Effects of Endophytic Bacillus Subtilis and Salicylic Acid on Postharvest Diseases (Phytophthora infestans, Fusarium oxysporum) Development in Stored Potato Tubers

 ,
,  and
and 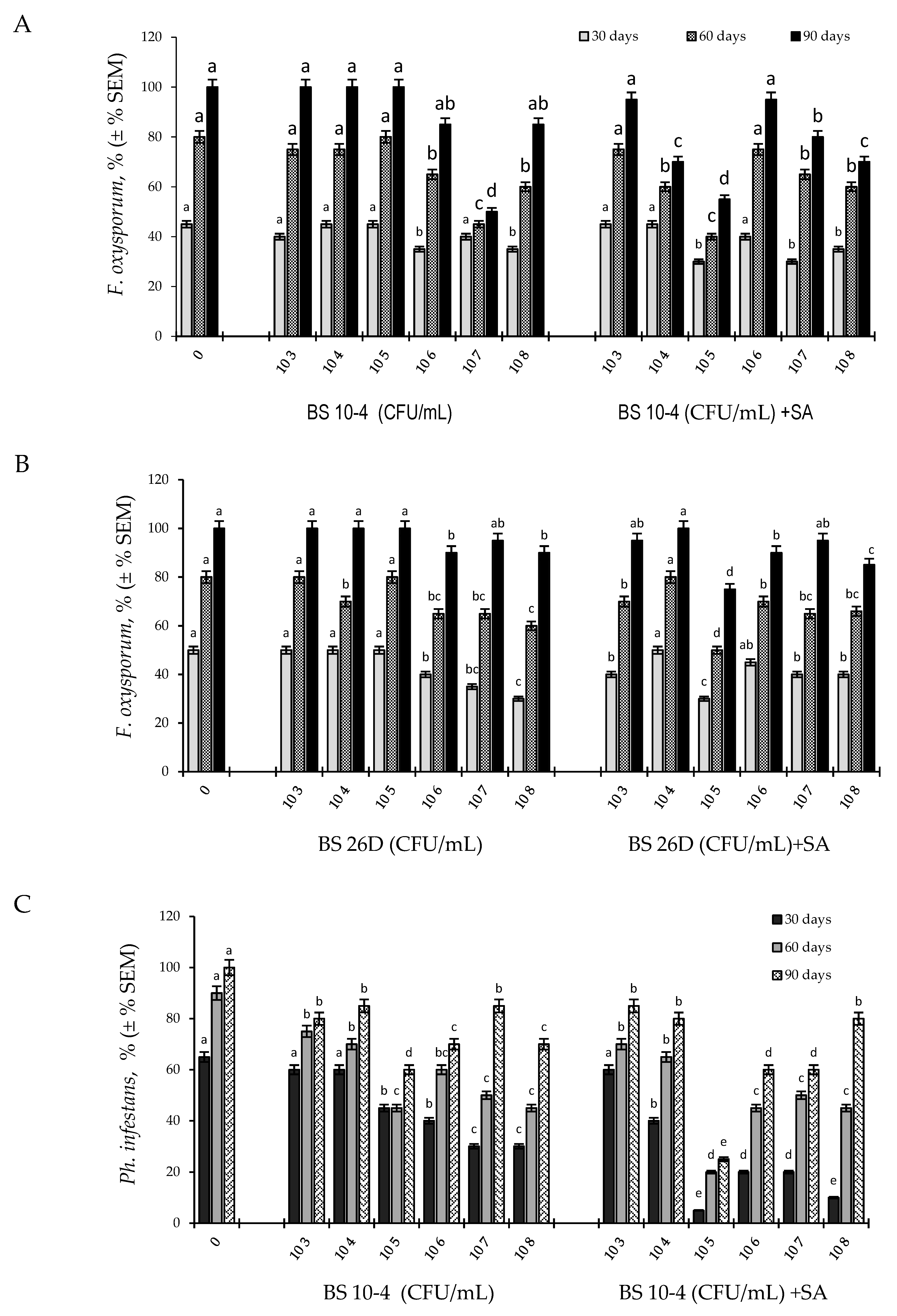{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
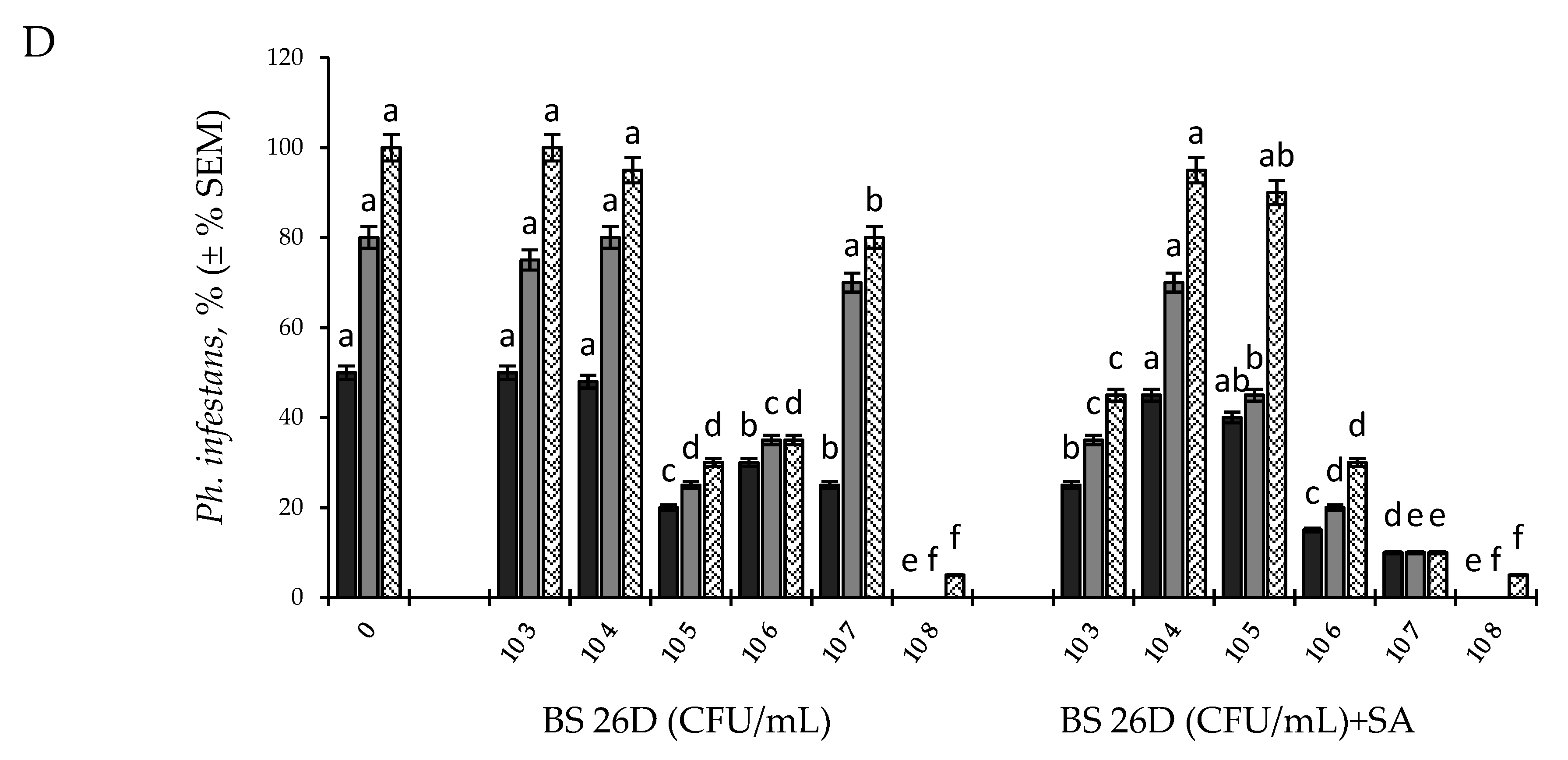
2.1. Effect of Endophytic B. subtilis (Strains 10-4, 26D) at Different Concentrations, Individually and in Compositions with SA, on the Development of Ph. infestans and F. oxysporum in Potato Tubers during Storage
