Isolation and Biophysical Characterisation of Bioactive Polysaccharides from Cucurbita Moschata (Butternut Squash)

 ,
,  ,
,
Abstract
:
1. Introduction
2. Methods
2.1. Isolation of Polysaccharide
2.2. Gel Chromatography
2.3. Hydrodynamic Characterisation
2.3.1. Sample Preparation
2.3.2. Intrinsic Viscosity
2.3.3. Sedimentation Velocity
2.3.4. Sedimentation Equilibrium
2.3.5. MSM Estimates
2.3.6. Conformational Flexibility
2.4. Complement Fixation
2.5. Methanolysis for Monosaccharide Composition Determination Using Gas Chromatography
2.6. GCMS and Linkage Determination
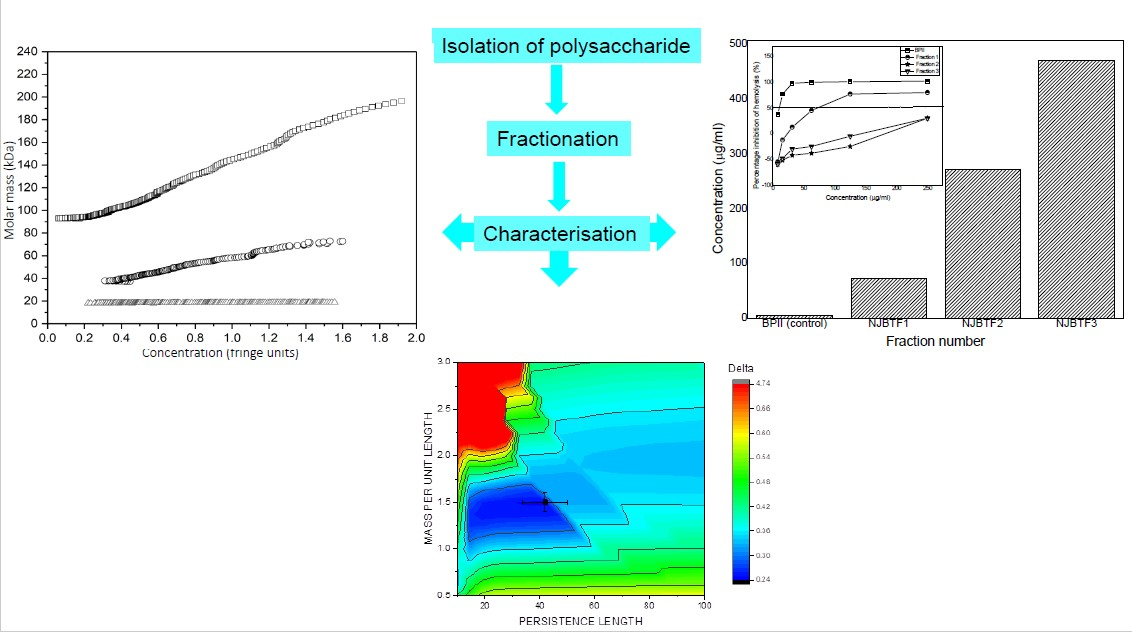
3. Results and Discussion
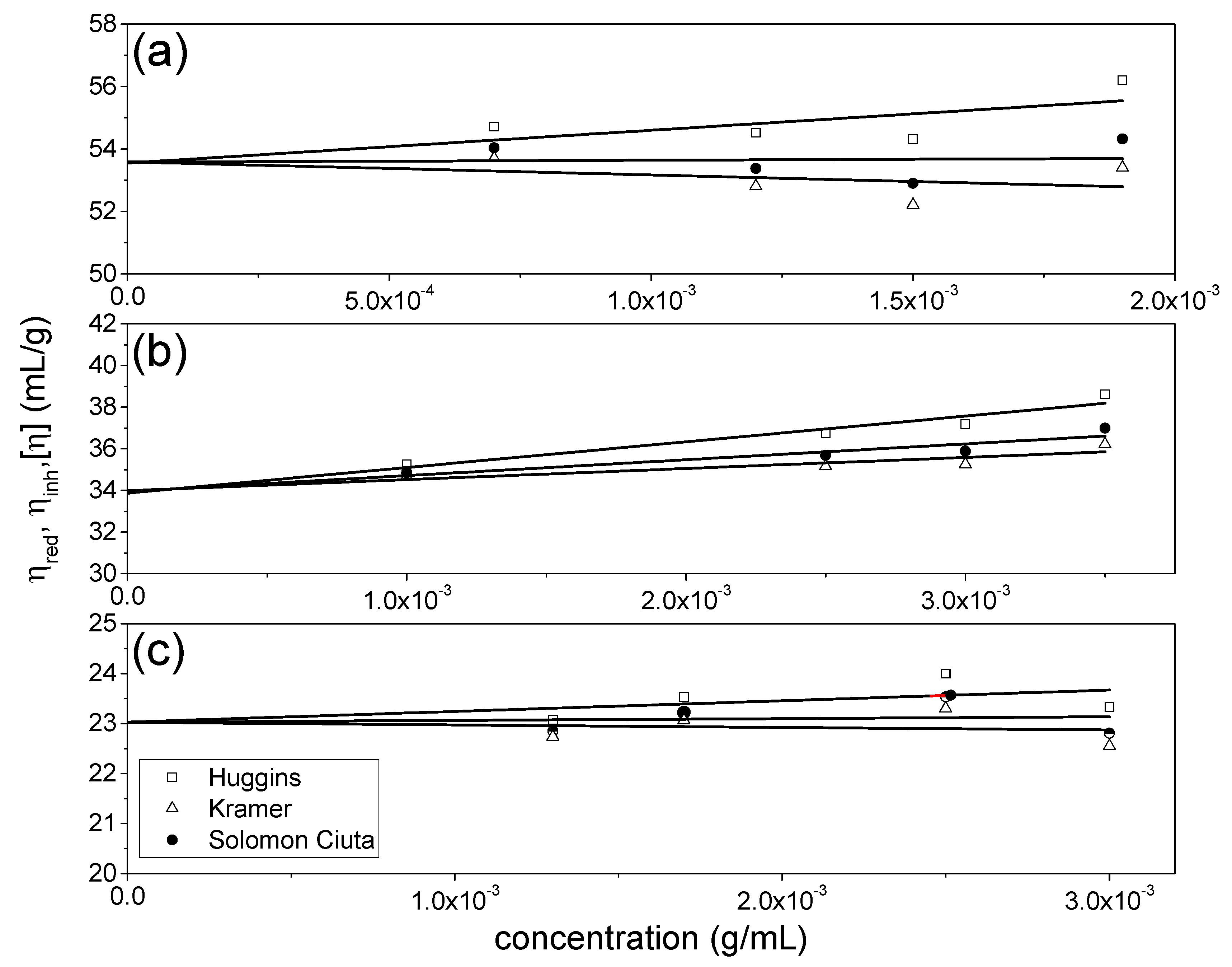
3.1. Intrinsic Viscosities
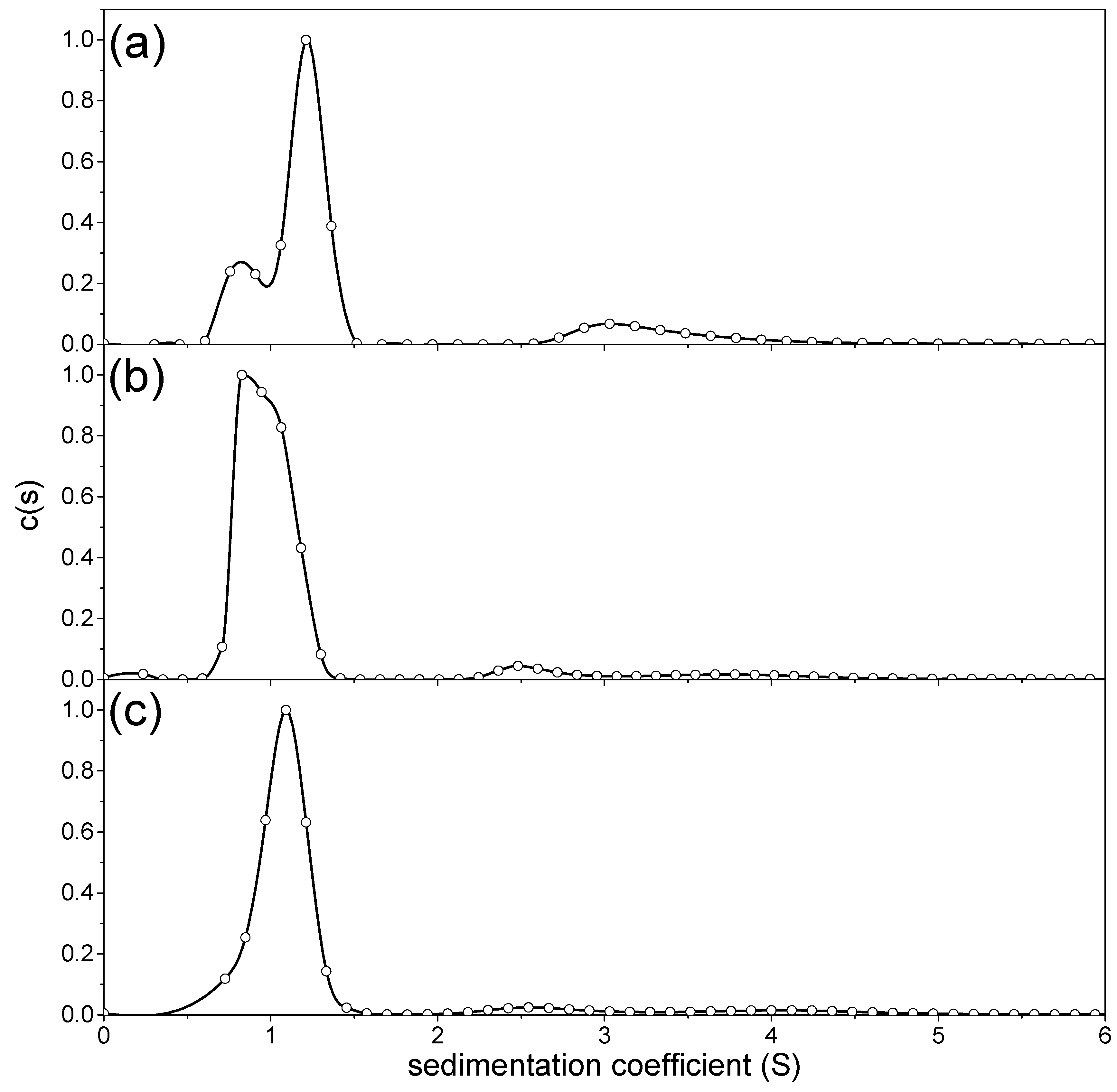
3.2. Sedimentation Coefficients and Distributions
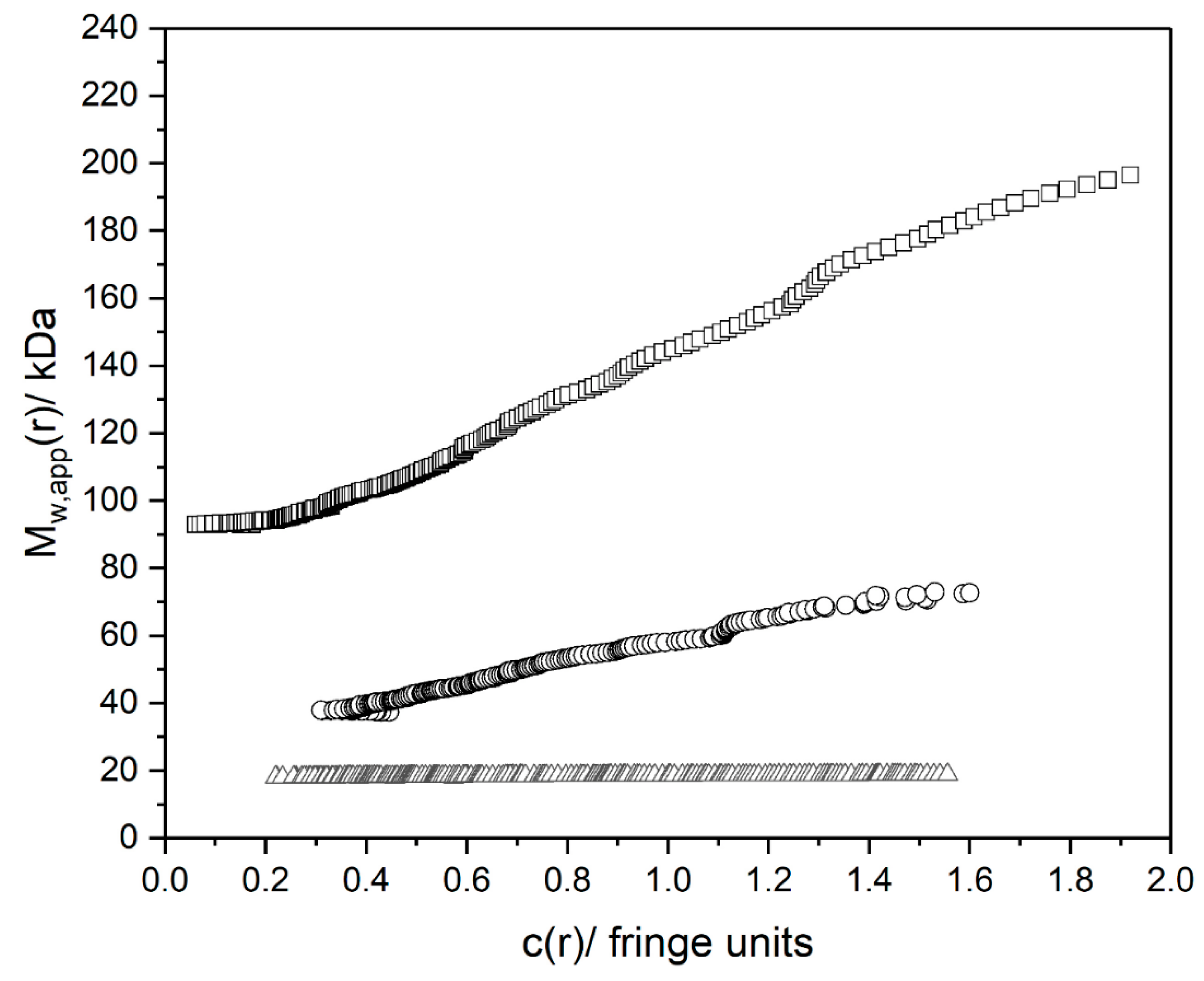
3.3. Molecular Weights
3.4. Persistence Length
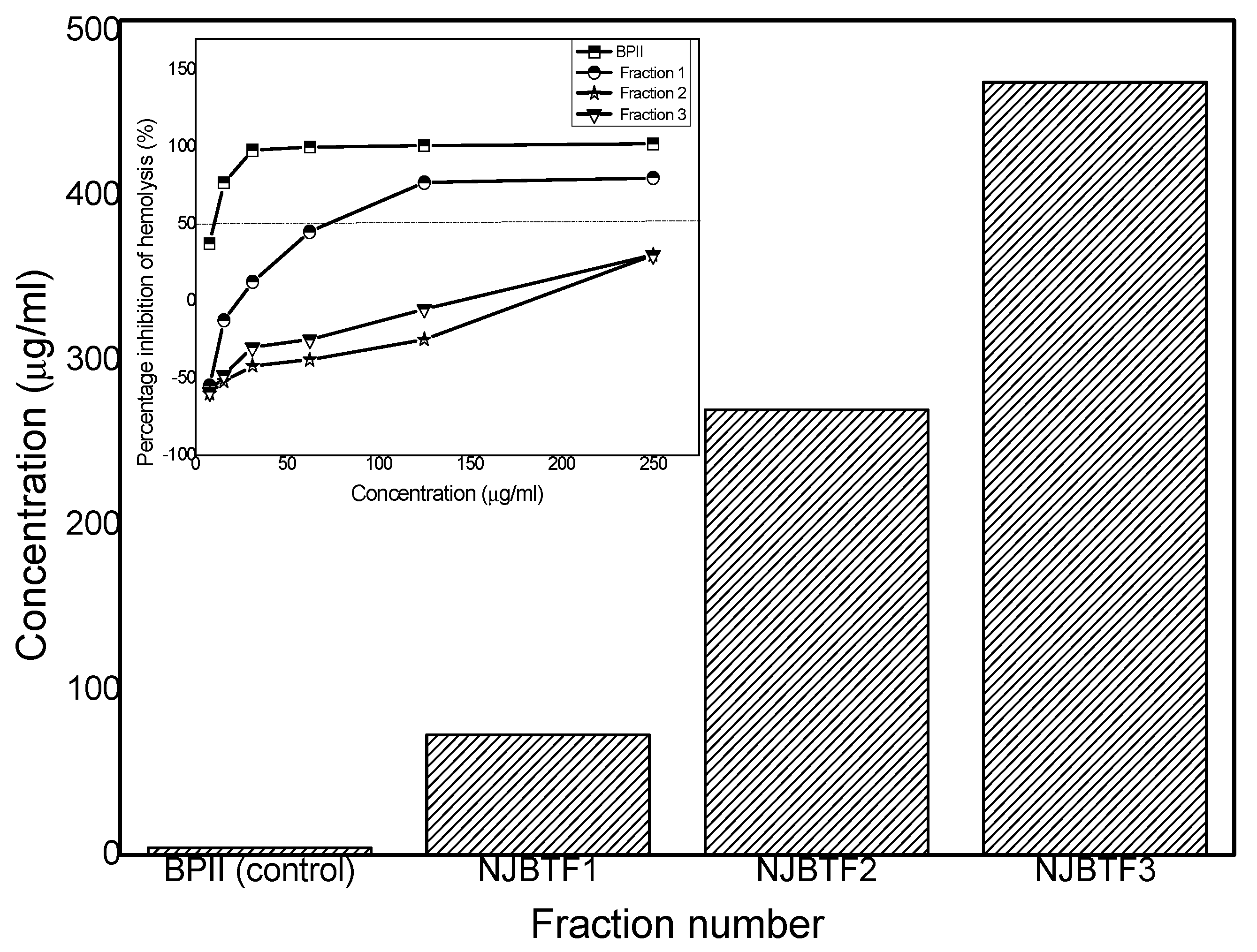
3.5. Complement Fixation
3.6. Monosaccharide Composition and Linkages
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| C | (g/mL or Rayleigh fringe units), concentration |
| ICH50 | concentration of the polysaccharide samples required to inhibit 50% lysis |
| Lp | persistence length (measure of flexibility) (cm) |
| Mw,app, | apparent weight-average molecular weight (Da) |
| Mw | weight average molecular weight (Da) |
| MSM | Scheraga-Mandelkern molecular weight (Da) |
| ML | mass per unit length (Da. cm−1) |
| NJBTF1 | fraction 1 (high molecular weight fraction) |
| NJBTF2 | fraction 2 (medium molecular weight fraction) |
| NJBTF3 | fraction 3 (low molecular weight fraction) |
| H | viscosity (Poise) |
| [η] | intrinsic viscosity (ml/g) |
| ῡ | partial specific volume |
| s20,w | weight average sedimentation coefficient, normalised to standard solvent conditions (density and viscosity of water at 20.0 °C) |
| so20,w, s20,w | corrected for non-ideality (extrapolation to c = 0) |
References
