High Below-Ground Productivity Allocation of Alpine Grasslands on the Northern Tibet

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Field Biomass Sampling
2.3. Laboratory Analysis
2.4. Productivity Estimation and Allocation
2.5. Statistical Analysis
3. Results
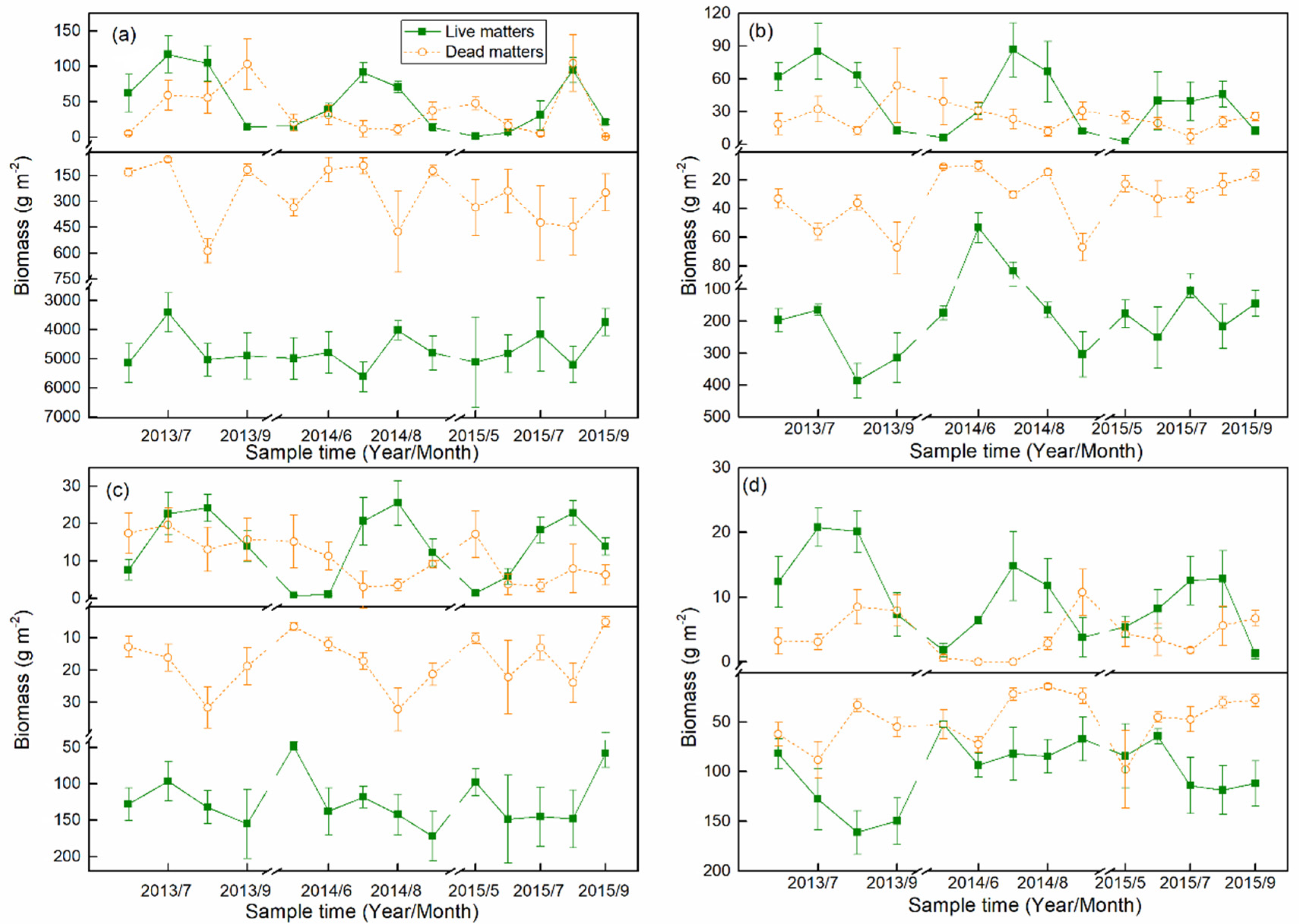
3.1. Biomass Dynamics
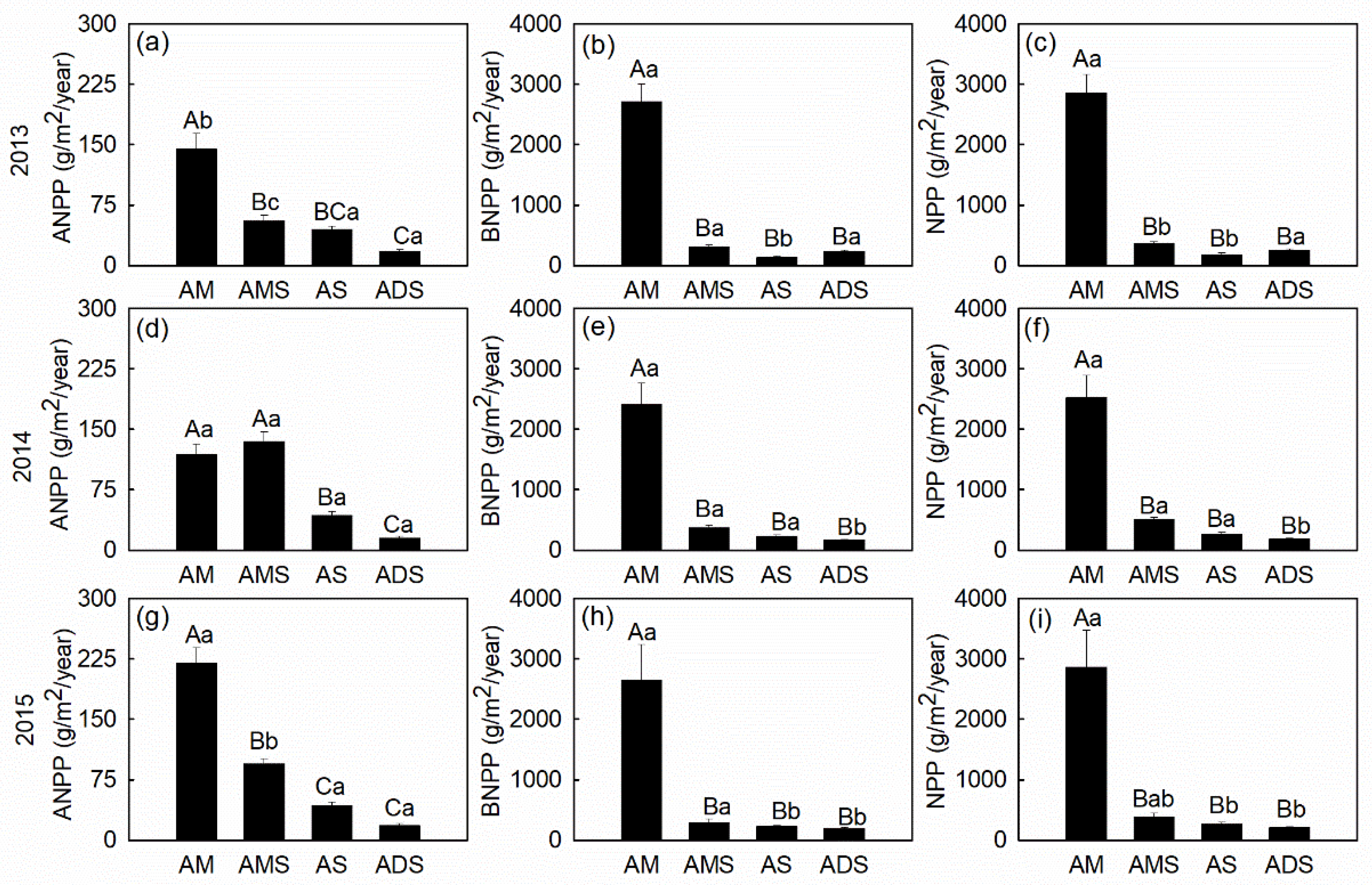
3.2. Productivity Dynamics
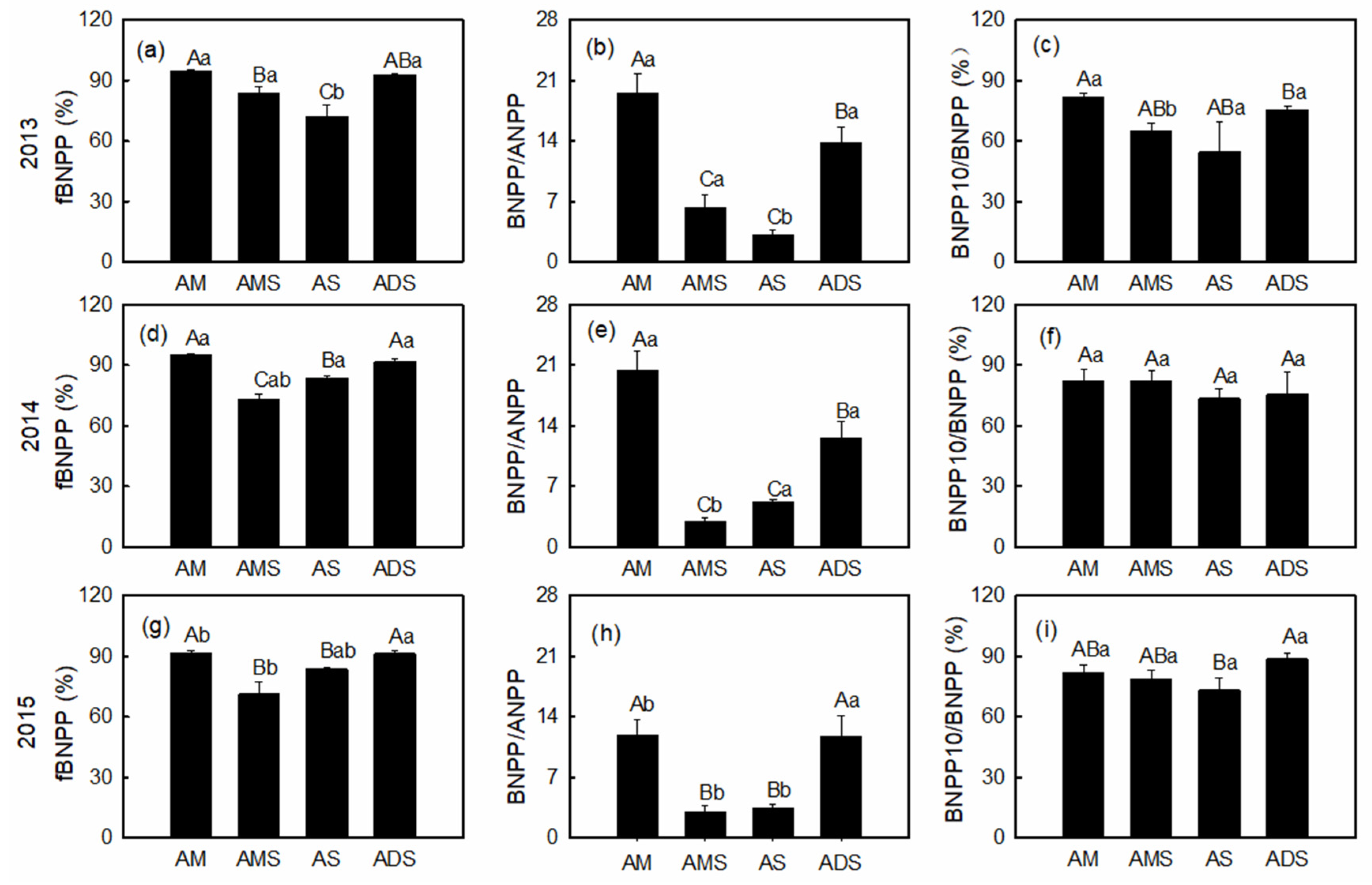
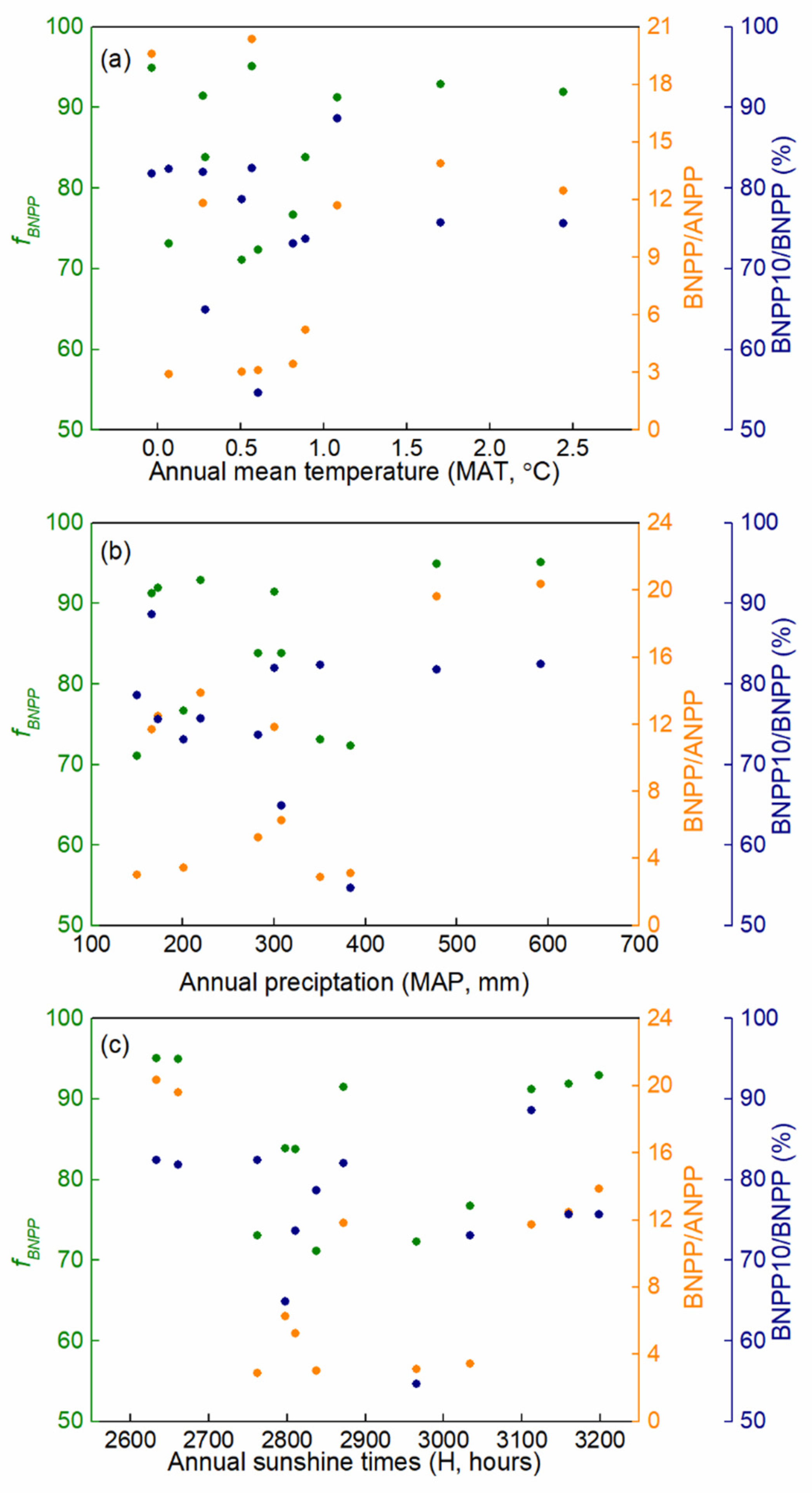
3.3. Productivity Allocation
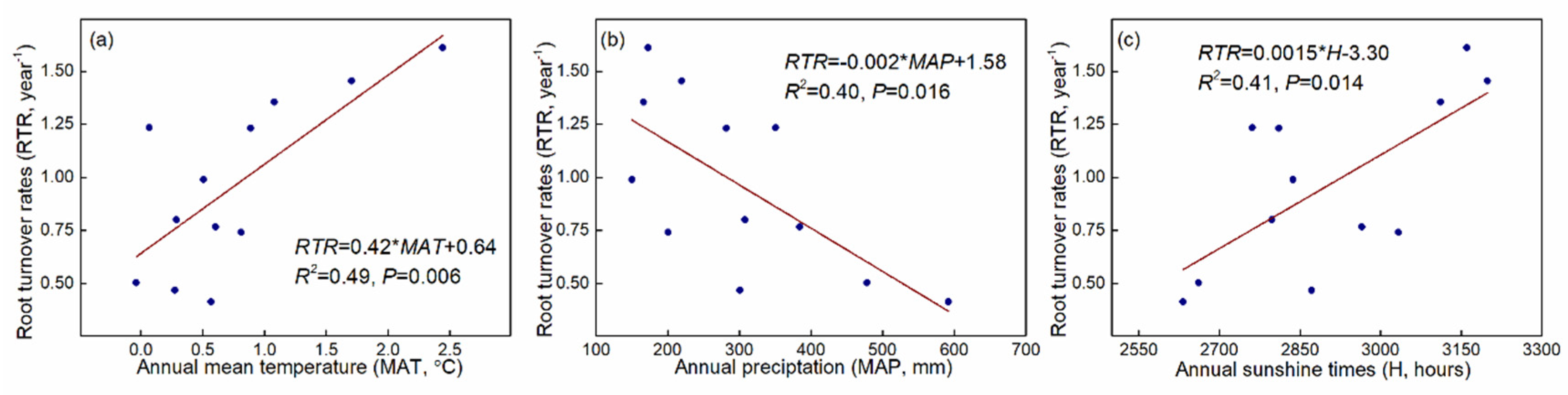
3.4. Root Turnover Rates
4. Discussion
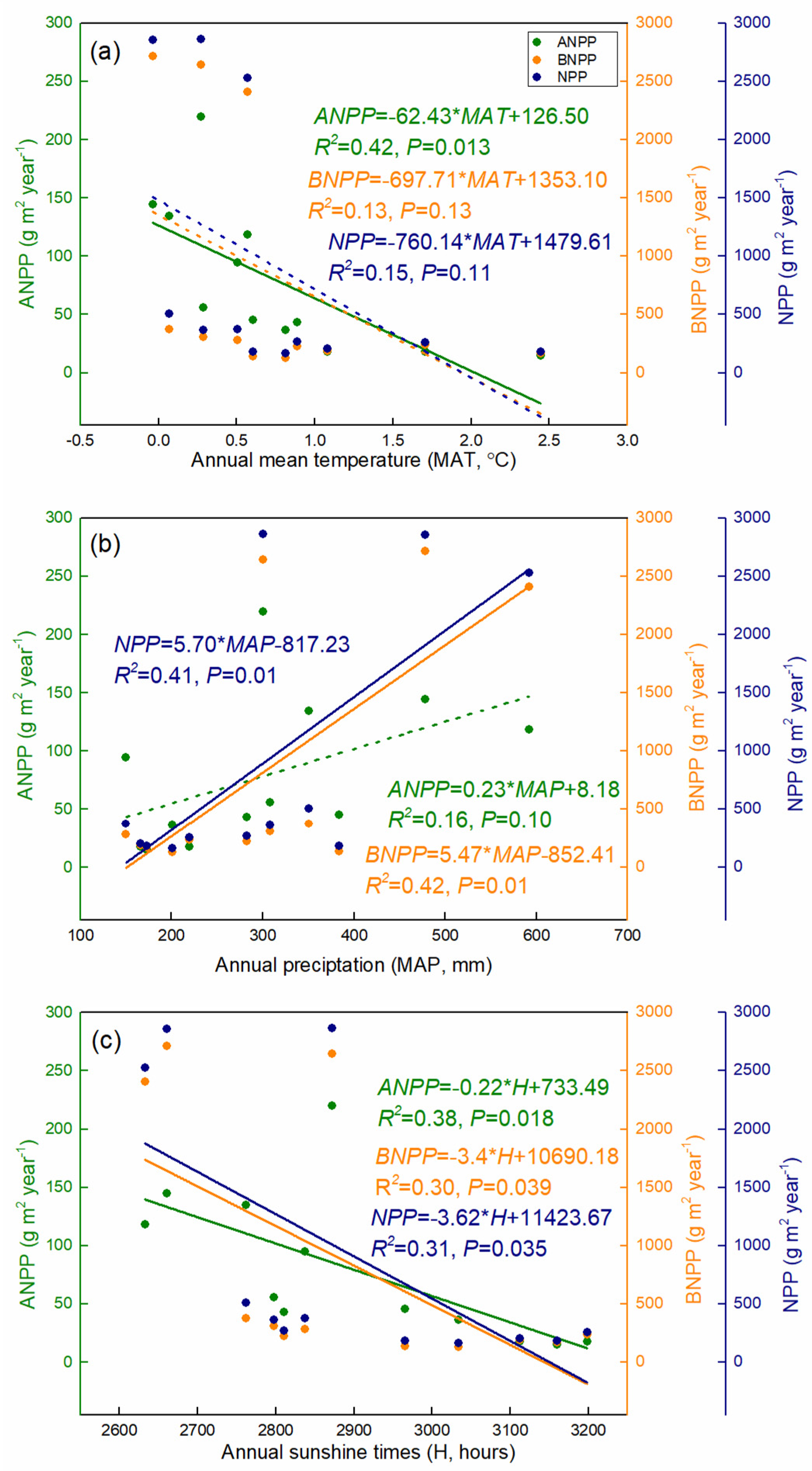
4.1. Productivity Dynamics
4.2. Productivity Allocation
4.3. Root Turnover Rates
4.4. Limitations and Applications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scurlock, J.M.O.; Olson, R.J. Terrestrial net primary productivity: A brief history and a new worldwide database. Environ. Rev. 2002, 10, 91–109. [Google Scholar] [CrossRef]
- Ni, J. Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China. Plant Ecol. 2004, 174, 217–234. [Google Scholar] [CrossRef]
- Litton, C.M.; Raich, J.W.; Ryan, M.G. Carbon allocation in forest ecosystems. Glob. Chang. Biol. 2007, 13, 2089–2109. [Google Scholar] [CrossRef]
- Hui, D.; Jackson, R.B. Geographical and interannual variability in biomass partitioning in grassland ecosystems: A synthesis of field data. New Phytol. 2006, 169, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Zerihun, A.; Montagu, K.D.; Hoffmann, M.B.; Bray, S.G. Patterns of below-and aboveground biomass in eucalyptus populnea woodland communities of northeast australia along a rainfall gradient. Ecosystems 2006, 9, 501–515. [Google Scholar] [CrossRef]
- Xia, J.; Yuan, W.; Wang, Y.-P.; Zhang, Q. Adaptive carbon allocation by plants enhances the terrestrial carbon sink. Sci. Rep. 2017, 7, 3341. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Kelly, R.H.; Parton, W.J.; Day, K.A.; Jackson, R.B.; Morgan, J.A.; Scurlock, J.M.O.; Tieszen, L.L.; Castle, J.V.; Ojima, D.S.; et al. Using simple environmental variables to estimate below-ground productivity in grasslands. Glob. Ecol. Biogeogr. 2002, 11, 79–86. [Google Scholar] [CrossRef]
- Sala, O.; Austin, A. Methods of estimating aboveground net primary productivity. In Methods in Ecosystem Science; Sala, O., Jackson, R., Mooney, H., Howarth, R., Eds.; Springer: New York, NY, USA, 2000; pp. 31–43. [Google Scholar]
- Lauenroth, W. Methods of estimating belowground net primary production. In Methods in Ecosystem Science; Sala, O., Jackson, R., Mooney, H., Howarth, R., Eds.; Springer: New York, NY, USA, 2000; pp. 58–71. [Google Scholar]
- Luo, T.X.; Li, W.H.; Zhu, H.Z. Estimated biomass and productivity of natural vegetation on the tibetan plateau. Ecol. Appl. 2002, 12, 980–997. [Google Scholar] [CrossRef]
