Transcription Factors in Plant Stress Responses: Challenges and Potential for Sugarcane Improvement
by
 , , ,
, , ,
Talha Javed
1,2,† ,
,
Rubab Shabbir
1,2,† ,
,
Ahmad Ali
1,
Irfan Afzal
2 ,
,
Uroosa Zaheer
1 and
San-Ji Gao
1,* 1
National Engineering Research Center for Sugarcane, Fujian Agriculture and Forestry University, Fuzhou 350002, China
2
Seed Physiology Lab., Department of Agronomy, University of Agriculture, Faisalabad-38040, Pakistan
*
Author to whom correspondence should be addressed.
†
Equally contributed authors.
Plants 2020, 9(4), 491; https://doi.org/10.3390/plants9040491
Submission received: 3 March 2020
/
Revised: 8 April 2020
/
Accepted: 8 April 2020
/
Published: 10 April 2020
(This article belongs to the Section Plant Response to Abiotic Stress and Climate Change)
Abstract
:Increasing vulnerability of crops to a wide range of abiotic and biotic stresses can have a marked influence on the growth and yield of major crops, especially sugarcane (Saccharum spp.). In response to various stresses, plants have evolved a variety of complex defense systems of signal perception and transduction networks. Transcription factors (TFs) that are activated by different pathways of signal transduction and can directly or indirectly combine with cis-acting elements to modulate the transcription efficiency of target genes, which play key regulators for crop genetic improvement. Over the past decade, significant progresses have been made in deciphering the role of plant TFs as key regulators of environmental responses in particular important cereal crops; however, a limited amount of studies have focused on sugarcane. This review summarizes the potential functions of major TF families, such as WRKY, NAC, MYB and AP2/ERF, in regulating gene expression in the response of plants to abiotic and biotic stresses, which provides important clues for the engineering of stress-tolerant cultivars in sugarcane.
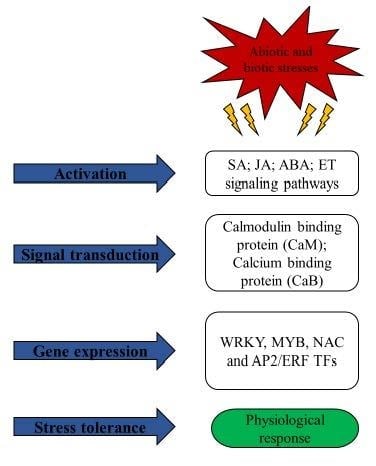
1. Introduction
During the life cycle of plant species, abiotic (temperature, waterlogging, drought, oxidative, salinity, ultraviolet) and biotic (viruses, bacteria, fungi, insects etc.) stresses are major threats to global crop productivity [1]. To cope with various stresses, plants have evolved complex rapid responses, but crop efficiency is still severely hampered. The prevailing alarming climate change scenario and future changing weather provide challenges for the researchers to better understanding plant responses such as the activation of different signal cascades, followed by signal transduction, and the respective gene expression in response to single or combined biotic or abiotic stresses, ultimately enhancing crop yield [2]. Sugarcane (Saccharum spp.) plays a crucial role in sugar and biofuel production, accounting for 80% of sugar production in the world. Genetic improvement of modern sugarcane cultivars hits a bottleneck through conventional hybrid breeding as a result of sugarcane complex genome, heterogenous and polyploid-aneuploid nature [3].
Under unfavorable environmental conditions, various stress-response mechanisms have been evolved by plants at different layers, such as cellular signal perception and transduction, inducing expression of specific sub-sets of defense genes, and therefore activating the overall defense reaction, ultimately contributing to phenotype [4]. Transcription factors (TFs) are important regulators for the control of gene expression in all living organisms and play crucial roles in plant development, cell cycling, cell signaling and stress response [5]. TFs modulate gene expression through binding with their distal and local cis-elements of target gene, which might be influenced by genomic features, DNA structure and TF interactions [6]. In plants, TFs are encoded by approximately 10% of genes at different stages/points to regulate specific signaling mediated function [5]. Various TFs databases specific for various crops are available, such as the plant transcription factor database and grass TF database. Major TF families, such as WRKY, MYB, NAC and AP2/ERF, are crucial regulators of various genes related to different stresses, which contribute to the ideal choice for genetic engineering in order to enhance resistance of plants against different stress stimuli [7]. According to the plant TF database, numerous TFs have been reported in Oryza sativa (2389), Hordeum vulgare (2620), Triticum aestivum (3437) and Saccharum spp. (672). It is noteworthy that, to date, 39 WRKY, 44 NAC, 38 MYB and 73 AP2/ERF gene families of TFs in sugarcane have been identified.
In last decade, numerous TFs involved in response to abiotic and biotic stress in different crops including sugarcane were investigated using modern molecular tools, such as functional genomics, transcriptomics and proteomics [8]. This review provides recently updated regulatory functions of major TFs families in response to stresses to understand the stress-responsive mechanisms in various crops, including sugarcane.
2. WRKY TFs in Response to Stresses
2.1. Classification and Diversification of WRKY Genes Family
WRKY being as one of the most characterized class of plant TFs, regulate diverse plants developmental, physiological and metabolic processes [9]. The first WRKY TF was identified in Ipomoea batatas in 1990s [10], but WRKY genes were considered plant specific TFs at prehistoric times [2]. A myriad of studies documented that other eukaryotic organisms, such fungi, amoebae and diplomonads, also have WRKY proteins in their genetic make-up. Ancient gene transfer events were considered as the origin of these non-plant WRKYs based on distribution pattern analysis [11]. Numerous WRKY TFs have been experimentally revealed in various plant species, including (Arabidopsis thaliana) [12], sugarcane (Saccharum spontaneum) [13], barley (Hordeum vulgare) [14], rice (Oryza sativa) [15], physic nut (Jatropha curcas) [16], maize (Zea mays) [17], wheat (Triticum aestivum) [18], cotton (Gossypium hirsutum) [19] and so on (Table 1).
Based on the genomic characterization of Arabidopsis, the most authenticated and widely accepted classification system stipulates that plant specific WRKY TFs are denoted by 60 amino acids in their highly conserved DNA binding region (called as WRKY domain) [20]. Furthermore, WRKY domains compromised of a highly conserved motif WRKYGQK at N terminus that provides a protein-protein interaction interface and a zinc finger region either C-X4-5-C-X22-23-H-X-H or C-X7-C-X23-H-X-C at the C-terminus having affinity towards DNA binding [21]. WRKY proteins, based on WRKY domains present and zinc finger region, have been classified into three major groups (Group I, II and III) [22]. Group I have two WRKY domains with a zinc finger motif (C-X4-5-C-X22-23-H-X-H) adjacent to atypical. Group II and III WRKYs have one WRKY domain. WRKYs in group I and II have the same type of finger motif, whereas group III WRKYs consists of a C-X7-C-X23-H-X-C motif at the C-terminus. WRKYs in Group II can be further categorized into five sub-groups (IIa, IIb, IIc, IId and IIe), while WRKYs in Group III can be categorized into two groups, IIIa (C-X7-C-X23-H-X-C) and IIIb (C-X7-C-Xn-H-X-C, n ≥ 24), based on the zinc-finger motif structure [21]. However, an additional WRKYs Group IV was proposed in S. spontaneum, as evidenced by these genes containing an incomplete domain (only the WRKYGQK motif was identified), which indicated they may have lost their function as WRKYs [13]. Taking together, the biological functions of all WRKYs groups have been linked with their ability to bind specifically in the promoter region of target genes to the W-box, i.e., (C/T) TGAC (C/T) and modulate these gene expressions [23].
2.2. Function and Expression Pattern of WRKYs under Abiotic Stresses
High or low temperature stress has significant influence on a myriad of processes including physiological, enzymatic, metabolic, growth and developmental in plant system, hence leading to low yield with poor quality of produce [9]. More specifically, decreased enzymatic activity due to denatured entire protein structure under temperature extremes lead to the halt of entire pathway, thereby resulting in wilting or senescence of the plant [24]. Stimulation of heat-induced signal transduction pathway by the WRKYs group I proteins (such as AtWRKY25, AtWRKY26 and AtWRKY33) contributed to significantly higher thermo-tolerance. While double (wrky25/26) and triple (wrky25/26/33) mutants depict poor seed germination, decreased membrane integrity and increased susceptibility to temperature stress [25]. The constitutive synergistic expression of TaWRKY1 and TaWRKY33 in Arabidopsis [26] and TaWRKY70 [27] in T. aestivum exhibits enhanced tolerance to heat stress. Furthermore, overexpression of TaWRKY008, TaWRKY122 and TaWRKY45 in T. aestivum also enhanced tolerance to extreme temperature stress [28]. Overexpression of VaWRKY12 that was localized in the nucleus inhibited the cellular damage in Arabidopsis and grapevine after cold stress [29], while overexpression of ZmWRKY106 in Zea mays showed higher activities of antioxidants under heat stress and improved vigor of transgenic plants [30].
Waterlogging is one of the serious threats to agriculture, as it affects the aeration ability of roots and alters the physical and chemical nature of soil, thus triggering the adverse effects on productivity of crop species worldwide [31]. Overexpression of WRKY22 in Arabidopsis, by targeting genes encoding a toll/interleukin-1 receptor (TIR) domain–containing protein, lead to cope the threat associated with excess soil moisture [32]. A sunflower TF HaWRKY76 was upregulated in Arabidopsis transgenic plants in response to excess soil moisture, imparting enhanced resistance through the repression of fermentation pathways that ensue to carbohydrates reservation [33]. In contrast, PlWRKY70, a Paeonia lactiflora TF, was considerably suppressed under waterlogging stress [34].
Drought, due to reduced potential water content in plants, can directly limit the normal functioning of crop plants by averting the enzymatic activities, accumulation of soluble matters, formation of ROS (reactive oxygen species) multiplexes and restricted metabolic pathways, thus making the agricultural efficiency at risk [35]. WRKY TFs has been frequently identified and reported to play optimistic role in regulating defense genes under water deficient environments. For example, AtWRKY28 in Arabidopsis, positively regulated in response to drought stress [36], whereas WRKY46/54/70 acted as negative regulators under drought stress [37]. In sugarcane, a WRKY-IIc TF gene ScWRKY3 played positive regulation by sodium chloride (NaCl), polyethylene glycol (PEG), and abscisic acid (ABA), but it was suppressed by salicylic acid (SA) and methyl jasmonate (MeJA) [38]. In other crops, overexpression of TaWRKY93 [39], ZmWRKY40 [17], TaWRKY2 [40], GhWRKY91 [41] and GmWRKY54 [42] conferred drought tolerance through ABA, Ca2+-mediated signal transduction pathways and osmotic adjustments.