- Niture, S.K.; Refai, L. Plant pectin: A potential source for cancer suppression. Am. J. Pharmacol. Toxicol. 2013, 8, 9. [Google Scholar] [CrossRef] [Green Version]
- Dutta, P.K.; Tripathi, S.; Mehrotra, G.K.; Dutta, J. Perspectives for chitosan based antimicrobial films in food applications. Food Chem. 2009, 114, 1173–1182. [Google Scholar] [CrossRef]
- Espitia, P.J.P.; Wu, W.X.; Avena-Bustillos, R.J.; Soares, N.F.; McHugh, T.H. Edible films from pectin: Physical-mechanical and antimicrobial properties-A review. Food Hydrocoll. 2014, 35, 287–296. [Google Scholar] [CrossRef]
- Beneke, C.E.; Viljoen, A.M.; Hamman, J.H. Polymeric plant-derived excipients in drug delivery. Molecules 2009, 14, 2602–2620. [Google Scholar] [CrossRef]
- Huang, H.J.; Yuan, W.K.; Chen, X.D. Microencapsulation Based on Emulsification for Producing Pharmaceutical Products: A Literature Review. Dev. Chem. Eng. Miner. Process. 2006, 14, 515–544. [Google Scholar] [CrossRef]
- Villanova, J.C.O.; Ayres, E.; Oréfice, R.L. Design, characterization and preliminary in vitro evaluation of a mucoadhesive polymer based on modified pectin and acrylic monomers with potential use as a pharmaceutical excipient. Carbohydr. Polym. 2015, 121, 372–381. [Google Scholar] [CrossRef]
- Hsieh, Y.S.Y.; Liao, S.F.; Yang, W.B. Biologically Active Polysaccharides in Medicinal Plants. N. Z. J. For. Sci. 2009, 39, 217–233. [Google Scholar] [CrossRef]
- Giosafatto, C.V.L.; Mariniello, L.; Ring, S. Extraction and Characterization of Foeniculum vulgare Pectins and Their Use for Preparing Biopolymer Films in the Presence of Phaseolin Protein. J. Agric. Food Chem. 2007, 55, 1237–1240. [Google Scholar] [CrossRef]
- Fissore, E.N.; Ponce, N.M.; Stortz, C.A.; Rojas, A.M.; Gerschenson, L.N. Characterisation of Fiber Obtained from Pumpkin (cucumis moschata duch.) Mesocarp Through Enzymatic Treatment. Food Sci. Technol. Int. 2007, 13, 141–151. [Google Scholar] [CrossRef]
