Molecular Characterization and Identification of Calnexin 1 As a Radiation Biomarker from Tradescantia BNL4430

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
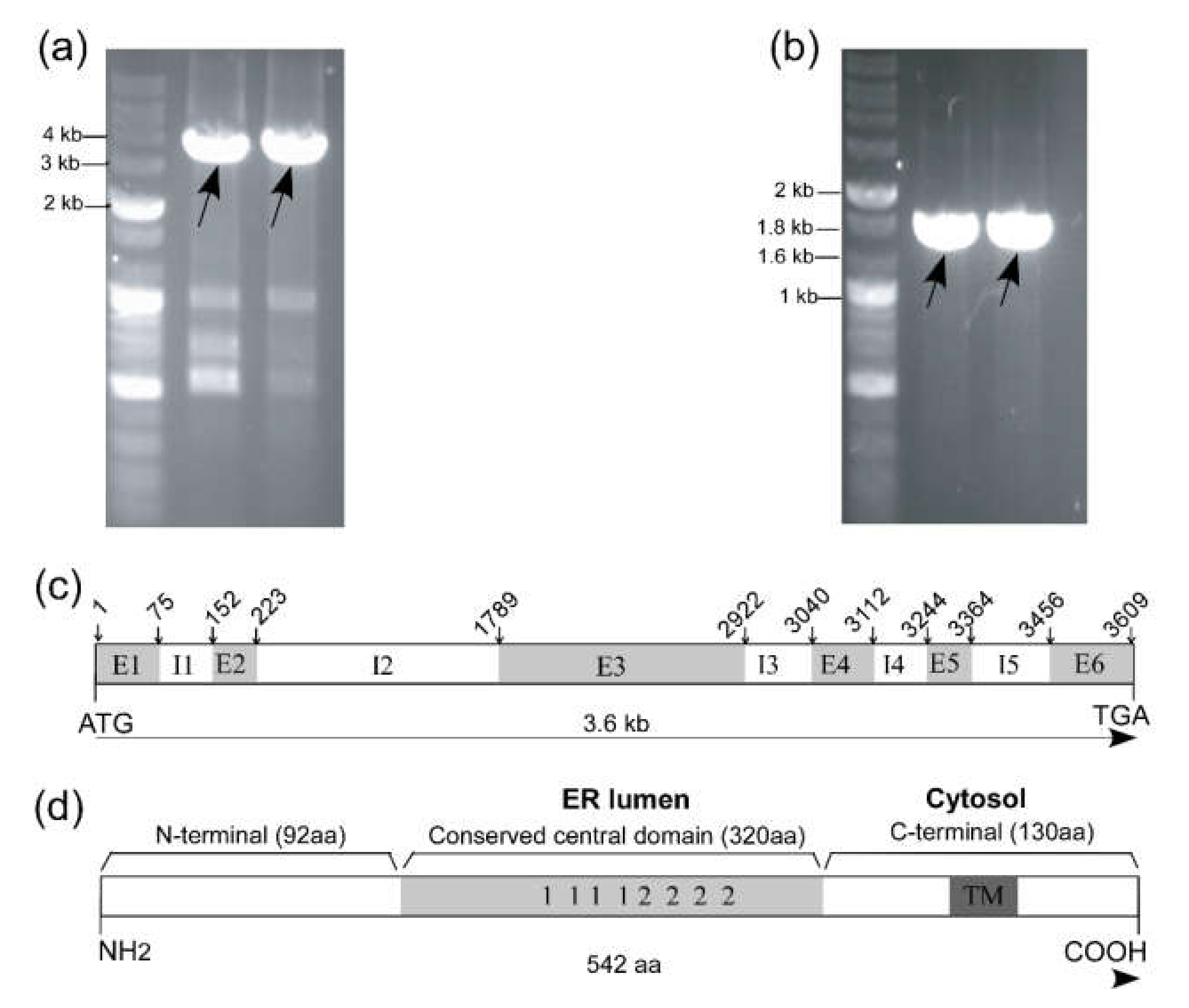
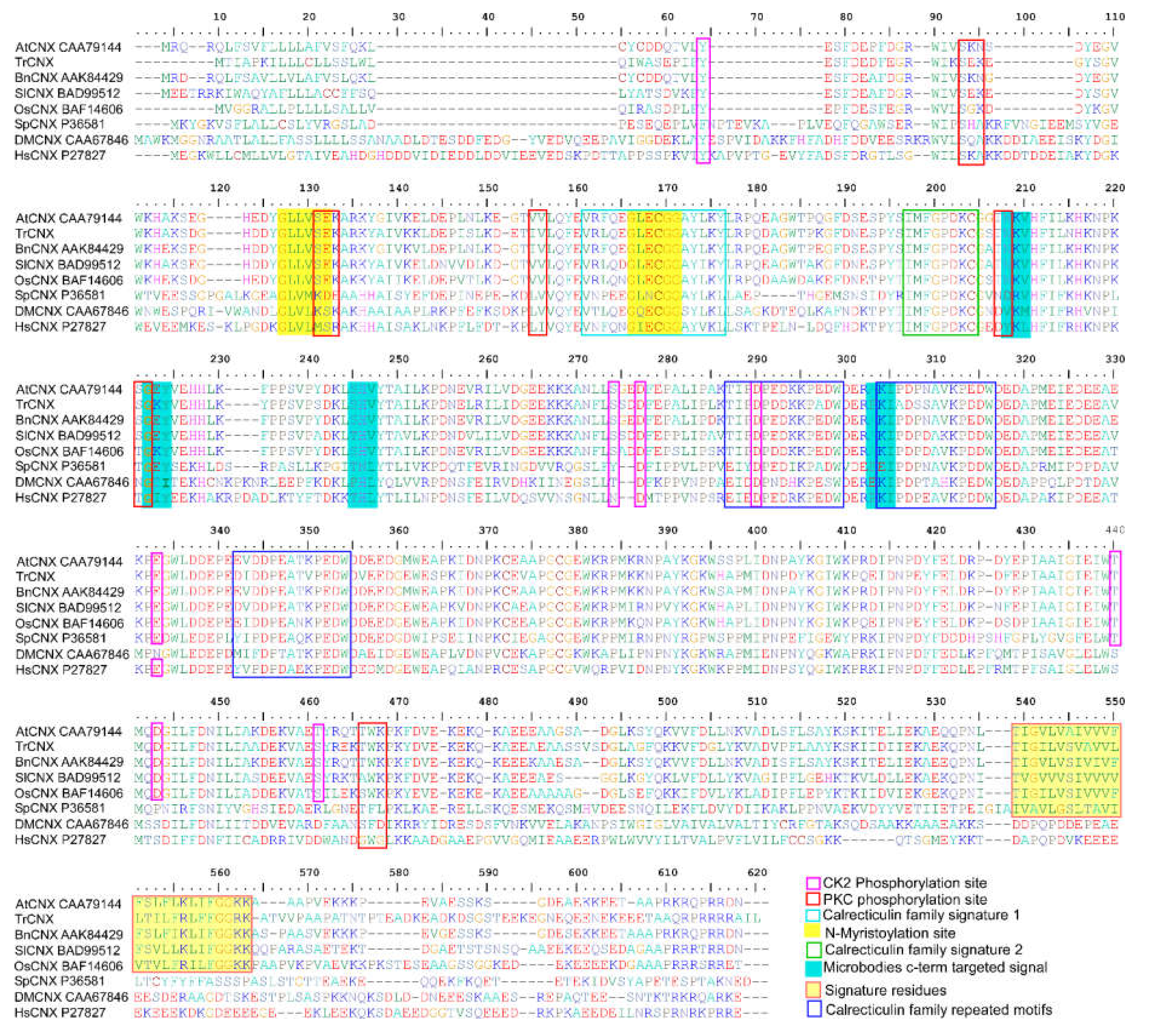
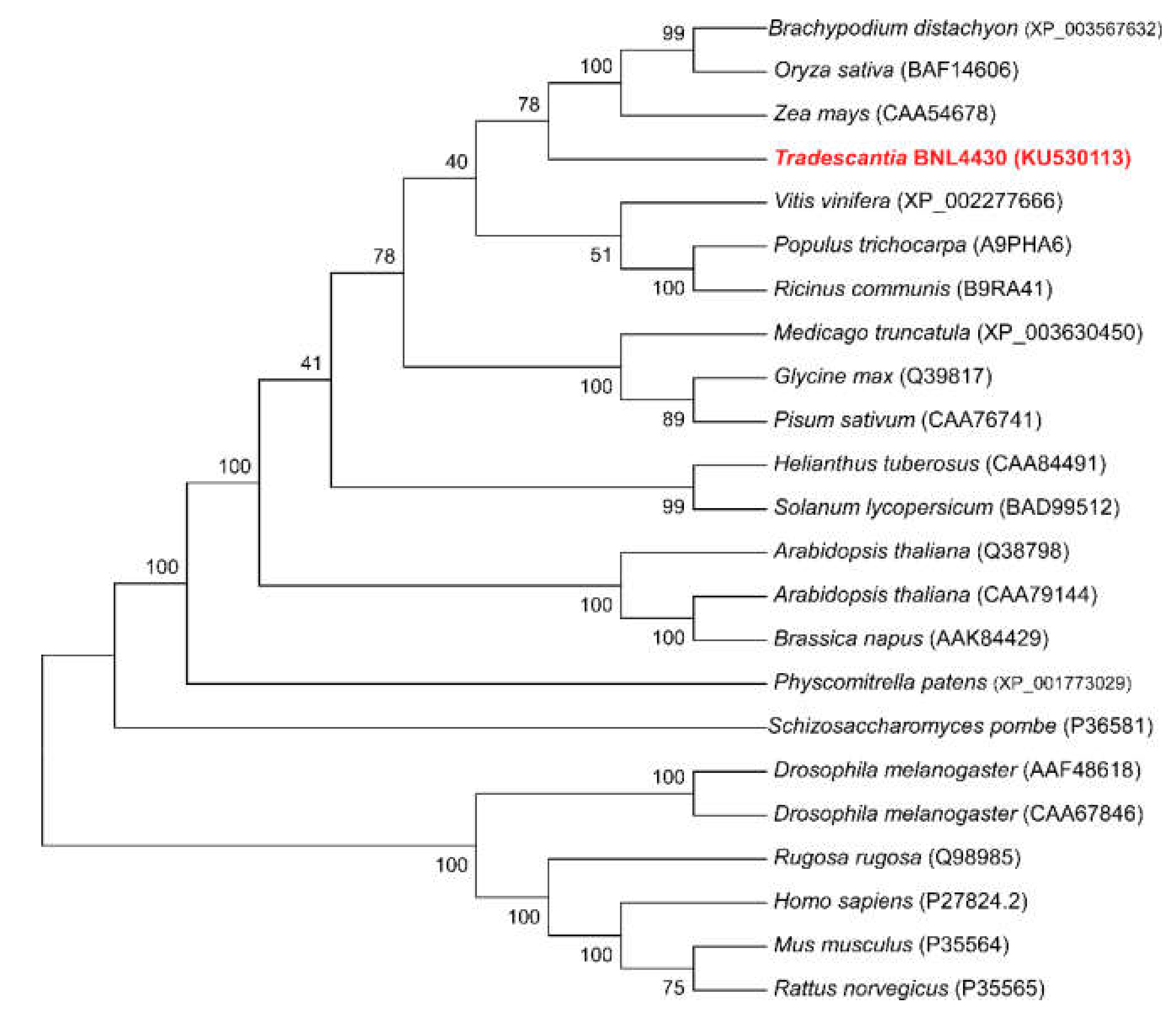
2.1. Identification and In Silico Analysis of Putative CNX1 Gene from Tradescantia
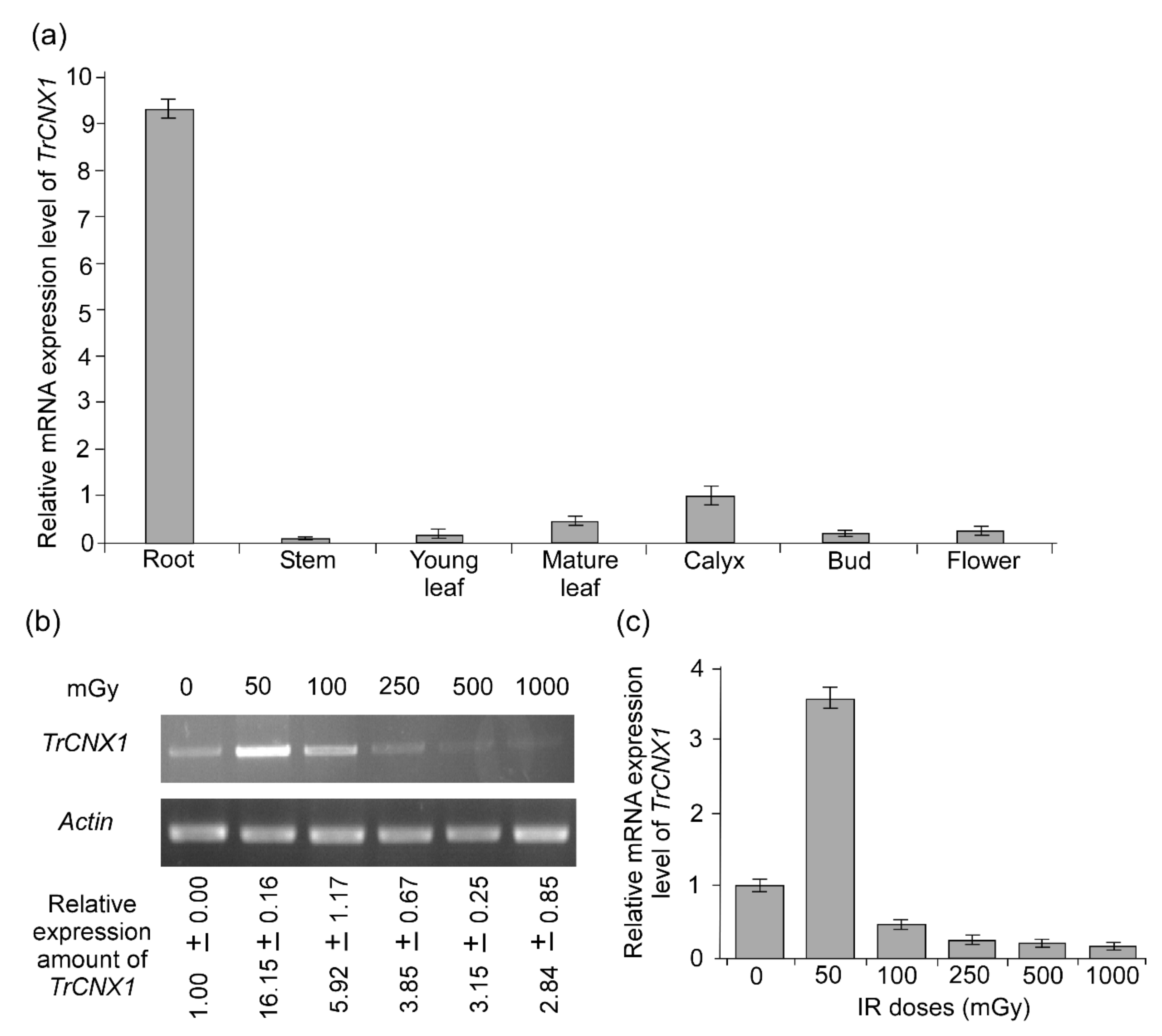
2.2. Differential Expression of TrCNX1 in Various Organs of Tradescantia
2.3. Accumulation of TrCNX1 Transcript in Tradescantia Flowers upon Ɣ-IR Stress
2.4. Expression and Analysis of TrCNX1 Fusion Protein in Recombinant E. coli
2.5. Overexpressed TrCNX1 in E. coli Enhanced Resistance to Irradiation Stresses
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Radiation Treatment
4.2. Transcriptome Resource, in Silico Identification, and Sequence Analysis
4.3. mRNA Isolation, c-DNA Cloning, RT-PCR, and RT-qPCR Analysis
4.4. Cloning of TrCNX into the pET28(a) Expression Vector and Recombinant Protein Expression
4.5. Spot Culture Assay to Analyze the Effects of UV-B and Ɣ-Radiation Stress
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jordan, B.R. The effects of ultraviolet-B radiation on plants: A molecular perspective. Adv. Bot. Res. 1996, 22, 97–161. [Google Scholar]
- Vardhan, P.V.; Shukla, L.I. Gamma irradiation of medicinally important plants and the enhancement of secondary metabolite production. Int. J. Radiat. Biol. 2017, 93, 967–979. [Google Scholar] [CrossRef]
- Esnault, M.A.; Legue, F.; Chenal, C. Ionizing radiation: Advances in plant response. Environ. Exp. Bot. 2010, 68, 231–237. [Google Scholar] [CrossRef]
