Impact of Fungal Endophyte Colonization of Maize (Zea mays L.) on Induced Resistance to Thrips- and Aphid-Transmitted Viruses


Abstract
:1. Introduction
2. Results
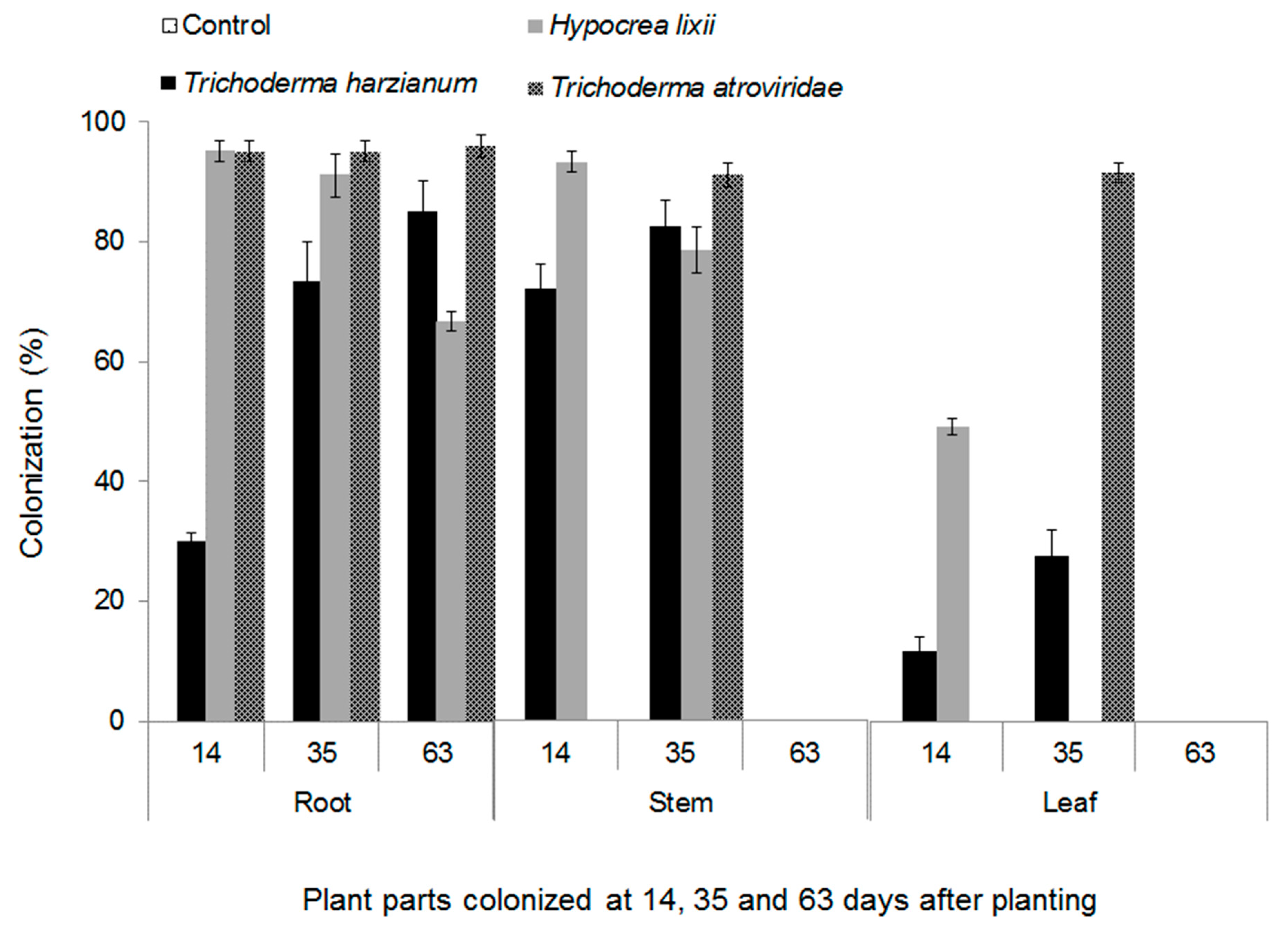
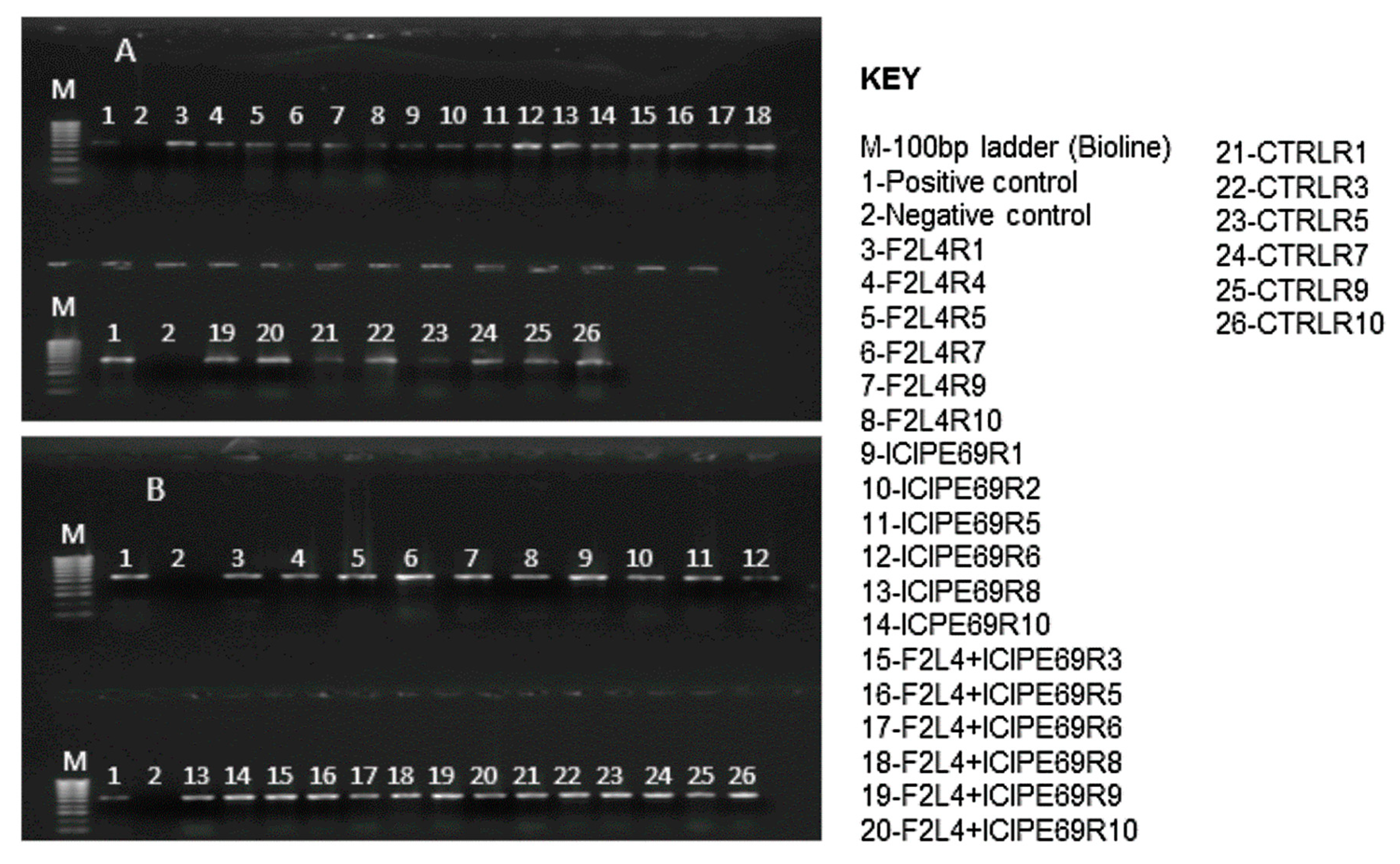
2.1. Endophytic Colonization of Maize Plants by Fungal Isolates
2.2. Effects of Fungus-Treated Maize Plants on Susceptibility to Sugarcane Mosaic Virus Disease
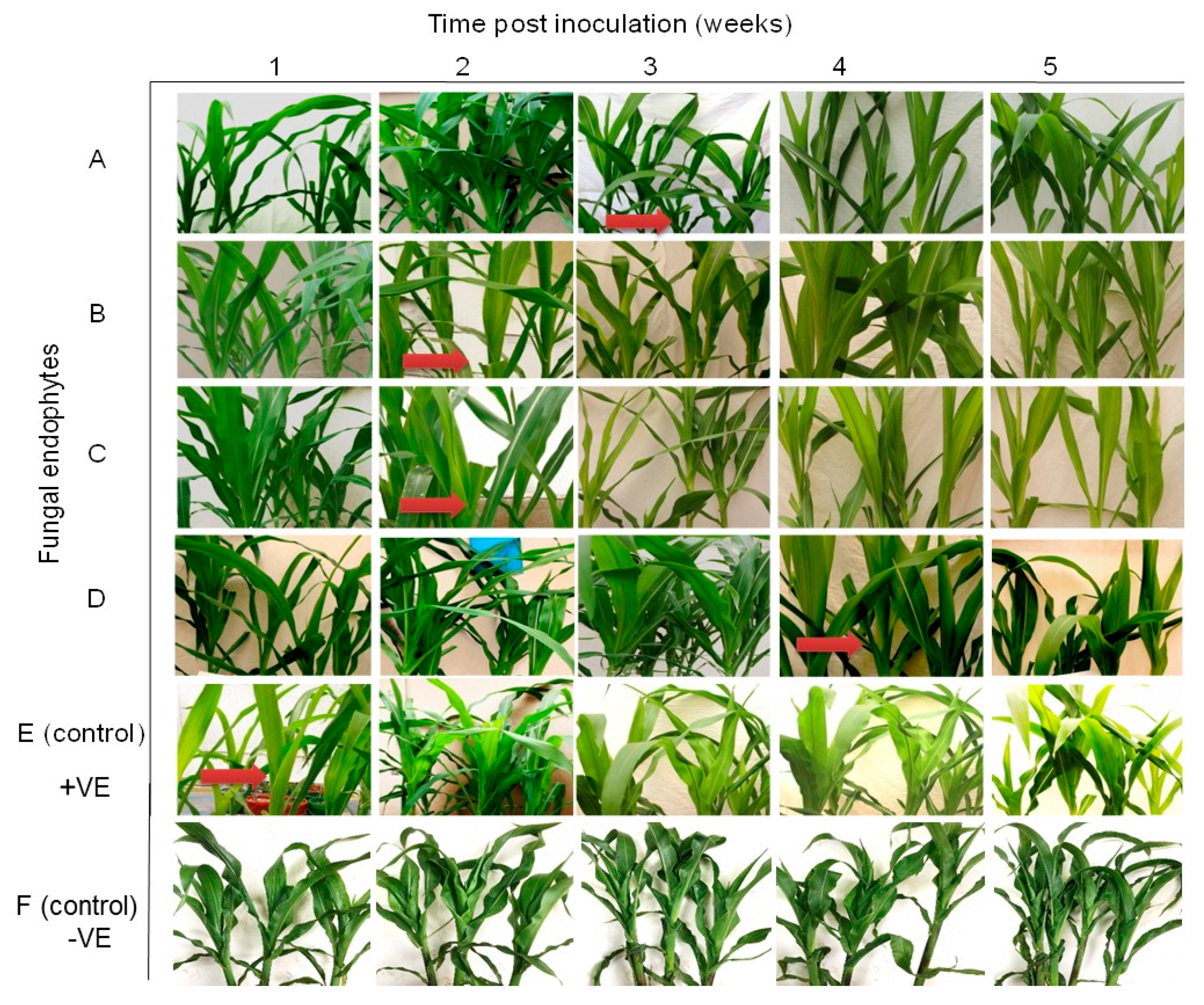
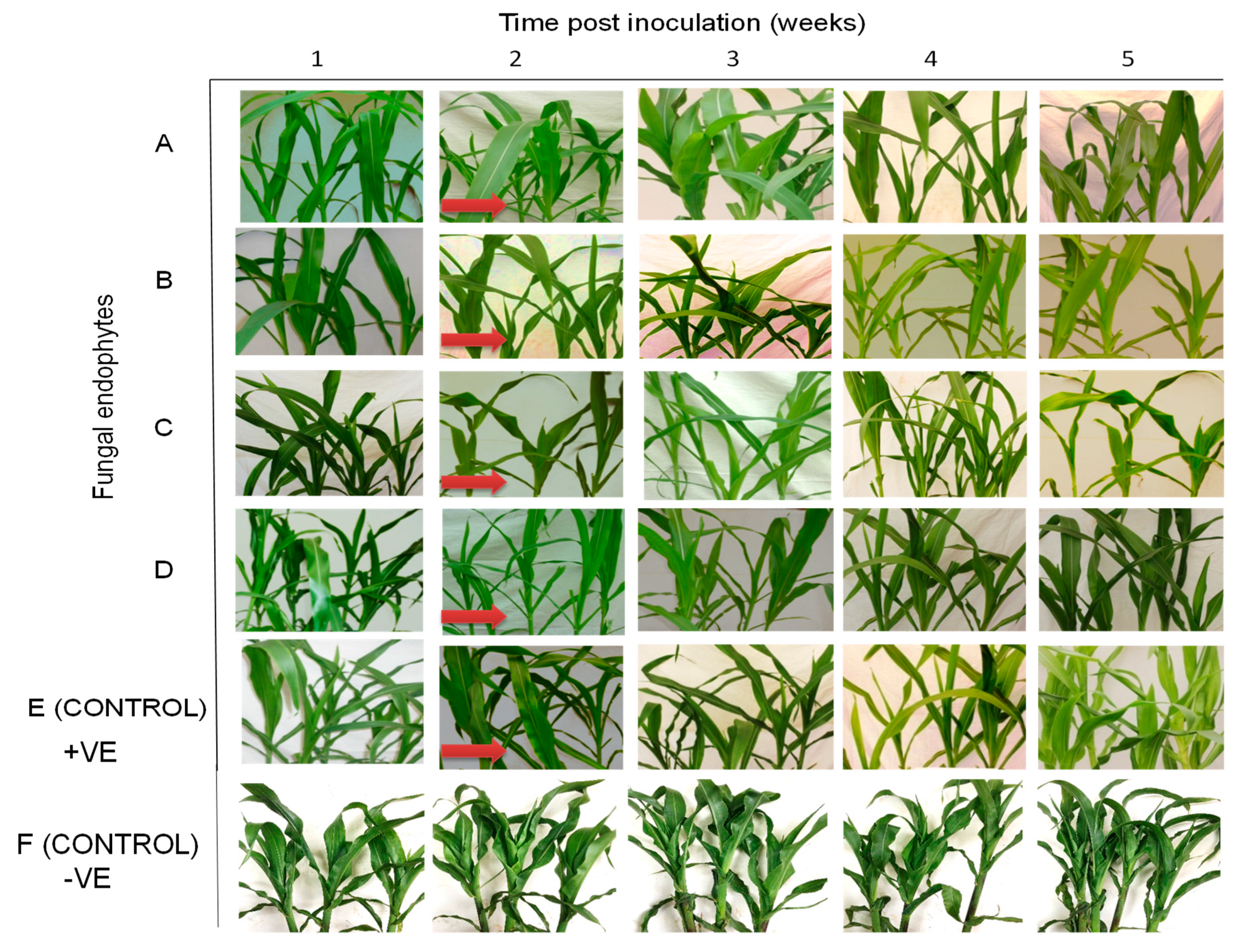
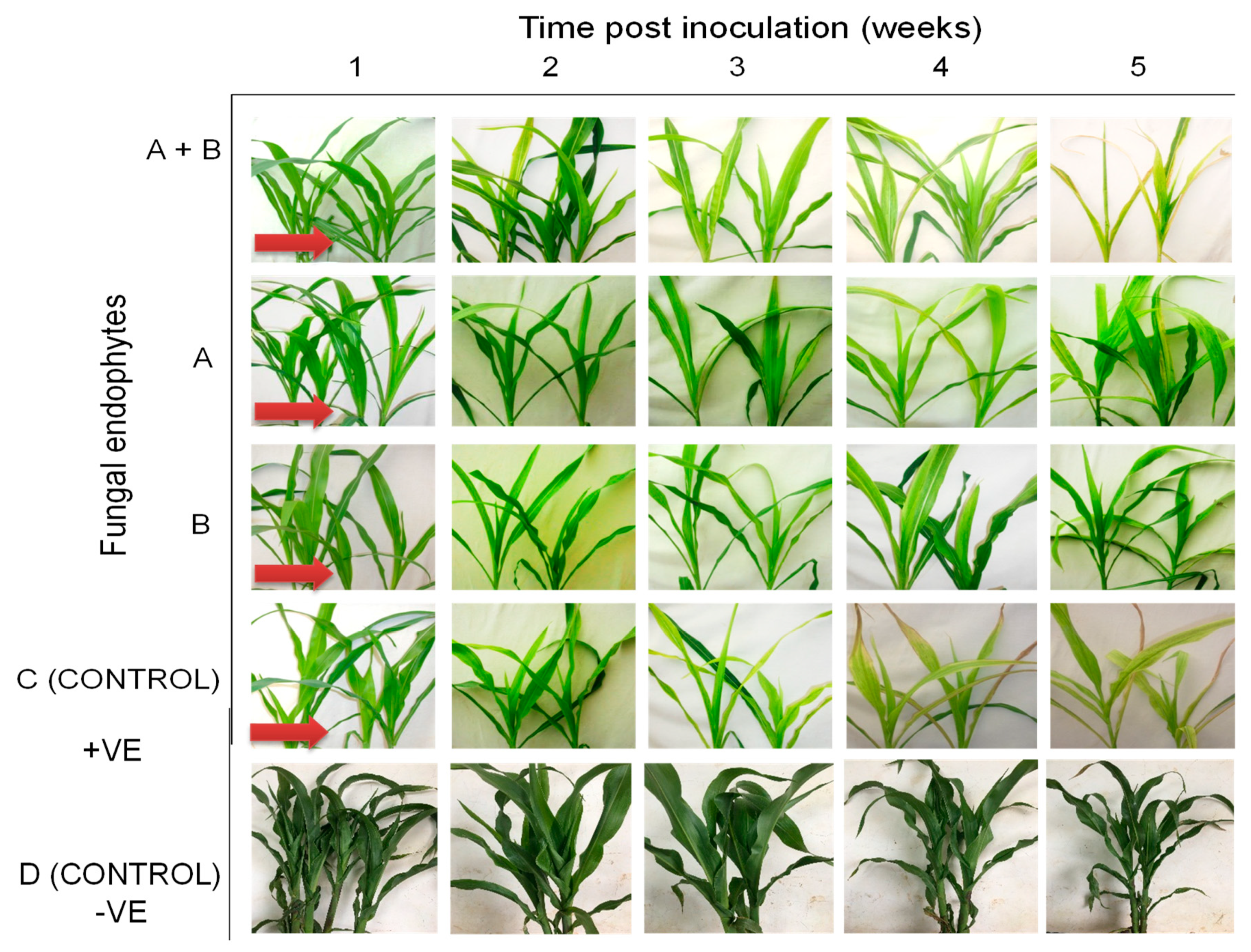
