Genome-Wide Characterization and Expression of Two-Component System Genes in Cytokinin-Regulated Gall Formation in Zizania latifolia

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Exogenous Cytokinin Treatment on Z. latifolia Gall Formation
2.2. Identification of TCS Genes in Z. latifolia
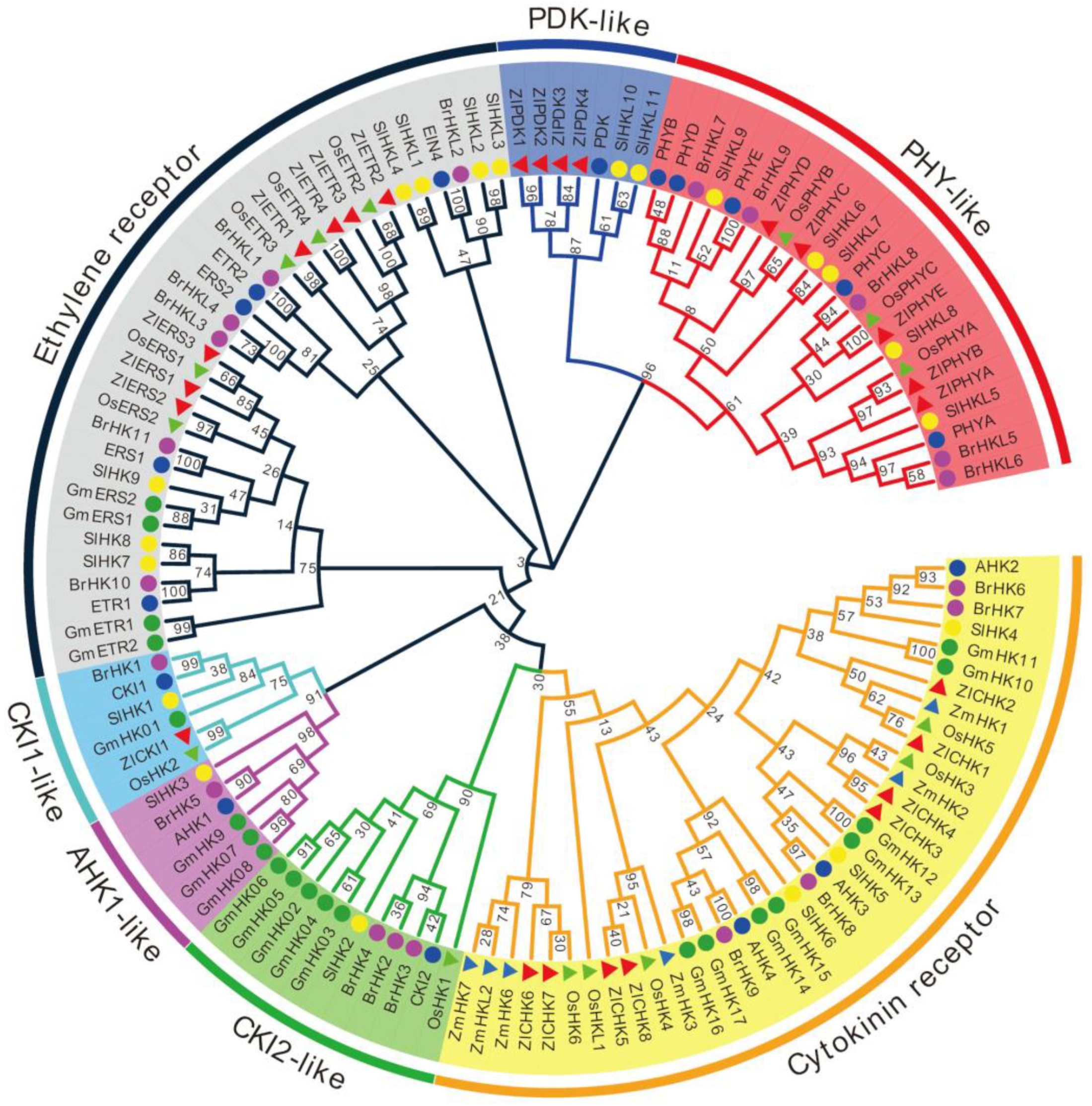
2.3. HK Proteins in Z. latifolia
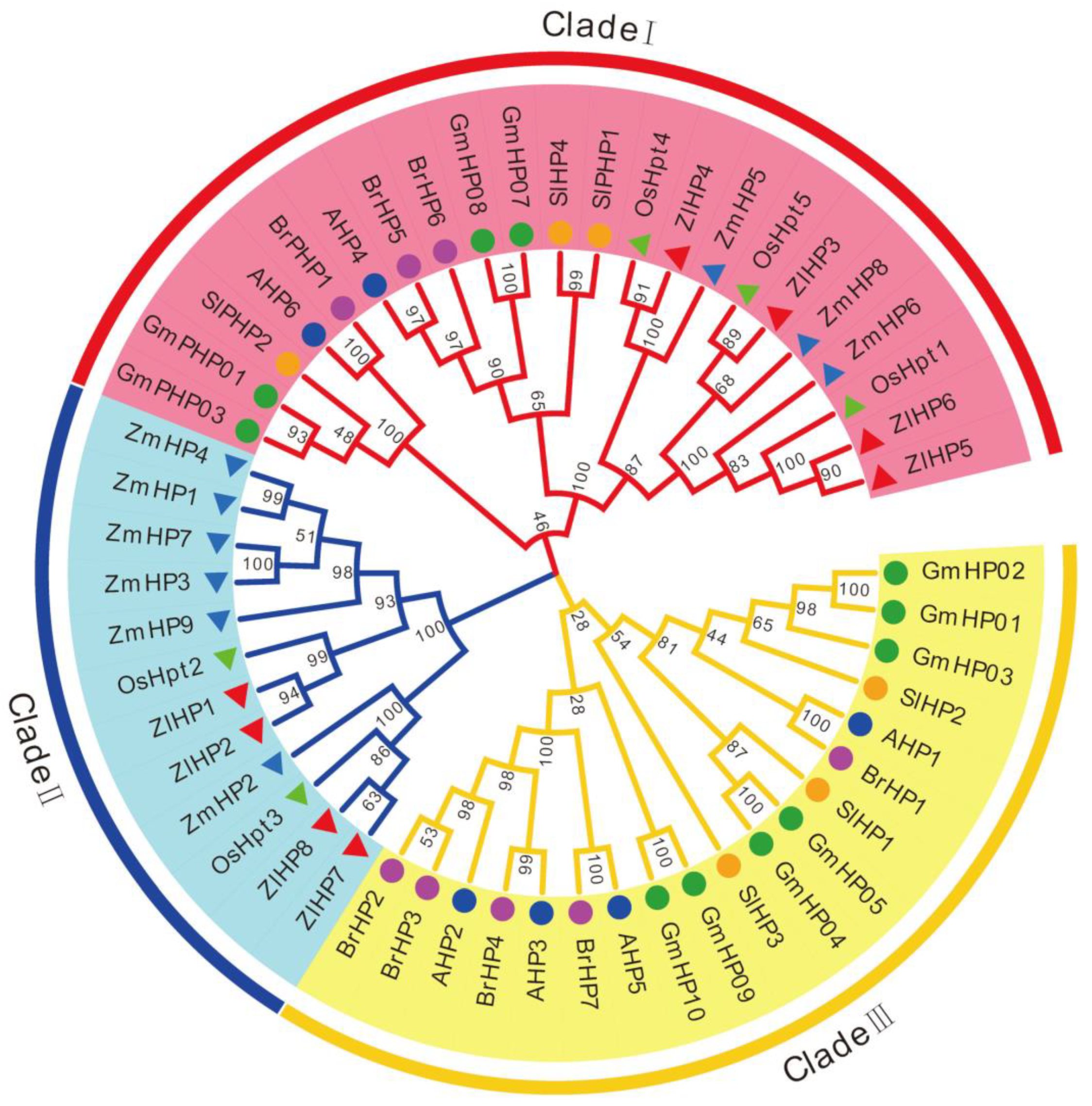
2.4. HP Proteins in Z. latifolia
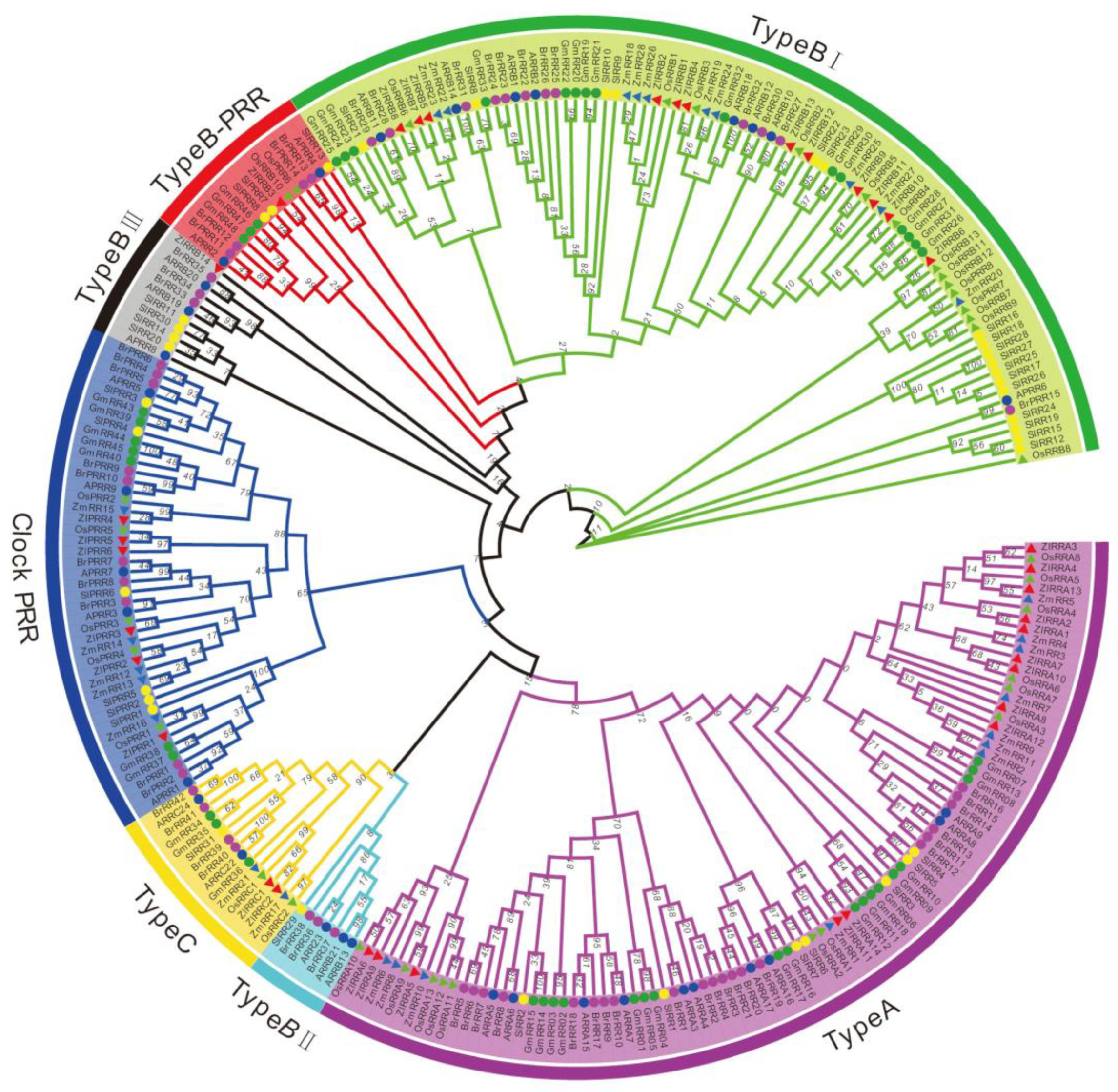
2.5. RR Proteins in Z. latifolia
2.6. Phylogenetic Relationship of TCS Members in Z. latifolia
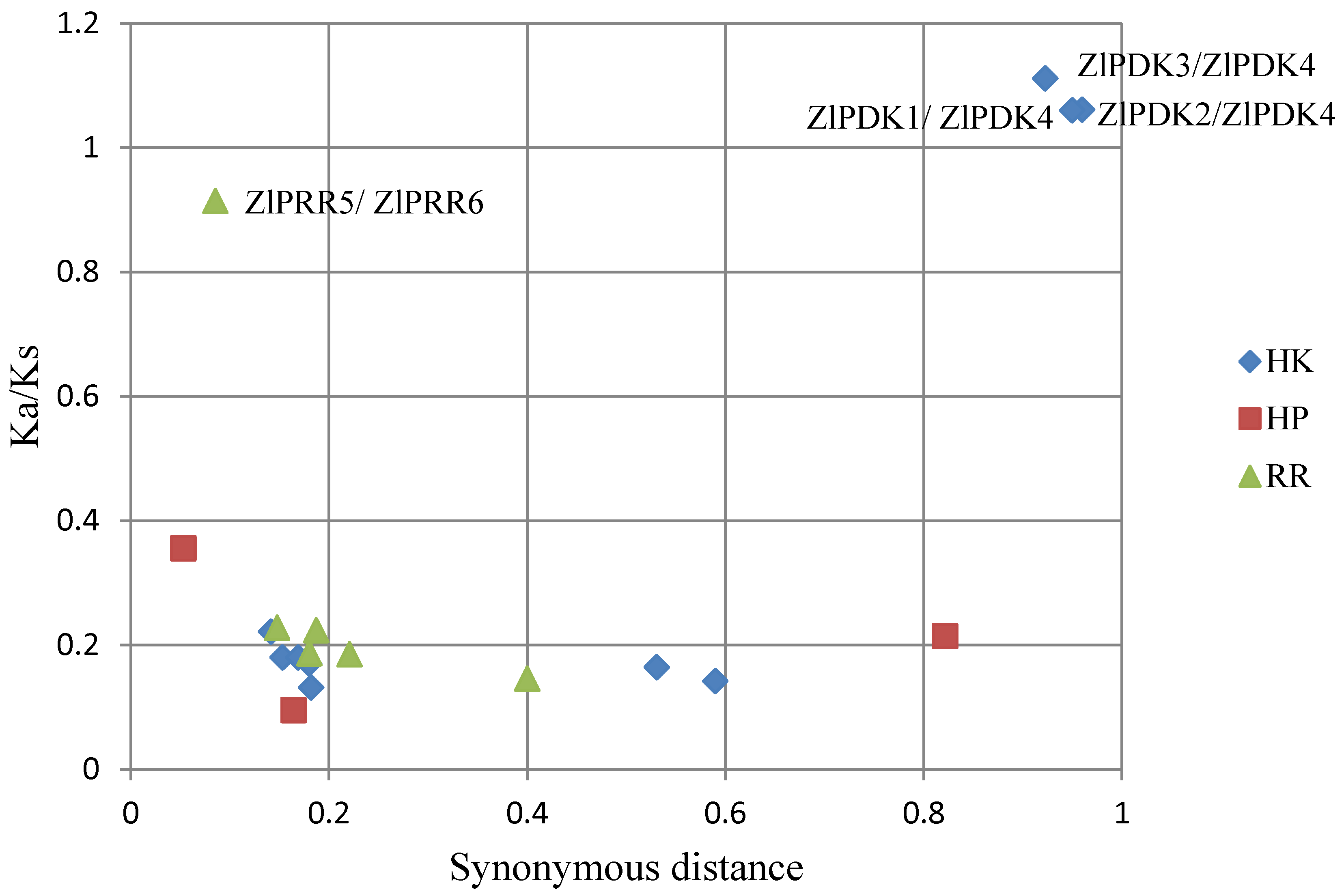
2.7. Strong Purifying Selection for TCS Genes in Z. latifolia
2.8. TCS Gene Expression During Z. latifolia Shoots Swelling
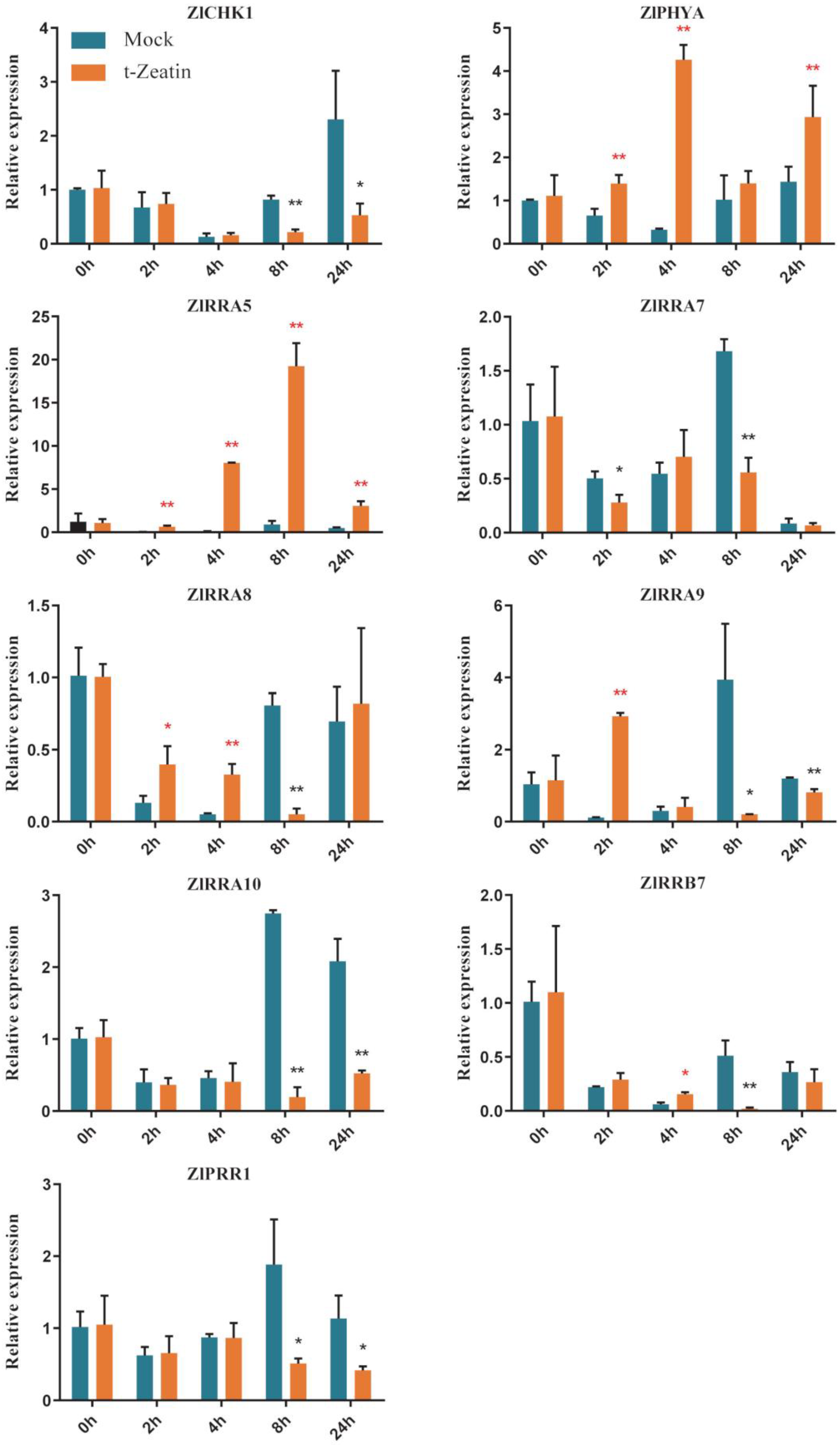
2.9. TCS Gene Response to Exogenous Cytokinin
3. Materials and Methods
3.1. Plant Growth and Treatments
3.2. Identification of TCS Genes in Z. latifolia
3.3. Genes Structure Prediction, Protein Sequence Analysis, and Phylogenetic Analysis
3.4. Calculation of Nonsynonymous (Ka) to Synonymous (Ks) Substitutions
3.5. Transcriptome Analysis and Expression of TCS Genes
3.6. RNA Isolation and Real-Time PCR
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guo, L.; Qiu, J.; Han, Z.; Ye, Z.; Chen, C.; Liu, C.; Xin, X.; Ye, C.Y.; Wang, Y.Y.; Xie, H.; et al. A host plant genome (Zizania latifolia) after a century-long endophyte infection. Plant J. Cell Mol. Biol. 2015, 83, 600–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.B.; Li, S.M.; Peng, J.; Ke, W.D. Zizania latifolia Turcz. cultivated in China. Genet. Resour. Crop Evol. 2007, 54, 1211–1217. [Google Scholar] [CrossRef]
- Chilton, M.D.; Drummond, M.H.; Merio, D.J.; Sciaky, D.; Montoya, A.L.; Gordon, M.P.; Nester, E.W. Stable incorporation of plasmid DNA into higher plant cells: The molecular basis of crown gall tumorigenesis. Cell 1977, 11, 263–271. [Google Scholar] [CrossRef]
- Barry, G.F.; Rogers, S.G.; Fraley, R.T.; Brand, L. Identification of a cloned cytokinin biosynthetic gene. Proc. Natl. Acad. Sci. USA 1984, 81, 4776–4780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crespi, M.; Messens, E.; Caplan, A.B.; van Montagu, M.; Desomer, J. Fasciation induction by the phytopathogen Rhodococcus fascians depends upon a linear plasmid encoding a cytokinin synthase gene. EMBO J. 1992, 11, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, O.; Oden, S.; Mol, A.; Vereecke, D.; Goethals, K.; El Jaziri, M.; Prinsen, E. Biosynthesis of auxin by the gram-positive phytopathogen Rhodococcus fascians is controlled by compounds specific to infected plant tissues. Appl. Environ. Microbiol. 2005, 71, 1169–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, Y.S.; Thrower, L. The host-parasite relationship between Zizania caduciflora Turcz. and Ustilago esculebta p. Heen. IV. Growth substances in the host-parasite combination. New Phytol. 1980, 85, 225–233. [Google Scholar] [CrossRef]
- Lin, Y.L.; Lin, C.H. Involvement of tRNA bound cytokinin on the gall formation in Zizania. J. Exp. Bot. 1990, 41, 277–281. [Google Scholar] [CrossRef]