- Guillon, F.; Champ, M. Structural And Physical Properties of Dietary Fibres, and Consequences of Processing on Human Physiology. Food Res. Int. 2000, 33, 233–245. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, P.; Zhang, H. Pectin in cancer therapy: A review. Trends Food Sci. Technol. 2015, 44, 258–271. [Google Scholar] [CrossRef]
- Grønhaug, T.E.; Ghildyal, P.; Barsett, H.; Michaelsen, T.E.; Morris, G.; Diallo, D.; Inngjerdingen, M.; Paulsen, B.S. Bioactive Arabinogalactans from the Leaves of OPILIA Celtidifolia Endl. ex Walp. (Opiliaceae). Glycobiology 2010, 20, 1654–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobo, V.N.; Maróstica, M.R.; Zazueta, J.J.; Gallegos, J.A. Physicochemical, technological properties, and health-benefits of Cucurbita moschata Duchense vs. Cehualca: A Review. Food Res. Int. 2011, 44, 2587–2593. [Google Scholar] [CrossRef]
- Asgary, S.; Moshtaghian, S.J.; Setorki, M.; Kazemi, S.; Rafieian-Kopaei, M.; Adelnia, A.; Shamsi, F. Hypoglycaemic and Hypolipidemic Effects of Pumpkin (Cucurbita pepo L.) on Alloxan-Induced Diabetic Rats. Afr. J. Pharm. Pharmacol. 2011, 5, 2620–2626. [Google Scholar] [CrossRef]
- Nosáľová, G.; Prisenžňáková, Ľ.; Košťálová, Z.; Ebringerová, A.; Hromádková, Z. Suppressive effect of pectic polysaccharides from Cucurbita pepo L. var. Styriaca on citric acid-induced cough reflex in guinea pigs. Fitoterapia 2011, 82, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Košťálová, Z.; Hromádková, Z.; Ebringerová, A. Chemical evaluation of seeded fruit biomass of oil pumpkin (Cucurbita pepo L. var. Styriaca). Chem. Pap. 2009, 63, 406–413. [Google Scholar] [CrossRef]
- Quanhong, L.; Caili, F.; Yukui, R.; Guanghui, H.; Tongyi, C. Effects of Protein-Bound Polysaccharide Isolated from Pumpkin on Insulin in Diabetic Rats. Plant Foods Hum. Nutr. 2005, 60, 13–16. [Google Scholar] [CrossRef]
- Li, X.; Zhao, R.; Zhou, H.L.; Wu, D.H. Deproteinization of Polysaccharide from the Stigma Maydis by Sevag Method. Adv. Mater. Res. 2012, 340, 416–420. [Google Scholar] [CrossRef]
- Sam, K.C.C. Compositional analysis of foods. In Food Analysis, 2nd ed.; Neilson, S., Ed.; Plenum Publishers: New York, NY, USA, 2003; p. 136. [Google Scholar]