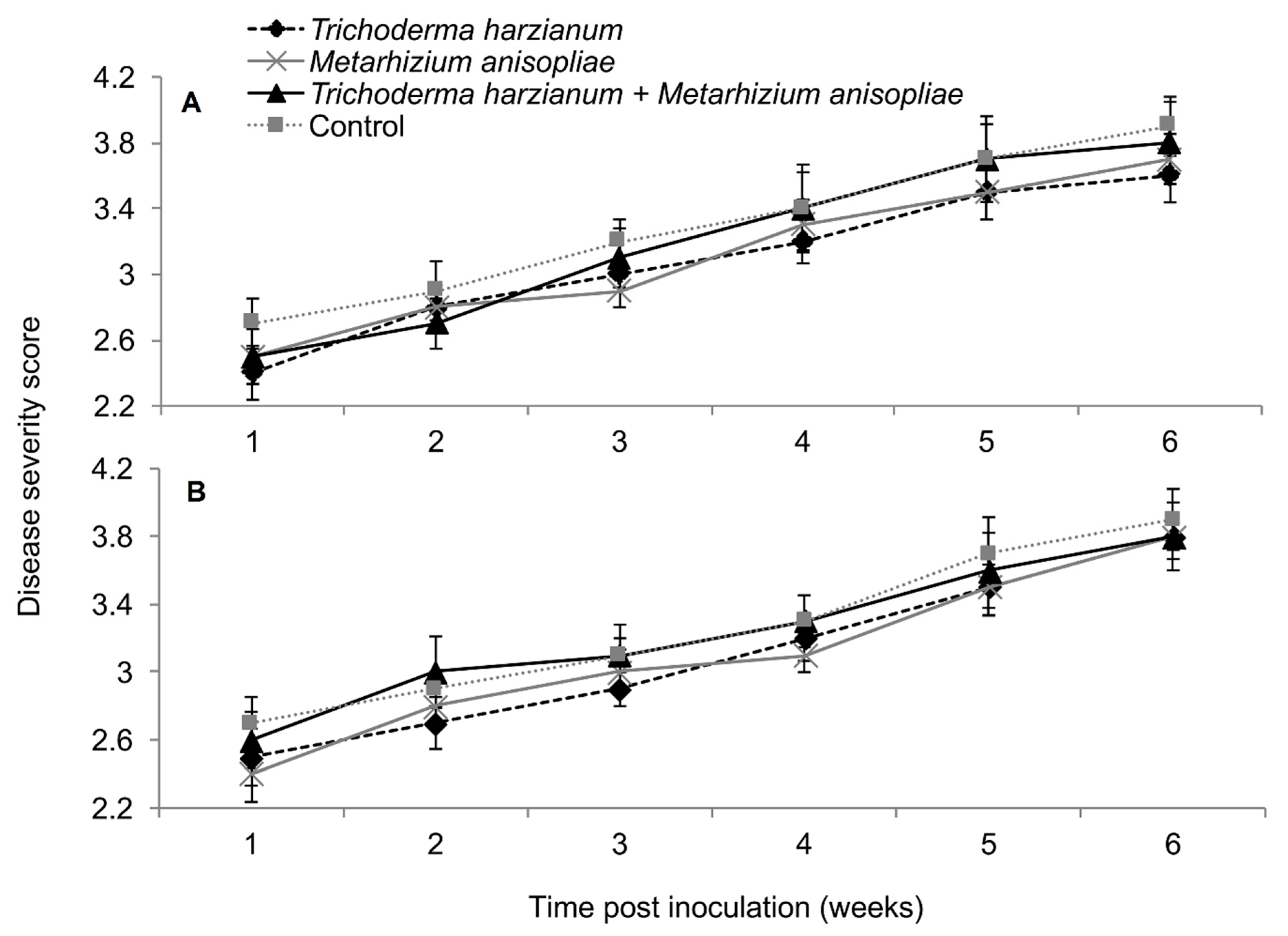
2.3. Symptom Development of Sugarcane Mosaic Virus Disease in Fungus-Treated Maize Plants
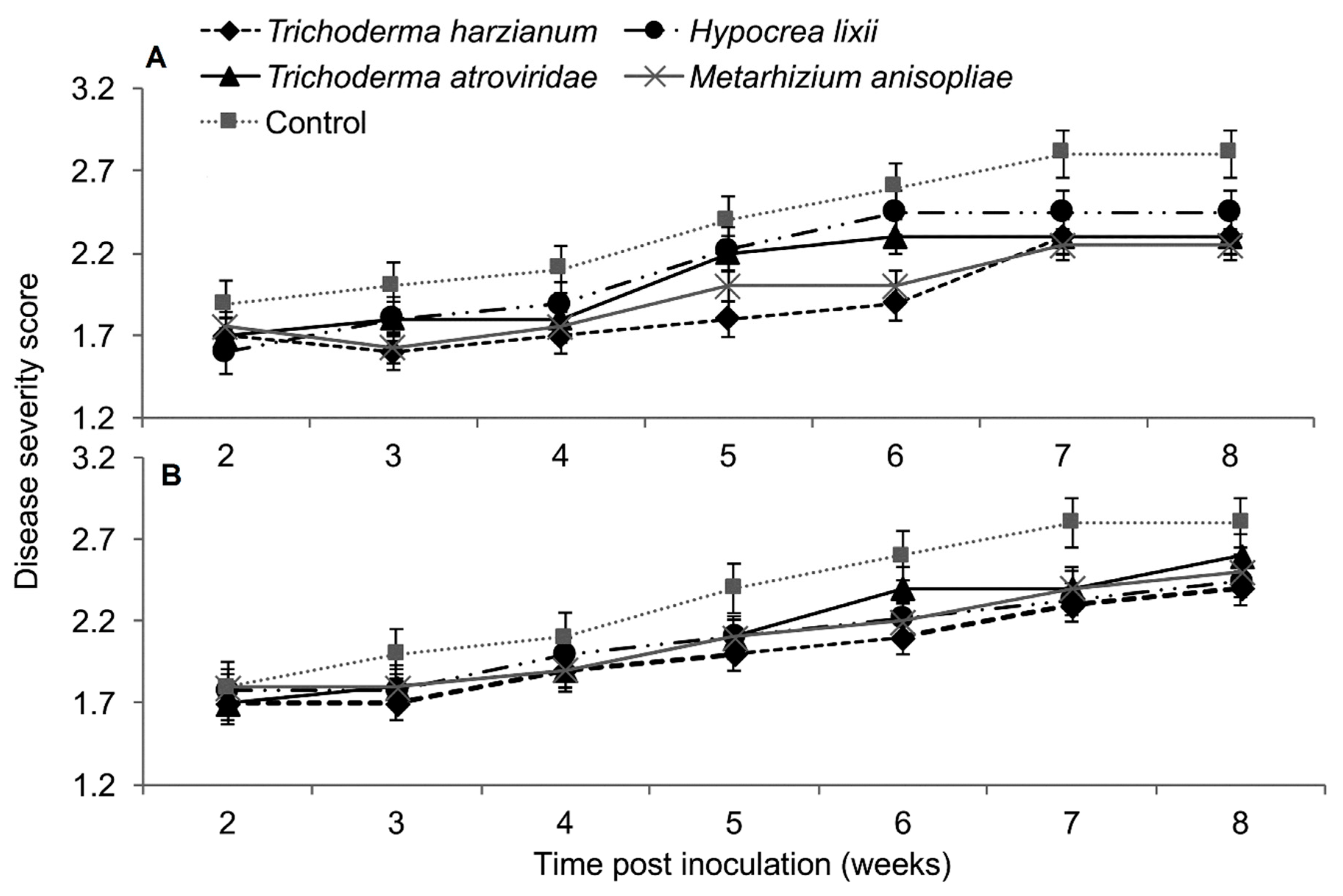
2.4. Severity of Sugarcane Mosaic Virus Disease
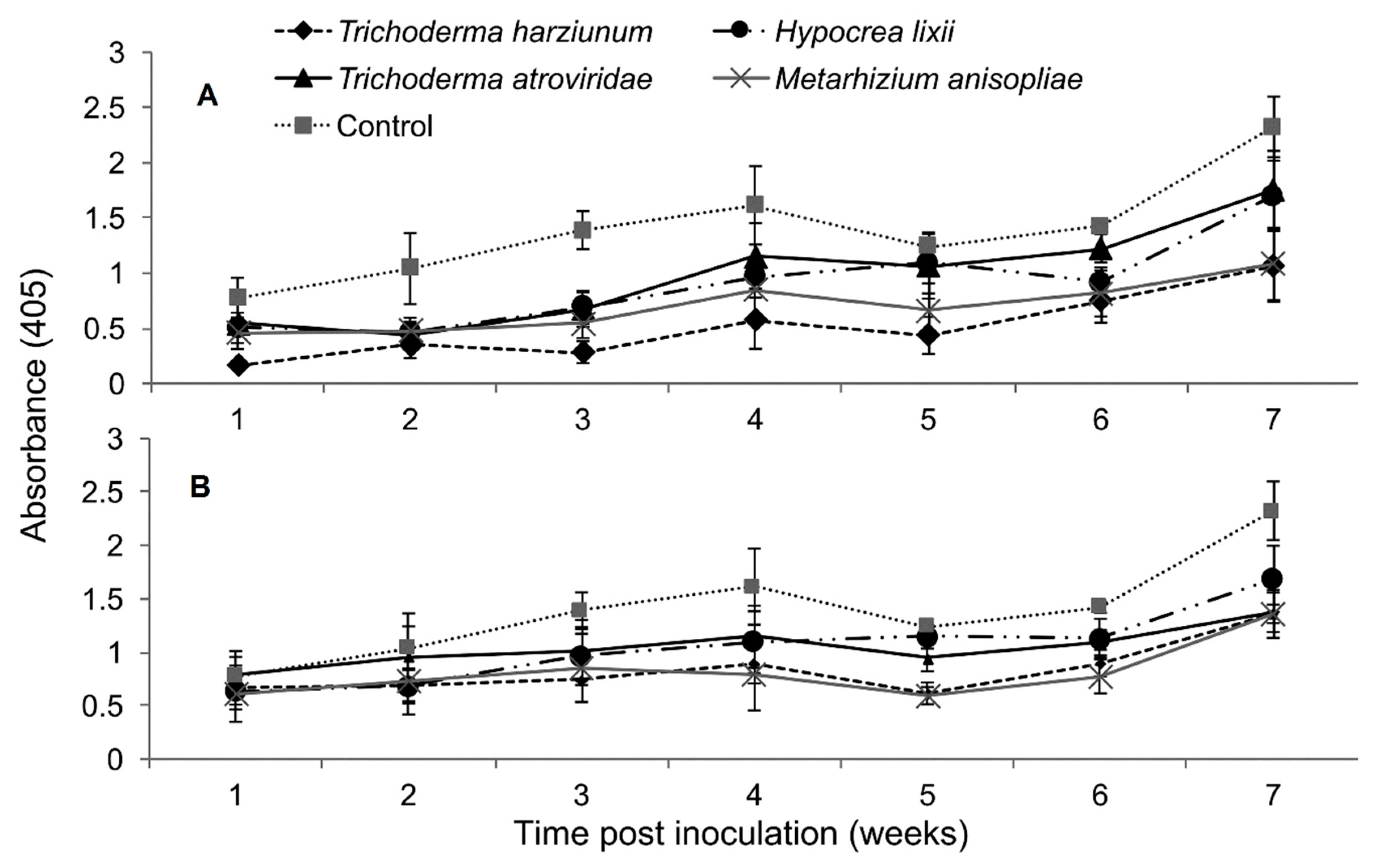
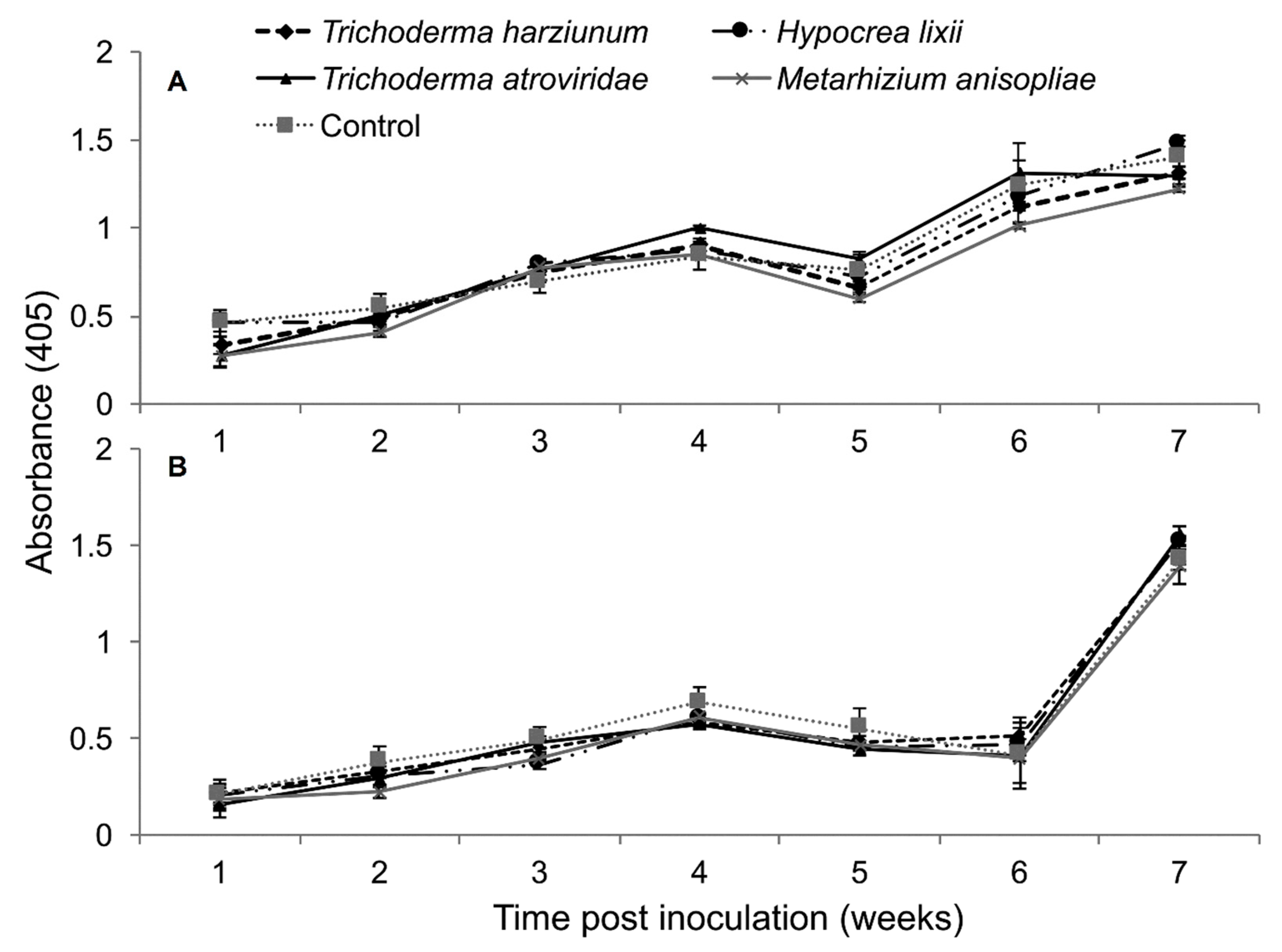
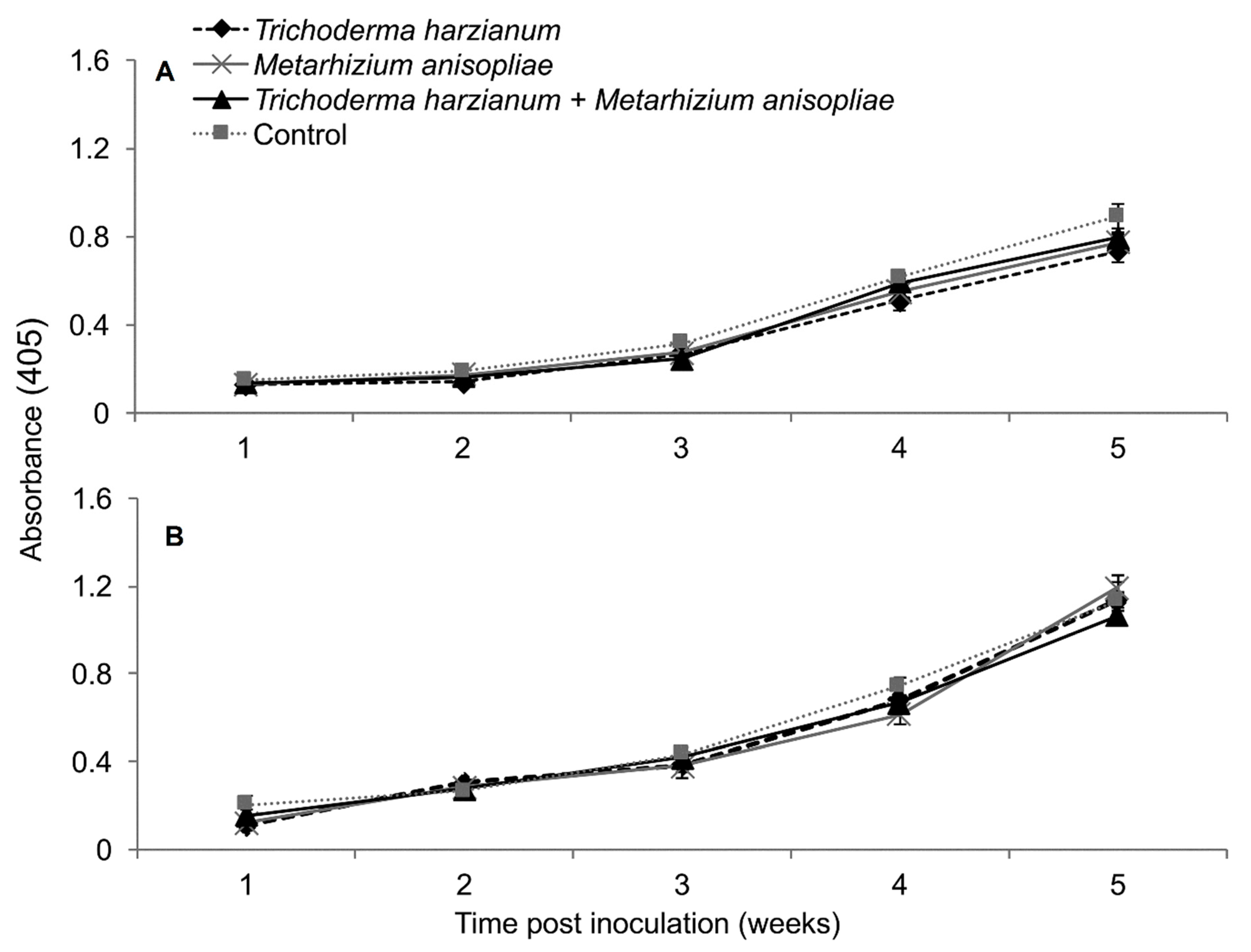
2.5. Titer Levels of Sugarcane Mosaic Virus
2.6. Effects of Fungus-Treated Maize Plants on Susceptibility to Maize Chlorotic Mottle Virus Disease
