benavent, M. Application of in silico approaches for the generation of milk protein-derived bioactive peptides. J. Funct. Foods 2020, 64, 103636. [Google Scholar] [CrossRef]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Two peptides from soy β-conglycinin induce a hypocholesterolemic effect in HepG2 Cells by a statin-like mechanism: Comparative in vitro and in silico modeling studies. J. Agric. Food Chem. 2015, 63, 7945–7951. [Google Scholar] [CrossRef] [Green Version]
- Heres, A.; Mora, L.; Toldrá, F. Inhibition of 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase enzyme by dipeptides identified in dry-cured ham. Food Prod. Processing Nutr. 2021, 3, 18. [Google Scholar] [CrossRef]
- Silva, M.; Philadelpho, B.; Santos, J.; Souza, V.; Souza, C.; Santiago, V.; Silva, J.; Souza, C.; Azeredo, F.; Castilho, M. IAF, QGF, and QDF Peptides Exhibit Cholesterol-Lowering Activity through a Statin-like HMG-CoA Reductase Regulation Mechanism: In Silico and In Vitro Approach. Int. J. Mol. Sci. 2021, 22, 11067. [Google Scholar] [CrossRef]
- Silva, M.B.D.C.E.; Souza, C.A.D.C.; Philadelpho, B.O.; Cunha, M.M.N.D.; Batista, F.P.R.; Silva, J.R.D.; Druzian, J.I.; Castilho, M.S.; Cilli, E.M.; Ferreira, E.S. In vitro and in silico studies of 3-hydroxy-3-methyl-glutaryl coenzyme A reductase inhibitory activity of the cowpea Gln-Asp-Phe peptide. Food Chem. 2018, 259, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Ashok, A.; Brijesha, N.; Aparna, H. Discovery, synthesis, and in vitro evaluation of a novel bioactive peptide for ACE and DPP-IV inhibitory activity. Eur. J. Med. Chem. 2019, 180, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Lin, G.; Zhang, R.; Liang, Z.; Wu, L.; Wu, W. Studies on molecular mechanism between ACE and inhibitory peptides in different bioactivities by 3D-QSAR and MD simulations. J. Mol. Liq. 2020, 304, 112702. [Google Scholar] [CrossRef]
- Wang, L.; Ding, L.; Du, Z.; Yu, Z.; Liu, J. Hydrolysis and transport of egg white-derived peptides in Caco-2 cell monolayers and everted rat sacs. J. Agric. Food Chem. 2019, 67, 4839–4848. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Dellafiora, L.; Paolella, S.; Galaverna, G.; Cozzini, P.; FitzGerald, R.J. In silico approaches applied to the study of peptide analogs of Ile-Pro-Ile in relation to their dipeptidyl peptidase IV inhibitory properties. Front. Endocrinol. 2018, 9, 329. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Huang, L.; Zhang, C.; Xie, P.; Cheng, J.; Wang, X.; Liu, L. Skin-care functions of peptides prepared from Chinese quince seed protein: Sequences analysis, tyrosinase inhibition and molecular docking study. Ind. Crop Prod. 2020, 148, 112331. [Google Scholar] [CrossRef]
- Tao, X.; Huang, Y.; Wang, C.; Chen, F.; Yang, L.; Ling, L.; Che, Z.; Chen, X. Recent developments in molecular docking technology applied in food science: A review. Int. J. Food Sci. Technol. 2020, 55, 33–45. [Google Scholar] [CrossRef]
- Qian, B.; Tian, C.; Huo, J.; Ding, Z.; Xu, R.; Zhu, J.; Yu, L.; Villarreal, O.D. Design and evaluation of four novel tripeptides as potent angiotensin converting enzyme (ACE) inhibitors with anti-hypertension activity. Peptides 2019, 122, 170171. [Google Scholar] [CrossRef]
- Natesh, R.; Schwager, S.; Sturrock, E.; Acharya, K. Crystal Structure of Human Angiotensin Converting Enzyme in Complex with Lisinopril. Nature 2003, 421, 551–554. [Google Scholar] [CrossRef] [Green Version]
- Istvan, E.S.; Deisenhofer, J. Structural mechanism for statin inhibition of HMG-CoA reductase. Science 2001, 292, 1160–1164. [Google Scholar] [CrossRef] [Green Version]