- Scurlock, J.M.O.; Johnson, K.; Olson, R.J. Estimating net primary productivity from grassland biomass dynamics measurements. Glob. Chang. Biol. 2002, 8, 736–753. [Google Scholar] [CrossRef]
- McNaughton, S.; Milchunas, D.; Frank, D. How can net primary productivity be measured in grazing ecosystems? Ecology 1996, 77, 974–977. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; He, J. Variations in vegetation net primary production in the qinghai-xizang plateau, China, from 1982 to 1999. Clim. Chang. 2006, 74, 253–267. [Google Scholar] [CrossRef]
- Zhuang, Q.; He, J.; Lu, Y.; Ji, L.; Xiao, J.; Luo, T. Carbon dynamics of terrestrial ecosystems on the tibetan plateau during the 20th century: An analysis with a process-based biogeochemical model. Glob. Ecol. Biogeogr. 2010, 19, 649–662. [Google Scholar] [CrossRef]
- Scurlock, J.M.O.; Cramer, W.; Olson, R.J.; Parton, W.J.; Prince, S.D. Terrestrial npp: Toward a consistent data set for global model evaluation. Ecol. Appl. 1999, 9, 913–919. [Google Scholar] [CrossRef]
- Knapp, A.K.; Carroll, C.J.; Fahey, T.J. Patterns and controls of terrestrial primary production in a changing world. Ecol. Environ. 2014, 205–246. [Google Scholar] [CrossRef]
- Niu, B.; He, Y.; Zhang, X.; Fu, G.; Shi, P.; Du, M.; Zhang, Y.; Zong, N. Tower-based validation and improvement of modis gross primary production in an alpine swamp meadow on the tibetan plateau. Remote Sens. 2016, 8, 592. [Google Scholar] [CrossRef]
- Sun, J.; Du, W. Effects of precipitation and temperature on net primary productivity and precipitation use efficiency across China’s grasslands. GISci. Remote Sens. 2017, 54, 881–897. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, Y.; Ma, Y.; Ma, W.; Liang, C.; Flynn, D.F.B.; Schmid, B.; Fang, J.; He, J.S. Field-based observations of regional-scale, temporal variation in net primary production in tibetan alpine grasslands. Biogeosciences 2014, 11, 2003–2016. [Google Scholar] [CrossRef]
- Wen, L.; Dong, S.; Li, Y.; Wang, X.; Li, X.; Shi, J.; Dong, Q. The impact of land degradation on the c pools in alpine grasslands of the qinghai-tibet plateau. Plant Soil 2013, 368, 329–340. [Google Scholar] [CrossRef]
- Fisk, M.C.; Schmidt, S.K.; Seastedt, T.R. Topographic patterns of above- and belowground production and nitrogen cycling in alpine tundra. Ecology 1998, 79, 2253–2266. [Google Scholar] [CrossRef]
- Xu, X.; Sherry, R.A.; Niu, S.; Li, D.; Luo, Y. Net primary productivity and rain-use efficiency as affected by warming, altered precipitation, and clipping in a mixed-grass prairie. Glob. Chang. Biol. 2013, 19, 2753–2764. [Google Scholar] [CrossRef]
- Xu, X.; Niu, S.; Sherry, R.A.; Zhou, X.; Zhou, J.; Luo, Y. Interannual variability in responses of belowground net primary productivity (npp) and npp partitioning to long-term warming and clipping in a tallgrass prairie. Glob. Chang. Biol. 2012, 18, 1648–1656. [Google Scholar] [CrossRef]