- Werner, T.; Schmulling, T. Cytokinin action in plant development. Curr. Opin. Plant Biol. 2009, 12, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, C.; Yang, H.; Jiao, Y. Cytokinin pathway mediates APETALA1 function in the establishment of determinate floral meristems in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 6840–6845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, K.D.; Takata, N.; Johansson, M.; Jurca, M.; Novak, O.; Henykova, E.; Liverani, S.; Kozarewa, I.; Strnad, M.; Millar, A.J.; et al. Circadian clock components control daily growth activities by modulating cytokinin levels and cell division-associated gene expression in Populus trees. Plant Cell Environ. 2018, 41, 1468–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, I.; Sheen, J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Schaller, G.E.; Kieber, J.J.; Shiu, S.H. Two-component signaling elements and histidyl-aspartyl phosphorelays. Arab. Book 2008, 6, e0112. [Google Scholar] [CrossRef] [Green Version]
- Yamada, H.; Suzuki, T.; Terada, K.; Takei, K.; Ishikawa, K.; Miwa, K.; Yamashino, T.; Mizuno, T. The Arabidopsis AHK4 Histidine Kinase is a Cytokinin-Binding Receptor that Transduces Cytokinin Signals Across the Membrane. Plant Cell Physiol. 2001, 42, 1017–1023. [Google Scholar] [CrossRef]
- Wurgler-Murphy, S.M.; Saito, H. Two-component signal transducers and MAPK cascades. Trends Biochem. Sci. 1997, 22, 172–176. [Google Scholar] [CrossRef]
- Hwang, I. Two-Component Signal Transduction Pathways in Arabidopsis. Plant Physiol. 2002, 129, 500–515. [Google Scholar] [CrossRef] [Green Version]
- Grefen, C.; Harter, K. Plant two-component systems: Principles, functions, complexity and cross talk. Planta 2004, 219, 733–742. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; De Peer, Y.V.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Mizuno, T. Two-component phosphorelay signal transduction systems in plants: From hormone responses to circadian rhythms. Biosci. Biotechnol. Biochem. 2005, 69, 2263–2276. [Google Scholar] [CrossRef]
- Pareek, A.; Singh, A.; Kumar, M.; Kushwaha, H.R.; Lynn, A.M.; Singla-Pareek, S.L. Whole-genome analysis of Oryza sativa reveals similar architecture of two-component signaling machinery with Arabidopsis. Plant Physiol 2006, 142, 380–397. [Google Scholar] [CrossRef] [Green Version]
- Ishida, K.; Niwa, Y.; Yamashino, T.; Mizuno, T. A genome-wide compilation of the two-component systems in Lotus japonicus. DNA Res. 2009, 16, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Yamashino, T.; Nakanishi, H.; Mizuno, T. Classification of the Genes Involved in the Two-Component System of the MossPhyscomitrella patens. Biosci. Biotechnol. Biochem. 2010, 74, 2542–2545. [Google Scholar] [CrossRef] [PubMed]
- Mochida, K.; Yoshida, T.; Sakurai, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S. Genome-wide analysis of two-component systems and prediction of stress-responsive two-component system members in soybean. DNA Res. 2010, 17, 303–324. [Google Scholar] [CrossRef] [Green Version]
