Does Storage under Gene Bank Conditions Affect Seed Germination and Seedling Growth? The Case of Senecio morisii (Asteraceae), a Vascular Plant Exclusive to Sardinian Water Meadows



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
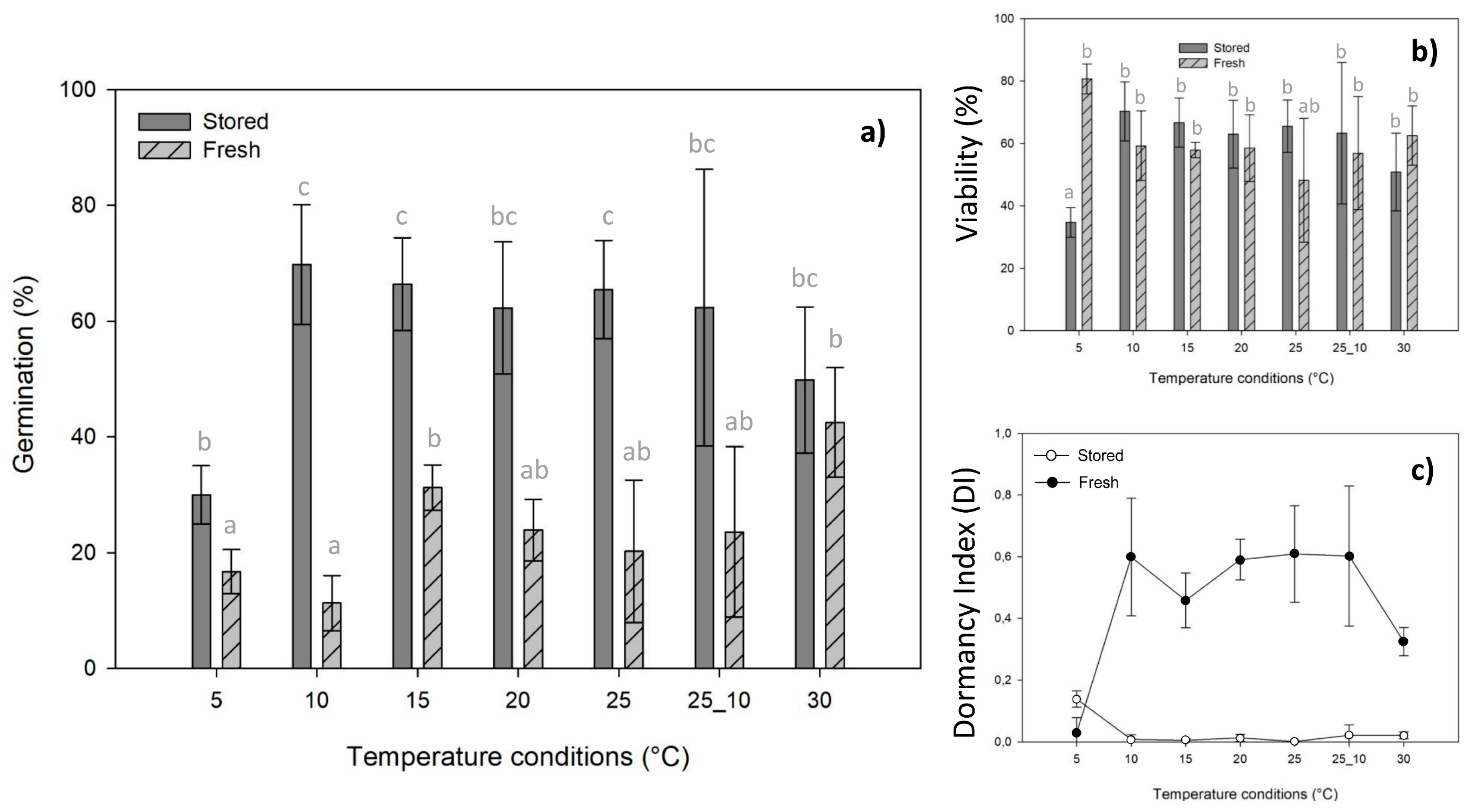
2.1. Seed Germination
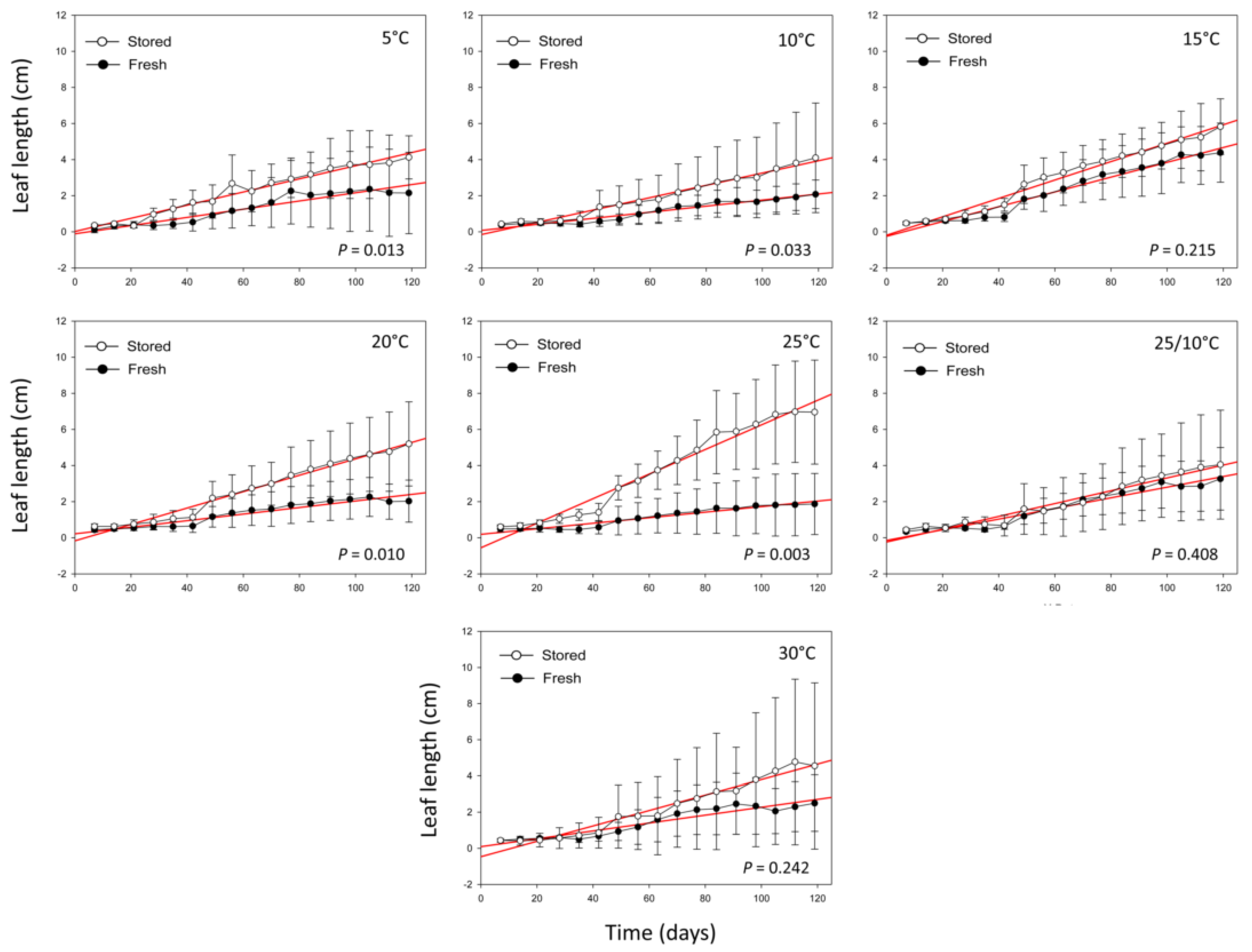
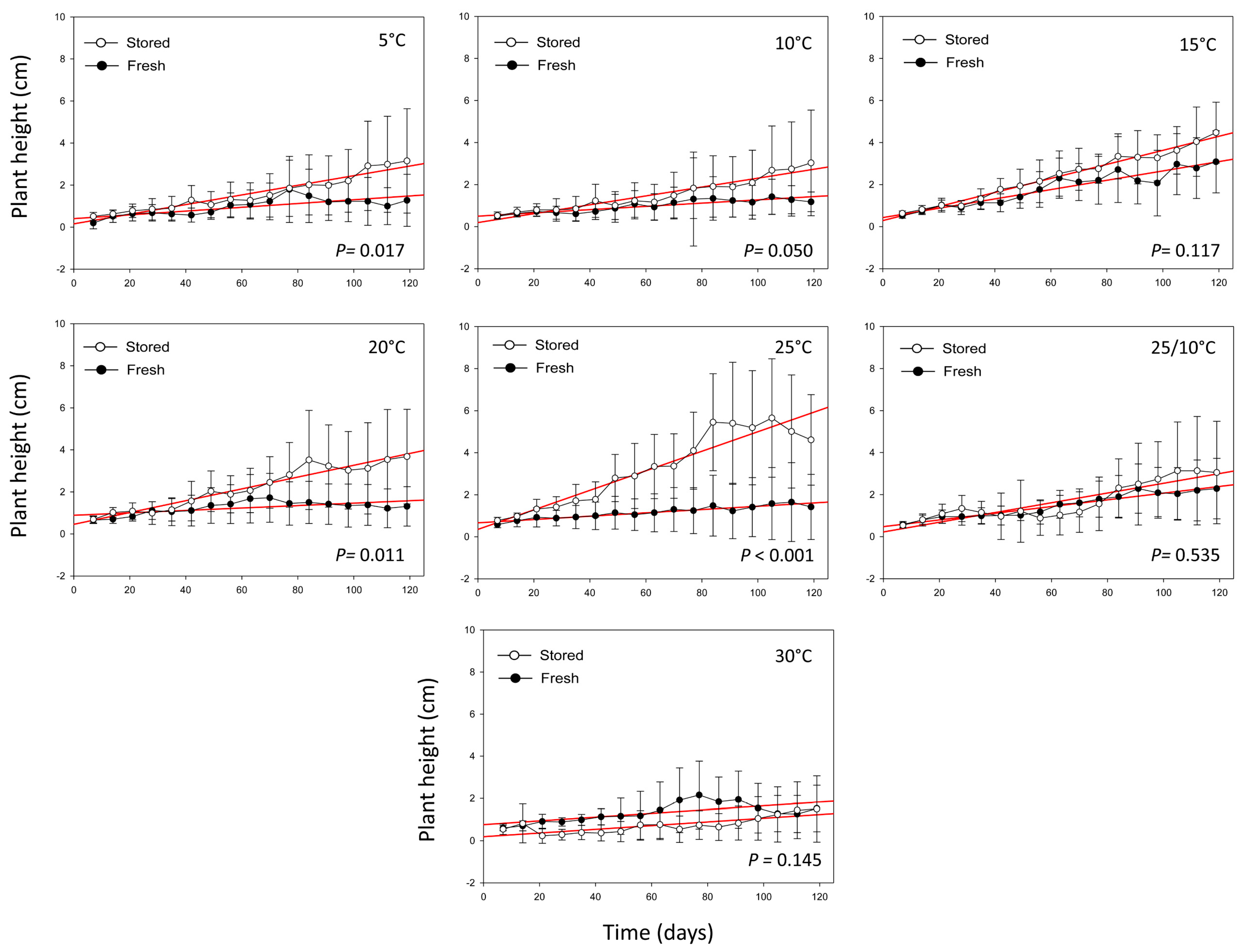
2.2. Seedling Growth and Survival Percentage
3. Discussion
4. Materials and Methods
4.1. Seed Lots Details
4.2. Seed Germination Trials
4.3. Seedling Growth and Survival Percentage