2.7. Symptom Development of Maize Chlorotic Mottle Virus Disease in Fungus-Treated Maize Plants
2.8. Severity of Maize Chlorotic Mottle Virus Disease
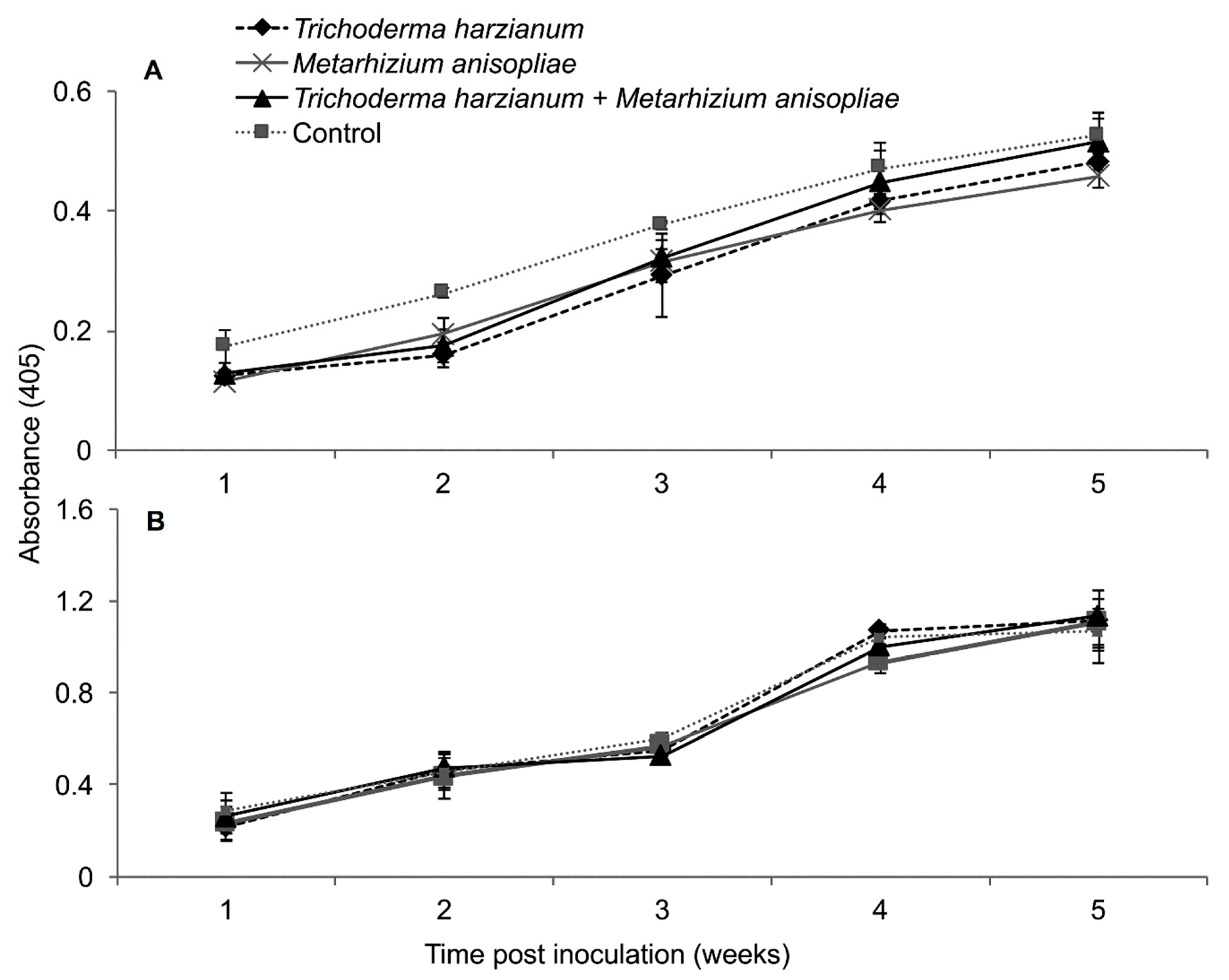
2.9. Titer Levels of Maize Chlorotic Mottle Virus
2.10. Effects of Fungus-Treated Maize Plants on Susceptibility to Maize Lethal Necrosis
2.11. Symptom Development of Maize Lethal Necrosis in Fungus-Treated Maize Plants
2.12. Severity of Maize Lethal Necrosis Disease
2.13. Titer Levels of Sugarcane Mosaic Virus and Maize Chlorotic Mottle Virus in Dually Infected Maize Plants