- Chu, Z.X.; Ma, Q.; Lin, Y.X.; Tang, X.L.; Zhou, Y.Q.; Zhu, S.W.; Fan, J.; Cheng, B.J. Genome-wide identification, classification, and analysis of two-component signal system genes in maize. Genet. Mol. Res. 2011, 10, 3316–3330. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, M.; Kong, L.; Lv, Y.; Zou, M.; Lu, G.; Cao, J.; Yu, X. Genome-Wide Identification, Phylogeny, Duplication, and Expression Analyses of Two-Component System Genes in Chinese Cabbage (Brassica rapa ssp. pekinensis). DNA Res. 2014, 21, 379–396. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Liu, X.; Ye, L.; Pan, C.; Chen, L.; Zou, T.; Lu, G. Genome-Wide Identification and Expression Analysis of Two-Component System Genes in Tomato. Int. J. Mol. Sci. 2016, 17, 1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Liu, X.; Zou, T.; Pan, C.; Qin, L.; Chen, L.; Lu, G. Genome-Wide Identification of Two-Component System Genes in Cucurbitaceae Crops and Expression Profiling Analyses in Cucumber. Front. Plant Sci. 2016, 7, 899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Yang, X.; Zhang, Y.; Wang, S.; Ge, Q.; Li, Q.; Wang, C.; Shi, Q.; Ren, Z.; Wang, L. Genome-Wide Identification of Two-Component Signal Transduction System Genes in Melon (Cucumis melon L.). Agric. Sci. 2018, 9, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Wang, S.; Wang, X.; Yang, X.; Li, Q.; Wang, C.; Chen, C.; Shi, Q.; Ren, Z.; Wang, L. Genome-wide characterization of two-component system (TCS) genes in melon (Cucumis melo L.). Plant Physiol. Biochem. 2020, 151, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.D.; Yan, N.; Wang, Z.H.; Zhang, X.H.; Zhang, J.Z.; Xue, H.M.; Wang, L.X.; Zhan, Q.; Xu, Y.P.; Guo, D.P. RNA-seq analysis provides insight into reprogramming of culm development in Zizania latifolia induced by Ustilago esculenta. Plant Mol. Biol. 2017, 95, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Heyl, A.; Brault, M.; Frugier, F.; Kuderova, A.; Lindner, A.C.; Motyka, V.; Rashotte, A.M.; Schwartzenberg, K.V.; Vankova, R.; Schaller, G.E. Nomenclature for members of the two-component signaling pathway of plants. Plant Physiol. 2013, 161, 1063–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-Component Signal Transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueguchi, C.; Sato, S.; Kato, T.; Tabata, S. The AHK4 Gene Involved in the Cytokinin-Signaling Pathway as a Direct Receptor Molecule in Arabidopsis thaliana. Plant Cell Physiol. 2001, 42, 751–755. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Higuchi, M.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Kato, T.; Tabata, S.; Shinozaki, K.; Kakimoto, T. Identification of CRE1 as a cytokinin receptor from Arabidopsis. Nature 2001, 409, 1060–1063. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Miwa, K.; Ishikawa, K.; Yamada, H.; Aiba, H.; Mizuno, T. The Arabidopsis Sensor His-kinase, AHK4, Can Respond to Cytokinins. Plant Cell Physiol. 2001, 42, 107–113. [Google Scholar] [CrossRef]
- Spíchal, L.; Rakova, N.Y.; Riefler, M.; Mizuno, T.; Romanov, G.A.; Strnad, M.; Schmülling, T. Two Cytokinin Receptors of Arabidopsis thaliana, CRE1/AHK4 and AHK3, Differ in their Ligand Specificity in a Bacterial Assay. Plant Cell Physiol. 2004, 45, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Romanov, G.A.; Lomin, S.N.; Schmulling, T. Biochemical characteristics and ligand-binding properties of Arabidopsis cytokinin receptor AHK3 compared to CRE1/AHK4 as revealed by a direct binding assay. J. Exp. Botany 2006, 57, 4051–4058. [Google Scholar] [CrossRef] [Green Version]