- Theisen, A.; Johann, C.; Deacon, M.P.; Harding, S.E. Refractive Increment Data-Book for Polymer and Biomolecular Scientists; Nottingham University Press: Nottingham, UK, 2000. [Google Scholar]
- Harding, S.E. The intrinsic viscosity of Biological Macromolecules. Progress in Measurement, Interaction and Application to Structure in Dilute Solution. Prog. Biophys. Mol. Biol. 1997, 68, 207–262. [Google Scholar] [CrossRef]
- Schuck, P. Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and lamm equation modeling. Biophys. J. 2000, 78, 1606–1619. [Google Scholar] [CrossRef] [Green Version]
- Schuck, P.; Harding, S.E.; Gillis, R.B.; Besong, T.M.D.; Almutairi, F.; Adams, G.G.; Rowe, A.J. SEDFIT-MSTAR: Molecular weight and molecular weight distribution analysis of polymers by sedimentation equilibrium in the ultracentrifuge. Analyst 2014, 139, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheraga, H.A.; Mandelkern, L. Consideratioi of the hydrodynamic properties of proteins. J. Am. Chem. Soc. 1953, 75, 175–184. [Google Scholar] [CrossRef]
- Ortega, A. Metodologías Computacionales Para Propiedades en Disolución de Macromoléculas Rígidas y Flexibles. Ph.D. Thesis, Universidad de Murcia, Murcia, Spain, 2005. [Google Scholar]
- Ortega, A.; Garcı´a de la Torre, J. Equivalent radii and ratios of radii from solution properties as indicators of macromolecular conformation, shape, and flexibility. Biomacromolecules 2007, 8, 2464–2475. [Google Scholar] [CrossRef] [PubMed]
- Bushin, S.; Tsvetkov, V.; Lysenko, E.; Emelianov, V. The sedimentation diffusion and viscometric analysis of the conformation properties and molecular rigidity of ladder-like polyphenyl siloxane in solution. Vysok. Soedin 1981, 23A, 2494–2503. [Google Scholar] [CrossRef]
- Bohdanecky, M. New method for estimating the parameters of the wormlike chain model from the intrinsic viscosity of stiff-chain polymers. Macromolecules 1983, 16, 1483–1492. [Google Scholar] [CrossRef]