3. Discussion
4. Materials and Methods
4.1. Fungal Isolates
4.2. Seed Inoculation
4.3. Evaluation of Colonization by Fungal Isolates
4.4. Virus Inocula
4.5. Mechanical Inoculation with Sugarcane Mosaic Virus and Maize Chlorotic Mottle Virus on Fungus-Treated Maize Plants
4.6. Mechanical Inoculation with Dual Viruses (SCMV and MCMV) on Fungus-Treated Maize Plants
4.7. Disease Assessment
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kibaara, B.W. Technical Efficiency in Kenya’s Maize Production: An Application of the Stochastic Frontier Approach; Colorado State University: Fort Collins, CO, USA, 2005. [Google Scholar]
- Kang’ethe, E. Situation Analysis: Improving Food Safety in the Maize Value Chain in Kenya; College of Agriculture and Veterinary Science, University of Nairobi: Nairobi, Kenya, 2011. [Google Scholar]
- Jayne, T.S.; Yamano, T.; Nyoro, J.; Awuor, T. Do farmers really benefit from high good prices? Balancing rural interests in Kenya’s maize pricing and marketing policy. Tegemeo Inst. Agric. Policy Dev. 2001, 28. [Google Scholar] [CrossRef]
- Wangai, A.W.; Redinbaugh, M.G.; Kinyua, Z.M.; Miano, D.W.; Leley, P.K.; Kasina, M.; Mahuku, G.; Scheets, K.; Jeffers, D. First report of Maize chlorotic mottle virus and maize lethal necrosis in Kenya. Plant Dis. 2012, 96, 1582. [Google Scholar] [CrossRef] [PubMed]
- CIMMYT. Maize Lethal Necrosis (MLN) Disease in Kenya and Tanzania: Facts and Actions. Available online: https://www.cimmyt.org/news/maize-lethal-necrosis-mln-disease-in-kenya-and-tanzania-facts-and-actions/ (accessed on 31 July 2013).