- Stolz, A.; Riefler, M.; Lomin, S.N.; Achazi, K.; Romanov, G.A.; Schmülling, T. The specificity of cytokinin signalling in Arabidopsis thaliana is mediated by differing ligand affinities and expression profiles of the receptors. Plant J. 2011, 67, 157–168. [Google Scholar] [CrossRef]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Arkhipov, D.V.; Osolodkin, D.I.; Schmulling, T.; Romanov, G.A. Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands. J. Exp. Botany 2015, 66, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Bartrina, I.; Jensen, H.; Novak, O.; Strnad, M.; Werner, T.; Schmulling, T. Gain-of-Function Mutants of the Cytokinin Receptors AHK2 and AHK3 Regulate Plant Organ Size, Flowering Time and Plant Longevity. Plant Physiol. 2017, 173, 1783–1797. [Google Scholar] [CrossRef] [Green Version]
- Rashotte, A.M.; Carson, S.D.B.; To, J.P.C.; Kieber, J.J. Expression Profiling of Cytokinin Action in Arabidopsis. Plant Physiol. 2003, 132, 1998–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Hall, A.E.; O’Malley, R.; Bleecker, A.B. Canonical histidine kinase activity of the transmitter domain of the ETR1 ethylene receptor from Arabidopsis is not required for signal transmission. Proc. Natl. Acad. Sci. USA 2003, 100, 352–357. [Google Scholar] [CrossRef] [Green Version]
- Binder, B.M.; O’Malley, R.C.; Wang, W.; Moore, J.M.; Parks, B.M.; Spalding, E.P.; Bleecker, A.B. Arabidopsis seedling growth response and recovery to ethylene. A kinetic analysis. Plant Physiol. 2004, 136, 2913–2920. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Schaller, G.E. Requirement of the histidine kinase domain for signal transduction by the ethylene receptor ETR1. Plant Physiol. 2004, 136, 2961–2970. [Google Scholar] [CrossRef] [Green Version]
- Rockwell, N.C.; Su, Y.-S.; Lagarias, J.C. Phytochrome structure and signaling mechanisms. Annu. Rev. Plant Biol. 2006, 57, 837–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Y.; Offringa, R. PDK1 regulates auxin transport and Arabidopsis vascular development through AGC1 kinase PAX. Nat. Plants 2020, 6, 544–555. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, Q.; Wu, J.; Zhang, L.; Jiao, X.; Zhang, S.; Zhang, Z.; Sun, D.; Lu, T.; Sun, Y. Two rice authentic histidine phosphotransfer proteins, OsAHP1 and OsAHP2, mediate cytokinin signaling and stress responses in rice. Plant Physiol. 2014, 165, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fassler, J.S.; West, A.H. Histidine phosphotransfer proteins in fungal two-component signal transduction pathways. Eukaryot. Cell 2013, 12, 1052–1060. [Google Scholar] [CrossRef] [Green Version]