- Yamakawa, H.; Fujii, M. Translational friction coefficient of wormlike chains. Macromolecules 1973, 6, 407–415. [Google Scholar] [CrossRef]
- Kök, M.S.; Abdelhameed, A.S.; Ang, S.; Morris, G.A.; Harding, S.E. A novel global hydrodynamic analysis of the molecular flexibility of the dietary fibre polysaccharide konjac glucomannan. Food Hydrocoll. 2009, 23, 1910–1917. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.A.; Garcia de la Torre, J.; Ortega, A.; Castile, J.; Smith, A.; Harding, S.E. Molecular flexibility of citrus pectins by combined sedimentation and viscosity analysis. Food Hydrocoll. 2008, 22, 1435–1442. [Google Scholar] [CrossRef] [Green Version]
- Michaelsen, T.E.; Gilje, A.; Samuelsen, A.B.; Høgåsen, K.; Paulsen, B.S. Interaction Between Human Complement and a Pectin Type Polysaccharide Fraction, PMII, from the Leaves of Plantago major L. Scand. J. Immunol. 2000, 52, 483–490. [Google Scholar] [CrossRef]
- Chambers, R.E.; Clamp, J.R. An assessment of methanolysis and other factors used in the analysis of carbohydrate-containing materials. Biochem. J. 1971, 125, 1009–1018. [Google Scholar] [CrossRef] [Green Version]
- Sims, I.M.; Bacic, A. Extracellular polysaccharides from suspension cultures of Nicotiana plumbaginifolia. Phytochemistry 1995, 38, 1397–1405. [Google Scholar] [CrossRef]
- Fissore, E.N.; Ponce, N.M.; de Escalada Pla, M.; Stortz, C.A.; Rojas, A.M.; Gerschenson, L.N. Characterization of Acid-Extracted Pectin-Enriched Products Obtained from Red Beet (Beta vulgaris L. var. conditiva) and Butternut (Cucurbita moschata Duch ex Poiret). J. Agric.Food Chem. 2010, 58, 3793–3800. [Google Scholar] [CrossRef] [PubMed]
- Roura, S.I.; Valle, C.E.D.; Aguero, L.; Davidovich, L.A. Changes in Apparent Viscosity and Vitamin C Retention During Thermal Treatment Of Butternut Squash (Cucurbita Moschata Duch) Pulp: Effect of Ripening Stage. J. Food Qual. 2007, 30, 538–551. [Google Scholar] [CrossRef]