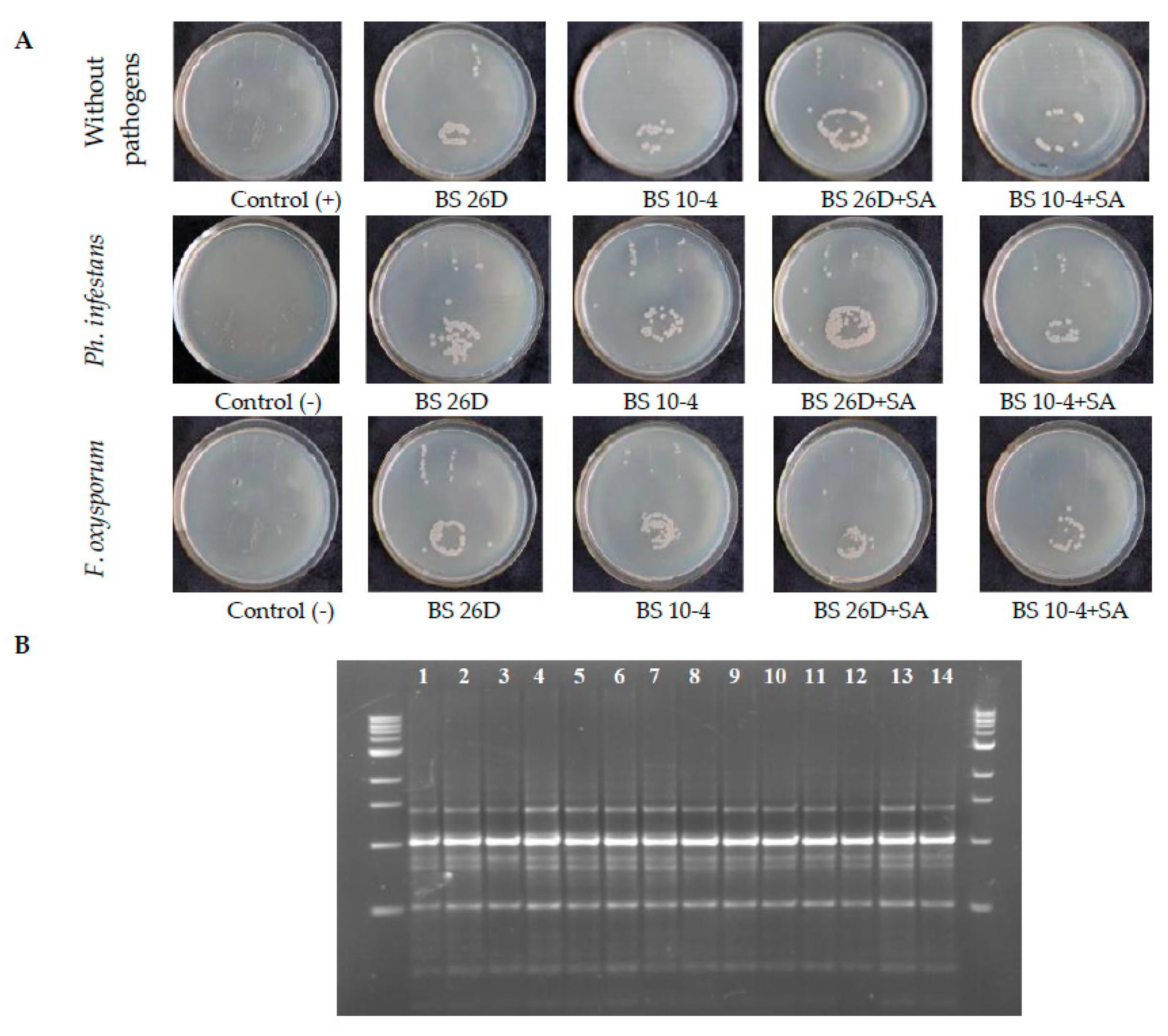
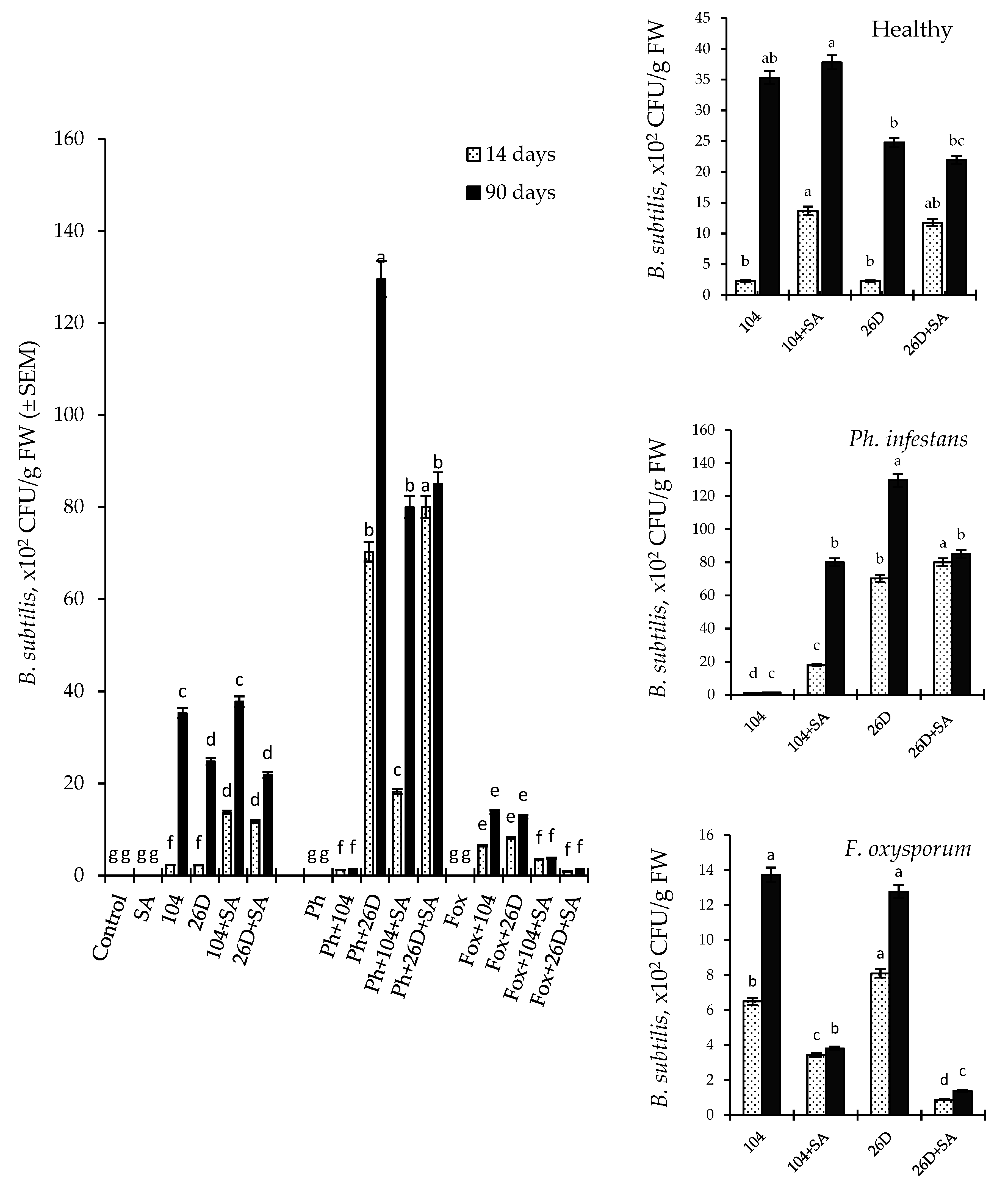
2.2. The Ability of B. subtilis (10-4 and 26D) to Colonize the Internal Tissues of Potato Tubers Treated before Storage with B. subtilis and B. subtilis + SA under Normal and Pathogens (Ph. infestans and F. oxysporum)-Infected Stored Conditions
2.3. Change in Count of B. subtilis in Healthy and Infected (by Ph. infestans and F. oxysporum) Tubers over Storage Time and under Exogenous SA Influence