- Miyata, S.-i.; Urao, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of genes for two-component phosphorelay mediators with a single HPt domain in Arabidopsis thaliana. FEBS Lett. 1998, 437, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Imamura, A.; Ueguchi, C.; Mizuno, T. Histidine-Containing Phosphotransfer (HPt) Signal Transducers Implicated in His-to-Asp Phosphorelay in Arabidopsis. Plant Cell Physiol. 1999, 39, 1258–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urao, T.; Miyata, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Possible His to Asp phosphorelay signaling in an Arabidopsis two-component system. FEBS Lett. 2000, 478, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Sakurai, K.; Ueguchi, C.; Mizuno, T. Two Types of Putative Nuclear Factors that Physically Interactwith Histidine-Containing Phosphotransfer (Hpt) Domains, Signaling Mediators in His-to-Asp Phosphorelay, in Arabidopsisthaliana. Plant Cell Physiol. 2001, 42, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Niwa, Y.; Ito, S.; Nakamichi, N.; Mizoguchi, T.; Niinuma, K.; Yamashino, T.; Mizuno, T. Genetic linkages of the circadian clock-associated genes, TOC1, CCA1 and LHY, in the photoperiodic control of flowering time in Arabidopsis thaliana. Plant Cell Physiol. 2007, 48, 925–937. [Google Scholar] [CrossRef]
- Swapan, K.S.; Alak, C.D.; Raju, B. Morphogenesis of edible gall in Zizania latifolia (Griseb.) Turcz. ex Stapf due to Ustilago esculenta Henn. infection in India. Afr. J. Microbiol. Res. 2016, 10, 1215–1223. [Google Scholar] [CrossRef] [Green Version]
- Franklin, K.A.; Allen, T.; Whitelam, G.C. Phytochrome A is an irradiance-dependent red light sensor. Plant J. 2007, 50, 108–117. [Google Scholar] [CrossRef]
- Monte, E.; Alonso, J.M.; Ecker, J.R.; Zhang, Y.; Li, X.; Young, J.; Austin-Phillips, S.; Quail, P.H. Isolation and Characterization of phyC Mutants in Arabidopsis Reveals Complex Crosstalk between Phytochrome Signaling Pathways. Plant Cell 2003, 15, 1962–1980. [Google Scholar] [CrossRef] [Green Version]
- Devlin, P.F.; Patel, S.R.; Whitelam, G.C. Phytochrome E influences internode elongation and flowering time in Arabidopsis. Plant Cell 1998, 10, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Reed, J.W.; Nagpal, P.; Poole, D.S.; Furuya, M.; Chory, J. Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell 1993, 5, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Gahlaut, V.; Mathur, S.; Dhariwal, R.; Khurana, J.P.; Tyagi, A.K.; Balyan, H.S.; Gupta, P.K. A multi-step phosphorelay two-component system impacts on tolerance against dehydration stress in common wheat. Funct. Integr. Genom. 2014, 14, 707–716. [Google Scholar] [CrossRef]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, D302–D305. [Google Scholar] [CrossRef]
- Punta, M.; Coggill, P.; Eberhardt, R.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, S.; Ceric, G.; Clements, J.; et al. The Pfam protein families database. Nucleic Acids Res. 2011, 40, D290–D301. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef]
- Marchler-Bauer, A.; Lu, S.; Anderson, J.B.; Chitsaz, F.; Derbyshire, M.K.; DeWeese-Scott, C.; Fong, J.H.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; et al. CDD: A Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 2011, 39, D225–D229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, A.; Zhu, Q.; Chen, X.; Luo, J. GSDS: A gene structure display server. Yi Chuan 2007, 29, 1023–1026. [Google Scholar] [CrossRef]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef] [Green Version]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Hurst, L.D. The K a/K s ratio:diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