- Harding, S.E. Challenges for the modern analytical ultracentrifuge analysis of polysaccharides. Carbohydr. Res. 2005, 340, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Dam, J.; Schuck, P. Calculating sedimentation coefficient distributions by direct modeling of sedimentation velocity concentration profiles. Methods Enzymol. 2004, 384, 185–212. [Google Scholar] [CrossRef]
- Creeth, J.M.; Pain, R.H. The determination of molecular weights of biological macromolecules by ultracentrifuge methods. Prog. Biophys. Mol. Biol. 1967, 17, 217–277. [Google Scholar] [CrossRef]
- Kabat, E.A.; Bezer, A.E. The effect of variation in molecular weight on the antigenicity of dextran in man. Arch. Biochem. Biophys. 1958, 78, 306–318. [Google Scholar] [CrossRef]
- Du, B.; Song, Y.; Hu, X.; Liao, X.; Ni, Y.; Li, Q. Oligosaccharides prepared by acid hydrolysis of polysaccharides from pumpkin (Cucurbita moschata) pulp and their prebiotic activities. Int. J. Food Sci. Technol. 2011, 46, 982–987. [Google Scholar] [CrossRef]
- Yang, X.; Zhao, Y.; Lv, Y. Chemical Composition and Antioxidant Activity of an Acidic Polysaccharide Extracted from Cucurbita moschata Duchesne ex Poiret. J. Agric. Food Chem. 2007, 55, 4684–4690. [Google Scholar] [CrossRef]
- Izydorczyk, M.; Cui, S.W.; Wang, Q. Polysaccharide gums: Structures, functional properties, and applications. In Food Carbohydrates: Chemistry, Physical Properties, and Applications; Cui, S.W., Ed.; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Voragen, A.G.J.; Coenen, G.J.; Verhoef, R.P.; Schols, H.A. Pectin, a versatile polysaccharide present in plant cell walls. Struct. Chem. 2009, 20, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, Y.; Luo, J. Structure elucidation of a pectin from Dendrobium nobile Lindl and its immunological activity. Biotechnol. Biotechno. Equip. 2018, 32, 744–750. [Google Scholar] [CrossRef] [Green Version]
- Öner, E.T.; Hernández, L.; Combie, J. Review of Levan polysaccharide: From a century of past experiences to future prospects. Biotechnol. Adv. 2016, 34, 827–844. [Google Scholar] [CrossRef] [PubMed]