- Adams, I.P.; Harju, V.A.; Hodges, T.; Hany, U.; Skelton, A.; Rai, S.; Deka, M.K.; Smith, J.; Fox, A.; Uzayisenga, B.; et al. First report of maize lethal necrosis disease in Rwanda. New Dis. Rep. 2014, 29, 22. [Google Scholar] [CrossRef] [Green Version]
- Mahuku, G.; Wangai, A.W.; Sadessa, K.; Teklewold, A.; Wegary, D.; Ayalneh, D.; Adams, I.; Smith, J.; Bottomley, E.; Bryce, S.; et al. First report of Maize chlorotic mottle virus and maize lethal necrosis on maize in Ethiopia. Plant Dis. 2015, 99, 1870. [Google Scholar] [CrossRef]
- Kagoda, F.; Gidoi, R.; Isabirye, E.B. Status of Maize lethal necrosis in Eastern Uganda. Afr. J. Agric. Res. 2016, 11, 652–660. [Google Scholar] [CrossRef] [Green Version]
- Lukanda, M.; Owati, A.; Ogunsanya, P.; Valimunzigha, K.; Katsongo, K.; Ndemere, H.; Kumar, P.L. First report of Maize chlorotic mottle virus infecting maize in the Democratic Republic of Congo. Plant Dis. 2014, 98, 1448. [Google Scholar] [CrossRef]
- Mahuku, G.; Lockhart, B.E.; Wanjala, B.; Jones, M.W.; Kimunye, J.N.; Stewart, L.R.; Cassone, B.J.; Sevgan, S.; Nyasani, J.O.; Kusia, E.; et al. Maize lethal necrosis (MLN), an emerging threat to maize-based food security in Sub-Saharan Africa. Phytopathology 2015, 105, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Adams, I.P.; Miano, D.W.; Kinyua, Z.M.; Wangai, A.; Kimani, E.; Phiri, N.; Reeder, R.; Harju, V.; Glover, R.; Hany, U.; et al. Use of next-generation sequencing for the identification and characterization of Maize chlorotic mottle virus and Sugarcane mosaic virus causing maize lethal necrosis in Kenya. Plant Pathol. 2013, 62, 741–749. [Google Scholar] [CrossRef]
- Nyvall, R.F. Field Crops Diseases; Nyvall, R.F., Ed.; Amazon. Com: Seattle, WA, USA, 1999. [Google Scholar]
- Nyasani, J.O.; Kusia, E.; Subramanian, S. Thrips as pests and vectors of Maize chlorotic mottle virus in maize. In Proceedings of the Xth International Symposium on Thysanoptera and Tospoviruses, Asilomar Conference Grounds, Pacific Grove, CA, USA, 16–20 May 2015. [Google Scholar]
- Mwando, N.L.; Tamiru, A.; Nyasani, J.O.; Obonyo, M.A.O.; Caulfield, J.C.; Bruce, T.J.A.; Subramanian, S. Maize chlorotic mottle virus induces changes in host plant volatiles that attract vector thrips species. J. Chem. Ecol. 2018, 44, 681–689. [Google Scholar] [CrossRef]
- Hassan, M.; Sahi, G.M.; Wakil, W.; Imanat, Y. Aphid transmission of Sugarcane sosaic virus (SCMV). Pak. J. Agric. 2003, 40, 74–76. [Google Scholar]
- Uyemoto, J.K. Biology and control of Maize chlorotic mottle virus. Plant Dis. 1983, 67, 7–10. [Google Scholar] [CrossRef]
- Nelson, S.; Brewbaker, J.; Hu, J. Maize chlorotic mottle. In Plant Disease; University of Hawaii: Honolulu, HI, USA, 2011; Volume 79, pp. 1–6. [Google Scholar]
- Kibaki, J.J.; Francis, M. Controlling Maize Lethal Necrosis Disease via Vector Management; Bayer Crop Science: Monheim, Germany, 2013. [Google Scholar]
- Albajes, R.; Gullino, M.L.; van Lenteren, J.C.; Elad, Y. Integrated Pest and Disease Management in Greenhouse Crops; Albajes, R., Gullino, M.L., Eds.; Kluwer Academic Publishers: Berlin/Heidelberg, Germany, 2002; Volume 14. [Google Scholar] [CrossRef]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaber, L.R.; Salem, N.M. Endophytic colonisation of squash by the fungal entomopathogen, Beauveria bassiana (Ascomycota: Hypocreales) for managing Zucchini yellow mosaic virus in Cucurbits. Biocontrol. Sci. Technol. 2014, 24, 1096–1109. [Google Scholar] [CrossRef]
- Istifadah, N.; McGee, P.A. Endophytic Chaetomium globosum reduces development of tan spot in wheat caused by Pyrenophora tritici-repentis. Australas. Plant Pathol. 2006, 35, 411. [Google Scholar] [CrossRef]
- Serfling, A.; Wirsel, S.G.R.; Lind, V.; Deising, H.B. Performance of the biocontrol fungus Piriformospora Indica on wheat under greenhouse and field conditions. Phytopathology 2007, 97, 523–531. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Pan, J.J.; May, G. Endophytic Fusarium verticillioides reduces disease severity caused by Ustilago maydis on maize. FEMS Microbiol. Lett. 2009, 299, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Lehtonen, P.T.; Helander, M.; Siddiqui, S.A.; Lehto, K.; Saikkonen, K. Endophytic fungus decreases plant virus infections in meadow ryegrass (Lolium pratense). Biol. Lett. 2006, 2, 620–623. [Google Scholar] [CrossRef] [Green Version]
- Muvea, A.M.; Subramanian, S.; Maniania, N.K.; Poehling, H.M.; Ekesi, S.; Meyhöfer, R. Endophytic colonization of onions induces resistance against viruliferous thrips and virus replication. Front. Plant Sci. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Akello, J. Biodiversity of Fungal Endophytes Associated with Maize, Sorghum and Napier Grass and the Influence of Biopriming on Resistance to Leaf Mining, Stem Boring and Sap Sucking Insect Pests; University of Bonn: Bonn, Germany, 2012. [Google Scholar]
- Akutse, K.S.; Maniania, N.K.; Fiaboe, K.K.M.; Van den Berg, J.; Ekesi, S. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life-history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae). Fungal Ecol. 2013, 6, 293–301. [Google Scholar] [CrossRef]
- Wyrebek, M.; Huber, C.; Sasan, R.K.; Bidochka, M.J. Three sympatrically occurring species of Metarhizium show plant rhizosphere specificity. Microbiology 2011, 157, 2904–2911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras-cornejo, H.A.; Mac, L.; Larsen, J. Ecological functions of Trichoderma Spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2018, 92, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.D.; Huang, G.R.; Wang, Y. Seasonal and tissue age influences on endophytic fungi of Pinus tabulaeformis (Pinaceae) in the Dongling mountains, Beijing. J. Integr. Plant Biol. 2008, 50, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Gurulingappa, P.; Sword, G.A.; Murdoch, G.; McGee, P.A. Colonization of crop Plants by Fungal Entomopathogens and Their Effects on Two Insect Pests When in Planta. Biol. Control. 2010, 55, 34–41. [Google Scholar] [CrossRef]