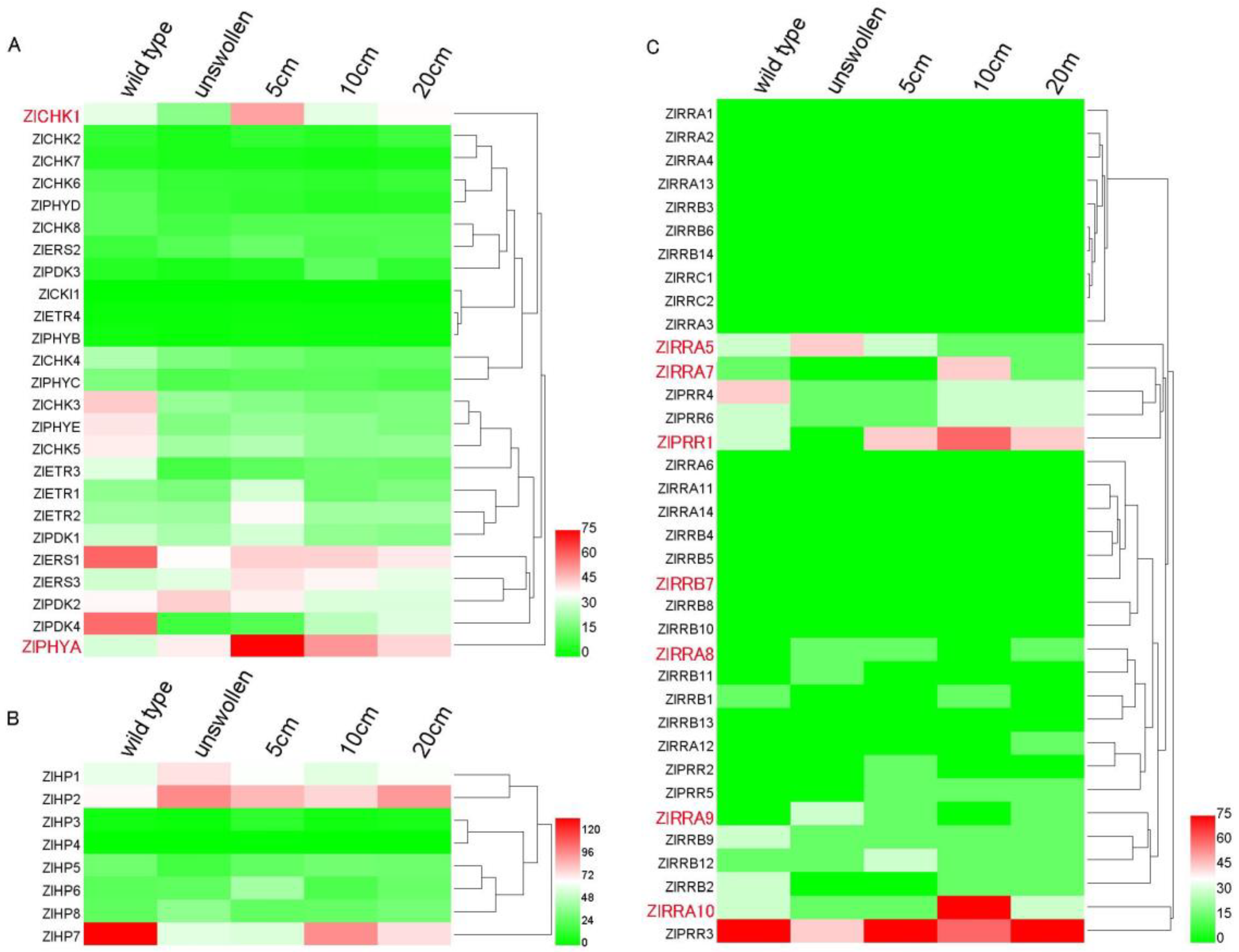{kind=link}
{kind=link}
| Gene ID | Gene Name | Character a | Gene Family | Counterpart | Score (bits) | Identity (%) | Length (aa) | Molecular Weight (kDa) | Isoelectric Point (PI) | Number of TM | Subcellular localization b |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Zlat_10043954 | ZlCHK1 | HisKA, HATPase_c, CHASE, REC, TM | Cytokinin Receptor | AHK2 | 532 | 65 | 609 | 67.56 | 7.26 | 4 | Endomembrane system |
| Zlat_10043955 | ZlCHK2 | HisKA, HATPase_c, REC | Cytokinin Receptor | AHK2 | 207 | 46 | 445 | 49.66 | 6.65 | 0 | Nucleus |
| Zlat_10033100 | ZlCHK3 | HisKA, HATPase_c, REC | Cytokinin Receptor | AHK3 | 673 | 61 | 570 | 63.2 | 6.72 | 0 | Nucleus |
| Zlat_10033475 | ZlCHK4 | HisKA, HATPase_c, CHASE, REC, TM | Cytokinin Receptor | AHK3 | 1045 | 63 | 861 | 95.89 | 8.55 | 1 | Nucleus |
| Zlat_10002572 | ZlCHK5 | HisKA, HATPase_c, CHASE, REC, TM | Cytokinin Receptor | AHK4 | 746 | 68 | 907 | 99.75 | 6.02 | 2 | Endomembrane system |
| Zlat_10008874 | ZlCHK6 | HisKA, HATPase_c, CHASE, REC, TM | Cytokinin Receptor | AHK4 | 202 | 72 | 430 | 48.69 | 5.58 | 1 | Organelle membrane |
| Zlat_10009428 | ZlCHK7 | HisKA, HATPase_c, CHASE, REC, TM | Cytokinin Receptor | AHK4 | 620 | 69 | 618 | 69.42 | 5.27 | 1 | Nucleus |
| Zlat_10007870 | ZlCHK8 | HisKA, HATPase_c, CHASE, REC, TM | Cytokinin Receptor | AHK4 | 335 | 55 | 716 | 78.44 | 5.41 | 1 | Nucleus |
| Zlat_10005586 | ZlCKI1 | HisKA, HATPase_c, REC | CKI1 | 330 | 36 | 810 | 89.62 | 5.81 | 0 | Nucleus | |
| Zlat_10007295 | ZlERS1 | HisKA, HATPase_c, GAF, TM | Ethylene Receptor | ETR1 | 900 | 74 | 636 | 70.93 | 6.86 | 3 | Endomembrane system |
| Zlat_10019946 | ZlERS2 | HisKA, HATPase_c, GAF | Ethylene Receptor | ETR1 | 380 | 65 | 314 | 34.48 | 5.29 | 0 | Nucleus |
| Zlat_10028651 | ZlERS3 | HisKA, HATPase_c, GAF, TM | Ethylene Receptor | ETR1 | 879 | 75 | 635 | 70.79 | 6.82 | 3 | Endomembrane system |
| Zlat_10043320 | ZlETR1 | HisKA, HATPase_c, GAF, REC, TM | Ethylene Receptor | ETR2 | 549 | 47 | 842 | 93.21 | 6.89 | 4 | Plasma membrane |
| Zlat_10010084 | ZlETR2 | HisKA, HATPase_c, GAF, REC, TM | Ethylene Receptor | EIN4 | 669 | 51 | 759 | 85.01 | 6.21 | 3 | Endomembrane system |
| Zlat_10043113 | ZlETR3 | HisKA, HATPase_c, GAF, REC, TM | Ethylene Receptor | EIN4 | 662 | 51 | 758 | 84.99 | 6.18 | 3 | Plasma membrane |
| Zlat_10044950 | ZlETR4 | HisKA, HATPase_c, GAF, REC | Ethylene Receptor | EIN4 | 302 | 39 | 507 | 54.79 | 7.52 | 0 | Nucleus |
| Zlat_10007862 | ZlPHYA | HisKA, HATPase_c, GAF, PAS | Phytochrome | PHYA | 1456 | 64 | 1128 | 125.1 | 5.78 | 0 | Nucleus |
| Zlat_10002747 | ZlPHYB | HisKA, HATPase_c, GAF, PAS | Phytochrome | PHYA | 1439 | 63 | 1129 | 125.24 | 5.83 | 0 | Nucleus |
| Zlat_10036932 | ZlPHYC | HisKA, HATPase_c, GAF, PAS | Phytochrome | PHYB | 1736 | 75 | 1190 | 130.73 | 5.72 | 0 | Organelle membrane |