- Ali, H.; Ghori, Z.; Sheikh, S.; Gul, A. Effects of Gamma Radiation on Crop Production. In Crop Production and Global Environmental Issues; Hakeem, K., Ed.; Springer: Cham, Switzerland, 2016; pp. 27–78. [Google Scholar]
- Kovalchuk, O.; Arkhipov, A.; Barylyak, I.; Karachov, I.; Titov, V.; Hohn, B.; Kovalchuk, I. Plants experiencing chronic internal exposure to ionizing radiation exhibit higher frequency of homologous recombination than acutely irradiated plants. Mutat. Res. 2000, 449, 47–56. [Google Scholar] [CrossRef]
- Majeed, A.; Khan, A.R.; Ahmad, H.; Muhammad, Z. Gamma irradiation effects on some growth parameters of Lepidium sativum L. ARPN J. Agric. Biol. Sci. 2009, 5, 39–42. [Google Scholar]
- Maity, J.P.; Kar, S.; Banerjee, S.; Chakraborty, A.; Santra, S.C. Effects of gamma irradiation on long-storage seeds of Oryza sativa (cv. 2233) and their surface infecting fungal diversity. Radiat. Phys. Chem. 2009, 78, 1006–1010. [Google Scholar] [CrossRef]
- Bala, M.; Singh, K.P. In vitro mutagenesis of rose (Rosa hybrida L.) explants using gamma-radiation to induce novel flower colour mutations. J. Hortic. Sci. Biotech. 2013, 88, 462–468. [Google Scholar] [CrossRef]
- Zaichkina, S.I.; Rozanova, O.M.; Aptikaeva, G.F.; Achmadieva, A.C.; Klokov, D.Y. Low Doses of Gamma-Radiation Induce Nonlinear Dose Responses in Mammalian and Plant Cells. Nonlinearity Biol. Toxicol. Med. 2004, 2, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, B.; Belinsky, S.; Leng, S.; Lin, Y.; Wilder, J.; Damiani, L. Radiation- stimulated epigenetic reprogramming of adaptive-response genes in the