- Lateef, T.; Naeem, S.; Qureshi, S.A. In-silico studies of HMG-Co A reductase inhibitors present in fruits of Withania coagulans Dunal (Solanaceae). Trop. J. Pharm. Res. 2020, 19, 305–312. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Bester, M.J.; Neitz, A.W.; Gaspar, A.R. Rational in silico design of novel α-glucosidase inhibitory peptides and in vitro evaluation of promising candidates. Biomed. Pharmacother. 2018, 107, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. Inhibition of dipeptidyl peptidase IV and xanthine oxidase by amino acids and dipeptides. Food Chem. 2013, 141, 644–653. [Google Scholar] [CrossRef] [Green Version]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. In silico approaches to predict the potential of milk protein-derived peptides as dipeptidyl peptidase IV (DPP-IV) inhibitors. Peptides 2014, 57, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Liu, Z.; Pan, D.; Zhao, Y.; Dang, Y.; Gao, X. Transport, Stability, and In Vivo Hypoglycemic Effect of a Broccoli-Derived DPP-IV Inhibitory Peptide VPLVM. J. Agric. Food Chem. 2022, 70, 4934–4941. [Google Scholar] [CrossRef]
- Pei, J.; Hua, Y.; Zhou, T.; Gao, X.; Dang, Y.; Wang, Y. Transport, in vivo antihypertensive effect, and pharmacokinetics of an Angiotensin-Converting Enzyme (ACE) inhibitory peptide LVLPGE. J. Agric. Food Chem. 2021, 69, 2149–2156. [Google Scholar] [CrossRef]
- Chen, M.; Pan, D.; Zhou, T.; Gao, X.; Dang, Y. Novel Umami Peptide IPIPATKT with Dual Dipeptidyl Peptidase-IV and Angiotensin I-Converting Enzyme Inhibitory Activities. J. Agric. Food Chem. 2021, 69, 5463–5470. [Google Scholar] [CrossRef]
- Martin, M.; Hagemann, D.; Nguyen, T.T.; Schwarz, L.; Khedr, S.; Moskopp, M.L.; Henle, T.; Deussen, A. Plasma concentrations and ACE-inhibitory effects of tryptophan-containing peptides from whey protein hydrolysate in healthy volunteers. Eur. J. Nutr. 2020, 59, 1135–1147. [Google Scholar] [CrossRef]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Álvarez-Ríos, A.I.; Santos-Sánchez, G.; Pedroche, J.; Millán, F.; Sánchez, C.C.; Fernández-Pachón, M.S.; Millán-Linares, M.C.; Martínez-López, A. Safety and Efficacy of a Beverage Containing Lupine Protein Hydrolysates on the Immune, Oxidative and Lipid Status in Healthy Subjects: An Intervention Study (the Lupine-1 Trial). Mol. Nutr. Food Res. 2021, 65, 2100139. [Google Scholar] [CrossRef]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; Crowe, W.; Allsopp, P.J.; McSorley, E.M.; Devaney, M.; Whooley, J.; McGovern, B.; Parthsarathy, V.; O’Harte, F.P. Stability to thermal treatment of dipeptidyl peptidase-IV inhibitory activity of a boarfish (Capros aper) protein hydrolysate when incorporated into tomato-based products. Int. J. Food Sci. Technol. 2021, 56, 158–165. [Google Scholar] [CrossRef]
- Harnedy-Rothwell, P.A.; McLaughlin, C.M.; O’Keeffe, M.B.; Gouic, A.V.L.; Allsopp, P.J.; McSorley, E.M.; Sharkey, S.; Whooley, J.; McGovern, B.; O’Harte, F.P. Identification and characterisation of peptides from a boarfish (Capros aper) protein hydrolysate displaying in vitro dipeptidyl peptidase-IV (DPP-IV) inhibitory and insulinotropic activity. Food Res. Int. 2020, 131, 108989. [Google Scholar] [CrossRef] [PubMed]
- Pontonio, E.; Montemurro, M.; Gennaro, G.V.D.; Miceli, V.; Rizzello, C.G. Antihypertensive Peptides from Ultrafiltration and Fermentation of the Ricotta Cheese Exhausted Whey: Design and Characterization of a Functional Ricotta Cheese. Foods 2021, 10, 2573. [Google Scholar] [CrossRef] [PubMed]
- Ghalamara, S.; Silva, S.; Brazinha, C.; Pintado, M. Valorization of fish by-products: Purification of bioactive peptides from codfish blood and sardine cooking wastewaters by membrane processing. Membranes 2020, 10, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Workflow for identifying peptide sequences from dietary protein sources.