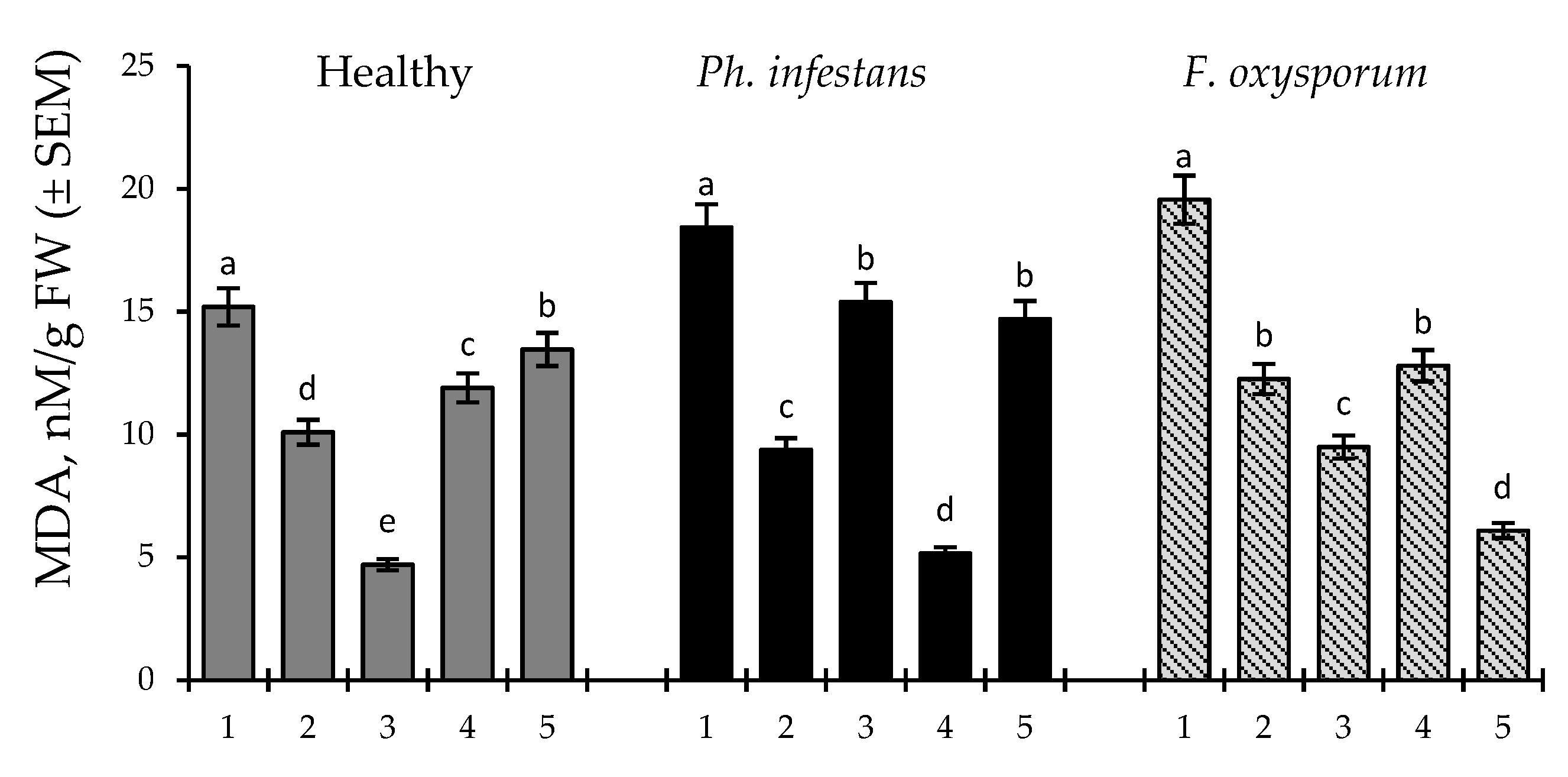
2.4. Effect of B. subtilis (10-4 and 26D), Individually and in Compositions with SA, on Lipid Peroxidation (MDA) in Stored Healthy and Pathogens (Ph. infestans and F. oxysporum)-Infected Potato Tubers
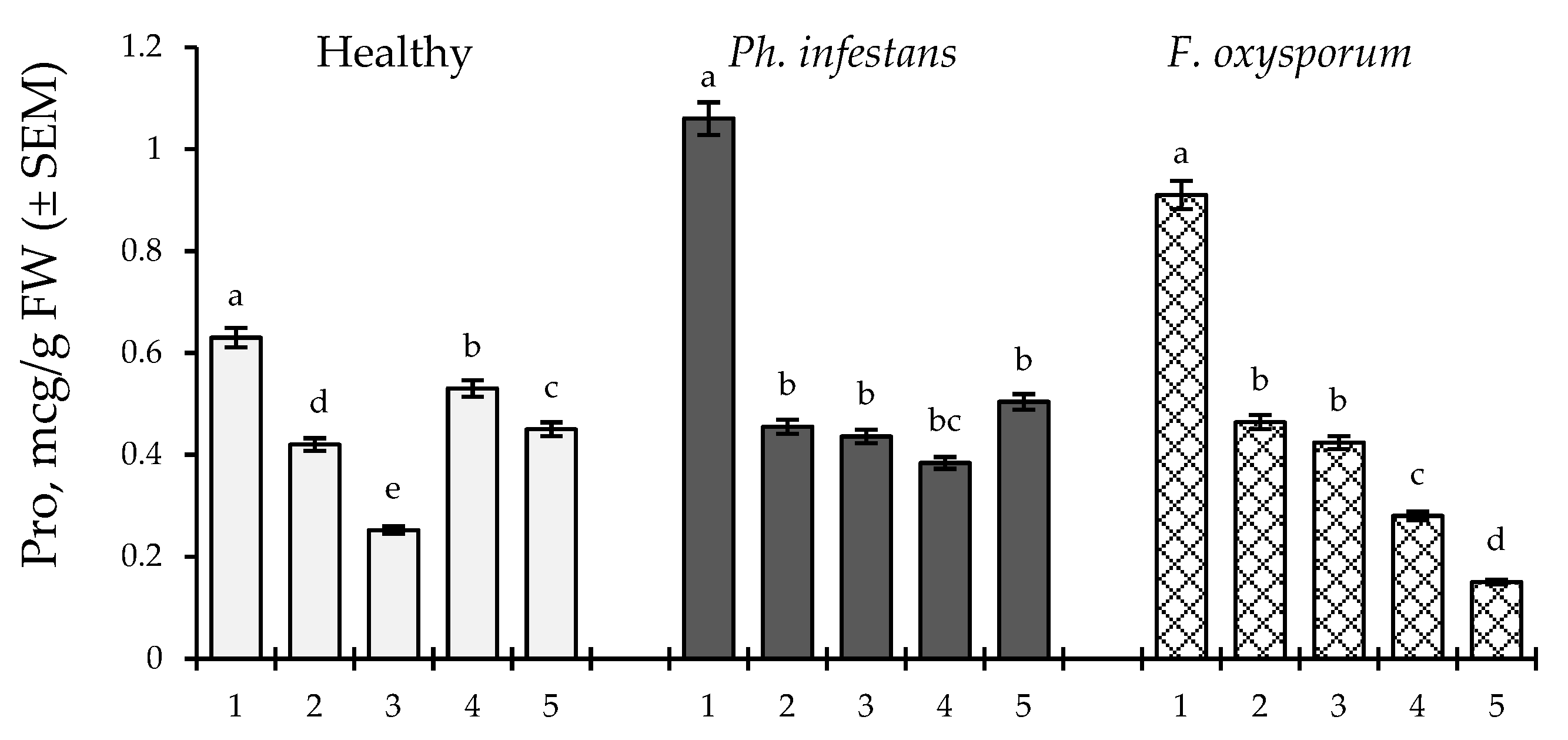
2.5. Effect of B. subtilis (10-4 and 26D) and B. subtilis + SA on Proline (Pro) Content in Healthy and Pathogens (Ph. infestans and F. oxysporum)-Infected Stored Tubers