lung: An evolutionary gift for mounting adaptive protection against lung cancer. Dose Response 2009, 7, 104–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, G.; Shibato, J.; Imanaka, T.; Cho, K.; Kubo, A.; Kikuchi, S.; Satoh, K.; Kimura, S.; Ozawa, S.; Fukutani, S.; et al. Unraveling low-level gamma radiation--responsive changes in expression of early and late genes in leaves of rice seedlings at Iitate Village, Fukushima. J. Hered. 2014, 105, 7237–7238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Neubeck, C.; Shankaran, H.; Karin, N.J.; Kauer, P.M.; Chrisler, W.B.; Wang, X.; Robinson, R.J.; Waters, K.M.; Tilton, S.C.; Sowa, M.B. Cell type-dependent gene transcription profile in a three-dimensional human skin tissue model exposed to low doses of ionizing radiation: Implications for medical exposures. Environ. Mol. Mutagen. 2012, 53, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Datta, P.S. Gamma rradiation to improve plant vigour, grain development, and yield attributes of wheat. Radiat. Phys. Chem. 2010, 79, 139–143. [Google Scholar] [CrossRef]
- Kovalchuk, I.; Kovalchuk, O. Transgenic Plants as Sensors of Environmental Pollution Genotoxicity. Sensors 2008, 8, 1539–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.H.; Lee, Y.I.; Chung, K.H.; Oh, J.H. Mutation frequency of Tradescantia (BNL clone 4430) stamen hairs exposed to low dose of gamma ray in KAERI γ-field. J. Korean Nucl. Soc. 1981, 13, 162–167. [Google Scholar]
- Hwang, J.E.; Hwang, S.G.; Kim, S.H.; Lee, K.J.; Jang, C.S.; Kim, J.B.; Kim, S.H.; Ha, B.K.; Ahn, J.W.; Kang, S.Y.; et al. Transcriptome profiling in response to different types of ionizing radiation and identification of multiple radio marker genes in rice. Physiol. Plant. 2013, 150, 604–619. [Google Scholar] [CrossRef]