- Gao, Y.; Giese, M.; Lin, S.; Sattelmacher, B.; Zhao, Y.; Brueck, H. Belowground net primary productivity and biomass allocation of a grassland in inner mongolia is affected by grazing intensity. Plant Soil 2008, 307, 41–50. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Matamala, R.; Gonzàlez-Meler, M.A.; Jastrow, J.D.; Norby, R.J.; Schlesinger, W.H. Impacts of fine root turnover on forest npp and soil c sequestration potential. Science 2003, 302, 1385–1387. [Google Scholar] [CrossRef]
- McCormack, M.L.; Crisfield, E.; Raczka, B.; Schnekenburger, F.; Eissenstat, D.M.; Smithwick, E.A.H. Sensitivity of four ecological models to adjustments in fine root turnover rate. Ecol. Model. 2015, 297, 107–117. [Google Scholar] [CrossRef]
- Lukac, M. Fine root turnover. In Measuring Roots; Mancuso, S., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 363–373. [Google Scholar]
- Wu, Y.B.; Wu, J.; Deng, Y.C.; Tan, H.C.; Du, Y.G.; Gu, S.; Tang, Y.H.; Cui, X.Y. Comprehensive assessments of root biomass and production in a kobresia humilis meadow on the qinghai-tibetan plateau. Plant Soil 2011, 338, 497–510. [Google Scholar] [CrossRef]
- Lopez-Marsico, L.; Altesor, A.; Oyarzabal, M.; Baldassini, P.; Paruelo, J.M. Grazing increases below-ground biomass and net primary production in a temperate grassland. Plant Soil 2015, 392, 155–162. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Lauenroth, W.K.; Gill, R. Turnover of root systems. In Root Ecology; de Kroon, H., Visser, E.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; Volume 168, pp. 61–89. [Google Scholar]
- Stewart, A.; Frank, D. Short sampling intervals reveal very rapid root turnover in a temperate grassland. Oecologia 2008, 157, 453–458. [Google Scholar] [CrossRef]
- West, J.B.; Espeleta, J.F.; Donovan, L.A. Fine root production and turnover across a complex edaphic gradient of a pinus palustris-aristida stricta savanna ecosystem. For. Ecol. Manag. 2004, 189, 397–406. [Google Scholar] [CrossRef]
- Maeght, J.L.; Gonkhamdee, S.; Clement, C.; Ayutthaya, S.I.N.; Stokes, A.; Pierret, A. Seasonal patterns of fine root production and turnover in a mature rubber tree (hevea brasiliensis mull. Arg.) stand- differentiation with soil depth and implications for soil carbon stocks. Front. Plant Sci. 2015, 6, 1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Li, B.; Zheng, D. A discussion on the boundary and area of the tibetan plateau in China. Geogr. Res. 2002, 21, 1–8. [Google Scholar]
- Kato, T.; Tang, Y.; Gu, S.; Hirota, M.; Du, M.; Li, Y.; Zhao, X. Temperature and biomass influences on interannual changes in co2 exchange in an alpine meadow on the qinghai-tibetan plateau. Glob. Chang. Biol. 2006, 12, 1285–1298. [Google Scholar] [CrossRef]
- Zhang, Y.; Qi, W.; Zhou, C.; Ding, M.; Liu, L.; Gao, J.; Bai, W.; Wang, Z.; Zheng, D. Spatial and temporal variability in the net primary production of alpine grassland on the tibetan plateau since 1982. J. Geogr. Sci. 2014, 24, 269–287. [Google Scholar] [CrossRef]