- Dahech, I.; Belghith, K.S.; Hamden, K.; Feki, A.; Belghith, H.; Mejdoub, H. Oral administration of levan polysaccharide reduces the alloxan-induced oxidative stress in rats. Int. J. Biol. Macromol. 2011, 49, 942–947. [Google Scholar] [CrossRef]
- Aspinall, G.O.; Percival, E.; Rees, D.A.; Rennie, M. Oligosaccharides, Polysaccharides and Related Compounds. In Rodd’s Chemistry of CarbonCompounds; Coffey, S., Ed.; Elsevier Publishing Company: Amesterdam, The Netherlands, 1967; Volume 1. [Google Scholar]
- Banguela, A.; Arrieta, J.G.; Rodríguez, R.; Trujillo, L.E.; Menéndez, C.; Hernández, L. High levan accumulation in transgenic tobacco plants expressing the Gluconacetobacter diazotrophicus levansucrase gene. J. Biotechnol. 2011, 154, 93–98. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [η]H (mL/g) | [η]K (mL/g) | [η]SC (mL/g) | |
|---|---|---|---|
| NJBTF1 | 53.6 ± 1.3 | 53.6 ± 1.2 | 53.6 ± 1.3 |
| NJBTF2 | 33.9 ± 0.6 | 34.0 ± 0.5 | 34.0 ± 0.6 |
| NJBTF3 | 23.0 ± 0.7 | 23.0 ± 0.7 | 23.0 ± 0.7 |
| Peak 1 | Peak 2 | Peak 3 | Weight Average | ||||
|---|---|---|---|---|---|---|---|
| so20,w (S) | wt frac (%) | so20,w (S) | wt frac (%) | so20,w (S) | wt frac (%) | so20,w (S) | |
| NJBTF1 | 1.3 ± 0.1 | 81 | 3.5 ± 0.2 | 19 | - | - | 1.7 ± 0.2 |
| NJBTF2 | 1.3 + 0.1 | 93 | 2.9 ± 0.6 | 5 | 4.1 ± 0.8 | 2 | 1.4 ± 0.2 |
| NJBTF3 | 1.2 + 0.2 | 83 | 1.7 ± 0.7 | 17 | - | - | 1.3 ± 0.2 |
| Mw (kDa) | MSM (kDa) | |
|---|---|---|
| NJBTF1 | 90 ± 5 | 30 ± 4 |
| NJBTF2 | 30 ± 2 | 20 ± 2 |
| NJBTF3 | 19 ± 2 | 12 ± 2 |
| NJBTF 1 | NJBTF 2 | NJBTF3 | ||
|---|---|---|---|---|
| Neutral monosaccharides | Arabinose | 3.6 | 4.4 | 2.8 |
| Rhamnose | 11.7 | 4.8 | 2.8 | |
| Fucose | tr | 1.2 | 1.3 | |
| Xylose | - | 0.4 | 0.2 | |
| Galactose | 5.0 | 5.4 | 5.0 | |
| Glucose | 11.4 | 8.8 | 13.1 | |
| Acidic monosaccharides | Glucuronic acid | 7.3 | - | - |
| Galacturonic acid | 61.0 | 75.0 | 74.8 |
| Polymer | Retention Time (min) | Primary Fragments-Mass to Charge Ratio (m/z) | Identity of Origin | Relative Amount |
|---|---|---|---|---|