| Zlat_10005292 | ZlPHYD | HisKA, HATPase_c, GAF, PAS | Phytochrome | PHYB | 1710 | 75 | 1178 | 123.76 | 5.69 | 0 | Nucleus |
| Zlat_10007618 | ZlPHYE | HisKA, HATPase_c, GAF, PAS | Phytochrome | PHYC | 1384 | 59 | 1137 | 125.65 | 5.62 | 0 | Chloroplast |
| Zlat_10046362 | ZlPDK1 | HATPase_c | Pyruvate dehydrogenase kinase | PDK | 530 | 74 | 363 | 40.77 | 6.08 | 0 | Chloroplast |
| Zlat_10041425 | ZlPDK2 | HATPase_c | Pyruvate dehydrogenase kinase | PDK | 521 | 72 | 363 | 40.73 | 6.33 | 0 | Chloroplast |
| Zlat_10030552 | ZlPDK3 | HATPase_c | Pyruvate dehydrogenase kinase | PDK | 551 | 76 | 363 | 40.91 | 6.68 | 0 | Nucleus |
| Zlat_10018898 | ZlPDK4 | HATPase_c | Pyruvate dehydrogenase kinase | PDK | 555 | 73 | 405 | 44.76 | 6.84 | 0 | Chloroplast |
| Gene ID | Gene Name | Character a | Gene Family | Counterpart | Score(bits) | Identity (%) | Length (aa) | Molecular Weight(kDa) | Isoelectric Point (PI) | Subcellular localization b |
|---|---|---|---|---|---|---|---|---|---|---|
| Zlat_10020936 | ZlHP1 | HPt | HPt | AHP1 | 117 | 20 | 210 | 23.93 | 8.57 | Organelle membrane |
| Zlat_10031305 | ZlHP2 | HPt | HPt | AHP1 | 115 | 50 | 146 | 16.63 | 5.28 | Nucleus |
| Zlat_10002667 | ZlHP3 | pseudo-HPt | pseudo-HPt | AHP4 | 162 | 61 | 276 | 30.89 | 7.11 | Extracellular space |
| Zlat_10013094 | ZlHP4 | HPt | HPt | AHP4 | 184 | 59 | 151 | 17.84 | 8.33 | Nucleus |
| Zlat_10015279 | ZlHP5 | pseudo-HPt | pseudo-HPt | AHP4 | 187 | 59 | 158 | 18.03 | 8.2 | Nucleus |
| Zlat_10032102 | ZlHP6 | pseudo-HPt | pseudo-HPt | AHP4 | 197 | 63 | 151 | 17.42 | 7.55 | Nucleus |
| Zlat_10034774 | ZlHP7 | HPt | HPt | AHP5 | 125 | 46 | 149 | 16.78 | 4.71 | Nucleus |
| Zlat_10034845 | ZlHP8 | HPt | HPt | AHP5 | 125 | 46 | 149 | 16.79 | 4.66 | Nucleus |
| Gene ID | Gene Name | Character a | Gene Family | Counterpart | Score(bits) | Identity (%) | Length (aa) | Molecular Weight(kDa) | Isoelectric Point (PI) | Subcellular Localization b |
|---|---|---|---|---|---|---|---|---|---|---|
| Zlat_10022933 | ZlRRA1 | REC | Type A | ARR3 | 103 | 61 | 167 | 16.23 | 5.71 | Nucleus |
| Zlat_10026415 | ZlRRA2 | REC | Type A | ARR3 | 110 | 65 | 175 | 16.16 | 6.84 | Chloroplast |
| Zlat_10028329 | ZlRRA3 | REC | Type A | ARR3 | 129 | 64 | 143 | 15.11 | 9.46 | Nucleus |
| Zlat_10028974 | ZlRRA4 | REC | Type A | ARR3 | 129 | 64 | 137 | 14.75 | 6.59 | Nucleus |
| Zlat_10030869 | ZlRRA5 | REC | Type A | ARR3 | 124 | 54 | 258 | 28.13 | 5.99 | Extracellular space |
| Zlat_10032021 | ZlRRA6 | REC | Type A | ARR3 | 121 | 52 | 269 | 29.31 | 7.6 | Plasma membrane |
| Zlat_10033406 | ZlRRA7 | REC | Type A | ARR6 | 152 | 64 | 165 | 17.89 | 9.1 | Chloroplast outer membrane |
| Zlat_10001454 | ZlRRA8 | REC | Type A | ARR8 | 200 | 72 | 234 | 25.56 | 4.83 | Nucleus |
| Zlat_10017478 | ZlRRA9 | REC | Type A | ARR8 | 99.4 | 59 | 187 | 21.08 | 6.81 | Nucleus |
| Zlat_10003241 | ZlRRA10 | REC | Type A | ARR9 | 112 | 72 | 76 | 8.31 | 8.71 | Extracellular space |
| Zlat_10016677 | ZlRRA11 | REC | Type A | ARR9 | 204 | 57 | 215 | 24.01 | 6.06 | Nucleus |
| Zlat_10020468 | ZlRRA12 | REC | Type A | ARR9 | 137 | 81 | 176 | 19.42 | 6.74 | Nucleus |
| Zlat_10029578 | ZlRRA13 | REC | Type A | ARR9 | 134 | 57 | 231 | 25.92 | 4.07 | Extracellular space |