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Falk, D.A. Endangered forest resources in the US: Integrated strategies for conservation of rare species and genetic diversity. Forest Ecol. Manag. 1990, 35, 91–107. [Google Scholar] [CrossRef]
- Offord, C.A.; McKensy, M.L.; Cuneo, P.V. Critical review of threatened species collections in the New South Wales Seedbank: Implications for ex situ conservation of biodiversity. Pac. Conserv. Biol. 2004, 10, 221–236. [Google Scholar] [CrossRef]
- Mondoni, A.; Rossi, G.; Orsenigo, S.; Probert, R.J. Climate warming could shift the timing of seed germination in alpine plants. Ann. Bot. 2012, 110, 155–164. [Google Scholar] [CrossRef]
- Maunder, M.; Guerrant, E.O.; Havens, K.; Dixon, K.W. Realizing the full potential of ex situ contributions to global plant conservation. In Ex Situ Plant Conservation: Supporting Species Survival in the Wild; Guerrant, E.O., Havens, K., Maunder, M., Eds.; Island Press: Washington, DC, USA, 2004; pp. 389–418. [Google Scholar]
- Cochrane, J.A.; Crawford, A.D.; Monks, L.T. The significance of ex situ seed conservation to reintroduction of threatened plants. Austral. J. Bot. 2007, 55, 356–361. [Google Scholar] [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011.