- Mao, S.; Wu, Q.; Zhu, J.; Li, H.; Zhang, F.; Li, Y. Response of the maintain performance in alpine grassland to enclosure on the northern tibetan plateau. Acta Prataculturae Sin. 2015, 24, 21–23. [Google Scholar]
- Zeng, C.; Wu, J.; Zhang, X. Effects of grazing on above- vs. Below-ground biomass allocation of alpine grasslands on the northern tibetan plateau. PLoS ONE 2015, 10, e0135173. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, X.; Shen, Z.; Shi, P.; Yu, C.; Chen, B. Effects of livestock exclusion and climate change on aboveground biomass accumulation in alpine pastures across the northern tibetan plateau. Chin. Sci. Bull. 2014, 59, 4332–4340. [Google Scholar] [CrossRef]
- Fu, G.; Sun, W.; Yu, C.-Q.; Zhang, X.-Z.; Shen, Z.-X.; Li, Y.-L.; Yang, P.-W.; Zhou, N. Clipping alters the response of biomass production to experimental warming: A case study in an alpine meadow on the tibetan plateau, China. J. Mt. Sci. 2015, 12, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Vogt, K.; Persson, H. Measuring growth and development of roots. In Techniques and Approaches in Forest Tree Ecophysiology; CRC: Boca Raton, FL, USA, 1991; pp. 477–501. [Google Scholar]
- Dahlman, R.C.; Kucera, C.L. Root productivity and turnover in native prairie. Ecology 1965, 46, 84–89. [Google Scholar] [CrossRef]
- Niu, B.; He, Y.; Zhang, X.; Du, M.; Shi, P.; Sun, W.; Zhang, L. Co2 exchange in an alpine swamp meadow on the central tibetan plateau. Wetlands 2017, 37, 1–19. [Google Scholar] [CrossRef]
- Kabacoff, R.I. R in Action, Data Analysis and Graphics with R; Manning Publications Co.: Shelter Island, NY, USA, 2011; p. 472. [Google Scholar]
- Johnson, J.W. Factors affecting relative weights: The influence of sampling and measurement error. Organ. Res. Methods 2004, 7, 283–299. [Google Scholar] [CrossRef]
- Wu, G.-L.; Du, G.-Z.; Liu, Z.-H.; Thirgood, S. Effect of fencing and grazing on a kobresia-dominated meadow in the qinghai-tibetan plateau. Plant Soil 2009, 319, 115–126. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Ji, C.; Han, W. Above- and belowground biomass allocation in tibetan grasslands. J. Veg. Sci. 2009, 20, 177–184. [Google Scholar] [CrossRef]
- Li, Y.; Dong, S.; Liu, S.; Zhou, H.; Gao, Q.; Cao, G.; Wang, X.; Su, X.; Zhang, Y.; Tang, L.; et al. Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the qinghai-tibetan plateau of China. Soil Biol. Biochem. 2015, 80, 306–314. [Google Scholar] [CrossRef]
- Wu, J.; Shen, Z.; Zhang, X. Precipitation and species composition primarily determine the diversity–productivity relationship of alpine grasslands on the northern tibetan plateau. Alp. Bot. 2014, 124, 13–25. [Google Scholar] [CrossRef]