- Huang, L.; Franklin, A.E.; Hoffman, N.E. Primary structure and characterization of an Arabidopsis thaliana calnexin-like protein. J. Biol. Chem. 1993, 268, 6560–6566. [Google Scholar]
- Parlati, F.; Dominguez, M.; Bergeron, J.J.; Thomas, D.Y. Saccharomyces cerevisiae CNE1 encodes an endoplasmic reticulum (ER) membrane protein with sequence similarity to calnexin and calreticulin and functions as a constituent of the ER quality control apparatus. J. Biol. Chem. 1995, 270, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowski, B.A.; Zielinkska-Kwiatkowska, A.G.; Migdalski, A.; Kleczkowski, L.A.; Wasilweska, L.D. Cloning of two cDNAs encoding calnexin-like proteins from maize (Zea mays) leaves; identification of potential calcium-binding domains. Gene 1995, 65, 219–222. [Google Scholar] [CrossRef]
- Goode, J.H.; Settlage, S.B.; Wilson, R.F.; Dewey, R.E. Isolation of a calnexin (GenBank U20502) homolog from developing soybean seeds (PGR95–005). Plant Physiol. 1995, 108, 1341–1343. [Google Scholar]
- Ehtesham, N.Z.; Phan, T.N.; Gaikwad, A.; Sopory, S.K.; Tuteja, N. Calnexin from Pisum sativum: Cloning of the cDNA and characterization of the encoded protein. DNA Cell Biol. 1999, 18, 853–862. [Google Scholar] [CrossRef] [Green Version]
- Sarwat, M.; Naqvi, A.R. Heterologous expression of rice calnexin (OsCNX) confers drought tolerance in Nicotiana tabacum. Mol. Biol. Rep. 2013, 40, 5451–5464. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R.; Gaxiola, R. Microbial models and salt stress tolerance in plants. Crit. Rev. Plant Sci. 2011, 13, 121–138. [Google Scholar] [CrossRef]