- Fenu, G.; Fois, M.; Cogoni, D.; Porceddu, M.; Pinna, M.S.; Cuena-Lombraña, A.; Nebot, A.; Sulis, E.; Picciau, R.; Santo, A.; et al. The Aichi Biodiversity Target 12 at regional level: An achievable goal? Biodiversity 2015, 16, 120–135. [Google Scholar] [CrossRef]
- Rivière, S.; Müller, J. Contribution of seed banks across Europe towards the 2020 Global Strategy for Plant Conservation targets, assessed through the ENSCONET database. Oryx 2018, 52, 464–470. [Google Scholar] [CrossRef]
- Roberts, E.H. Predicting the storage life of seeds. Seed Sci. Technol. 1973, 1, 499–514. [Google Scholar]
- ISTA. International Rules for Seed Testing, 2008 ed.; The International Seed Testing Association (ISTA): Bassersdorf, Germany, 2008. [Google Scholar]
- Probert, R.; Adams, J.; Coneybeer, J.; Crawford, A.; Hay, F. Seed quality for conservation is critically affected by pre-storage factors. Austral. J. Bot. 2007, 55, 326–335. [Google Scholar] [CrossRef]
- Probert, R.J.; Daws, M.I.; Hay, F.R. Ecological correlates of ex situ seed longevity: A comparative study on 195 species. Ann. Bot. 2009, 104, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Godefroid, S.; Van de Vyver, A.; Lebrun, J.; Masengo Kalenga, W.; Handjila Minengo, G.; Rose, C.; Ngongo Luhembwe, M.; Vanderborght, T.; Mahy, G. Germination capacity and seed storage behaviour of threatened metallophytes from the Katanga copper belt (DR Congo): Implications for ex situ conservation. Plant. Ecol. Evol. 2013, 146, 183–192. [Google Scholar] [CrossRef]
- Bacchetta, G.; Fenu, G.; Mattana, E.; Piotto, B.; Virevaire, M. Manuale per la Raccolta, Studio, Conservazione e Gestione ex situ del Germoplasma; Manuali e Linee Guida (APAT): Roma, Italy, 2006. [Google Scholar]
- Bacchetta, G.; Bueno-Sánchez, A.; Fenu, G.; Jiménez-Alfaro, B.; Mattana, E.; Piotto, B.; Virevaire, M. Conservación ex situ de Plantas Silvestres; La Caixa: Principado de Asturias, Spain, 2008. [Google Scholar]
- Porceddu, M.; Santo, A.; Orrù, M.; Meloni, F.; Ucchesu, M.; Picciau, R.; Sarigu, M.; Cuena-Lombraña, A.; Podda, L.; Sau, S.; et al. Seed conservation actions for the preservation of plant diversity: The case of the Sardinian Germplasm Bank (BG-SAR). Plant Sociol. 2017, 54, 111–117. [Google Scholar] [CrossRef]
- FAO. Genebank Standards for Plant Genetic Resources for Food and Agriculture; FAO: Rome, Italy, 2014. [Google Scholar]
- Godefroid, S.; Van de Vyver, A.; Vanderborght, T. Germination capacity and viability of threatened species collections in seed banks. Biodivers. Conserv. 2010, 19, 1365–1383. [Google Scholar] [CrossRef]
- Liu, K.; Baskin, J.M.; Baskin, C.C.; Bu, H.; Liu, M.; Liu, W.; Du, G. Effect of storage conditions on germination of seeds of 489 species from high elevation grasslands of the Eastern Tibet plateau and some implications for climate change. Am. J. Bot. 2011, 98, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Baldos, O.C.; DeFrank, J.; Kramer, M.; Sakamoto, G.S. Storage humidity and temperature affect dormancy loss and viability of tanglehead (Heteropogon contortus) seeds. Hortscience 2014, 49, 1328–1334. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.B.; Ahmed, Z.; Diederichsen, A. Towards a better monitoring of seed ageing under ex situ seed conservation. Conserv. Physiol. 2015, 3, cov026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempewolf, H.; Eastwood, R.J.; Guarino, L.; Khoury, C.K.; Müller, J.V.; Toll, J. Adapting agriculture to climate change: A global initiative to collect, conserve, and use crop wild relatives. Agroecol. Sust. Food 2014, 38, 369–377. [Google Scholar] [CrossRef] [Green Version]
- de Melo, R.B.; Franco, A.C.; Silva, C.O.; Piedade, M.T.F.; Ferreira, C.S. Seed germination and seedling development in response to submergence in tree species of the Central Amazonian floodplains. AoB Plants 2015, 7, plv041. [Google Scholar] [CrossRef] [Green Version]
- Merritt, D.J.; Dixon, K.W. Restoration seed banks: A matter of scale. Science 2011, 332, 424–425. [Google Scholar] [CrossRef]