- Knapp, A.K.; Smith, M.D. Variation among biomes in temporal dynamics of aboveground primary production. Science 2001, 291, 481–484. [Google Scholar] [CrossRef] [Green Version]
- Pandey, C.B.; Singh, J.S. Rainfall and grazing effects on net primary productivity in a tropical savanna, india. Ecology 1992, 73, 2007–2021. [Google Scholar] [CrossRef]
- Fu, G.; Shen, Z.-X.; Zhang, X.-Z. Increased precipitation has stronger effects on plant production of an alpine meadow than does experimental warming in the northern tibetan plateau. Agric. For. Meteorol. 2018, 249, 11–21. [Google Scholar] [CrossRef]
- Wu, Y.; Tan, H.; Deng, Y.; Wu, J.; Xu, X.; Wang, Y.; Tang, Y.; Higashi, T.; Cui, X. Partitioning pattern of carbon flux in a kobresia grassland on the qinghai-tibetan plateau revealed by field 13c pulse-labeling. Glob. Chang. Biol. 2010, 16, 2322–2333. [Google Scholar] [CrossRef]
- Chen, B.X.; Zhang, X.Z.; Tao, J.; Wu, J.S.; Wang, J.S.; Shi, P.L.; Zhang, Y.J.; Yu, C.Q. The impact of climate change and anthropogenic activities on alpine grassland over the qinghai-tibet plateau. Agr. For. Meteorol. 2014, 189, 11–18. [Google Scholar] [CrossRef]
- Gao, Q.; Li, Y.; Wan, Y.; Qin, X.; Jiangcun, W.; Liu, Y. Dynamics of alpine grassland npp and its response to climate change in northern tibet. Clim. Chang. 2009, 97, 515. [Google Scholar] [CrossRef]
- Liu, D.; Li, Y.; Wang, T.; Peylin, P.; MacBean, N.; Ciais, P.; Jia, G.; Ma, M.; Ma, Y.; Shen, M.; et al. Contrasting responses of grassland water and carbon exchanges to climate change between tibetan plateau and inner mongolia. Agric. For. Meteorol. 2018, 249, 163–175. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Wu, J.; Shen, Z.; Zhang, Y.; Xu, X.; Fan, Y.; Zhao, Y.; Yan, W. Root biomass distribution in alpine ecosystems of the northern tibetan plateau. Environ. Earth Sci. 2011, 64, 1911–1919. [Google Scholar] [CrossRef]
- Fan, J.; Zhong, H.; Harris, W.; Yu, G.; Wang, S.; Hu, Z.; Yue, Y. Carbon storage in the grasslands of China based on field measurements of above- and below-ground biomass. Clim. Chang. 2008, 86, 375–396. [Google Scholar] [CrossRef]
- Yang, F.; Lu, G.; Shi, S. The structure characteristics and production of alpine kobresia humilis meadow. Acta Biol. Plateau Sin. 1985, 4, 49–56. [Google Scholar]
- Yang, E.; Wang, Q.; Shi, S.H. Seasonal and Annual Biomass Dynamics of Kobresia Humilis Meadow. In Proceedings of the International Symposium of an Alpine Meadow Ecosystem, Beijing, China; Academia Sinica: Taipei, Taiwan, 1989; Volume 71, p. 80. [Google Scholar]
- Yang, F.; Wang, Q.; Shi, S. The allocation of the biomass and energy in kobresia humilis meadow, haibei districi, qinghai province. Acta Phytoecol. Geobot. Sin. 1987, 11, 106–112. [Google Scholar]
- Wang, W.; Wang, Q.; Deng, Z. Communities structural characteristic and plant distribution pattern in alpine kobresia meadow, haibei region of qinghai province. Acta Phytoecol. Sin. 1998, 22, 336–343. [Google Scholar]