- Nakahara, Y.; Sawabe, S.; Kainuma, K.; Katsuhara, M.; Shibasaka, M.; Suzuki, M.; Yamamoto, K.; Oguri, S.; Sakamoto, H. Yeast functional screen to identify genes conferring salt stress tolerance in Salicornia europaea. Plant Sci. 2015, 6, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.M.; Chen, R.J.; Zhang, J.H.; Gao, X.L.; Li, L.H.; Wang, P.R.; Deng, X.J.; Xu, Z.J. OsPOP5, a prolyl oligopeptidase family gene from rice confers abiotic stress tolerance in Escherichia coli. Int. J. Mol. Sci. 2013, 14, 20204–20219. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.F.L.; Huang, W.K.; Lin, Y.H.; Chen, Y.; Chang, T.H. Protective function of the recombinant Oshsp18.0-CII protein, a class II small heat shock protein of rice, in Escherichia coli. Bot. Stud. 2012, 53, 291–299. [Google Scholar]
- Boston, R.S.; Viitanen, P.V.; Vierling, E. Molecular chaperones and protein folding in plants. Plant Mol. Biol. 1996, 32, 191–222. [Google Scholar] [CrossRef]
- Nouri, M.Z.; Hiraga, S.; Yanagawa, Y.; Sunohara, Y.; Matsumoto, H.; Komatsu, S. Characterization of calnexin in soybean roots and hypocotyls under osmotic stress. Phytochemistry 2012, 74, 20–29. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, Y.Y.; Kim, J.K. Somatic mutations of Tradescantia 4430 treated with radiation and mercury. J. Radiat. Ind. 2007, 1, 149–153. [Google Scholar]
- Ma, N.L.; Rahmat, Z.; Lam, S.S. A Review of the “Omics” Approach to Biomarkers of Oxidative Stress in Oryza sativa. Int. J. Mol. Sci. 2013, 14, 7515–7541. [Google Scholar] [CrossRef]