| PECTIN | 12.99 | 45, 118, 161 | T-Araf | 3.0 |
| 13.91 | 118, 131, 162, 175 | T-Rha | 1.8 | |
| 14.64 | 118, 131, 162, 175 | T-Fuc | Tr. | |
| 15.34 | 45, 190, 161 | 1,2 Araf | 0.3 | |
| 15.40 | 45, 118, 233 | 1,3-Araf | 0.5 | |
| 16.19 | 118, 189 | 1,5-Araf | Tr. | |
| 16.23 | 131, 190, | 1,2-Rha | 7.7 | |
| 17.28 | 47, 118, 162, 163, 207 | T-GlcA | 1.0 | |
| 17.83 | 47, 118, 162, 163, 207 | T-GalA | 2.0 | |
| 18,37 | 131, 262 | 1,2,3-Rha | 1.8 | |
| 18.47 | 118 | 1,3,5-Araf | 0.4 | |
| 18.58 | 190, 203 | 1,2,4-Rha | 2.0 | |
| 19.73 | 47, 118, 162, 235 | 1,4 GalA | 65.9 | |
| 19.89 | 47, 118, 162, 235 | 1,4 GlcA | 1.8 | |
| 20.13 | 45, 118, 161, 234, 277 | 1,3-Gal | 3.0 | |
| 21.07 | 118, 162, 189, 233 | 1,6-Gal | 0.3 | |
| 21.88 | 47, 190 235 | 1,2,4-GalA | 0.8 | |
| 23.33 | 118, 189, 2234 | 1,3,6-Gal | 2.3 | |
| LEVAN | 16.16; 16.32 | 45, 161, 162 | T-Fruf | 18 |
| 17.28 | 45, 118, 161, 162, 205 | T-Glc | 1 | |
| 19.37; 19.55 | 45, 162, 189 | 2,6-Fruf | 245 | |
| 22.61 | 189, 190 | 1,2,6-Fruf | 16 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiwani, S.I.; Gillis, R.B.; Besong, D.; Almutairi, F.; Erten, T.; Kök, M.S.; Harding, S.E.; Paulsen, B.S.; Adams, G.G. Isolation and Biophysical Characterisation of Bioactive Polysaccharides from Cucurbita Moschata (Butternut Squash). Polymers 2020, 12, 1650. https://doi.org/10.3390/polym12081650
Jiwani SI, Gillis RB, Besong D, Almutairi F, Erten T, Kök MS, Harding SE, Paulsen BS, Adams GG. Isolation and Biophysical Characterisation of Bioactive Polysaccharides from Cucurbita Moschata (Butternut Squash). Polymers. 2020; 12(8):1650. https://doi.org/10.3390/polym12081650
Chicago/Turabian StyleJiwani, Shahwar Imran, Richard B. Gillis, David Besong, Fahad Almutairi, Tayyibe Erten, M. Samil Kök, Stephen E. Harding, Berit S. Paulsen, and Gary G. Adams. 2020. "Isolation and Biophysical Characterisation of Bioactive Polysaccharides from Cucurbita Moschata (Butternut Squash)" Polymers 12, no. 8: 1650. https://doi.org/10.3390/polym12081650