- Pu, J.; Li, Y.; Zhao, L.; Yang, S. The relationship between seasonal changes of kobresia humilis meadow biomass and the meteorological factors. Acta Agrestia Sin. 2005, 13, 238–241. [Google Scholar]
- Hu, Z.; Sun, J.; Li, Y.; Long, R.; Yang, F. The characteristics of biomass and conversion efficiency of solar radiation for principal types of alpine grasslands in tianzhu, gansu province. Acta Phytoecol. Sin. 1994, 18, 121–131. [Google Scholar]
- Li, X.; Li, F.; Huang, B.; Qiao, Y.; Sun, H.; Sun, B. Seedling development and biomass accumulation of kobresia on qinghai -tibetan plateau. Acta Pratacultural Sci. 1996, 5, 48–54. [Google Scholar]
- Zhu, X.; Hu, Z.; Wan, C.; Chen, Q.; Ellis, J.E.; Swift, D.M.; Bartlett, T.E. The grassland type and production estimate of yongfeng rangeland. Acta Prataculturae Sin. 1993, 2, 72–78. [Google Scholar]
- Iversen, C.; Norby, R. Terrestrial plant productivity and carbon allocation in a changing climate. Glob. Environ. Chang. 2014, 297–316. [Google Scholar] [CrossRef]
- Ding, J.; Yang, T.; Zhao, Y.; Liu, D.; Wang, X.; Yao, Y.; Peng, S.; Wang, T.; Piao, S. Increasingly important role of atmospheric aridity on tibetan alpine grasslands. Geophys. Res. Lett. 2018, 45, 2852–2859. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distributions for terrestrial biomes. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef]
- Fitter, A.H.; Graves, J.D.; Self, G.K.; Brown, T.K.; Bogie, D.S.; Taylor, K. Root production, turnover and respiration under two grassland types along an altitudinal gradient: Influence of temperature and solar radiation. Oecologia 1998, 114, 20–30. [Google Scholar] [CrossRef]
- Ineson, P.; Self, G.K.; Bogie, D.S.; Benham, D.; Fitter, A.H.; Brown, T.K.; Graves, J.D. Root production and turnover in an upland grassland subjected to artificial soil warming respond to radiation flux and nutrients, not temperature. Oecologia 1999, 120, 575–581. [Google Scholar] [CrossRef]
- Shaver, G.R.; Billings, W.D. Root production and root turnover in a wet tundra ecosystem, barrow, alaska. Ecology 1975, 56, 401–409. [Google Scholar] [CrossRef]
- Fu, X.; Wang, J.; Wang, H.; Dai, X.; Yang, F.; Zhao, M. Response of the fine root production, phenology, and turnover rate of six shrub species from a subtropical forest to a soil moisture gradient and shading. Plant Soil 2015, 399, 135–146. [Google Scholar] [CrossRef]
- Deng, L.; Zhang, Z.N.; Shangguan, Z.P. Long-term fencing effects on plant diversity and soil properties in China. Soil Tillage Res. 2014, 137, 7–15. [Google Scholar] [CrossRef]
- Xiong, D.; Shi, P.; Sun, Y.; Wu, J.; Zhang, X. Effects of grazing exclusion on plant productivity and soil carbon, nitrogen storage in alpine meadows in northern tibet, China. Chin. Geogr. Sci. 2014, 24, 488–498. [Google Scholar] [CrossRef]