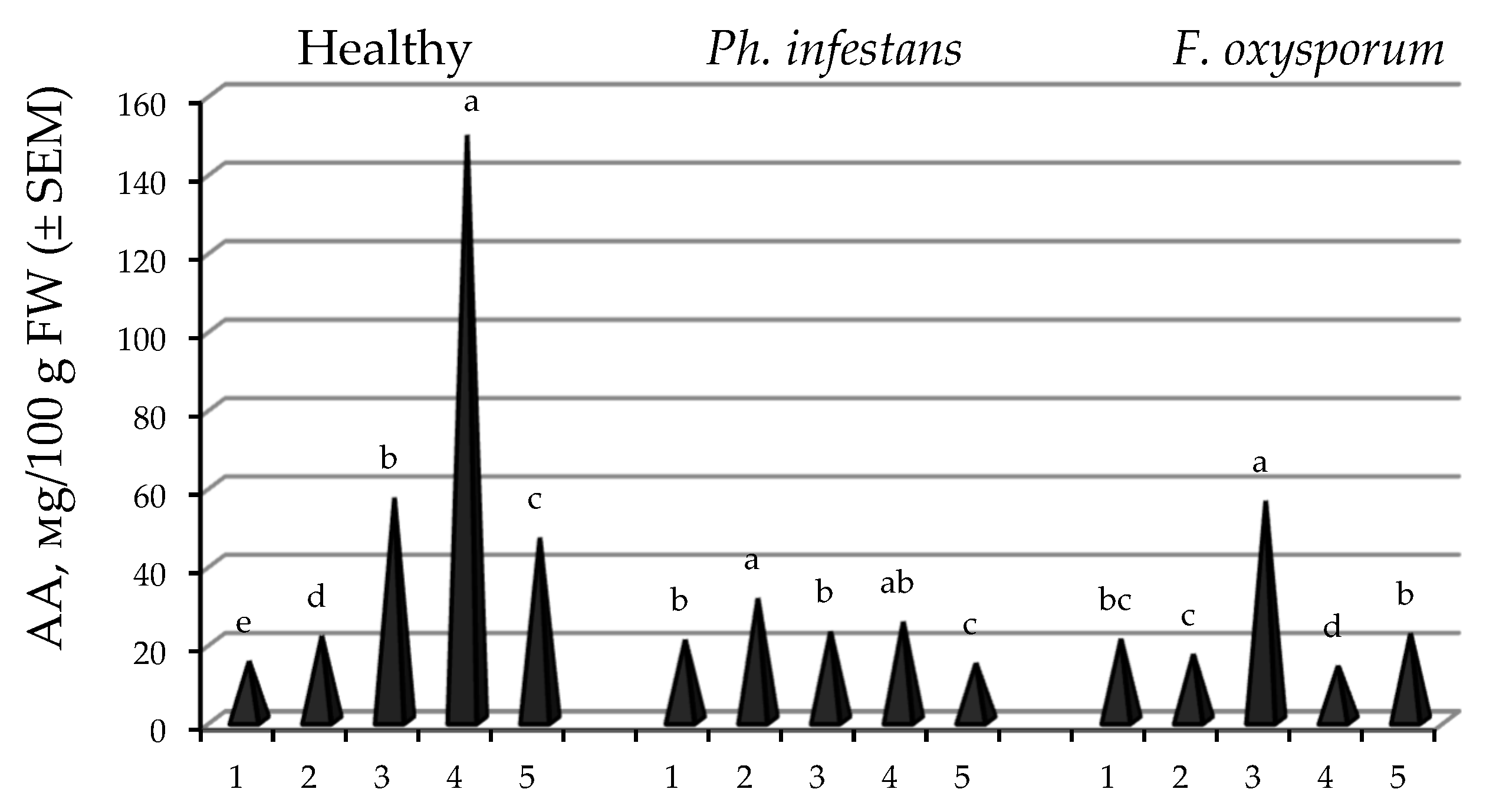
2.6. Effect of Endophytic B. subtilis (10-4, 26D), and Their Compositions with SA, on Ascorbic Acid (AA) Content in Stored Potato Tubers
3. Materials and Methods
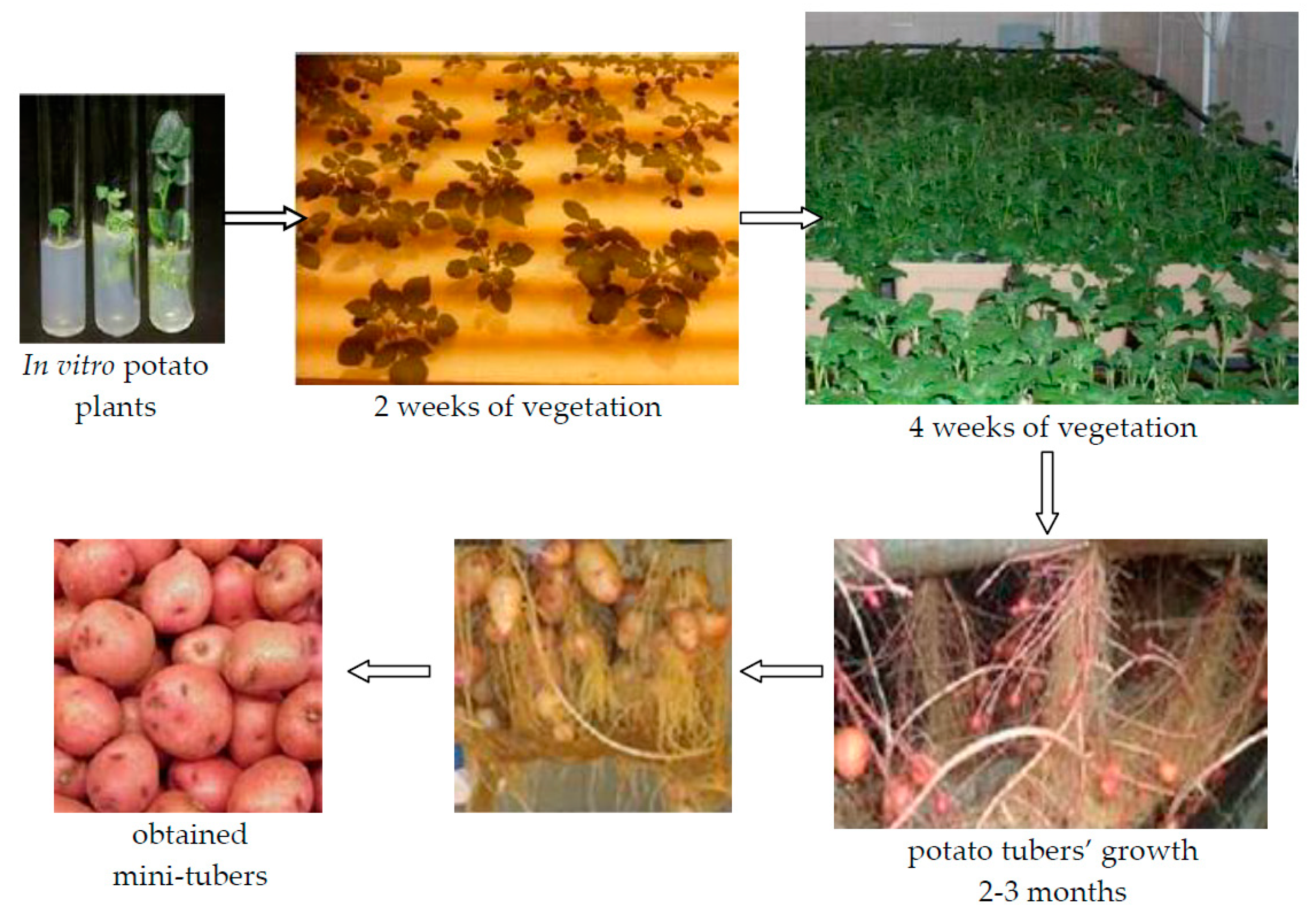
3.1. Plant Material, Scheme of Experiments, and Storage Conditions
3.2. Bacterial Strains and Phytopathogenic Fungi
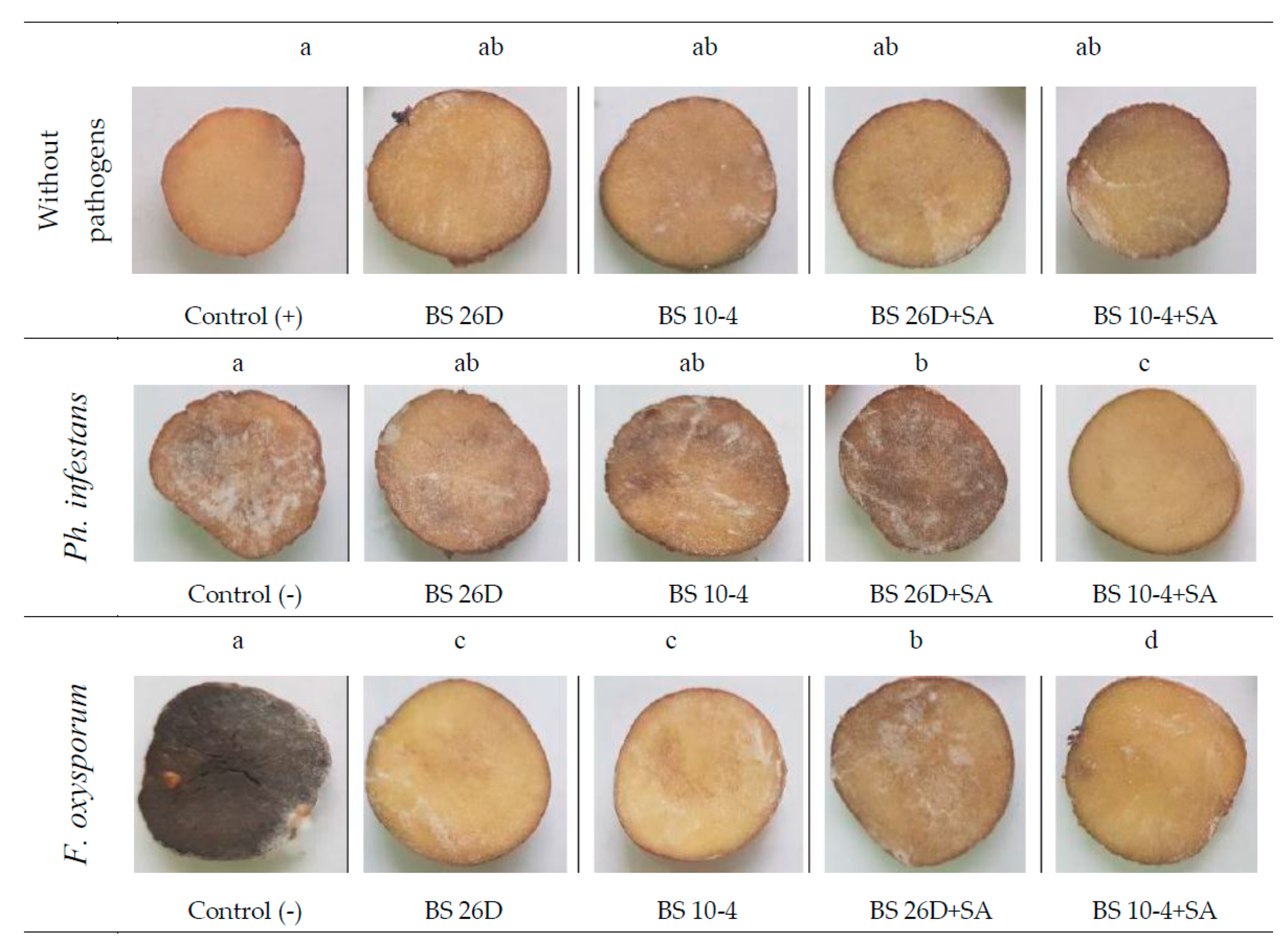
3.3. Assessment Visual Symptoms of Disease Development
3.4. Determination of the Ability of B. subtilis to Colonize Internal Tissues of Potato Tubers
3.5. Assessment the Quantitative Content (Titer) of B. subtilis Bacteria in Potato Tubers
3.6. Quantification of Proline (Pro) Content
3.7. Estimation of Lipid Peroxidation (MDA)
3.8. Ascorbic Acid (AA) Determination
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Food Losses and Waste. Available online: http://www.fao.org/food-loss-and-food-waste/en/ (accessed on 15 April 2019).