Figure 2.
Identified peptides by de novo sequencing and database sequencing from tandem mass spectra.
Figure 2.
Identified peptides by de novo sequencing and database sequencing from tandem mass spectra.

Figure 3.
Flowchart for molecular docking (i) preparation receptor and ligands; (ii) docking; (iii) simulation results, including determining interaction residues, interaction bonds, and calculation binding mode score of a ligand to a receptor.
Figure 3.
Flowchart for molecular docking (i) preparation receptor and ligands; (ii) docking; (iii) simulation results, including determining interaction residues, interaction bonds, and calculation binding mode score of a ligand to a receptor.


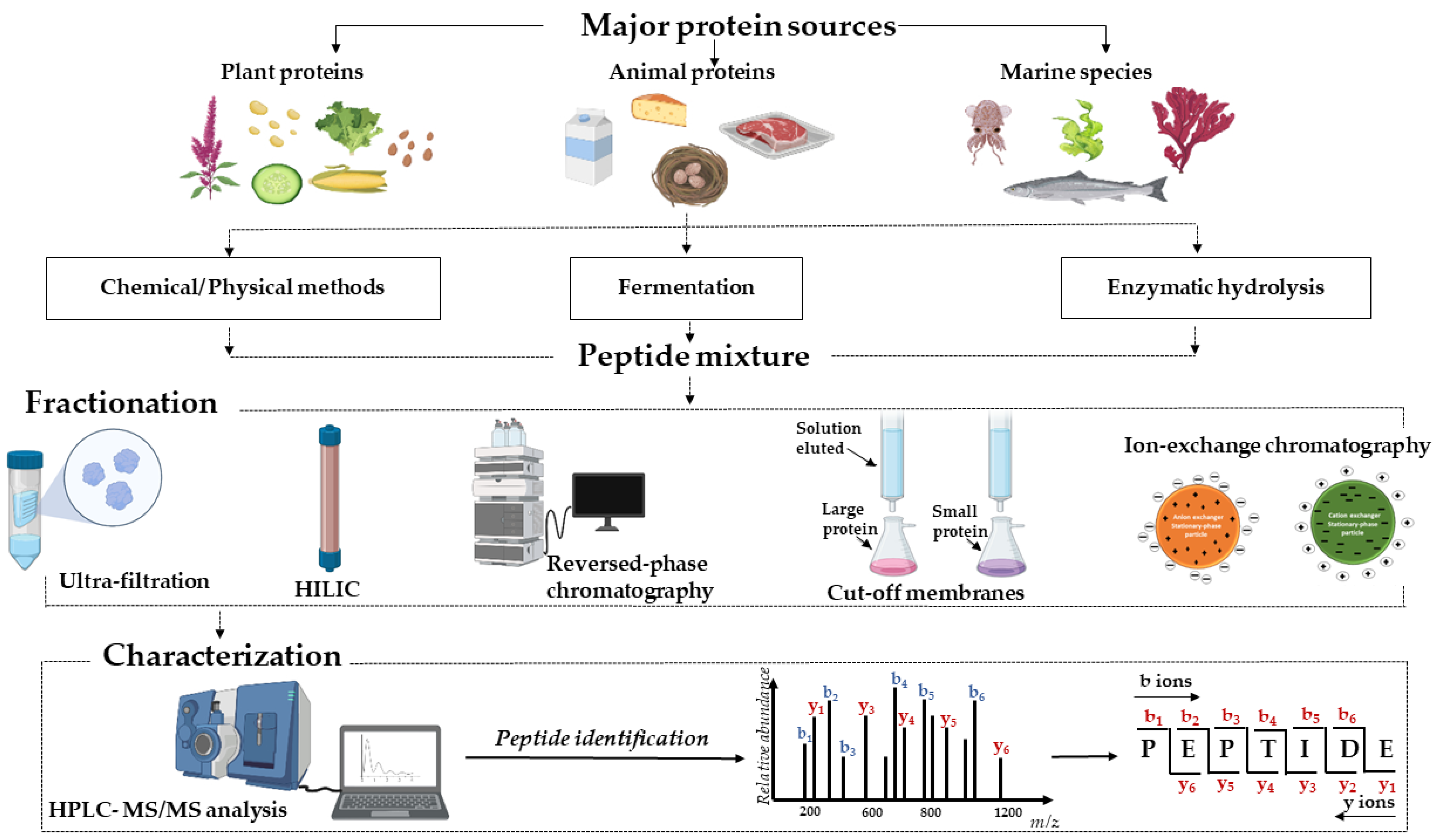{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary methods on functional properties of bioactive peptides fractionation and purification.