- Hu, Z.; Li, S.; Guo, Q.; Niu, S.; He, N.; Li, L.; Yu, G. A synthesis of the effect of grazing exclusion on carbon dynamics in grasslands in China. Glob. Chang. Biol. 2016, 22, 1385–1393. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Longitude (° E) | Latitude (° N) | Elevation (m) | MAP (mm) | MAT (°C) | Dominant Species * | Other Plant Genera |
|---|---|---|---|---|---|---|---|
| AM | 92.01 | 31.64 | 4532 | 458.15 | −0.41 | K. pygmaea | Potentilla, Saussurea, Gentiana |
| AMS | 90.31 | 31.39 | 4611 | 341.74 | −0.13 | S. purpurea | Oxytropis, Androsace, Edelweiss |
| AS | 86.91 | 32.08 | 4624 | 336.00 | 0.47 | S. purpurea | Rhodiola, Sinapis, Heteropappus |
| ADS | 83.25 | 33.17 | 4711 | 177.21 | 0.67 | S. purpurea S. glareosa | Oxytropis, Sinapis |
| NPP Dynamics ‡ | NPP Allocations ‡ | RTR ‡ | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ANPP | BNPP | NPP | fBNPP | BNPP/ANPP | BNPP10/BNPP † | ||||||||||
| Sources † | df | MS | P | MS | P | MS | P | MS | P | MS | P | MS | P | MS | P |
| AGT | 3 | 61038 | *** | 20976609 | *** | 23037555 | *** | 1311.9 | *** | 654.8 | *** | 661.7 | * | 2.58 | *** |
| MAT | 2 | 1410 | 0.22 | 55504 | 0.63 | 74614 | 0.58 | 10 | 0.65 | 3.4 | 0.62 | 149.3 | 0.42 | 0.09 | 0.18 |
| MAP | 2 | 10303 | ** | 29613 | 0.72 | 74851 | 0.58 | 41.7 | 0.36 | 129.9 | ** | 322.1 | 0.24 | 0.005 | 0.76 |
| H | 2 | 2617 | 0.1 | 5756 | 0.88 | 611 | 0.96 | 159.2 | 0.08 | 40.6 | 0.09 | 781.2 | 0.07 | 0.45 | ** |
| MAT * MAP | 4 | 230 | 0.62 | 81010 | 0.56 | 89880 | 0.55 | 3.6 | 0.79 | 0.5 | 0.85 | 24.3 | 0.75 | 0.05 | 0.32 |
| MAT *H | 4 | 924 | 0.32 | 96452 | 0.52 | 116265 | 0.50 | 27 | 0.46 | 10.3 | 0.39 | 0.40 | 0.97 | 0.20 | 0.05 |
| MAP *H | 4 | 7017 | ** | 41208 | 0.68 | 82231 | 0.57 | 0 | 0.99 | 35.8 | 0.11 | 470.2 | 0.16 | 0.12 | 0.12 |
| MAT | MAP | MAT *MAP | Intercept | R2 | P | |
|---|---|---|---|---|---|---|
| fBNPP | 20.18 *** (53.1) | 0.07 *** (46.9) | −0.06 * | 59.59 *** | 0.27 | *** |
| BNPP/ANPP | 14.30 *** (24.8) | 0.06 *** (75.2) | −0.04 ** | −10.84 | 0.41 | *** |
| BNPP10/BNPP | 12.62 | 0.03 | −0.06 | 70.11 | <0.1 | 0.57 |
| RTR | 0.30 *** (56.2) | −0.001 ** (43.8) | -- | 1.09 | 0.52 | *** |
| Year | AM (year−1) | AMS (year−1) | AS (year−1) | ADS (year−1) |
|---|---|---|---|---|
| 2013 | 0.51 ± 0.05 C a | 0.80 ± 0.09 B b | 0.77 ± 0.10 B b | 1.45 ± 0.10 A a |
| 2014 | 0.42 ± 0.06 C a | 1.23 ± 0.01 B a | 1.23 ± 0.07 B a | 1.61 ± 0.09 A a |
| 2015 | 0.47 ± 0.12 C a | 0.99 ± 0.16 B ab | 0.74 ± 0.05 BC b | 1.35 ± 0.11 A a |
| Mean | 0.46 ± 0.04 C | 1.01 ± 0.07 B | 0.91 ± 0.03 B | 1.47 ± 0.05 A |
| Site | Lon. (° E) | Lat. (° N) | Ele. (m) | MAP | MAT | Dominant Species | ANPP | BNPP | fBNPP | RTR | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Haibei | 101.38 | 37.22 | 3300 | 412.28 | 0.53 | K. humilis | 303.4 | 898.9 | 0.74 | / | [4] |
| Haibei | 101.38 | 37.22 | 3250 | 528.0 | −2.0 | K. humilis | 579.9 | 658.8 | 0.53 | 0.52 | [30] |