Ultraviolet (UV) light plays a crucial role from germination to the growth and development in plants [43]. However, UV type B light can restrict the normal enzymatic functioning and also cause significant damage to DNA bases. In rice, overexpressing OsWRKY89, a group III TF, through direct or indirect activation of SA mediated signal transduction pathway, not only intensified the plants resistance against UV irradiation but also imparted tolerance counter to insect pests and fungal infection [44]. Oxidative stress mediated through jasmonate (JA) signaling pathway is among one of the most detrimental stresses [45], while production and prompt scavenging of ROS are responsible for regulation of ROS-mediated signaling in plants [46]. Overexpression of WRKY4, WRKY5, WRKY11 and WRKY46 were observed during the exposure of oxidative stress in Salvia officinalis and play a pivotal role in the signaling mechanism during plant responses [45].
2.3. Function and Expression Pattern of WRKY Genes under Biotic Stresses
During biotic stress, plants undergo the partial or complete modulation of multiple signal transduction pathways such as plant hormones, which subsequently resulted in the induction of several related transcriptional genes, thus resulting in positive response to stressful environment [4]. Plant WRKYs have been involved in the microbe-associated molecular pattern-triggered immunity (PAMP-triggered immunity), effector-triggered immunity (ETI), or system acquired resistance (SAR) [9]. For example, CsWRKY50 plays crucial role against infection stress of Pseudopernospora cubensis in Cucumis sativus [47]. VvWRKY1, JA pathway-related gene, positively regulates the stress tolerance response to downy mildew in Vitis vinifera [48]. In addition, another JA-regulated gene, CaWRKY27, confers the resistance against Ralstonia solanacearum infection in tobacco (Nicotiana tabacum) [49]. GhWRKY44, a significant TF in cotton-pathogen interaction, it provides an enhanced tolerance in tobacco against bacterial and fungal pathogens. The plants exhibited a lower level of ROS accumulation in experimental units having overexpression of GhWRKY44 through SA and JA signal transduction pathways of disease resistance [50]. A significant increase in resistance by TaWRKY70 transcription factor, through SA and ET mediated signal transduction pathways, was noted in plants when exposed to pathogen (Puccinia striiformis sp. tritici) infection [27]. A versatile role in response to pathogen stimuli was indicated by WsWRKY1 [51], AcWRKYs [52] and GmWRKY31 [53] TFs in different crops. The sugarcane ScWRKY3 was stably expressed in the smut-resistant cultivar, whereas it was suppressed in the smut-susceptible cultivar at early stages (0–72 h) of infection with the smut pathogen (Sporisorium scitamineum). Meanwhile, this gene was proposed to act as a negative regulator under the pathogen infection either Fusarium solani var. coeruleum or Ralstonia solanacearum in N. benthamiana [38]. More recently, most of WRKY33 alleles were significantly upregulated in sugarcane against the Xanthomonas albilineans attacks [54]. Collectively, WRKY TFs through self-regulation or hormones mediated signal transduction pathways proved to be effective for alleviating the infection stress caused by biotic or abiotic agents.
3. NAC TFs in Response to Stresses
3.1. Classification and Diversification of NAC Gene Family
NAC (NAM: no apical meristem, ATAF, CUC: cup-shaped cotyledon) is one of the most important and largest family of plant-specific stress-responsive TFs [55]. NAM from Petunia hybrida [56] and Ataf1/2 from Arabidopsis [57] were the first reported NAC proteins. Numerous NAC genes have been identified in sugarcane and other important crops (Table 1). NAC TFs are characterized by diverse C terminal that contains a variable transcriptional regulatory region (TR), while N-terminal equipped with 150–160 amino acids and also harbor NAC domain for DNA binding [58,59]. The highly conserved NAC domain can be further classified into five different sub-domains (A–E). Furthermore, nuclear localization, formation of homo/heterodimers and DNA binding have been associated with the functions of conserved NAC domain, while TR region either as activator or repressor was proven to be associated with transcription regulation [59]. Based on structure, NAC TFs can be grouped into two sub classes, typical and atypical NAC TFs. Typical NAC TFs are characterized by the presence of NAC domain at N terminal and a divergent C-region [59], while in atypical NAC TFs, the C-terminal regions contain additional motifs/domains or the C-terminus can be absent [60]. NTLs (NAC with transmembrane motif1-like), atypical NAC TFs, are characterized by the occurrence of transmembrane (TM) motif in C-region [61]. Notably, TM motif is thought to be involved in anchoring to plasma membrane, where they could impart their function by releasing through proteolysis [62,63].
3.2. Function and Expression Pattern of NAC TFs under Abiotic Stresses
Crop plants employ various molecular mechanisms such as chaperon signaling, ROS scavenging, various compatible solutes accumulation, transcriptional activity, antioxidant production and induction of mitogen-activated protein kinase (MAPK) and calcium-dependent protein kinase (CDPK) cascades against heat stress [64,65]. NAC TFs were observed to be involved in molecular regulations during temperature fluctuations. For example, overexpression of NTL1 and NTL11 genes were observed after heat stress, while NTL4 and NTL7 were exhibited specifically by low temperature in Arabidopsis [62]. In sugarcane, SsNAC23 expression was induce in response to low temperature stress [66]. ANAC019, member of NAC TFs, has been observed to enhance the ability of Arabidopsis to cold tolerance by inducing the expression of cold responsive marker gene COR47 [55]. Transformation of Arabidopsis with GmNAC20 of Glycine max exhibits an increased tolerance against chilling and salinity stress by the activation of dehydration responsive element binding/C-repeat binding factor-cold responsive (DREB/CBF-COR) pathway [67]. Various members of NAC play a significant role against temperature stress in different crop species as evidenced by overexpression of OsNAP exhibit improved resistance in rice against cold stress at the vegetative stage through ABA-mediated signal transduction pathway [68]. Notably, interaction of LlNAC2 with LlZHFD4 and LlDREB1 from tiger lily enhanced the tolerance in Arabidopsis against cold, salinity and drought stresses by maintaining its membrane integrity [69]. CaNAC064 was found a positive modulator of plant resistance to cold stress by interacting with haplo-proteins that were induced by low temperature [70].
Aquaporins expression, stability of cell membrane, accumulation of osmo-protectants and scavenging of ROS are modulated in plants when exposed to drought stress [71]. NAC TFs have been reported to regulate drought stress in Arabidopsis as overexpressing ANAC019, ANAC055 and ANAC072 genes bind specifically to drought responsive cis-element [72]. Overexpression of OsNAC10 (root-specific) exhibited to improve root dynamics and to enhance drought tolerance, which ultimately leading towards sustainable plants physio-morphological and yield related attributes in rice under field conditions [73]. Another root specific OsNAC6 TF was proven to be effective for enhanced drought tolerance and root development in rice [74]. In transgenic rice, overexpression of EcNAC67 upon drought stress displayed higher seedling vigor, lower spikelet sterility and enhanced tolerance to drought stress [75]. Overexpression of TaNAC47 and ONAC066 enhanced drought tolerance in wheat [76] and rice [77], respectively. Moreover, GhNAC2 [78] showed the potential to enhance root growth and tolerance against limited water supply in transgenic cotton and Arabidopsis. In Pyrus betulifolia, overexpression of PbeNAC1 imparted resistance against chilling and drought stress by interacting with PbeDREBs [79]. Few overexpressing NAC genes such as NAC016 in Arabidopsis [80] and ShNAC1 in tomato [81] were also found to be negative regulators against drought stress.
Balazadeh et al. [82] demonstrated the disruptive role of ANAC092 for higher rate of seed germination, salinity inducing lower chlorophyll loss at later stages, and the delay of leaf and flower senescence in Arabidopsis. Significantly enhanced tolerance to osmotic stress in rice was attributed by OsSRO1c gene. This gene imparts stomatal closure and hydrogen peroxide (H2O2) accumulation by interacting with several stress responsive functional and regulatory proteins [83]. In soybean, lateral root development, alleviation of salt and freezing stress was resulted from the overexpression of GmNAC20. Stress alleviation by GmNAC20 was regulated by the activation of the DREB/CBF–COR pathway. Overexpressing GmNAC11 only led to decreased sensitivity towards salinity [67]. A nucleus localized TF TaNAC67 conferred significantly enhanced multi-abiotic stresses (drought, salt, freezing) tolerance in transgenic Arabidopsis [84]. Overexpression of TaNAC29 in transgenic Arabidopsis plants, grown under greenhouse conditions, showed decreased sensitivity towards drought and salinity stresses with increased activity of antioxidants and lower accumulation of harmful malondialdehyde (MDA) and H2O2 contents [85]. Enhanced tolerance to salinity and osmotic stresses was markedly confirmed by overexpression of ThNAC7 in Tamarix hispida seedlings [86].
3.3. Function and Expression Pattern of NAC TFs under Biotic Stresses
Many studies showed that a number of NAC TFs play dual roles in plant immunity against various pathogens through the hypersensitive responses and ETI [87]. A wheat TF TaNAC8 has a positive role to protect plants against the stripe rust pathogen infection [88]. ZmNAC41 and ZmNAC100 genes were induced by JA and SA pathways, respectively, to ensure tolerance in maize against Colletotrichum graminicola [89]. In contrast, TaNAC30 [90] and TaNAC2 [91] negatively regulated the defense mechanism against pathogen stress. Virus induced gene silencing (VIGS) analysis in tomato depicted the positive role of SlNAC61 against infection stress caused by the Tomato yellow leaf curl virus (TYLCV) [92]. In rice, ONAC122 and ONAC131 TFs have imperative roles in diseases tolerance responses through the regulated expression of defense- and signaling-related genes such as OsLOX, OsPR1a, OsWRKY45 and OsNH1 [93]. In tomato, two homologous NAC TFs (JA2 and JA2L) differentially regulated stomatal closure and reopening under pathogen attack through. Namely, JA2 played a positive role in stomatal closure by regulating the expression of an ABA biosynthetic gene, whereas JA2L served as a positive role in JA/COR (JA/coronatine)-mediated stomatal reopening by regulating the expression of genes involved in the JA metabolism [94]. Interestingly, hypersensitive cell death in response to bacterial pathogens was promoted by the overexpression of NAC4 in Arabidopsis [95]. The LsNAC069-silenced lettuce lines showed non-significant alteration in susceptibility to Bremia Lactucae, but increased resistance to Pseudomonas cichorii bacteria [96]. The characterization of TFs in sugarcane is still limited. Overall, NAC TFs through several signal mediated cascades play important role in protecting plants against different abiotic and biotic stresses.