- Alamar, M.C.; Tosetti, R.; Landahl, S.; Bermejo, A.; Terry, L.A. Assuring potato tuber quality during storage: A future perspective. Front. Plant Sci. 2017, 8, 2034. [Google Scholar] [CrossRef] [Green Version]
- Dale, F. Breeding for Storage. 2014. Available online: https://potatoes.ahdb.org.uk/sites/default/files/publication_upload/Breeding%20for%20storage%20-%20Finlay%20Dale.pdf (accessed on 15 April 2019).
- Fry, W.E.; Thurston, H.D.; Stevenson, W.R. Late blight. In Compendium of Potato Disease; Stevenson, W.R., Loria, R., Franc, G.D., Weingartncr, D.P., Eds.; APS Press: St. Paul, MN, USA, 2001; pp. 28–30. [Google Scholar]
- Fry, W.; Goodwin, S. Re-emergence of potato and tomato late blight in the United States. Plant Dis. 1997, 81, 1349–1357. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, W. Late blight control strategies in the United States. Acta Hortic. 2008, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Secor, G.A.; Salas, B. Fusarium dry rot and Fusarium wilt. In Compendium of Potato Disease; Stevenson, W.R., Loria, R., Franc, G.D., Weingartner, D.P., Eds.; APS Press: St. Paul, MN, USA, 2001; pp. 23–25. [Google Scholar]
- Bourke, A. The Visitation of God? The Potato and the Great Irish Famine; Hill, J., Grada, C.O., Eds.; The Lilliput Press: Dublin, Ireland, 1993. [Google Scholar]
- Desjardins, A.E.; Plattner, R.D. Trichothecene toxin production by strains of Gibberella pulicaris (Fusarium sambucinum) in liquid culture and in potato tubers. J. Agric. Food. Chem. 1989, 37, 388–392. [Google Scholar] [CrossRef]
- Desjardins, A.; Hohn, T.; McCormick, S. Trichothecene biosynthesis in Fusarium species: Chemistry, genetics, and significance. Microbiol. Mol. Biol. Rev. 1993, 57, 595–604. [Google Scholar] [CrossRef] [Green Version]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Wisniewski, M.; Droby, S.; John, N.; Liu, J.; Schena, L. Alternative management technologies for postharvest disease control: The journey fromsimplicity to complexity. Postharv. Biol. Technol. 2016, 122, 3–10. [Google Scholar] [CrossRef]
- Lastochkina, O.; Seifi Kalhor, M.; Aliniaeifard, S.; Baymiev, A.; Pusenkova, L.; Garipova, S.; Kulabuhova, D.; Maksimov, I. Bacillus spp.: Efficient biotic strategy to control postharvest diseases of fruits and vegetables. Plants 2019, 8, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, V.; Bochow, H.; Dolej, S.; Dolej, S.; Fischer, I. Plant growth-promoting Bacillus subtilis strain as potential inducer of systemic resistance in tomato against Fusarium wilt. J. Plant Dis. Protect. 2000, 107, 145–154. [Google Scholar]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Gutiérrez, L.; Zeriouh, H.; Romero, D.; Cubero, J.; Vicente, A.; Pérez-García, A. The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate—And salicylic acid-dependent defense responses. Microb. Biotechnol. 2013, 6, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egamberdieva, D.; Wirth, S.J.; Shurigin, V.V.; Hashem, A.; Abd Allah, E.F. Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by Fusarium solani under salt stress. Front. Microbiol. 2017, 8, 1887. [Google Scholar] [CrossRef] [PubMed]
- Shafi, O.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Lastochkina, O.V.; Pusenkova, L.I.; Yuldashev, R.A.; Il’yasova, E.Y.; Aliniaeifard, S. Effect of Bacillus subtilis based biologicals on physiological and biochemical parameters of sugar beet (Beta vulgaris l.) plants infected with Alternaria alternata. Biol. Agric. 2018, 53, 958–968. [Google Scholar]
- Saleh, S.A.; Heuberger, H.; Schnitzler, W.H. Alleviation of salinity effect on artichoke productivity by Bacillus subtilis FZB24, supplemental Ca and micronutrients. J. Appl. Bot. Food Qual. 2005, 79, 24–32. [Google Scholar]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria help plants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Dimkpa, C.O.; Merten, D.; Svatos, A.; Büchel, G.; Kothe, E. Siderophores mediate reduced and increased uptake of cadmium by Streptomyces tendae F4 and sunflower (Helianthus annuus), respectively. J. Appl. Microbiol. 2009, 107, 1687–1696. [Google Scholar] [CrossRef]
- Sayed, S.A.; Atef, A.S.; Soha, E. Response of three sweet basil cultivars to inoculation with Bacillus subtilis and arbuscular mycorrhizal fungi under salt stress conditions. Nat. Sci. 2011, 9, 31–36. [Google Scholar]
- Turan, M.; Ekinci, M.; Yıldırım, E.; Güneş, K.; Karagöz, K.; Kotan, R.; Dursun, A. Plant growth promoting rhizobacteria improved growth, nutrient, and hormone content in cabbage (Brassica oleracea) seedlings. Turk. J. Agric. For. 2014, 38, 327–333. [Google Scholar] [CrossRef]
- Lastochkina, O.; Pusenkova, L.; Yuldashev, R.; Babaev, M.; Garipova, S.; Blagova, D.; Khairullin, R.; Aliniaeifard, S. effects of Bacillus subtilis on some physiological and biochemical parameters of Triticum aestivum L. (wheat) under salinity. Plant Physiol. Biochem. 2017, 121, 80–88. [Google Scholar] [CrossRef]
- Seifikalhor, M.S.; Aliniaeifard, S.; Self, M.; Javadi, E.; Bernard, F.; Li, T.; Lastochkina, O. Rhisobacteria Bacillus subtilis reduces toxic effects of high electrical conductivity in soilless culture of lettuce. Acta Hortic. 2018, 1227, 471–478. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Chen, F.; Li, Y.B.; Liu, S.X. A preliminary study on the biological control of postharvest diseases of Litchi fruit. J. Fruit Sci. 2001, 14, 185–186. [Google Scholar]