Table 1.
Summary methods on functional properties of bioactive peptides fractionation and purification.
| Source | Hydrolysate Treatment | Fractionation Method * | Peptide Sequence | Health Effects ** | Validation *** | Ref. | |
|---|---|---|---|---|---|---|---|
| In Vitro | In Vivo | ||||||
| Plant | |||||||
| Chia Seed | Alcalase, Flavourzyme | GDVIAIR | Antimicrobial | Yes | No | [46] | |
| Cassia obtusifolia seed | Trypsin, α-chymotrypsin, pepsin, and thermolysin | 1. SCX 2. RP-HPLC | FHAPWK, LYIPH, LYLPH, IYIPH, and IYLPH | ACE inhibition | Yes | Yes | [11] |
| Wheat gluten hydrolysate | Alcalase | 1. FPLC | SAGGYIW and APATPSFW | ACE inhibition | Yes | No | [58] |
| Hazelnut | Alcalase | 1. Ion-exchange chromatography 2. GPC 3. RP-HPLC | AVKVL, YLVR, and TLVGR | ACE inhibition | Yes | Yes | [59] |
| Soybean hydrolysate | Extraction with water, alcalase, papain | 1. Ultrafiltration | Antioxidant | Yes | No | [25] | |
| ACE inhibition | |||||||
| α-glucosidase inhibition | |||||||
| Kunitz trypsin inhibition | |||||||
| Soybean hydrolysate | Alkaline, papain, trypsin | 1. SEC 2. Anion-exchange chromatography | LLPLPVLK, SWLRL, WLRL | Antioxidant | Yes | No | [26] |
| ACE inhibition | |||||||
| α-glucosidase inhibition | |||||||
| DPP-IV inhibition | |||||||
| Germinated soybean | Alkaline | 1. Ultrafiltration 2. RP-HPLC | DPP-IV inhibition | Yes | No | [27] | |
| α-amylase inhibition | |||||||
| α-glucosidase | |||||||
| Walnut hydrolysate | Alcalase | 1. Ultrafiltration 2. SEC 2 times 3. RP-HPLC | LPLLR | α-glucosidase | Yes | No | [55] |
| α-amylase | |||||||
| Alleviating insulin resistance | |||||||
| Animal and animal products | |||||||
| Chicken egg white hydrolysate | Thermolysin, pepsin | 1. Ultrafiltration 2. SCX 3. RP-HPLC | WEKAFKDED, QAMPFRVTEQE, ERYPIL, VFKGL | Adipogenic | Yes | No | [20] |
| Chicken egg white hydrolysate | Trypsin, pepsin, neutral proteinase, alkaline proteinase, acidic protease | 1. Ultrafiltration 2. SEC | LAPYK, SVIRW, PKSVIRW, ADWAK | Antioxidant | Yes | No | [45] |
| ACE inhibition | |||||||
| Chicken egg white hydrolysate | Ethanol | 1. SEC 2. RP-HPLC | STDVPRDPWVWGSAHPQAQHTR, GDPSAWSWGAEAHS, ALGEDIVDLDSFSEQH | Antioxidant | Yes | No | [60] |
| Soft-shelled turtle yolk hydrolysate | Pepsin, trypsin, chymotrypsin, and thermolysin | 1. SCX 2. RP-HPLC | LPLF, WLQL, LPSW, VPGLAL, LVGLPL | DPP-IV inhibition | Yes | No | [9] |
| Camel whey hydrolysates | Pepsin | 1. RP-UPLC | FCCLGPVPP | CE inhibition | Yes | No | [15] |
| PAGNFLPPVAAAPVM, MLPLMLPFTMGY, and LRFPL | PL inhibition | ||||||
| Camel and bovine casein hydrolysates | Alcalase and pronase E | 1. RP-UPLC | FLWPEYGAL | α-amylase inhibition | Yes | No | [16] |
| HLPGRG, QNVLPLH, and PLMLP | DPP-IV inhibition | ||||||
| LPTGWLM, MFE and GPAHCLL | α-glucosidase inhibition | ||||||
| α-lactalbumin-rich whey hydrolysate | Trypsin | 1. Ultrafiltration 2. SEC | LDQWLCEKL | DPP-IV inhibition | Yes | No | [61] |