4. MYB TFs in Response to Stresses
4.1. Classification and Diversification of MYB Gene Family
The MYB family is a major and functionally diverse protein class of eukaryotes. The proteins of this family as TFs are majorly associated with protein-protein interaction, DNA binding and regulatory activity management of proteins [97]. Several MYB proteins have also been identified in regulating various cellular processes such as the stress responses, cell morphogenesis and cell cycle of different crop species [98]. The MYB gene “colored1 (c1)” was identified in Zea mays, which encodes a MYB-protein domain involved in anthocyanin biosynthesis in aleurone layer of maize seed [99].
Based on the number of repeats (varying from 1–4) in sequence, MYB family proteins have been classified in four groups, i.e., 1R-MYB (one repeat), R2R3-MYB (two repeats), 3R-MYB (three repeats) and 4R-MYB (four repeats) [100]. Each repeat forms three α-helices containing approximately 50–53 amino-acids with second and third helices forming HTH fold (helix-turn-helix). HTH fold consists of three tryptophan (amino acids) spaced equally apart forming a hydrophobic core [101]. The R2R3-MYB subfamily has also been classified in 30–38 groups on the basis of diverse domains of N- and C-terminal [102]. The presence and role of MYB TFs have thoroughly been investigated in various plant species including sugarcane, thus making them key factors for regulating response to biotic and abiotic stresses (Table 1).
4.2. Function and Expression Pattern of MYB TFs under Abiotic Stresses
Plants regulate the stress related genes and signaling networks to overcome abiotic stresses [2]. Many of MYB TFs have been observed to play a significant role in regulating heat stress of various crops. Like in tomato, overexpression of LeAN2 improved the resistance of plants against heat stress through a high level of antioxidants activity (non-enzymatic) and a low level of ROS accumulation [103]. In rice, overexpressing OsMYB1 gene can increase the tolerance of plants to both heat and salinity stresses [104]. While in maize, OsMYB55 proved to be effective in enhancing tolerance against high temperature and drought stress with improved growth and development of seedlings [105]. In wheat, six MYB genes related to heat stress were identified out of which TaMYB80 was effective for resistance against heat and drought stresses in transgenic Arabidopsis [106]. Similarly, PbrMYB5 from Pyrus betulaefolia imparted tolerance against cold stress by modulating the ascorbic acid biosynthesis in tobacco [107].
MYB TFs have been observed to be involved in ABA signaling pathways in response to drought stress [108] in various crops particularly in Arabidopsis. Approximately 51% MYB proteins were upregulated (such as AtMYB2/74/102) and 41% were downregulated as a result of drought stress [109]. Overexpression of AtMYB44 (a member of R2R3-MYB TF subfamily) has been observed to enhance the tolerance against drought/salt-stress in soybean [110]. Similarly, BcMYB1, another member of R2R3-MYB TF subfamily, was induced strongly during water deficiency, high polyethylene glycol contents and salinity stress in Boea crassifolia. However, only marginal induction of BcMYB1 was observed in the case of low temperatures [111].
In Arabidopsis, overexpression of AtMYB37 could increase seed yield and tolerance of transgenic plants against drought stresses [112]. In sugarcane, an MYB TF gene PScMYBAS1 promoter was involved in positive regulation under abiotic stresses (dehydration, salt, cold and wounding) and hormone (SA and MeJA) treatments [113]. Recently, a study reported that the overexpression of ScMYBAS1-3 was associated with drought resistance and biomass accumulation in rice transgenic lines [114]. Another observation revealed that a sugarcane ScMYB2 was participated in an ABA-mediated leaf senescence signaling pathway and acted as a positive regulator in respond to PEG-mediated drought stress [115]. OsMYB3 from rice was observed to enhance tolerance against cold, while OsMYB2 was observed to enhance tolerance against dehydration, cold and salt stresses [116]. Expression of OsMYB48-1 can be distinctly induced through PEG, ABA, H2O2 and dehydration, whereas only slight induction was observed after cold and high salinity stresses. However, overexpression of OsMYB48-1 could enhance the drought and salinity tolerance in rice [117]. Similarly, wheat TaMYB1D was proven to be effective to enhance the tolerance against drought and oxidative stresses through regulating phenylpropanoid metabolism [118].
Among secondary metabolite, flavonoids and sinapate esters are key compounds in absorbing UV-B and enhance tolerance of plants from harmful UV-radiation effects in UV-B impaired mutant of Arabidopsis [119]. Initial targets of UV-B include lipids, proteins and nucleic acids and their damage is compromised by producing higher number of flavonoids and sinapate esters in plants under low exposure of UV-B light [119,120]. Many MYB TFs were observed to be regulated by mediating the phenylpropanoid metabolism to protect the plants against light and other stresses [120]. AtMYB4-mutated Arabidopsis plants showed enhanced tolerance to UV-B exposure because MYB4 represses the function of gene producing cinnamate 4-hydroxylase enzyme, which is involved in biosynthesis of hydroxycinnamate ester, while another protein AtMYB7 had a positive effect in producing phenylpropanoid compounds to absorb UV-B radiation [119]. Thus, it was concluded that both AtMYB4 and AtMYB7 are involved in regulating the balance for the production and accumulation of sunscreen compounds for radiation tolerance of Arabidopsis [119]. In addition, overexpression of GmMYB12B2, a soybean R2R3-MYB TF, rendered tolerance to UV radiation and salt stress in transgenic Arabidopsis [121].
4.3. Function and Expression Pattern of MYB TFs under Biotic Stresses
In addition to abiotic stress response, MYB TFs were also observed to be involved against biotic stresses. In Arabidopsis, AtMYB102 was observed to enhance susceptibility against GPA (green-peach aphids) infestation [122]. AtMYB96 acts as important molecular link between SA and ABA crosstalk, through which it can enhance the resistance against pathogens in Arabidopsis [123]. Beneficial microorganisms also trigger the defense responses of plant and in their response MYB72 (root specific) act as signaling pathways convergence node in Arabidopsis [124]. Overexpression of TaRIM1 increased the resistance against infestation of Rhizoctonia cerealis in transgenic wheat [125]. A MYB TF y1 (yellow seed 1) of sorghum produced the 3-deoxyanthocyanidin phytoalexins against the attack of Colletotrichum sublineolum in maize [126]. Overexpression of GmMYB12B2, a soybean R2R3-MYB TF, rendered tolerance to UV radiation and salt stress in transgenic Arabidopsis [121]. Moreover, induction of MYB TFs against insect infestation was also observed in chrysanthemum, such as the overexpression of CmMYB15 through lignin accumulation, which can reduce the aphid proliferation [127]. Similarly, another MYB TF, MdMYB30, could enhance disease resistance through the regulation of wax biosynthesis in apple [128]. CaPHL8, a novel MYB TF, was proven to be effective for increasing pepper plants immunity against Ralstonia solanacerum infection [129]. VdMYB1, a member of R2R3-MYB TF, was observed as a positive activator of the defense response by activating the expression of stilbene synthase gene2 (VdSTS2) against infection stress caused by Erysiphe necator fungus of grapevine [130]. Overall, MYB TFs play a crucial role in enhancing the tolerance of plants against all type of stresses via biotic stresses and abiotic stresses.
5. AP2/ERF TFs in Response to Stresses
5.1. Classification and Diversification of AP2/ERF Gene Family
The AP2/ERF (APETALA2/Ethylene response element binding factors) superfamily genes are characterized by the presence of highly conserved DNA-binding domain with 60–-70 amino acids in each domain [131]. In addition, presence of alanine and aspartate at position 9 and 14, respectively, governs the binding of the cis-element [132]. However, based on number of domains (single or double) existing in genes, AP2/ERF can be categorized into five major sub-groups: AP2, RAV (Related to ABI3/VP1), DREB (Dehydration responsive element binding protein), ERF (Ethylene responsive factors) and others [132]. Until now, numerous AP2/ERF TFs were identified in plant species, including six AP2/ERF TFs in sugarcane and 30 AP2/ERF TFs in Arabidopsis (Table 1).
5.2. Function and Expression Pattern of AP2/ERF TFs under Abiotic Stresses
AP2/ERF TFs play a significant role in several abiotic stresses tolerance most probably through ET mediated signaling cascade. The identified AP2/ERF TFs, known to be involved in abiotic tolerance, belonged to Arabidopsis [132]. Overexpression of wheat TaDREB3-A1 gene displayed the enhanced tolerance in transgenic Arabidopsis plants against heat, drought and salt stresses [133]. An observation revealed that Arabidopsis plants overexpressing RAP2.1 showed an enhanced sensitivity to cold and drought stresses, as evidenced that RAP2.1 acting as an active transcriptional repressor in defense responses to these stresses in Arabidopsis through the down-expression of the desiccation/Cold-regulated (RD/COR) genes [134]. Another study showed that overexpression of a BnaERF-B3-hy15mu3 mutant gene from a modified AP2/ERF TF from Brassica napus enhances freeze tolerance in transgenic Arabidopsis, as resulted from this mutant gene encoded for a factor exhibiting more binding activity with the GCC box element than the wild-type gene [135]. Two Groups DREBs (I and II) from Brassica napus regulated synergistically the drought-responsive element (DRE)-mediated signaling pathway by trans-active and trans-inactive at different stages of cold stress in a competitive manner [136]. Expression pattern of ERF2 and ERF3 impart the acclimation response against cold stress and transgenic Arabidopsis lines, also exhibiting low electrolyte leakage with improved root architecture [137]. Similarly, higher measurements for antioxidants were observed, while lower MDA contents were observed due to the upregulation of cold responsive genes (VaERF080 and VaERF087) in Arabidopsis [138]. A recent study also showed higher antioxidants, lower ROS, and improved cold tolerance by overexpression of BpERF13 in birch [139]. OsDREB1G overexpressed in rice exhibited strong cold tolerance, but did not exhibit significant increase in drought or salt tolerance [140].
AP2/ERF TFs have been identified and characterized for waterlogging in many plant species. Sugarcane aerial roots developed under waterlogging stress, contribute to maintaining higher root activity and a higher ethylene concentration that increased the sensitivity of adventitious root-forming tissues and plays a principal role in aerenchyma formation [141]. Transcriptomic profiling of maize illustrated that 38 out of 184 AP2/ERF genes were remarkably induced by waterlogging stress [142]. The Arabidopsis AtRAP2.12 is a key transcriptional regulator of the core anaerobic response controlled hydraulic conductivity of root 1 (HCR1), thereby regulating hydraulic conductivity (Lpr) and hypoxia responsive genes hence modulating the resilience of plants to multiple flooding scenarios [143]. Similarly, ZmERFB180 [144] and TaERFVII.1 [145] conferred waterlogging tolerance in maize and wheat, respectively. Three ThRAP2.3/15/39 genes also played a vital role to cope with the waterlogging stress in the Taxodium hybrid [146].