- Muvea, A.M.; Meyhöfer, R.; Subramanian, S.; Poehling, H.M.; Ekesi, S.; Maniania, N.K. Colonization of onions by endophytic fungi and their impacts on the biology of Thrips tabaci. PLoS ONE 2014, 9, e108242. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E.; Posada, F.; Catherine Aime, M.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.W.; Petrini, O.; Marais, G.F.; Pretorius, Z.A.; Rehder, F. Occurrence of fungal endophytes in cultivars of Triticum aestivum in South Africa. Mycoscience 1995, 36, 105–111. [Google Scholar] [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef] [Green Version]
- Fraser, R.S.S. Systemic consequences of the local lesion reaction to Tobacco mosaic virus in a tobacco variety lacking the N gene for hypersensitivity. Physiol. Plant Pathol. 1979, 14, 383–394. [Google Scholar] [CrossRef]
- Morris, S.W.; Vernooij, B.; Titatarn, S.; Starrett, M.; Thomas, S.; Wiltse, C.C.; Frederiksen, R.A.; Bhandhufalck, A.; Hulbert, S.; Uknes, S. Induced resistance responses in maize. Mol. Plant Microbe Interact. 1998, 11, 643–658. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.M.; Shimizu, M.; Takahashi, H.; Ozaki, K.; Hyakumachi, M. Induction of systemic resistance against Cucumber mosaic virus in Arabidopsis thaliana by Trichoderma asperellum SKT-1. Plant Pathol. J. 2013, 29, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Marina, M.A.; Silva-Flores, M.A.; Uresti-Rivera, E.E.; Castro-Longoria, E.; Herrera-Estrella, A.; Casas-Flores, S. Colonization of Arabidopsis roots by Trichoderma atroviride promotes growth and enhances systemic disease resistance through Jasmonic acid/Ethylene and Salicylic acid pathways. Eur. J. Plant Pathol. 2011, 131, 15–26. [Google Scholar] [CrossRef]
- Loebenstein, G. Localization and induced resistance in virus-infected plants. Annu. Rev. Phytopathol. 1997, 10, 177–206. [Google Scholar] [CrossRef]
- Maurhofer, M.; Hase, C.; Meuwly, P.; Metraux, J.P.; Defago, G. Induction of systemic resistance of tobacco to Tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens Strain CHA0: Influence of the GacA gene and of pyoverdine production. Phytopathology 1994, 84, 139–146. [Google Scholar] [CrossRef]
- Mutune, B.; Ekesi, S.; Niassy, S.; Matiru, V.; Bii, C.; Maniania, N.K. Fungal endophytes as promising tools for the management of Bean Stem Maggot Ophiomyia phaseoli on beans Phaseolus vulgaris. J. Pest Sci. 2016, 89, 993–1001. [Google Scholar] [CrossRef]
- Pava-ripoll, M.; Angelini, C.; Fang, W.; Wang, S.; Posada, F.J.; Leger, R.S. The rhizosphere-competent entomopathogen Metarhizium anisopliae expresses a specific subset of genes in plant root exudate. Microbiology 2011, 157, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Guy, P.L. Barley yellow dwarf viruses in Japanese pasture grasses and lack of correlation with the presence of fungal endophytes. Plant Pathol. 1993, 42, 1–5. [Google Scholar] [CrossRef]
- Goldberg, K.; Brakke, M.K. Concentration of Maize chlorotic mottle virus increased in mixed infections with Maize dwarf mosaic virus, Strain B. Pap. Plant Pathol. 1987. Paper 177. [Google Scholar] [CrossRef]
- Goettel, M.S.; Inglis, G.D. Chapter V-3—Fungi: Hyphomycetes. Man. Tech. Insect Pathol. 1997, 213–249. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; Boom, T.V.D.; Langelüddeke, P.; Stauss, R.; Weber, E.; Witzenberger, A. A Uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Dingle, J.; Mcgee, P.A. Some endophytic fungi reduce the density of pustules of Puccinia recondita f. Sp. Tritici in wheat. Mycol. Res. 2003, 107, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Guske, S.; Dammann, U.; Boyle, C. Endophyte-host interactions II. Defining symbiosis of the endophyte-host interaction. Symbiosis 1998, 25, 213–227. [Google Scholar]
- Hull, R. Mechanical inoculation of plant viruses. Curr. Protoc. Microbiol. 2009, 16, 16B. [Google Scholar] [CrossRef] [PubMed]
- Karanja, J.; Derera, J.; Gubba, A.; Mugo, S.; Wangai, A. Response of slected maize inbred germplasm to Maize lethal necrosis disease and its causative viruses (Sugarcane mosaic virus and Maize chlorotic mottle virus) in Kenya. Open Agric. J. 2018, 12, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of Enzyme-Linked Immunosorbent Assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Kiarie, S.; Nyasani, J.O.; Gohole, L.; Maniania, N.; Subramanian, S. Prospects of fungus-based biopesticides for management of insect vectors of Maize lethal necrosis disease. Afr. J. Educ. Sci. Technol. 2019, 5, 10–23. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Isolate | Source | Origin | Year of Isolation |
|---|---|---|---|---|
| Beauveria bassiana | ICIPE 35 | Coffee berry | Kenya | 2009 |
| ICIPE 273 | Coleopteran larvae | Kericho, Kenya | 2004 | |
| ICIPE 279 | soil | Mbita, Kenya | 2005 | |
| G1LU3 | Monocots | Kenya | 2012 | |
| Metarhizium anisopliae | ICIPE 20 | soil | Migori, Kenya | 1989 |
| ICIPE 30 | Coleopteran larvae | Kendu Bay, Kenya | 1989 | |
| ICIPE 69 | Soil | Matete, DRC | 1990 | |
| Trichoderma harzianum | F2L4 | Onion | Loitoktok, Kenya | 2014 |
| Trichoderma atroviride | F5S21 | Onion | Loitoktok, Kenya | 2014 |
| Hypocrea lixii | F3ST1 | Napier grass | Kenya | 2012 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiarie, S.; Nyasani, J.O.; Gohole, L.S.; Maniania, N.K.; Subramanian, S. Impact of Fungal Endophyte Colonization of Maize (Zea mays L.) on Induced Resistance to Thrips- and Aphid-Transmitted Viruses. Plants 2020, 9, 416. https://doi.org/10.3390/plants9040416
Kiarie S, Nyasani JO, Gohole LS, Maniania NK, Subramanian S. Impact of Fungal Endophyte Colonization of Maize (Zea mays L.) on Induced Resistance to Thrips- and Aphid-Transmitted Viruses. Plants. 2020; 9(4):416. https://doi.org/10.3390/plants9040416
Chicago/Turabian StyleKiarie, Simon, Johnson O. Nyasani, Linnet S. Gohole, Nguya K. Maniania, and Sevgan Subramanian. 2020. "Impact of Fungal Endophyte Colonization of Maize (Zea mays L.) on Induced Resistance to Thrips- and Aphid-Transmitted Viruses" Plants 9, no. 4: 416. https://doi.org/10.3390/plants9040416