| Milk protein hydrolysis (Yogurt) | 1. L. bulgaricus LB4 and S. thermophilus ST447; 2. L. bulgaricus LB4, S. thermophilus ST447, and L. paracasei IMC502; 3. S. thermophilus ST447, L. acidophilus NCFM, 4. L. rhamnosus GG and B. lactis HN019. | 1. Ultrafiltration | NENLLRFF | ACE inhibition | Yes | No | [17] |
| Antioxidative | |||||||
| Stimulating properties | |||||||
| Sprouted quinoa yogurt beverages | Lactobacilluscasei | 1. RP-HPLC | LAHMIVAGA, VAHPVF | α-glucosidase inhibition | Yes | No | [18] |
| ACE inhibition | |||||||
| DPP-IV inhibition | |||||||
| Antioxidant | |||||||
| Deer velvet antler | Pepsin | 1. SEC | LVVYPW, LVVYPWTQ and VVYPWTQ | ACE inhibition | Yes | No | [22] |
| Edible insect | Pepsin and pancreatin | 1. SEC | KVEGDLK, YETGNGIK, AIGVGAIR, IIAPPER, FDPFPK | α-glucosidase inhibition | Yes | No | [23] |
| Lipase inhibition | |||||||
| ACE inhibition | |||||||
| Marine species | |||||||
| Pearl oyster meat protein | Alkaline | 1. Ultrafiltration 2. GPC 3. RP-HPLC | HLHT and GWA | ACE inhibition | Yes | Yes | [29] |
| Salmo salar skin | Alcalase or protamex | 1. YMC ODS-A C18 separation | OGEFG, DEGP | Antiplatelet | Yes | Yes | [30] |
| Atlantic salmon skin | Pepsin, trypsin, papain, or Alcalase | 1. Ultrafiltration 2. Gel chromatography 3. RP-HPLC | LDKVFR, YYGYTGAFR, VLATSGPG | DPP-IV inhibition | Yes | No | [31] |
| Seahorse protein hydrolysate | Alcalase | 1. GPC 2. RP-HPLC | PAGPRGPA | ACE inhibitory | Yes | No | [62] |
| Tuna cooking juice hydrolysate | Alcalase | 1. SEC 2. RP-HPLC | FTGATKPEF, HMGGMTRHA, FTGATKPGF | ACE inhibition | Yes | No | [63] |
| Silver carp muscle hydrolysate | Alcalase 2.4 L, Neutrase, pepsin, trypsin, and Flavourzyme | 1. Ultrafiltration 2. SEC 3. RP-HPLC | LLDLGVP, AALEQTER, ILYGDFK, KAVGEPPLF, GPAGPQGPR | DPP-IV inhibition | Yes | No | [12] |
| ACE inhibition | |||||||
| Trachinus Draco (greater weever) myofibrillar | Trypsin and alcalase | 1. RP-HPLC | DLSGSTAMKEAVDNAYAR and FPGDHDR | DPP-IV inhibition | [32] | ||
| ACE inhibition | Yes | No | |||||
| Metal chelation activity | |||||||
| Antioxidant | |||||||
| Byproduct | |||||||
| Porcine liver | Papain | 1. RP-HPLC | FWG, MFLG, and SDPPLVFVG | Antioxidant | Yes | No | [64] |
| Bovine bone gelatin | Alcalase 2.4 L | 1. Ultrafiltration 2. SEC 3. RP-HPLC | RGL-(Hyp)-GL and RGM-(Hyp)-GF | ACE inhibition | Yes | Yes | [36] |
| Pearl oyster shell hydrolysate | Nucleicin and Orientase 22 BF | 1. SEC 2. RP-HPLC | GVGSPY | ACE inhibition | Yes | No | [38] |
| Almond oil manufacture residue | Prote Ax and Protease M | 1. Ultrafiltration 2. Gel chromatography 3. RP-HPLC | WH, WS | α-glucosidase inhibition | Yes | No | [40] |
| Baijiu vinasse | Corolase PP | 1. Ultrafiltration | KLPDHPKLPK, VDVPVKVPYS | Antioxidant | Yes | No | [65] |
| Anti-inflammatory | |||||||
| Orange seed protein | Pepsin | 1. SEC 2. RP-HPLC | α-glucosidase inhibition | No | No | [39] | |