- Nikitaki, Z.; Hellweg, C.E.; Georgakilas, A.G.; Ravanat, J.L. Stress-induced DNA damage biomarkers: Applications and limitations. Front Chem. 2015, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Sparrow, A.H.; Schairer, L.A.; Nawrocky, M.M.; Sautkulis, R.C. Effects of low temperature and low level chronic gamma radiation on somatic mutation rates in Tradescantia. Radiat. Res. 1971, 47, 273–274. [Google Scholar]
- Ma, T.H.; Cabrera, G.L.; Cebulska-Wasilewska, A.; Chen, R.; Loarca, F.; Vandenberg, A.L.; Salamone, M.F. Tradescantia stamen hair mutation bioassay. Mutat. Res. 1994, 310, 211–220. [Google Scholar] [CrossRef]
- Ellgaard, L.; Molinari, M.; Helenius, A. Setting the standards: Quality control in the secretory pathway. Science 1999, 286, 1882–1888. [Google Scholar] [CrossRef] [PubMed]
- Wada, I.; Rindress, D.; Cameron, P.H.; Ou, W.J.; Doherty, J.J., 2nd; Louvard, D.; Bell, A.W.; Dignard, D.; Thomas, D.Y.; Bergeron, J.J. SSR alpha and associated calnexin are major calcium binding proteins of the endoplasmic reticulum membrane. J. Biol. Chem. 1991, 266, 19599–19610. [Google Scholar] [PubMed]
- Hupp, T.R.; Meek, D.W.; Midgley, C.A.; Lane, D.P. Regulation of the specific DNA binding function of p53. Cell 1992, 71, 875–886. [Google Scholar] [CrossRef]
- Elagöz, A.; Callejo, M.; Armstrong, J.; Rokeach, L.A. Although calnexin is essential in S. pombe, its highly conserved central domain is dispensable for viability. J. Cell Sci. 1999, 112, 4449–4460. [Google Scholar]