The expression analysis in Populus euphratica depicted that upregulated expression of PeDREBa exhibited significantly higher values for physio-morphological traits and signal responsive regulation of drought and salt stress [147]. A soybean GmDREB2 overexpressed in Arabidopsis alleviated the drought stress and salt toxicity through binding specifically to the cis-element [148]. Another soybean GmDREB1 activated the expression of numerous soybean-specific stress-responsive genes i.e., an ABA receptor family protein (GmPYL21) and translation-related genes such as ribosomal proteins under diverse abiotic stress conditions [149]. Transcriptomic analysis revealed that AP2si6 appeared to be more dynamically expressed in transgenic Sesamum indicum to cope with signal responsive water deficiency [150]. Overexpression of OsDREP1 [151] and AtDREB2A CA [152] conferred drought endurance in rice and sugarcane, respectively. Transcriptional profiling of wheat plants has revealed that Dreb2 genes were differentially upregulated in the presence of ABA responsive cis-element to cope with drought stress [153]. TaDREB1 genes were induced as a result of DRE-binding protein (a transcript activator), thereby improving the tendency of wheat plants to tolerate the osmotic variations [154]. Transgenic wheat and barley carrying two DREB/CBF genes TaDREB3 and TaCBF5L under the stress-responsive promoters HDZI-3 and HDZI-4 showed enhanced tolerance towards frost in wheat and drought resistance in barley [155]. Expression of SodERF3 was induced by ABA, salt stress and wounding in sugarcane and overexpression of this gene in transgenic tobacco plants increased tolerance to drought and osmotic stresses [156]
5.3. Function and Expression Pattern of AP2/ERF TFs under Biotic Stresses
AP2/ERF TFs were associated with the regulation of disease resistance in plants. Overexpression of TaPIE1 [157], OsEREBP1 [158], Soly106 [159], GmERF113 [160] and OsERF83 [161] genes, in response to JA, SA or ET mediated signal transduction pathways, were believed to be effective against pathogen infection. HvRAF, a novel AP2/ERF TF in barley, had dual regulatory functions under biotic (R. solanacearum) and abiotic (salinity) stresses [162]. In wheat, overexpression of TaPIEP1 was significantly induced in response to Bipolaris sorokiniana infection. The transgenic plants with TaPIEP1 overexpression were exhibited significantly higher resistance against fungal pathogen [163]. In addition to NAC and WRKY, several AP2/ERF TFs were differently expressed in resistant (CLN2777A) and susceptible (TMXA48-4-0) tomato cultivars in response to TYLCV [164]. A pepper CaERF5 played a crucial role in protecting transgenic tobacco plants against R. solanacearum infection [165]. More recently, significantly higher resistance in Arabidopsis was observed by ectopic expression of an apple MdERF11 in response to Botryosphaeria dothidea [166]. Collectively, AP2/ERF TFs play a vital role in abiotic and biotic stresses endurance through different stress-mediated signal transduction pathways.
6. Conclusion and Future Prospects
TFs play pivotal roles at transcriptional level by either suppressing or activating genes under diverse stresses. Approximately 7% coding capacity of vascular plant genome is attributed to TFs for regulating genes at transcriptional level [167]. Thousands of TFs were identified in plants, while major TF families (WRKY, NAC, MYB, AP2/ERF, etc.), mediated through different signal transduction pathways, have been used to cope with abiotic and biotic stresses in various crops during the last two decades. However, more extensive field studies need to be conducted to identify the applications of TF genes in producing stress-resistant crops with high productivity to ensure food security. Although TFs as mediators of stresses have been used to produce stress-tolerant plants, advanced gene editing technologies such as CRISPR/Cas9 are powerful tools and can be explored in the future. To date, apart from the WRKY gene family [13], NAC, MYB and AP2/ERF TFs have not been identified in Saccharum species at a genome wide level due to its complex polyploid genome. There is an urgent need for employing molecular breeding tools to improve sugarcane cultivars, a vegetative propagation crop, with narrow genetic pools and a complicated genome. In addition, plant epigenetics, which is a conserved regulatory mechanism in gene expression, includes DNA methylation, histone modification, chromatin remodeling and noncoding RNA etc., which is an emerging tool to better understand the biological processes in sugarcane environmental responses.
Author Contributions
T.J. and R.S. equally contributed to original draft preparation. A.A. and U.Z. assisted in part data collection. I.A. helped the critical review and editing. S.-J.G. conceived the project and critical review of the manuscript. All authors have read and approved the final manuscript.
Funding
This work was funded by an earmark fund from the China Agriculture Research System, grant number (CARS-170302).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhadouria, R.; Singh, R.; Singh, V.K.; Borthakur, A.; Ahamad, A.; Kumar, G.; Singh, P. Agriculture in the era of climate change: Consequences and effects. In Climate Change and Agricultural Ecosystems; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–23. [Google Scholar]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Khan, M.; Sharif, R.; Mujtaba, M.; Gao, S.-J. Sugarcane Omics: An update on the current status of research and crop improvement. Plants 2019, 8, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraire-Velázquez, S.; Rodríguez-Guerra, R.; Sánchez-Calderón, L. Abiotic and biotic stress response crosstalk in plants. In Abiotic Stress Response in Plants—Physiological, Biochemical and Genetic Perspectives; IntechOpen: London, UK, 2011; pp. 3–26. [Google Scholar]
- Gonzalez, D.H. Introduction to transcription factor structure and function. In Plant Transcription Factors; Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–11. [Google Scholar]
- Inukai, S.; Kock, K.H.; Bulyk, M.L. Transcription factor–DNA binding: Beyond binding site motifs. Curr. Opin. Genet. Deve. 2017, 43, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant. Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, G.; Joyia, F.A.; Anwar, S.; Parvaiz, A.; Khan, M.S. Biotechnological interventions for the improvement of sugarcane crop and sugar production. In Sugarcane-Technology and Research; IntechOpen: London, UK, 2018; pp. 113–138. [Google Scholar]
- Chen, F.; Hu, Y.; Vannozzi, A.; Wu, K.; Cai, H.; Qin, Y.; Mullis, A.; Lin, Z.; Zhang, L. The WRKY transcription factor family in model plants and crops. Crit. Rev. Plant. Sci. 2017, 36, 311–335. [Google Scholar] [CrossRef]
- Ishiguro, S.; Nakamura, K. Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and β-amylase from sweet potato. Mol. Gen. Genet. 1994, 244, 563–571. [Google Scholar] [CrossRef]
- Rinerson, C.I.; Rabara, R.C.; Tripathi, P.; Shen, Q.J.; Rushton, P.J. The evolution of WRKY transcription factors. BMC Plant. Biol. 2015, 15, 66. [Google Scholar] [CrossRef] [Green Version]
- Bao, W.; Wang, X.; Chen, M.; Chai, T.; Wang, H. A WRKY transcription factor, PcWRKY33, from Polygonum cuspidatum reduces salt tolerance in transgenic Arabidopsis thaliana. Plant. Cell Rep. 2018, 37, 1033–1048. [Google Scholar] [CrossRef]
- Li, Z.; Hua, X.; Zhong, W.; Yuan, Y.; Wang, Y.; Wang, Z.; Ming, R.; Zhang, J. Genome-wide identification and expression profile analysis of WRKY family genes in the autopolyploid Saccharum spontaneum. Plant. Cell Physiol. 2020, 61, 616–630. [Google Scholar] [CrossRef]
- Uluhan, E.; Keleş, E.N.; Tufan, F. Analysis of WRKY transcription factors in barley cultivars infected with Fusarium culmorum. Int. J. Life Sci. Biotechnol. 2019, 2, 165–174. [Google Scholar] [CrossRef]
- Lilly, J.J.; Subramanian, B. Gene network mediated by WRKY13 to regulate resistance against sheath infecting fungi in rice (Oryza sativa L.). Plant. Sci. 2019, 280, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Xu, X.; Zhang, L.; Wu, P.; Chen, Y.; Li, M.; Jiang, H.; Wu, G. Genome-wide analysis of the WRKY gene family in physic nut (Jatropha curcas L.). Gene 2013, 524, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-T.; Ru, J.-N.; Liu, Y.-W.; Yang, J.-F.; Li, M.; Xu, Z.-S.; Fu, J.-D. The maize WRKY transcription factor ZmWRKY40 confers drought resistance in transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19, 2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wu, J.; Shang, X.; Geng, M.; Gao, J.; Zhao, S.; Yu, X.; Liu, D.; Kang, Z.; Wang, X. WRKY transcription factors shared by BTH-induced resistance and NPR1-mediated acquired resistance improve broad-spectrum disease resistance in wheat. Mol. Plant. Microbe Interact. 2019, 33, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, L.; Wei, H.; Wang, H.; Su, J.; Yu, S. Characterization and functional analysis of GhWRKY42, a group IId WRKY gene, in upland cotton (Gossypium hirsutum L.). BMC Genet. 2018, 19, 48. [Google Scholar] [CrossRef]
- Goyal, P.; Manzoor, M.M.; Vishwakarma, R.A.; Sharma, D.; Dhar, M.K.; Gupta, S. A comprehensive transcriptome-wide identification and screening of WRKY gene family engaged in abiotic stress in Glycyrrhiza glabra. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant. Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Brand, L.H.; Fischer, N.M.; Harter, K.; Kohlbacher, O.; Wanke, D. Elucidating the evolutionary conserved DNA-binding specificities of WRKY transcription factors by molecular dynamics and in vitro binding assays. Nucleic Acids Res. 2013, 41, 9764–9778. [Google Scholar] [CrossRef]