| α-amylase inhibition | |||||||
| ACE inhibition | |||||||
| Ferric-reducing antioxidant power | |||||||
| DPPH radical scavenging activity | |||||||
| Product | |||||||
| Wine lees hydrolysate | Flavourzyme®® | 1. Ultrafiltration 2. RP-HPLC | FKTTDQQTRTTVA, NPKLVTIV, TVTNPARIA, LDSPSEGRAPG, LDSPSEGRAPGAD | ACE inhibition | Yes | Yes | [34] |
| Iberian dry-cured ham | HCl | 1. SEC 2. RP-HPLC 3. HILIC-HPLC | AEEEYPDL, LGVGG | α-glucosidase inhibition | Yes | No | [33] |
* Size exclusion chromatography (SEC); gel permeation chromatography (GPC); strong cation exchange (SCX); reversed-phase high-performance liquid chromatography (RP-HPLC); fast protein liquid chromatography system (FPLC); ** cholesterol esterase (CE); pancreatic lipase (PL); dipeptidyl peptidase IV (DPP-IV); angiotensin-converting enzyme (ACE); *** yes/no, with or without analyses experiments.
Table 2.
Molecular software sources studied on the interaction of bioactive peptides and enzymes.
| Bioactivity | Receptors | PDB Code of Receptors | Software | Sequences | Ref. |
|---|---|---|---|---|---|
| AEC inhibitory | ACE | 1O86 | Discovery Studio | RGL-(Hyp)-GL and RGM-(Hyp)-GF | [36] |
| 1O86 | AutoDock | HLHT and GWA | [29] | ||
| 1O86 | AutoDock | LAPYK, ADWAK, SVIRW, PKSVIRW | [45] | ||
| 1O86 | Maestro | GIPLPLI | [133] | ||
| 1O86 | AutoDock | VKW and GTW | [134] | ||
| DPP-IV inhibitory | DPP-IV | 5Y7H | AutoDock | LDKVFR, YYGYTGAFR, and VLATSGPG | [31] |
| 1WCY | Maestro | GIPLPLI | [133] | ||
| 3Q8W, 3F8S, and 5T4B | Discovery Studio | IRDLLER, YAEERYP, IRNVQPS | [135] | ||
| 1ORW | Sybyl | APA, APF, APR, IPA, LPA, and FPF | [136] | ||
| Cholesterol esterase (CE) inhibitory | CE | 1AQL and 1F6W | PepSite | FCCLGPVPP | [15] |
| Pancreatic lipase (PL) inhibitory | PL | 1ETH | PepSite | PAGFLPPVAAAPVM, MLPLMLPFTMGY, and LRFPL | [15] |
| HMG-CoA inhibitory | HMG–CoA reductase | 1HW9 | AutoDock Vina | QGF, IAF, QDF, and PIY | [131] |
| 1DQ8 | AutoDock | AA, AL, AQ, DA, DD, EE, ES, LL, QQ, and VH | [130] | ||
| 1HW9 | AutoDock Vina | QDF | [132] | ||
| 3CCZ | PLANTS | YVVNPDNDEN and YVVNPDNNEN | [129] | ||
| Tyrosinase inhibition | Tyrosinase | 2Y9X | Discovery Studio | RHAKF and NYRRE | [137] |
| Antithrombotic | Thrombin | 2BVR | Discovery Studio | FQSEEQQQTEDELQDK | [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nong, N.T.P.; Hsu, J.-L. Bioactive Peptides: An Understanding from Current Screening Methodology. Processes 2022, 10, 1114. https://doi.org/10.3390/pr10061114
AMA Style
Nong NTP, Hsu J-L. Bioactive Peptides: An Understanding from Current Screening Methodology. Processes. 2022; 10(6):1114. https://doi.org/10.3390/pr10061114
Chicago/Turabian StyleNong, Nhung Thi Phuong, and Jue-Liang Hsu. 2022. "Bioactive Peptides: An Understanding from Current Screening Methodology" Processes 10, no. 6: 1114. https://doi.org/10.3390/pr10061114
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.