| Haibei | 101.38 | 37.22 | 3200–3400 | 528.0 | −2.95 | K. humilis | 282.6 | 654.0 | 0.70 | / | [62] |
| Haibei | 101.38 | 37.22 | 3200–3400 | 528.0 | −2.95 | K. humilis | 286.6 | 1134.1 | 0.80 | / | [63] |
| Haibei | 101.38 | 37.22 | 3200–3400 | 528.0 | −2.95 | K. humilis | 282.7 | 654.0 | 0.70 | / | [64] |
| Haibei | 101.38 | 37.22 | 3250 | 514.0 | −4.11 | K. pygmaea | 368.4 | 5604.8 | 0.94 | / | [65] |
| Haibei | 101.38 | 37.22 | 3250 | 514.0 | −4.11 | K. humilis | 418.5 | 2578.0 | 0.86 | / | [65] |
| Haibei | 101.38 | 37.22 | 3250 | 514.0 | −4.11 | K. tibetica | 518.4 | 11183.2 | 0.96 | / | [65] |
| Haibei | 101.20–101.38 | 37.48–37.45 | 3200–3600 | 546.1 | −1.29 | K. humilis | 309.4 | 1267.9 | 0.80 | 0.32 | [66] |
| Jinqiang | 103.53 | 37.62 | 2930–3200 | 236.0 | 4.31 | K. capillifolia | 373.0 | 5497.9 | 0.94 | / | [67] |
| Maqen | 100.23 | 34.65 | 3800 | 495.0 | −2.80 | K. pygmaea | 30.1 | 1704.1 | 0.98 | / | [68] |
| Maqen | 100.23 | 34.65 | 3800 | 495.0 | −2.81 | K. capillifolia | 26.4 | 2790.7 | 0.99 | / | [68] |
| Tianzhou | 103.53 | 37.67 | 2900–3700 | 236.0 | 4.31 | K. humilis | 373.0 | 5498.9 | 0.94 | / | [69] |
| Tianzhou | 103.53 | 37.67 | 2900–3100 | 236.0 | 4.31 | S. purpurea-K. spp | 535.8 | 3739.3 | 0.87 | / | [69] |
| Naqu | 92.01 | 31.64 | 4532 | 458.1 | −0.41 | K. pygmaea | 160.9 | 2588.9 | 0.94 | 0.46 | This study |
| Bangoin | 90.31 | 31.39 | 4611 | 341.7 | −0.13 | S. purpurea | 94.5 | 321.5 | 0.76 | 1.01 | This study |
| Nima | 86.91 | 32.08 | 4624 | 336.0 | 0.47 | S. purpurea | 41.7 | 163.1 | 0.77 | 0.91 | This study |
| Gaize | 83.25 | 33.17 | 4711 | 177.2 | 0.67 | S.purpurea-S.glareosa | 16.9 | 198.3 | 0.92 | 1.47 | This study |
| Northern Tibetan meadows | 73.6 | 683.4 | 0.90 | 0.32 | [41] | ||||||
| Northern Tibetan steppes | 30.2 | 122.4 | 0.80 | 0.28 | [41] | ||||||
| Northern Tibetan desert-steppes | 13.0 | 86.3 | 0.87 | 0.26 | [41] | ||||||
| Tibetan steppes | 9.8–267.4 | 44.6–1834.8 | 0.58–0.93 | / | [50] | ||||||
| Tibetan meadows | 31.8–347.5 | 82.9–2784.8 | 0.45–0.93 | / | [50] | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, B.; Zeng, C.; Zhang, X.; He, Y.; Shi, P.; Tian, Y.; Feng, Y.; Li, M.; Wang, Z.; Wang, X.; et al. High Below-Ground Productivity Allocation of Alpine Grasslands on the Northern Tibet. Plants 2019, 8, 535. https://doi.org/10.3390/plants8120535
Niu B, Zeng C, Zhang X, He Y, Shi P, Tian Y, Feng Y, Li M, Wang Z, Wang X, et al. High Below-Ground Productivity Allocation of Alpine Grasslands on the Northern Tibet. Plants. 2019; 8(12):535. https://doi.org/10.3390/plants8120535
Chicago/Turabian StyleNiu, Ben, Chaoxu Zeng, Xianzhou Zhang, Yongtao He, Peili Shi, Yuan Tian, Yunfei Feng, Meng Li, Zhipeng Wang, Xiangtao Wang, and et al. 2019. "High Below-Ground Productivity Allocation of Alpine Grasslands on the Northern Tibet" Plants 8, no. 12: 535. https://doi.org/10.3390/plants8120535