- Ciolkowski, I.; Wanke, D.; Birkenbihl, R.P.; Somssich, I.E. Studies on DNA-binding selectivity of WRKY transcription factors lend structural clues into WRKY-domain function. Plant. Mol. Biol. 2008, 68, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant. 2018, 162, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Fu, Q.; Chen, L.; Huang, W.; Yu, D. Arabidopsis thaliana WRKY25, WRKY26, and WRKY33 coordinate induction of plant thermotolerance. Planta 2011, 233, 1237–1252. [Google Scholar] [CrossRef] [PubMed]
- He, G.-H.; Xu, J.-Y.; Wang, Y.-X.; Liu, J.-M.; Li, P.-S.; Chen, M.; Ma, Y.-Z.; Xu, Z.-S. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis. BMC Plant. Biol. 2016, 16, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tao, F.; An, F.; Zou, Y.; Tian, W.; Chen, X.; Xu, X.; Hu, X. Wheat transcription factor TaWRKY70 is positively involved in high-temperature seedling plant resistance to Puccinia striiformis f. sp. tritici. Mol. Plant. Pathol. 2017, 18, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Mishra, V.K.; Kumari, S.; Chand, R.; Varadwaj, P.K. Deciphering genome-wide WRKY gene family of Triticum aestivum L. and their functional role in response to abiotic stress. Genes Genom. 2019, 41, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, T.; Sun, X.; Wang, Y.; Du, C.; Zhu, Z.; Gichuki, D.K.; Wang, Q.; Li, S.; Xin, H. Overexpression of VaWRKY12, a transcription factor from Vitis amurensis with increased nuclear localization under low temperature, enhances cold tolerance of plants. Plant. Mol. Biol. 2019, 100, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-T.; Ru, J.-N.; Liu, Y.-W.; Li, M.; Zhao, D.; Yang, J.-F.; Fu, J.-D.; Xu, Z.-S. Maize WRKY transcription factor ZmWRKY106 confers drought and heat tolerance in transgenic plants. Int. J. Mol. Sci. 2018, 19, 3046. [Google Scholar] [CrossRef] [Green Version]
- Yadav, G.; Jat, H. Integrated farming system approaches for managing saline and waterlogged ecologies. In Research Developments in Saline Agriculture; Springer: Berlin/Heidelberg, Germany, 2019; pp. 753–768. [Google Scholar]
- Hsu, F.-C.; Chou, M.-Y.; Chou, S.-J.; Li, Y.-R.; Peng, H.-P.; Shih, M.-C. Submergence confers immunity mediated by the WRKY22 transcription factor in Arabidopsis. Plant. Cell 2013, 25, 2699–2713. [Google Scholar] [CrossRef] [Green Version]
- Raineri, J.; Ribichich, K.F.; Chan, R.L. The sunflower transcription factor HaWRKY76 confers drought and flood tolerance to Arabidopsis thaliana plants without yield penalty. Plant. Cell Rep. 2015, 34, 2065–2080. [Google Scholar] [CrossRef]
- Han, C.; Li, J.; Ma, Y.; Guo, J.; Guo, X.; Xu, J. PlWRKY70: A Paeonia lactiflora transcription factor that sensitively responds to low-temperature, salt and waterlogging stresses. Can. J. Plant. Sci. 2019, 100, 146–155. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [Green Version]
- Babitha, K.; Ramu, S.; Pruthvi, V.; Mahesh, P.; Nataraja, K.N.; Udayakumar, M. Co-expression of AtbHLH17 and AtWRKY28 confers resistance to abiotic stress in Arabidopsis. Transgenic Res. 2013, 22, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought responses. Plant. Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, F.; Zhang, X.; Wang, W.; Sun, T.; Chen, Y.; Dai, M.; Yu, S.; Xu, L.; Su, Y.; et al. Expression characteristics and functional analysis of the ScWRKY3 gene from sugarcane. Int. J. Mol. Sci. 2018, 19, 4059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Tian, Y.; Liu, X. A wheat salinity-induced WRKY transcription factor TaWRKY93 confers multiple abiotic stress tolerance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 428–433. [Google Scholar] [CrossRef]
- Gao, H.; Wang, Y.; Xu, P.; Zhang, Z. Overexpression of a WRKY transcription factor TaWRKY2 enhances drought stress tolerance in transgenic wheat. Front. Plant. Sci. 2018, 9, 997. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Ma, Q.; Zhang, C.; Wang, C.; Wei, H.; Wang, H.; Yu, S. The Cotton GhWRKY91 Transcription factor mediates leaf senescence and responses to drought stress in transgenic Arabidopsis thaliana. Front. Plant. Sci. 2019, 10, 1352. [Google Scholar] [CrossRef]
- Wei, W.; Liang, D.W.; Bian, X.H.; Shen, M.; Xiao, J.H.; Zhang, W.K.; Ma, B.; Lin, Q.; Lv, J.; Chen, X. GmWRKY54 improves drought tolerance through activating genes in abscisic acid and Ca2+ signaling pathways in transgenic soybean. Plant. J. 2019, 100, 384–398. [Google Scholar] [CrossRef]
- Thomas, T.D.; Dinakar, C.; Puthur, J.T. Effect of UV-B priming on the abiotic stress tolerance of stress-sensitive rice seedlings: Priming imprints and cross-tolerance. Plant. Physiol. Biochem. 2020, 147, 21–30. [Google Scholar] [CrossRef]
- Wang, H.; Hao, J.; Chen, X.; Hao, Z.; Wang, X.; Lou, Y.; Peng, Y.; Guo, Z. Overexpression of rice WRKY89 enhances ultraviolet B tolerance and disease resistance in rice plants. Plant. Mol. Biol. 2007, 65, 799–815. [Google Scholar] [CrossRef]
- Marchica, A.; Lorenzini, G.; Papini, R.; Bernardi, R.; Nali, C.; Pellegrini, E. Signalling molecules responsive to ozone-induced oxidative stress in Salvia officinalis. Sci. Total Environ. 2019, 657, 568–576. [Google Scholar] [CrossRef]
- Garg, N.; Manchanda, G. ROS generation in plants: Boon or bane? Plant. Biosys. 2009, 143, 81–96. [Google Scholar] [CrossRef]
- Luan, Q.; Chen, C.; Liu, M.; Li, Q.; Wang, L.; Ren, Z. CsWRKY50 mediates defense responses to Pseudoperonospora cubensis infection in Cucumis sativus. Plant. Sci. 2019, 279, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Marchive, C.; Léon, C.; Kappel, C.; Coutos-Thévenot, P.; Corio-Costet, M.-F.; Delrot, S.; Lauvergeat, V. Over-expression of VvWRKY1 in grapevines induces expression of jasmonic acid pathway-related genes and confers higher tolerance to the downy mildew. PLoS ONE 2013, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, F.; Wang, Y.; She, J.; Lei, Y.; Liu, Z.; Eulgem, T.; Lai, Y.; Lin, J.; Yu, L.; Lei, D. Overexpression of CaWRKY27, a subgroup IIe WRKY transcription factor of Capsicum annuum, positively regulates tobacco resistance to Ralstonia solanacearum infection. Physiologia Plant. 2014, 150, 397–411. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Wang, N.; Guo, X.; Gao, Z. GhWRKY44, a WRKY transcription factor of cotton, mediates defense responses to pathogen infection in transgenic Nicotiana benthamiana. Plant. Cell. Tiss. Org. Cult. 2015, 121, 127–140. [Google Scholar] [CrossRef]
- Singh, A.K.; Kumar, S.R.; Dwivedi, V.; Rai, A.; Pal, S.; Shasany, A.K.; Nagegowda, D.A. A WRKY transcription factor from Withania somnifera regulates triterpenoid withanolide accumulation and biotic stress tolerance through modulation of phytosterol and defense pathways. New Phytol. 2017, 215, 1115–1131. [Google Scholar] [CrossRef] [Green Version]
- Jing, Z.; Liu, Z. Genome-wide identification of WRKY transcription factors in kiwifruit (Actinidia spp.) and analysis of WRKY expression in responses to biotic and abiotic stresses. Genes Genom. 2018, 40, 429–446. [Google Scholar] [CrossRef]
- Dong, H.; Tan, J.; Li, M.; Yu, Y.; Jia, S.; Zhang, C.; Wu, Y.; Liu, Y. Transcriptome analysis of soybean WRKY TFs in response to Peronospora manshurica infection. Genomics 2019, 111, 1412–1422. [Google Scholar] [CrossRef]
- Ntambo, M.S.; Meng, J.-Y.; Rott, P.C.; Henry, R.J.; Zhang, H.-L.; Gao, S.-J. Comparative transcriptome profiling of resistant and susceptible sugarcane cultivars in response to infection by Xanthomonas albilineans. Int. J. Mol. Sci. 2019, 20, 6138. [Google Scholar] [CrossRef] [Green Version]
- Jensen, M.K.; Kjaersgaard, T.; Nielsen, M.M.; Galberg, P.; Petersen, K.; O’shea, C.; Skriver, K. The Arabidopsis thaliana NAC transcription factor family: Structure–function relationships and determinants of ANAC019 stress signalling. Biochem. J. 2010, 426, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Sablowski, R.W.; Meyerowitz, E.M. A homolog of NO APICAL MERISTEM is an immediate target of the floral homeotic genes APETALA3/PISTILLATA. Cell 1998, 92, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: An analysis of the cup-shaped cotyledon mutant. Plant. Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant. Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC proteins: Regulation and role in stress tolerance. Trends Plant. Sci. 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Ernst, H.A.; Olsen, A.N.; Skriver, K.; Larsen, S.; Leggio, L.L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kim, S.-G.; Kim, Y.-S.; Seo, P.J.; Bae, M.; Yoon, H.-K.; Park, C.-M. Exploring membrane-associated NAC transcription factors in Arabidopsis: Implications for membrane biology in genome regulation. Nucleic Acids Res. 2007, 35, 203–213. [Google Scholar] [CrossRef]
- Liang, M.; Li, H.; Zhou, F.; Li, H.; Liu, J.; Hao, Y.; Wang, Y.; Zhao, H.; Han, S. Subcellular distribution of NTL transcription factors in Arabidopsis thaliana. Traffic 2015, 16, 1062–1074. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Bita, C.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant. Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [Green Version]