| Zlat_10044807 | ZlRRA14 | REC | Type A | ARR9 | 220 | 70 | 196 | 22.08 | 6.05 | Nucleus |
| Zlat_10024295 | ZlRRB1 | REC, Myb | Type B | ARR1 | 352 | 57 | 689 | 73.95 | 5.99 | Nucleus |
| Zlat_10041210 | ZlRRB2 | REC, Myb | Type B | ARR1 | 356 | 57 | 688 | 73.83 | 6.13 | Nucleus |
| Zlat_10019278 | ZlRRB3 | REC, Myb | Type B | ARR2 | 186 | 39 | 353 | 40.14 | 7.07 | Nucleus |
| Zlat_10010976 | ZlRRB4 | REC, Myb | Type B | ARR10 | 278 | 42 | 627 | 68.56 | 5.95 | Nucleus |
| Zlat_10006425 | ZlRRB5 | REC, Myb | Type B | ARR11 | 128 | 62 | 179 | 20.47 | 4.81 | Nucleus |
| Zlat_10007685 | ZlRRB6 | REC, Myb | Type B | ARR11 | 47.4 | 30 | 340 | 36.56 | 5.61 | Cytoplasm |
| Zlat_10010099 | ZlRRB7 | REC, Myb | Type B | ARR11 | 328 | 50 | 583 | 65.13 | 5.04 | Nucleus |
| Zlat_10033001 | ZlRRB8 | REC, Myb | Type B | ARR11 | 348 | 54 | 580 | 65.07 | 5.16 | Nucleus |
| Zlat_10007027 | ZlRRB9 | REC, Myb | Type B | ARR12 | 321 | 49 | 707 | 76.01 | 6.25 | Nucleus |
| Zlat_10018529 | ZlRRB10 | REC, Myb | Type B | ARR12 | 343 | 55 | 694 | 76.02 | 5.87 | Nucleus |
| Zlat_10018563 | ZlRRB11 | REC, Myb | Type B | ARR12 | 322 | 66 | 693 | 75.34 | 6.2 | Nucleus |
| Zlat_10022449 | ZlRRB12 | REC, Myb | Type B | ARR12 | 336 | 59 | 621 | 68.29 | 5.88 | Nucleus |
| Zlat_10028769 | ZlRRB13 | REC, Myb | Type B | ARR12 | 348 | 60 | 626 | 68.84 | 5.8 | Nucleus |
| Zlat_10041971 | ZlRRB14 | REC, Myb | Type B | ARR14 | 106 | 46 | 299 | 31.15 | 6.61 | Nucleus |
| Zlat_10027036 | ZlRRC1 | REC | Type C | ARR24 | 66.6 | 40 | 128 | 13.85 | 5.69 | Nucleus |
| Zlat_10029496 | ZlRRC2 | REC | Type C | ARR24 | 67.8 | 41 | 129 | 13.8 | 5.68 | Nucleus |
| Zlat_10006383 | ZlPRR1 | Pseudo-REC, CCT | Pseudo | APRR1 | 306 | 39 | 518 | 57.68 | 6.34 | Nucleus |
| Zlat_10008561 | ZlPRR2 | Pseudo-REC, CCT | Pseudo | APRR7 | 318 | 41 | 764 | 82.49 | 8.52 | Chloroplast outer membrane |
| Zlat_10024918 | ZlPRR3 | Pseudo-REC, CCT | Pseudo | APRR7 | 347 | 40 | 742 | 81.17 | 6.23 | Nucleus |
| Zlat_10017908 | ZlPRR4 | Pseudo-REC, CCT | Pseudo | APRR5 | 228 | 39 | 629 | 70.06 | 6.78 | Nucleus |
| Zlat_10023948 | ZlPRR5 | Pseudo-REC, CCT | Pseudo | APRR5 | 212 | 53 | 684 | 74.99 | 8.21 | Nucleus |
| Zlat_10035513 | ZlPRR6 | Pseudo-REC, CCT | Pseudo | APRR5 | 215 | 54 | 682 | 75.18 | 8.44 | Nucleus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Zhang, F.; Wu, X.; Hu, Y.; Dong, L.; Dewitte, W.; Wen, B. Genome-Wide Characterization and Expression of Two-Component System Genes in Cytokinin-Regulated Gall Formation in Zizania latifolia. Plants 2020, 9, 1409. https://doi.org/10.3390/plants9111409
He L, Zhang F, Wu X, Hu Y, Dong L, Dewitte W, Wen B. Genome-Wide Characterization and Expression of Two-Component System Genes in Cytokinin-Regulated Gall Formation in Zizania latifolia. Plants. 2020; 9(11):1409. https://doi.org/10.3390/plants9111409
Chicago/Turabian StyleHe, Lili, Feng Zhang, Xiaozhen Wu, Yanmei Hu, LiLi Dong, Walter Dewitte, and Bo Wen. 2020. "Genome-Wide Characterization and Expression of Two-Component System Genes in Cytokinin-Regulated Gall Formation in Zizania latifolia" Plants 9, no. 11: 1409. https://doi.org/10.3390/plants9111409