- Danilczyk, U.G.; Williams, D.B. The lectin chaperone calnexin utilizes polypeptide-based interactions to associate with many of its substrates in vivo. J. Biol. Chem. 2001, 276, 25532–25540. [Google Scholar] [CrossRef] [Green Version]
- Tuteja, N.; Mahajan, S. Calcium Signaling Network in Plants: An Overview. Plant Signal Behav. 2007, 2, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Martínez, I.M.; Chrispeels, M.J. Genomic Analysis of the Unfolded Protein Response in Arabidopsis Shows Its Connection to Important Cellular Processes. Plant Cell 2003, 15, 561–576. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Subburaj, S.; Ha, H.J.; Park, N.; Choi, S.H.; Lee, G.J. Chalcone Isomerase-like genes in Tradescantia BNL4430: Identification, molecular characterization, and differential expression profiles under Ɣ-radiation stress. J. Plant Biochem. Biot. 2017, 26, 330–345. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, H.-J.; Subburaj, S.; Kim, Y.-S.; Kim, J.-B.; Kang, S.-Y.; Lee, G.-J. Molecular Characterization and Identification of Calnexin 1 As a Radiation Biomarker from Tradescantia BNL4430. Plants 2020, 9, 387. https://doi.org/10.3390/plants9030387
Ha H-J, Subburaj S, Kim Y-S, Kim J-B, Kang S-Y, Lee G-J. Molecular Characterization and Identification of Calnexin 1 As a Radiation Biomarker from Tradescantia BNL4430. Plants. 2020; 9(3):387. https://doi.org/10.3390/plants9030387
Chicago/Turabian StyleHa, Hye-Jeong, Saminathan Subburaj, Young-Sun Kim, Jin-Baek Kim, Si-Yong Kang, and Geung-Joo Lee. 2020. "Molecular Characterization and Identification of Calnexin 1 As a Radiation Biomarker from Tradescantia BNL4430" Plants 9, no. 3: 387. https://doi.org/10.3390/plants9030387