- Ditt, R.F.; Gentile, A.; Tavares, R.G.; Camargo, S.R.; Fernandez, J.H.; Silva, M.J.d.; Menossi, M. Analysis of the stress-inducible transcription factor SsNAC23 in sugarcane plants. Sci. Agric. 2011, 68, 454–461. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant. J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant. Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, Y.; Zhang, Y.; Lyu, Y. A Stress-Responsive NAC transcription factor from tiger lily (LlNAC2) interacts with LlDREB1 and LlZHFD4 and enhances various abiotic stress tolerance in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 3225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.-m.; Zhang, H.-f.; Liu, S.-y.; Wang, X.-k.; Zhang, Y.-m.; Meng, Y.-c.; Luo, D.; Chen, R.-g. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant. Sci. 2020, 291, 110346. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.; Shukla, A. Acclimation and tolerance strategies of rice under drought stress. Rice Sci. 2015, 22, 147–161. [Google Scholar] [CrossRef] [Green Version]
- Tran, L.-S.P.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant. Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.-H.; Do Choi, Y.; Kim, M.; Reuzeau, C.; Kim, J.-K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant. Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.K.; Chung, P.J.; Jeong, J.S.; Jang, G.; Bang, S.W.; Jung, H.; Kim, Y.S.; Ha, S.H.; Choi, Y.D.; Kim, J.K. The rice Os NAC 6 transcription factor orchestrates multiple molecular mechanisms involving root structural adaptions and nicotianamine biosynthesis for drought tolerance. Plant. Biotechnol. J. 2017, 15, 754–764. [Google Scholar] [CrossRef]
- Rahman, H.; Ramanathan, V.; Nallathambi, J.; Duraialagaraja, S.; Muthurajan, R. Over-expression of a NAC 67 transcription factor from finger millet (Eleusine coracana L.) confers tolerance against salinity and drought stress in rice. BMC Biotechnol. 2016, 16, 35. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, L.; Xia, C.; Zhao, G.; Jia, J.; Kong, X. The novel wheat transcription factor TaNAC47 enhances multiple abiotic stress tolerances in transgenic plants. Front. Plant. Sci. 2016, 6, 1174. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant. Biol. 2019, 19, 278. [Google Scholar] [CrossRef] [PubMed]
- Gunapati, S.; Naresh, R.; Ranjan, S.; Nigam, D.; Hans, A.; Verma, P.C.; Gadre, R.; Pathre, U.V.; Sane, A.P.; Sane, V.A. Expression of GhNAC2 from G. herbaceum, improves root growth and imparts tolerance to drought in transgenic cotton and Arabidopsis. Sci. Rep. 2016, 6, 24978. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Li, K.-Q.; Xu, X.-Y.; Zhang, H.-P.; Chen, H.-X.; Chen, Y.-H.; Hao, J.; Wang, Y.; Huang, X.-S.; Zhang, S.-L. A novel NAC transcription factor, PbeNAC1, of Pyrus betulifolia confers cold and drought tolerance via interacting with PbeDREBs and activating the expression of stress-responsive genes. Front. Plant. Sci. 2017, 8, 1049. [Google Scholar] [CrossRef] [Green Version]
- Sakuraba, Y.; Kim, Y.-S.; Han, S.-H.; Lee, B.-D.; Paek, N.-C. The Arabidopsis transcription factor NAC016 promotes drought stress responses by repressing AREB1 transcription through a trifurcate feed-forward regulatory loop involving NAP. Plant. Cell 2015, 27, 1771–1787. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhou, Y.; Li, H.; Wang, T.; Zhang, J.; Ouyang, B.; Ye, Z. Molecular and functional characterization of ShNAC1, an NAC transcription factor from Solanum habrochaites. Plant. Sci. 2018, 271, 9–19. [Google Scholar] [CrossRef]
- Balazadeh, S.; Siddiqui, H.; Allu, A.D.; Matallana-Ramirez, L.P.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Köhler, B.; Mueller-Roeber, B. A gene regulatory network controlled by the NAC transcription factor ANAC092/AtNAC2/ORE1 during salt-promoted senescence. Plant. J. 2010, 62, 250–264. [Google Scholar] [CrossRef]
- You, J.; Zong, W.; Li, X.; Ning, J.; Hu, H.; Li, X.; Xiao, J.; Xiong, L. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Chen, S.; Li, A.; Zhai, C.; Jing, R. Novel NAC transcription factor TaNAC67 confers enhanced multi-abiotic stress tolerances in Arabidopsis. PLoS ONE 2014, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Wang, Y.; Li, B.; Chang, J.; Chen, M.; Li, K.; Yang, G.; He, G. TaNAC29, a NAC transcription factor from wheat, enhances salt and drought tolerance in transgenic Arabidopsis. BMC Plant. Biol. 2015, 15, 268. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Li, Z.; Lu, H.; Huo, L.; Wang, Z.; Wang, Y.; Ji, X. The NAC protein from Tamarix hispida, ThNAC7, confers salt and osmotic stress tolerance by increasing reactive oxygen species scavenging capability. Plants 2019, 8, 221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Wang, H.; Cai, J.; Li, D.; Song, F. NAC transcription factors in plant immunity. Phytopath. Res. 2019, 1, 3. [Google Scholar] [CrossRef]
- Xia, N.; Zhang, G.; Sun, Y.-F.; Zhu, L.; Xu, L.-S.; Chen, X.-M.; Liu, B.; Yu, Y.-T.; Wang, X.-J.; Huang, L.-L. TaNAC8, a novel NAC transcription factor gene in wheat, responds to stripe rust pathogen infection and abiotic stresses. Physiol. Mol. Plant. Pathol. 2010, 74, 394–402. [Google Scholar] [CrossRef]
- Voitsik, A.-M.; Muench, S.; Deising, H.B.; Voll, L.M. Two recently duplicated maize NAC transcription factor paralogs are induced in response to Colletotrichum graminicola infection. BMC Plant. Biol. 2013, 13, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Wei, J.; Song, N.; Wang, N.; Zhao, J.; Kang, Z. A novel wheat NAC transcription factor, TaNAC30, negatively regulates resistance of wheat to stripe rust. J. Integr. Plant. Biol. 2018, 60, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-M.; Zhang, Q.; Pei, C.-L.; Li, X.; Huang, X.-L.; Chang, C.-Y.; Wang, X.-J.; Huang, L.-L.; Kang, Z.-S. TaNAC2 is a negative regulator in the wheat-stripe rust fungus interaction at the early stage. Physiol. Mol. Plant. P. 2018, 102, 144–153. [Google Scholar] [CrossRef]
- Huang, Y.; Li, T.; Xu, Z.-S.; Wang, F.; Xiong, A.-S. Six NAC transcription factors involved in response to TYLCV infection in resistant and susceptible tomato cultivars. Plant. Physiol. Biochem. 2017, 120, 61–74. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, H.; Li, D.; Huang, L.; Hong, Y.; Ding, X.S.; Nelson, R.S.; Zhou, X.; Song, F. Functions of rice NAC transcriptional factors, ONAC122 and ONAC131, in defense responses against Magnaporthe grisea. Plant. Mol. Biol. 2013, 81, 41–56. [Google Scholar] [CrossRef]
- Du, M.; Zhai, Q.; Deng, L.; Li, S.; Li, H.; Yan, L.; Huang, Z.; Wang, B.; Jiang, H.; Huang, T. Closely related NAC transcription factors of tomato differentially regulate stomatal closure and reopening during pathogen attack. Plant. Cell 2014, 26, 3167–3184. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Jeon, H.S.; Kim, H.G.; Park, O.K. An Arabidopsis NAC transcription factor NAC4 promotes pathogen-induced cell death under negative regulation by microRNA164. New Phytol. 2017, 214, 343–360. [Google Scholar] [CrossRef]
- Meisrimler, C.N.; Pelgrom, A.J.; Oud, B.; Out, S.; Van den Ackerveken, G. Multiple downy mildew effectors target the stress-related NAC transcription factor LsNAC069 in lettuce. Plant. J. 2019, 99, 1098–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, S. Function of MYB domain transcription factors in abiotic stress and epigenetic control of stress response in plant genome. Plant. Signal. Behav. 2016, 11, e1117723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambawat, S.; Sharma, P.; Yadav, N.R.; Yadav, R.C. MYB transcription factor genes as regulators for plant responses: An overview. Physiol. Mol. Biol. Plants 2013, 19, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz-Ares, J.; Ghosal, D.; Wienand, U.; Peterson, P.; Saedler, H. The regulatory c1 locus of Zea mays encodes a protein with homology to myb proto-oncogene products and with structural similarities to transcriptional activators. EMBO J. 1987, 6, 3553–3558. [Google Scholar] [CrossRef]
- Hajiebrahimi, A.; Owji, H.; Hemmati, S. Genome-wide identification, functional prediction, and evolutionary analysis of the R2R3-MYB superfamily in Brassica napus. Genome 2017, 60, 797–814. [Google Scholar] [CrossRef]
- Ogata, K.; Kanei-Ishii, C.; Sasaki, M.; Hatanaka, H.; Nagadoi, A.; Enari, M.; Nakamura, H.; Nishimura, Y.; Ishii, S.; Sarai, A. The cavity in the hydrophobic core of Myb DNA-binding domain is reserved for DNA recognition and trans-activation. Nat. Struct. Biol. 1996, 3, 178. [Google Scholar] [CrossRef]
- Mmadi, M.A.; Dossa, K.; Wang, L.; Zhou, R.; Wang, Y.; Cisse, N.; Sy, M.O.; Zhang, X. Functional characterization of the versatile MYB gene family uncovered their important roles in plant development and responses to drought and waterlogging in sesame. Genes 2017, 8, 362. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Wang, J.-R.; Wang, G.-D.; Liang, X.-Q.; Li, X.-D.; Meng, Q.-W. An R2R3-MYB gene, LeAN2, positively regulated the thermo-tolerance in transgenic tomato. J. Plant. Physiol. 2015, 175, 1–8. [Google Scholar] [CrossRef]