- Morales-de la Pena, M.; Welti-Chanes, J.; Martín-Belloso, O. Novel technologies to improve food safety and quality. Curr. Opin. Plant Sci. 2019, 30, 1–7. [Google Scholar] [CrossRef]
- Arroyave-Toroa, J.J.; Mosquera, S.; Villegas-Escobar, V. Biocontrol activity of Bacillus subtilis EA-CB0015 cells and lipopeptides against postharvest fungal pathogens. Biol. Control 2017, 114, 195–200. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Z.; Zhu, P.; Liu, P.; Zhang, Z.; Mastuda, Y.; Toyoda, H.; Xu, L. Postharvest biological control of melon pathogens using Bacillus subtilis EXWB1. J. Plant Pathol. 2010, 92, 645–652. [Google Scholar]
- Kim, G.H.; Koh, Y.J.; Jung, J.S.; Hur, J.S. Control of postharvest fruit rot diseases of kiwifruit by antagonistic bacterium Bacillus subtilis. Acta Hortic. 2015, 1096, 377–382. [Google Scholar] [CrossRef]
- Kim, Y.S.; Balaraju, K.; Jeon, Y. Effects of rhizobacteria Paenibacillus polymyxa APEC136 and Bacillus subtilis APEC170 on biocontrol of postharvest pathogens of apple fruits. J. Zhejiang Univ. Sci. B 2016, 17, 931–940. [Google Scholar] [CrossRef]
- Singh, V.; Deverall, B.J. Bacillus subtilis as a control agent against fungal pathogens of citrus fruit. Trans. Br. Mycol. Soc. 1984, 83, 487–490. [Google Scholar] [CrossRef]
- Mari, M.; Guizzardi, M.; Pratella, G.C. Biological control of gray mold in pears by antagonistic bacteria. Biol. Control 1996, 7, 30–37. [Google Scholar] [CrossRef]
- Touré, Y.; Ongena, M.; Jacques, P.; Guiro, A.; Thonart, P. Role of lipopeptides produced by Bacillus subtilis GA1 in the reduction of grey mould disease caused by Botrytis cinerea on apple. J. Appl. Microbiol. 2004, 96, 1151–1160. [Google Scholar] [CrossRef]
- Zhao, Y.; Shao, X.F.; Tu, K.; Chen, J.K. Inhibitory effect of Bacillus subtilis B10 on the diseases of postharvest strawberry. J. Fruit Sci. 2007, 24, 339–343. [Google Scholar]
- Jamalizadeh, M.; Etebarian, H.R.; Aminian, H.; Alizadeh, A. Biological control of Botrytis mali on apple fruit by use of Bacillus bacteria, isolated from the rhizosphere of wheat. Arch. Phytopathol. Plant Protect. 2010, 43, 1836–1845. [Google Scholar] [CrossRef]
- Kilani-Feki, O.; Ben Khedher, S.; Dammak, M.; Kamoun, A.; Jabnoun-Khiareddine, H.; Daami-Remadi, M.; Touns, S. Improvement of antifungal metabolites production by Bacillus subtilis V26 for biocontrol of tomato postharvest disease. Biol. Control 2016, 95, 73–82. [Google Scholar] [CrossRef]
- Miller, A.R. Harvest and Handling Injury: Physiology, Biochemistry, and Detection. In Postharvest Physiology and Pathology of Vegetables; Marcel Dekker Inc.: New York, NY, USA, 2003. [Google Scholar]
- Nagorska, K.; Bikowski, M.; Obuchowski, M. Multicellular behaviour and production of a wide variety of toxic substances support usage of Bacillus subtilis as a powerful biocontrol agent. Acta Biochim. Pol. 2007, 54, 495–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loon, L.C. Plant responses to plant growth-promoting rhizobacteria. Eur. J. Plant Pathol. 2007, 119, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Berg, G. Plant-microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef]
- Pandey, P.K.; Singh, M.C.; Singh, S.S.; Kumar, A.K.; Pathak, M.M.; Shakywar, R.C.; Pandey, A.K. Inside the plants: Endophytic bacteria and their functional attributes for plant growth promotion. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 11–21. [Google Scholar] [CrossRef]
- Sarma, B.K.; Yadav, K.S.; Singh, D.P.; Singh, H.B. Rhizobacteria mediated induced systemic tolerance in plants: Prospects for abiotic stress management. In Bacteria in Agrobiology: Stress Management; Springer: Berlin/Heidelberg, Germany, 2012; pp. 225–238. [Google Scholar]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Gao, H.; Xu, X.; Dai, Y.; He, H. Isolation, identification and characterization of Bacillus subtilis CF-3, a bacterium from fermented bean curd for controlling postharvest diseases of peach fruit. Food Sci. Technol. Res. 2016, 22, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Maksimov, I.V.; Khairullin, R.M. The role of Bacillus bacterium in formation of plant defense: Mechanism and reaction. In The Handbook of Microbial Bioresourses; Gupta, V.K., Sharma, G.D., Tuohy, M.G., Gaur, R., Eds.; CAB International: Galway, Ireland, 2016; Chapter 4; pp. 56–80. [Google Scholar]
- Maksimov, I.V.; Veselova, S.V.; Nuzhnaya, T.V.; Sarvarova, E.R.; Khairullin, R.M. Plant growth promoting bacteria in regulation of plant resistance to stress factors. Rus. J. Plant Physiol. 2015, 62, 715–726. [Google Scholar] [CrossRef]
- Yang, D.M.; Bi, Y.; Chen, X.R.; Ge, Y.H.; Zhao, J. Biological control of postharvest diseases with Bacillus subtilis (B1 strain) on muskmelons (Cucumis melo L. cv. Yindi). Acta Hortic. 2006, 712, 735–739. [Google Scholar] [CrossRef]
- Ahn, I.P.; Lee, S.W.; Kim, M.G.; Park, S.R.; Hwang, D.J.; Bae, S.C. Priming by rhizobacterium protects tomato plants from biotrophic and necrotrophic pathogen infections through multiple defense mechanisms. Mol. Cells 2011, 32, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Pusenkova, L.I.; Il’yasova, E.Y.; Lastochkina, O.V.; Maksimov, I.V.; Leonova, S.A. Changes in the species composition of the rhizosphereand phyllosphere of sugar beet under the impact of biological preparations based on endophytic bacteria and their metabolites. Eurasian Soil Sci. 2016, 49, 1136–1144. [Google Scholar] [CrossRef]