- Clemente, A.S.; Müller, J.V.; Almeida, E.; Costa, C.A.; Lobo-Dias, S.; Brehm, J.M.; Rebelo, R.; Martins-Loução, M.A. What can routine germination tests in seed banks tell us about the germination ecology of endemic and protected species? Botany 2017, 95, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Pelser, P.B.; Nordenstam, B.; Kadereit, J.W.; Watson, L.E. An ITS phylogeny of tribe Senecioneae (Asteraceae) and a new delimitation of Senecio L. Taxon 2007, 56, 1077–1104. [Google Scholar] [CrossRef] [Green Version]
- Calvo, J.; Aedo, C. A taxonomic revision of the Eurasian/Northwestern African Senecio doria group (Compositae). Syst. Bot. 2015, 40, 900–913. [Google Scholar] [CrossRef]
- Fois, M.; Cuena-Lombraña, A.; Fenu, G.; Bacchetta, G. Using species distribution models at local scale to guide the search of poorly known species: Review, methodological issues and future directions. Ecol. Model. 2018, 385, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Orsenigo, S.; Montagnani, C.; Fenu, G.; Gargano, D.; Peruzzi, L.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Bovio, M.; et al. Red Listing plants under full national responsibility: Extinction risk and threats in the vascular flora endemic to Italy. Biol. Conserv. 2018, 224, 213–222. [Google Scholar] [CrossRef]
- Mira, S.; González-Benito, M.E.; Ibars, A.M.; Estrelles, E. Dormancy release and seed ageing in the endangered species Silene diclinis. Biodivers. Conserv. 2011, 20, 345–358. [Google Scholar] [CrossRef]
- Abeli, T.; Dixon, K. Translocation ecology: The role of ecological sciences in plant translocation. Plant. Ecol. 2016, 217, 123–125. [Google Scholar] [CrossRef] [Green Version]
- Sans, F.X.; García-Serrano, H.; Afán, I. Life-history traits of alien and native Senecio species in the Mediterranean region. Acta Oecol. 2004, 26, 167–178. [Google Scholar] [CrossRef]
- Hilton, J.R. The influence of light on the germination of Senecio vulgaris L. New Phytol. 1983, 94, 29–37. [Google Scholar] [CrossRef]
- Ren, Z.; Abbott, R.J. Seed dormancy in Mediterranean Senecio vulgaris L. New Phytol. 1991, 117, 673–678. [Google Scholar] [CrossRef]
- Fenner, M. The effects of the parent environment on seed germinability. Seed Sci. Res. 1991, 1, 75–84. [Google Scholar] [CrossRef]
- Fernández-Pascual, E.; Jiménez-Alfaro, B. Phenotypic plasticity in seed germination relates differentially to overwintering and flowering temperatures. Seed Sci. Res. 2014, 24, 273–280. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Mira, S.; Estrelles, E.; González-Benito, M.E. Effect of water content and temperature on seed longevity of seven Brassicaceae species after 5 years of storage. Plant. Biol. 2015, 17, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Crawford, A.D.; Steadman, K.J.; Plummer, J.A.; Cochrane, A.; Probert, R.J. Analysis of seed-bank data confirms suitability of international seed-storage standards for the Australian flora. Austral. J. Bot. 2007, 55, 18–29. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; 3.2.2 R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuena-Lombraña, A.; Sanna, M.; Porceddu, M.; Bacchetta, G. Does Storage under Gene Bank Conditions Affect Seed Germination and Seedling Growth? The Case of Senecio morisii (Asteraceae), a Vascular Plant Exclusive to Sardinian Water Meadows. Plants 2020, 9, 581. https://doi.org/10.3390/plants9050581
Cuena-Lombraña A, Sanna M, Porceddu M, Bacchetta G. Does Storage under Gene Bank Conditions Affect Seed Germination and Seedling Growth? The Case of Senecio morisii (Asteraceae), a Vascular Plant Exclusive to Sardinian Water Meadows. Plants. 2020; 9(5):581. https://doi.org/10.3390/plants9050581
Chicago/Turabian StyleCuena-Lombraña, Alba, Martina Sanna, Marco Porceddu, and Gianluigi Bacchetta. 2020. "Does Storage under Gene Bank Conditions Affect Seed Germination and Seedling Growth? The Case of Senecio morisii (Asteraceae), a Vascular Plant Exclusive to Sardinian Water Meadows" Plants 9, no. 5: 581. https://doi.org/10.3390/plants9050581