- Deeba, F.; Sultana, T.; Javaid, B.; Mahmood, T.; Naqvi, S. Molecular characterization of a MYB protein from Oryza sativa for its role in abiotic stress tolerance. Braz. Arch. Biol. Technol. 2017, 60, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Casaretto, J.A.; El-kereamy, A.; Zeng, B.; Stiegelmeyer, S.M.; Chen, X.; Bi, Y.-M.; Rothstein, S.J. Expression of OsMYB55 in maize activates stress-responsive genes and enhances heat and drought tolerance. BMC Genom. 2016, 17, 312. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Tian, X.; Wang, F.; Zhang, L.; Xin, M.; Hu, Z.; Yao, Y.; Ni, Z.; Sun, Q.; Peng, H. Characterization of wheat MYB genes responsive to high temperatures. BMC Plant. Biol. 2017, 17, 208. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Liu, Y.; Zhao, L.; Zhang, S.; Huang, X. A novel MYB transcription factor regulates ascorbic acid synthesis and affects cold tolerance. Plant. Cell Environ. 2019, 42, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Baldoni, E.; Genga, A.; Cominelli, E. Plant MYB transcription factors: Their role in drought response mechanisms. Int. J. Mol. Sci. 2015, 16, 15811–15851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katiyar, A.; Smita, S.; Lenka, S.K.; Rajwanshi, R.; Chinnusamy, V.; Bansal, K.C. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis. BMC Genom. 2012, 13, 544. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.S.; Sohn, H.B.; Noh, K.; Jung, C.; An, J.H.; Donovan, C.M.; Somers, D.A.; Kim, D.I.; Jeong, S.-C.; Kim, C.-G. Expression of the Arabidopsis AtMYB44 gene confers drought/salt-stress tolerance in transgenic soybean. Mol. Breed. 2012, 29, 601–608. [Google Scholar] [CrossRef]
- Chen, B.-J.; Wang, Y.; Hu, Y.-L.; Wu, Q.; Lin, Z.-P. Cloning and characterization of a drought-inducible MYB gene from Boea crassifolia. Plant. Sci. 2005, 168, 493–500. [Google Scholar] [CrossRef]
- Yu, Y.-T.; Wu, Z.; Lu, K.; Bi, C.; Liang, S.; Wang, X.-F.; Zhang, D.-P. Overexpression of the MYB37 transcription factor enhances abscisic acid sensitivity, and improves both drought tolerance and seed productivity in Arabidopsis thaliana. Plant. Mol. Biol. 2016, 90, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Prabu, G.; Prasad, D.T. Functional characterization of sugarcane MYB transcription factor gene promoter (PScMYBAS1) in response to abiotic stresses and hormones. Plant. Cell Rep. 2012, 31, 661–669. [Google Scholar] [CrossRef]
- Fávero, P.J.R.; Mara, D.A.L.; Dos, S.B.M. Overexpression of ScMYBAS1 alternative splicing transcripts differentially impacts biomass accumulation and drought tolerance in rice transgenic plants. PLoS ONE 2018, 13, e0207534. [Google Scholar]
- Guo, J.; Ling, H.; Ma, J.; Chen, Y.; Su, Y.; Lin, Q.; Gao, S.; Wang, H.; Que, Y.; Xu, L. A sugarcane R2R3-MYB transcriprion factor gene is alternatively spliced during drought stress. Sci. Rep. 2017, 7, 41922. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Dai, X.; Zhang, W.-H. A R2R3-type MYB gene, OsMYB2, is involved in salt, cold, and dehydration tolerance in rice. J. Exp. Bot. 2012, 63, 2541–2556. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Li, J.; Liu, P.; Duan, J.; Zhao, Y.; Guo, X.; Li, Y.; Zhang, H.; Ali, J.; Li, Z. Overexpression of OsMYB48-1, a novel MYB-related transcription factor, enhances drought and salinity tolerance in rice. PLoS ONE 2014, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Zhang, F.; Sun, F.; Luo, Q.; Wang, R.; Hu, R.; Chen, M.; Chang, J.; Yang, G.; He, G. A wheat MYB transcriptional repressor TaMyb1D regulates phenylpropanoid metabolism and enhances tolerance to drought and oxidative stresses in transgenic tobacco plants. Plant. Sci. 2017, 265, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Cominelli, E.; Bailey, P.; Parr, A.; Mehrtens, F.; Jones, J.; Tonelli, C.; Weisshaar, B.; Martin, C. Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in Arabidopsis. EMBO J. 2000, 19, 6150–6161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Osbourn, A.; Ma, P. MYB transcription factors as regulators of phenylpropanoid metabolism in plants. Mol. Plant. 2015, 8, 689–708. [Google Scholar] [CrossRef] [Green Version]
- Li, X.W.; Wang, Y.; Yan, F.; Li, J.W.; Zhao, Y.; Zhao, X.; Zhai, Y.; Wang, Q.Y. Overexpression of soybean R2R3-MYB transcription factor, GmMYB12B2, and tolerance to UV radiation and salt stress in transgenic Arabidopsis. Genet. Mol. Res. 2016, 15, 6573. [Google Scholar] [CrossRef]
- Zhu, L.; Guo, J.; Ma, Z.; Wang, J.; Zhou, C. Arabidopsis transcription factor MYB102 increases plant susceptibility to aphids by substantial activation of ethylene biosynthesis. Biomolecules 2018, 8, 39. [Google Scholar] [CrossRef] [Green Version]
- Seo, P.J.; Park, C.M. MYB96-mediated abscisic acid signals induce pathogen resistance response by promoting salicylic acid biosynthesis in Arabidopsis. New Phytol. 2010, 186, 471–483. [Google Scholar] [CrossRef]
- Segarra, G.; Van der Ent, S.; Trillas, I.; Pieterse, C. MYB72, a node of convergence in induced systemic resistance triggered by a fungal and a bacterial beneficial microbe. Plant. Biol. 2009, 11, 90–96. [Google Scholar] [CrossRef] [Green Version]
- Shan, T.; Rong, W.; Xu, H.; Du, L.; Liu, X.; Zhang, Z. The wheat R2R3-MYB transcription factor TaRIM1 participates in resistance response against the pathogen Rhizoctonia cerealis infection through regulating defense genes. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Ibraheem, F.; Gaffoor, I.; Tan, Q.; Shyu, C.-R.; Chopra, S. A sorghum MYB transcription factor induces 3-deoxyanthocyanidins and enhances resistance against leaf blights in maize. Molecules 2015, 20, 2388–2404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Sheng, L.; Du, X.; Wang, Y.; Zhang, Y.; Song, A.; Jiang, J.; Guan, Z.; Fang, W.; Chen, F. Overexpression of CmMYB15 provides chrysanthemum resistance to aphids by regulating the biosynthesis of lignin. Hortic. Res. 2019, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.-L.; Zhang, C.-L.; Wang, G.-L.; Wang, Y.-X.; Qi, C.-H.; Zhao, Q.; You, C.-X.; Li, Y.-Y.; Hao, Y.-J. The R2R3 MYB transcription factor MdMYB30 modulates plant resistance against pathogens by regulating cuticular wax biosynthesis. BMC Plant. Biol. 2019, 19, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noman, A.; Hussain, A.; Adnan, M.; Khan, M.I.; Ashraf, M.F.; Zainab, M.; Khan, K.A.; Ghramh, H.A.; He, S. A novel MYB transcription factor CaPHL8 provide clues about evolution of pepper immunity againstsoil borne pathogen. Microb. Pathogen. 2019, 137, 103758. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Guo, D.; Li, G.; Yang, Y.; Zhang, G.; Li, S.; Liang, Z. The grapevine R2R3-type MYB transcription factor VdMYB1 positively regulates defense responses by activating the stilbene synthase gene 2 (VdSTS2). BMC Plant. Biol. 2019, 19, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.; Li, Y.; Hou, X. Genome-wide analysis of the AP2/ERF transcription factor superfamily in Chinese cabbage (Brassica rapa ssp. pekinensis). BMC Genom. 2013, 14, 573. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, Y.; Liu, Q.; Dubouzet, J.G.; Abe, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration-and cold-inducible gene expression. Biochem. Biophys. Res. Commun. 2002, 290, 998–1009. [Google Scholar] [CrossRef]
- Niu, X.; Luo, T.; Zhao, H.; Su, Y.; Ji, W.; Li, H. Identification of wheat DREB genes and functional characterization of TaDREB3 in response to abiotic stresses. Gene 2020, 740. [Google Scholar] [CrossRef]
- Dong, C.-J.; Liu, J.-Y. The Arabidopsis EAR-motif-containing protein RAP2.1 functions as an active transcriptional repressor to keep stress responses under tight control. BMC Plant. Biol. 2010, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Xiong, A.-S.; Jiang, H.-H.; Zhuang, J.; Peng, R.-H.; Jin, X.-F.; Zhu, B.; Wang, F.; Zhang, J.; Yao, Q.-H. Expression and function of a modified AP2/ERF transcription factor from Brassica napus enhances cold tolerance in transgenic Arabidopsis. Mol. Biotechnol. 2013, 53, 198–206. [Google Scholar] [CrossRef]
- Zhao, T.-J.; Sun, S.; Liu, Y.; Liu, J.-M.; Liu, Q.; Yan, Y.-B.; Zhou, H.-M. Regulating the drought-responsive element (DRE)-mediated signaling pathway by synergic functions of trans-active and trans-inactive DRE binding factors in Brassica napus. J. Biol. Chem. 2006, 281, 10752–10759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Illgen, S.; Zintl, S.; Zuther, E.; Hincha, D.; Schmulling, T. Characterisation of the ERF102 to ERF105 genes of Arabidopsis thaliana and their role in the response to cold stress. Plant Mol. Biol. 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhu, Z.; Zhang, L.; Fang, L.; Zhang, J.; Wang, Q.; Li, S.; Liang, Z.; Xin, H. Overexpression of ethylene response factors VaERF080 and VaERF087 from Vitis amurensis enhances cold tolerance in Arabidopsis. Sci. Hortic. 2019, 243, 320–326. [Google Scholar] [CrossRef]
- Lv, K.; Li, J.; Zhao, K.; Chen, S.; Nie, J.; Zhang, W.; Liu, G.; Wei, H. Overexpression of an AP2/ERF family gene, BpERF13, in birch enhances cold tolerance through upregulating CBF genes and mitigating reactive oxygen species. Plant. Sci. 2020, 292, 110375. [Google Scholar] [CrossRef]
- Moon, S.-J.; Min, M.K.; Kim, J.; Kim, D.Y.; Yoon, I.S.; Kwon, T.R.; Byun, M.O.; Kim, B.-G. Ectopic expression of OsDREB1G, a member of the OsDREB1 subfamily, confers cold stress tolerance in rice. Front. Plant. Sci. 2019, 10, 297. [Google Scholar] [CrossRef]
- Gomathi, R.; Rao, P.N.G.; Chandran, K.; Selvi, A. Adaptive responses of sugarcane to waterlogging stress: An over view. Sugar Tech 2015, 17, 325–338. [Google Scholar] [CrossRef]
- Du, H.; Huang, M.; Zhang, Z.; Cheng, S. Genome-wide analysis of the AP2/ERF gene family in maize waterlogging stress response. Euphytica 2014, 198, 115–126. [Google Scholar] [CrossRef]