- Buchholz, F.; Kostic, T.; Sessitsch, A.; Mitter, B. The potential of plant microbiota in reducing postharvest food loss. Microb. Biotechnol. 2018, 11, 971–975. [Google Scholar] [CrossRef] [Green Version]
- Shakirova, F.M.; Avalbaev, A.M.; Bezrukova, M.V.; Fatkhutdinova, R.A.; Maslennikova, D.R.; Yuldashev, R.A.; Allagulova, C.R.; Lastochkina, O.V. Hormonal intermediates in the protective action of exogenous phytohormones in wheat plants under salinity: A case study on wheat. In Phytohormones and Abiotic Stress Tolerance in Plants; Khan, N., Nazar, R., Iqbal, N., Anjum, N., Eds.; Springer: Berlin, Germany, 2012; pp. 185–228. [Google Scholar]
- Fung, R.; Wang, C.; Smith, D.; Gross, K.; Tian, M. MeSA and MeJA increase steady-state transcript levels of alternative oxidase and resistance against chilling injury in sweet peppers (Capsicum annuum L.). Plant Sci. 2004, 166, 711–719. [Google Scholar] [CrossRef]
- Cai, C.; Xu, C.J.; Li, X.; Ferguson, I.; Chen, K.S. Accumulation of lignin in relation to change in activities of lignification enzymes in loquat fruit flesh after harvest. Postharvest Biol. Technol. 2006, 40, 163–169. [Google Scholar] [CrossRef]
- Asghari, M.; Aghdam, M.S. Impact of salicylic acid on post-harvest physiology of horticultural crops. Trends Food Sci. Technol. 2010, 21, 502–509. [Google Scholar] [CrossRef]
- El-Ghaouth, A.; Wilson, C.L.; Wisniewski, M.E. Biologically based alternatives to synthetic fungicides for the postharvest diseases of fruit and vegetables. In Diseases of Fruit and Vegetables; Samh, N., Ed.; Kluwen Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 511–535. [Google Scholar]
- Droby, S.; Chalutz, E.; Wilson, C.L.; Wisniewski, M.E. Biological control of postharvest diseases: A promising alternative to the use of synthetic fungicides. Phytoparasitica 1992, 20, 1495–1503. [Google Scholar] [CrossRef]
- Dokhanieh, A.Y.; Aghdam, M.S.; Fard, J.R.; Hassanpour, H. Postharvest salicylic acid treatment enhances antioxidant potential of cornelian cherry fruit. Sci. Hortic. 2013, 154, 31–36. [Google Scholar] [CrossRef]
- Wilson, C.L.; Wisniewski, M.E. Biological control of postharvest diseases of fruit and vegetables: An emerging technology. Ann. Rev. Phytopathol. 1989, 27, 425–441. [Google Scholar] [CrossRef]
- Droby, S.; Chalutz, E. Mode of action of biological agents of postharvest diseases. In Biological Control of Postharvest Diseases—Theory and Practice; Wilson, C.L., Wisniewski, M.E., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 63–75. [Google Scholar]
- Droby, S.; Chalutz, E.; Wilson, C.L.; Wisniewski, M. Characterization of the biocontrol activity of Debaryomyces hansenii in the control of Penicillium digitatum on grapefruit. Can. J. Microbiol. 1989, 35, 794–800. [Google Scholar] [CrossRef]
- Castoria, R.; de Curtis, F.; Lima, G.; Caputo, L.; Pacifico, S.; de Cicco, V. Aureobasidium pullulans (LS-30), an antagonist of postharvest pathogens of fruits: Study on its mode of action. Postharvest Biol. Technol. 2001, 32, 717–724. [Google Scholar] [CrossRef]
- Mardanova, A.M.; Hadieva, G.F.; Lutfullin, M.T.; Khilyas, I.V.; Minnullina, L.F.; Gilyazeva, A.G.; Bogomolnaya, L.M.; Sharipova, M.R. Bacillus subtilis strains with antifungal activity against the phytopathogenic fungi. Agric. Sci. 2017, 8, 1–20. [Google Scholar]
- Chebotar, V.K.; Kiprushkina, E.I. Application of microbial preparations in potato storage technologies. Dostizheniya nauki i tekhniki APK 2015, 29, 33–35. (In Russian) [Google Scholar]
- Netrusov, A.I.; Egorova, M.A.; Zakharchuk, L.M. Praktikum Po Mikrobiologii (A Practical Course in Microbiology); Tsentr “Akademiya”: Moscow, Russia, 2005. [Google Scholar]
- Maksimov, I.V.; Pusenkova, L.I.; Abizgildina, R.R. Biopreparation with endophytic bacterium Bacillus subtilis 26D created postharvest protecting effect in potato tubers. Agrochemistry 2011, 6, 43–48. (In Russian) [Google Scholar]
- Bates, l.S.; Waldern, R.P.; Teare, D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- GOST 24556-89. Products of Fruits and Vegetables Processing. Methods for Determination of Vitamin C. Izdatelstvo Standartov. 2003. Available online: http://docs.cntd.ru/document/gost-24556-89 (accessed on 18 January 2018).











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lastochkina, O.; Baymiev, A.; Shayahmetova, A.; Garshina, D.; Koryakov, I.; Shpirnaya, I.; Pusenkova, L.; Mardanshin, I.; Kasnak, C.; Palamutoglu, R. Effects of Endophytic Bacillus Subtilis and Salicylic Acid on Postharvest Diseases (Phytophthora infestans, Fusarium oxysporum) Development in Stored Potato Tubers. Plants 2020, 9, 76. https://doi.org/10.3390/plants9010076
Lastochkina O, Baymiev A, Shayahmetova A, Garshina D, Koryakov I, Shpirnaya I, Pusenkova L, Mardanshin I, Kasnak C, Palamutoglu R. Effects of Endophytic Bacillus Subtilis and Salicylic Acid on Postharvest Diseases (Phytophthora infestans, Fusarium oxysporum) Development in Stored Potato Tubers. Plants. 2020; 9(1):76. https://doi.org/10.3390/plants9010076
Chicago/Turabian StyleLastochkina, Oksana, Andrey Baymiev, Aysylu Shayahmetova, Darya Garshina, Igor Koryakov, Irina Shpirnaya, Liudmila Pusenkova, Il’dar Mardanshin, Cemal Kasnak, and Recep Palamutoglu. 2020. "Effects of Endophytic Bacillus Subtilis and Salicylic Acid on Postharvest Diseases (Phytophthora infestans, Fusarium oxysporum) Development in Stored Potato Tubers" Plants 9, no. 1: 76. https://doi.org/10.3390/plants9010076