- Shahzad, Z.; Canut, M.; Tournaire-Roux, C.; Martiniere, A.; Boursiac, Y.; Loudet, O.; Maurel, C. A potassium-dependent oxygen sensing pathway regulates plant root hydraulics. Cell 2016, 167, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Liang, K.; Fang, T.; Zhao, H.; Han, X.; Cai, M.; Qiu, F. A group VII ethylene response factor gene, ZmEREB180, coordinates waterlogging tolerance in maize seedlings. Plant. Biotechnol. J. 2019, 17, 2286–2298. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Xu, H.; Rong, W.; Ye, X.; Zhang, Z. Constitutive expression of a stabilized transcription factor group VII ethylene response factor enhances waterlogging tolerance in wheat without penalizing grain yield. Plant. Cell Environ. 2019, 42, 1471–1485. [Google Scholar] [CrossRef]
- Fan, W.; Yang, Y.; Wang, Z.; Yin, Y.; Yu, C.; Shi, Q.; Guo, J.; Xuan, L.; Hua, J. Molecular cloning and expression analysis of three ThERFs involved in the response to waterlogging stress of Taxodium ‘Zhongshanshan406’, and subcellular localization of the gene products. PeerJ 2018, 6, e4434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.-L.; Ma, J.-T.; Zhao, Y.-M.; Wei, Y.-H.; Tang, Y.-X.; Wu, Y.-M. Improvement of drought and salt tolerance in Arabidopsis and Lotus corniculatus by overexpression of a novel DREB transcription factor from Populus euphratica. Gene 2012, 506, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, Q.-Y.; Cheng, X.-G.; Xu, Z.-S.; Li, L.-C.; Ye, X.-G.; Xia, L.-Q.; Ma, Y.-Z. GmDREB2, a soybean DRE-binding transcription factor, conferred drought and high-salt tolerance in transgenic plants. Biochem. Biophys. Res. Commun. 2007, 353, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Kidokoro, S.; Watanabe, K.; Ohori, T.; Moriwaki, T.; Maruyama, K.; Mizoi, J.; Myint Phyu Sin Htwe, N.; Fujita, Y.; Sekita, S.; Shinozaki, K. Soybean DREB 1/CBF-type transcription factors function in heat and drought as well as cold stress-responsive gene expression. Plant. J. 2015, 81, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Dossa, K.; Wei, X.; Li, D.; Fonceka, D.; Zhang, Y.; Wang, L.; Yu, J.; Boshou, L.; Diouf, D.; Cissé, N. Insight into the AP2/ERF transcription factor superfamily in sesame and expression profiling of DREB subfamily under drought stress. BMC Plant. Biol. 2016, 16, 171. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Wang, Y.; Wang, W.; Zhao, X.; Qin, Q.; Sun, F.; Hu, F.; Zhao, Y.; Li, Z.; Fu, B. Characterization of transcription factor gene OsDRAP1 conferring drought tolerance in rice. Front. Plant. Sci. 2018, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- de Souza, W.R.; de Oliveira, N.G.; Vinecky, F.; Ribeiro, A.P.; Basso, M.F.; Casari, R.A.d.C.N.; da Cunha, B.A.D.B.; Duarte, K.E.; Santiago, T.R.; Martins, P.K. Field evaluation of AtDREB2A CA overexpressing sugarcane for drought tolerance. J. Agron. Crop. Sci. 2019, 205, 545–553. [Google Scholar] [CrossRef]
- Tavakol, E.; Sardaro, M.L.S.; Shariati, V.; Rossini, L.; Porceddu, E. Isolation, promoter analysis and expression profile of Dreb2 in response to drought stress in wheat ancestors. Gene 2014, 549, 24–32. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Z.; Xiao, H.-m.; Yang, Y. Characterization of TaDREB1 in wheat genotypes with different seed germination under osmotic stress. Hereditas 2018, 155, 26. [Google Scholar] [CrossRef]
- Yang, Y.; Al-Baidhani, H.H.J.; Harris, J.; Riboni, M.; Li, Y.; Mazonka, I.; Bazanova, N.; Chirkova, L.; Sarfraz Hussain, S.; Hrmova, M. DREB/CBF expression in wheat and barley using the stress-inducible promoters of HD-Zip I genes: Impact on plant development, stress tolerance and yield. Plant. Biotechnol. J. 2020 18, 829–844. [CrossRef] [Green Version]
- Trujillo, L.E.; Sotolongo, M.; Menendez, C.; Ochogavia, M.E.; Coll, Y.; Hernandez, I.; Borras, H.O.; Thomma, B.P.H.J.; Vera, P.; Hernandez, L. SodERF3, a novel sugarcane ethylene responsive factor (ERF) enhances salt and drought tolerance when overexpressed in tobacco plants. Plant. Cell Physiol. 2008, 49, 512–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Qi, L.; Liu, X.; Cai, S.; Xu, H.; Huang, R.; Li, J.; Wei, X.; Zhang, Z. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant. Physiol. 2014, 164, 1499–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jisha, V.; Dampanaboina, L.; Vadassery, J.; Mithöfer, A.; Kappara, S.; Ramanan, R. Overexpression of an AP2/ERF type transcription factor OsEREBP1 confers biotic and abiotic stress tolerance in rice. PLoS ONE 2015, 10, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Zhang, B.-L.; Sun, S.; Xing, G.-M.; Wang, F.; Li, M.-Y.; Tian, Y.-S.; Xiong, A.-S. AP2/ERF transcription factors involved in response to tomato yellow leaf curly virus in tomato. Plant. Genom. 2016, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chang, X.; Qi, D.; Dong, L.; Wang, G.; Fan, S.; Jiang, L.; Cheng, Q.; Chen, X.; Han, D. A novel soybean ERF transcription factor, GmERF113, increases resistance to Phytophthora sojae infection in soybean. Front. Plant. Sci. 2017, 8, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tezuka, D.; Kawamata, A.; Kato, H.; Saburi, W.; Mori, H.; Imai, R. The rice ethylene response factor OsERF83 positively regulates disease resistance to Magnaporthe oryzae. Plant. Physiol. Biochem. 2019, 135, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Won, S.Y.; Suh, S.C.; Kim, H.; Wing, R.; Jeong, Y.; Hwang, I.; Kim, M. The barley ERF-type transcription factor HvRAF confers enhanced pathogen resistance and salt tolerance in Arabidopsis. Planta 2007, 225, 575–588. [Google Scholar] [CrossRef]
- Dong, N.; Liu, X.; Lu, Y.; Du, L.; Xu, H.; Liu, H.; Xin, Z.; Zhang, Z. Overexpression of TaPIEP1, a pathogen-induced ERF gene of wheat, confers host-enhanced resistance to fungal pathogen Bipolaris sorokiniana. Funct. Integr. Genomic. 2010, 10, 215–226. [Google Scholar] [CrossRef]
- Chen, T.; Lv, Y.; Zhao, T.; Li, N.; Yang, Y.; Yu, W.; He, X.; Liu, T.; Zhang, B. Comparative transcriptome profiling of a resistant vs. susceptible tomato (Solanum lycopersicum) cultivar in response to infection by tomato yellow leaf curl virus. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef]
- Lai, Y.; Dang, F.; Lin, J.; Yu, L.; Lin, J.; Lei, Y.; Chen, C.; Liu, Z.; Qiu, A.; Mou, S. Overexpression of a pepper CaERF5 gene in tobacco plants enhances resistance to Ralstonia solanacearum infection. Funct. Plant. Biol. 2014, 41, 758–767. [Google Scholar] [CrossRef]
- Wang, J.H.; Gu, K.D.; Han, P.L.; Yu, J.Q.; Wang, C.K.; Zhang, Q.Y.; You, C.X.; Hu, D.G.; Hao, Y.J. Apple ethylene response factor MdERF11 confers resistance to fungal pathogen Botryosphaeria dothidea. Plant. Sci. 2020, 291, 110351. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Bokowiec, M.T.; Laudeman, T.W.; Brannock, J.F.; Chen, X.; Timko, M.P. TOBFAC: The database of tobacco transcription factors. BMC Bioinform. 2008, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]

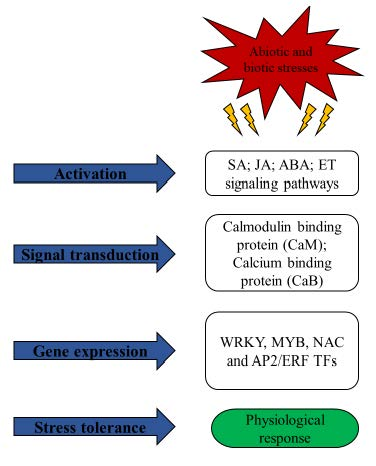{kind=link}
Table 1.
Gene numbers of WRYK, NAC, MYB and AP2/ERF transcription factors in land organisms a.
| Plant Species | Type of Organism | WRKY | NAC | MYB | AP2/ERF |
|---|---|---|---|---|---|
| Saccharum spp. (Sugarcane) | Monocot plant | 39 | 44 | 36 | 6 |
| Sorghum bicolor (Sorghum) | Monocot plant | 134 | 180 | 145 | 42 |
| Triticum aestivum (Wheat) | Monocot plant | 171 | 263 | 263 | 43 |
| Oryza sativa subsp. japonica (Rice) | Monocot plant | 128 | 170 | 130 | 22 |
| Hordeum vulgare (Barley) | Monocot plant | 126 | 150 | 99 | 34 |
| Zea mays (Maize) | Monocot plant | 161 | 189 | 203 | 54 |
| Zoysia matrella (Manila grass) | Monocot plant | 269 | 313 | 293 | 49 |
| Brachypodium distachyon (False brome) | Monocot plant | 134 | 186 | 163 | 64 |
| Beta vulgaris (Sugar beet) | Dicot Plant | 49 | 59 | 68 | 14 |
| Arabidopsis thaliana | Dicot Plant | 90 | 138 | 168 | 30 |
| Nicotiana tabacum (Tobacco) | Dicot Plant | 210 | 280 | 319 | 93 |
| Jatropha curcas (Physic nut | Dicot Plant | 61 | 97 | 115 | 19 |
| Cucumis sativus (Cucumber) | Dicot Plant | 88 | 102 | 134 | 27 |
| Solanum lycopersicum (Tomato) | Dicot Plant | 81 | 101 | 140 | 27 |
| Vitis Vinifers (Grape) | Dicot Plant | 59 | 71 | 138 | 19 |
| Gossypium hirsutum (Cotton) | Dicot Plant | 238 | 306 | 441 | 59 |
| Glycine max (Soybean) | Dicot Plant | 296 | 269 | 430 | 99 |
| Sesamum indicum (Sesame) | Dicot Plant | 88 | 105 | 168 | 43 |
| Camelina sativa (Flax) | Dicot Plant | 224 | 350 | 402 | 93 |
| Brassica napus (Rapeseed) | Dicot Plant | 285 | 411 | 489 | 57 |
| Chlamydomonas reinhardtii | Single-celled green alga | 2 | 0 | 16 | 12 |
| Gonium pectoral | Single-celled green alga | 1 | 0 | 12 | 3 |
| Ostreococcus lucimarinus | Single-celled green alga | 2 | 0 | 11 | 3 |
| Selaginella moellendorffii | Moss | 19 | 22 | 24 | 10 |
| Physcomitrella patens | Moss | 117 | 142 | 180 | 44 |
a The data was summarized from plant transcription factor database, 2020.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Javed, T.; Shabbir, R.; Ali, A.; Afzal, I.; Zaheer, U.; Gao, S.-J. Transcription Factors in Plant Stress Responses: Challenges and Potential for Sugarcane Improvement. Plants 2020, 9, 491. https://doi.org/10.3390/plants9040491
AMA Style
Javed T, Shabbir R, Ali A, Afzal I, Zaheer U, Gao S-J. Transcription Factors in Plant Stress Responses: Challenges and Potential for Sugarcane Improvement. Plants. 2020; 9(4):491. https://doi.org/10.3390/plants9040491
Chicago/Turabian StyleJaved, Talha, Rubab Shabbir, Ahmad Ali, Irfan Afzal, Uroosa Zaheer, and San-Ji Gao. 2020. "Transcription Factors in Plant Stress Responses: Challenges and Potential for Sugarcane Improvement" Plants 9, no. 4: 491. https://doi.org/10.3390/plants9040491
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.