Vegetation Heterogeneity Effects on Soil Macro-Arthropods in an Alpine Tundra of the Changbai Mountains, China

 ,
,
Abstract
:1. Introduction
2. Results
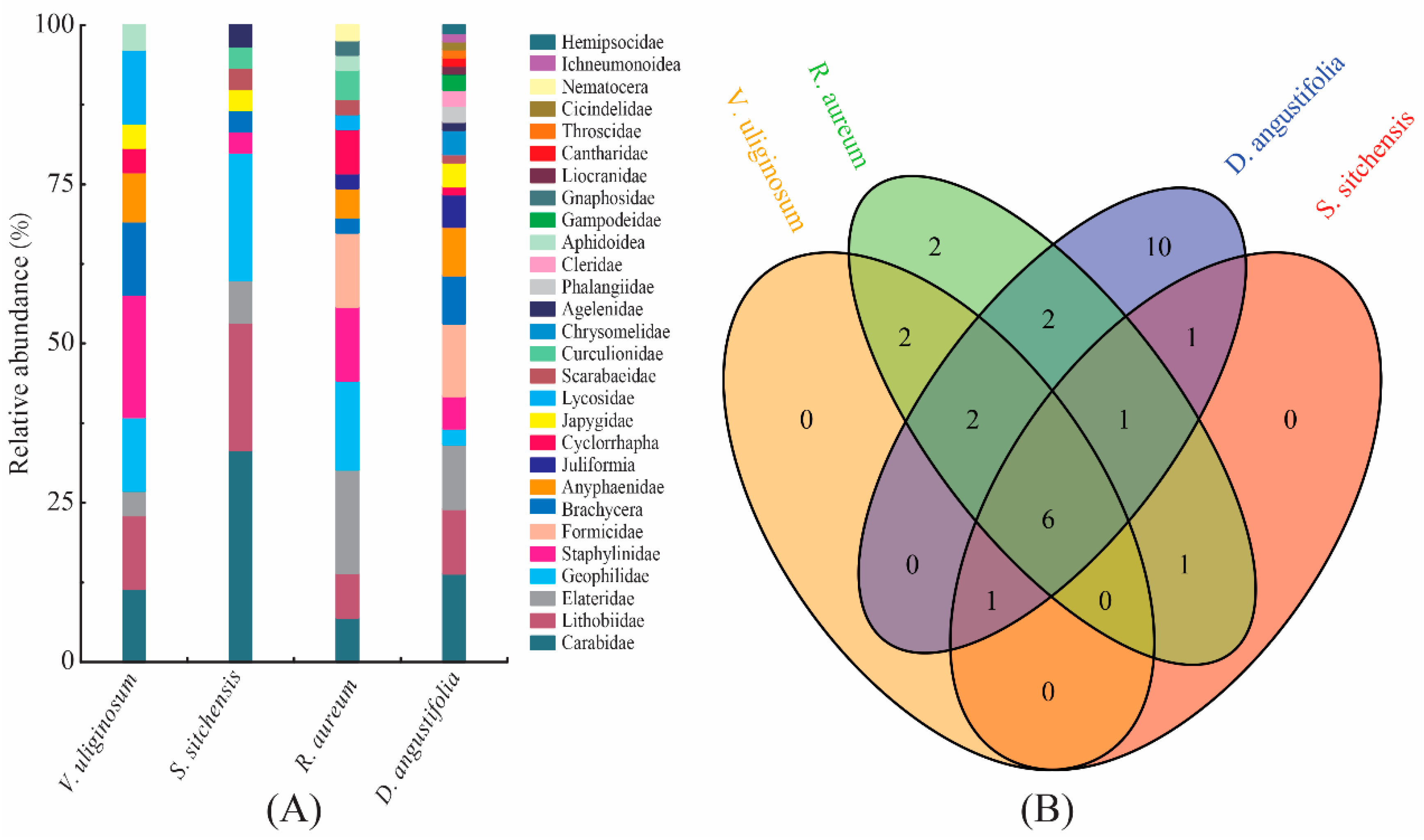
2.1. Taxonomic Composition of Soil Macro-Arthropod Communities
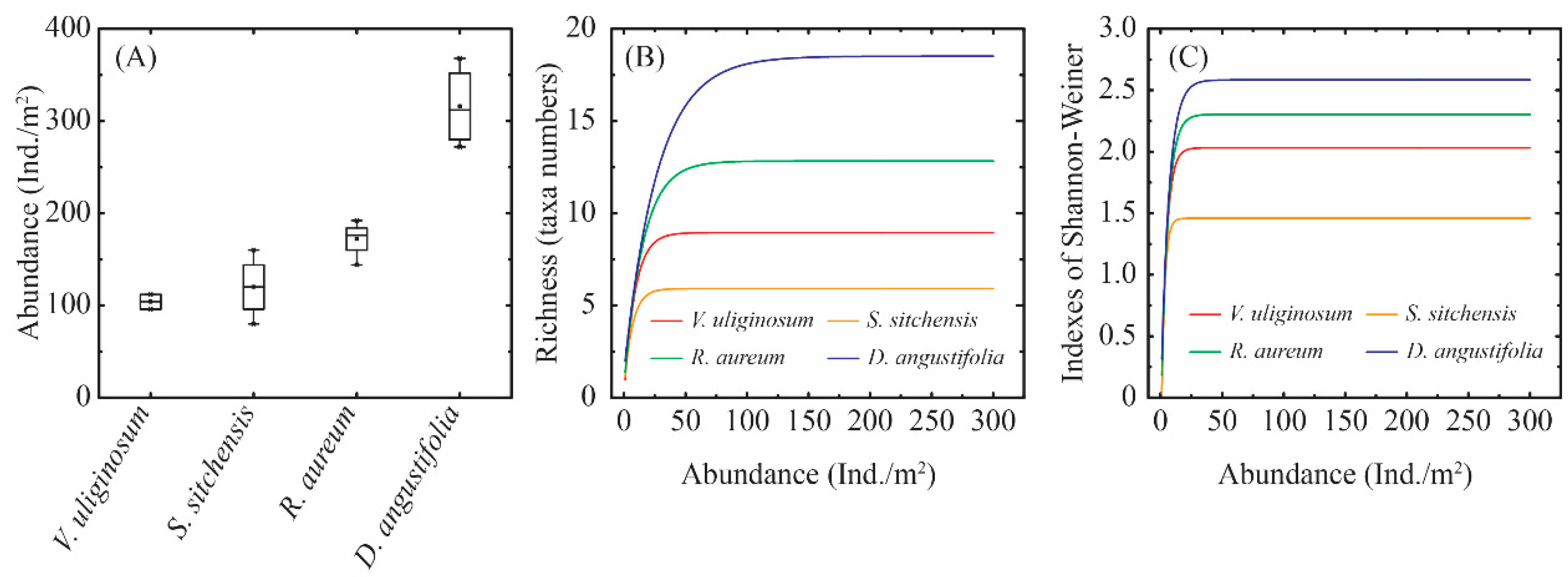
2.2. Diversity Characteristics of Soil Macro-Arthropods
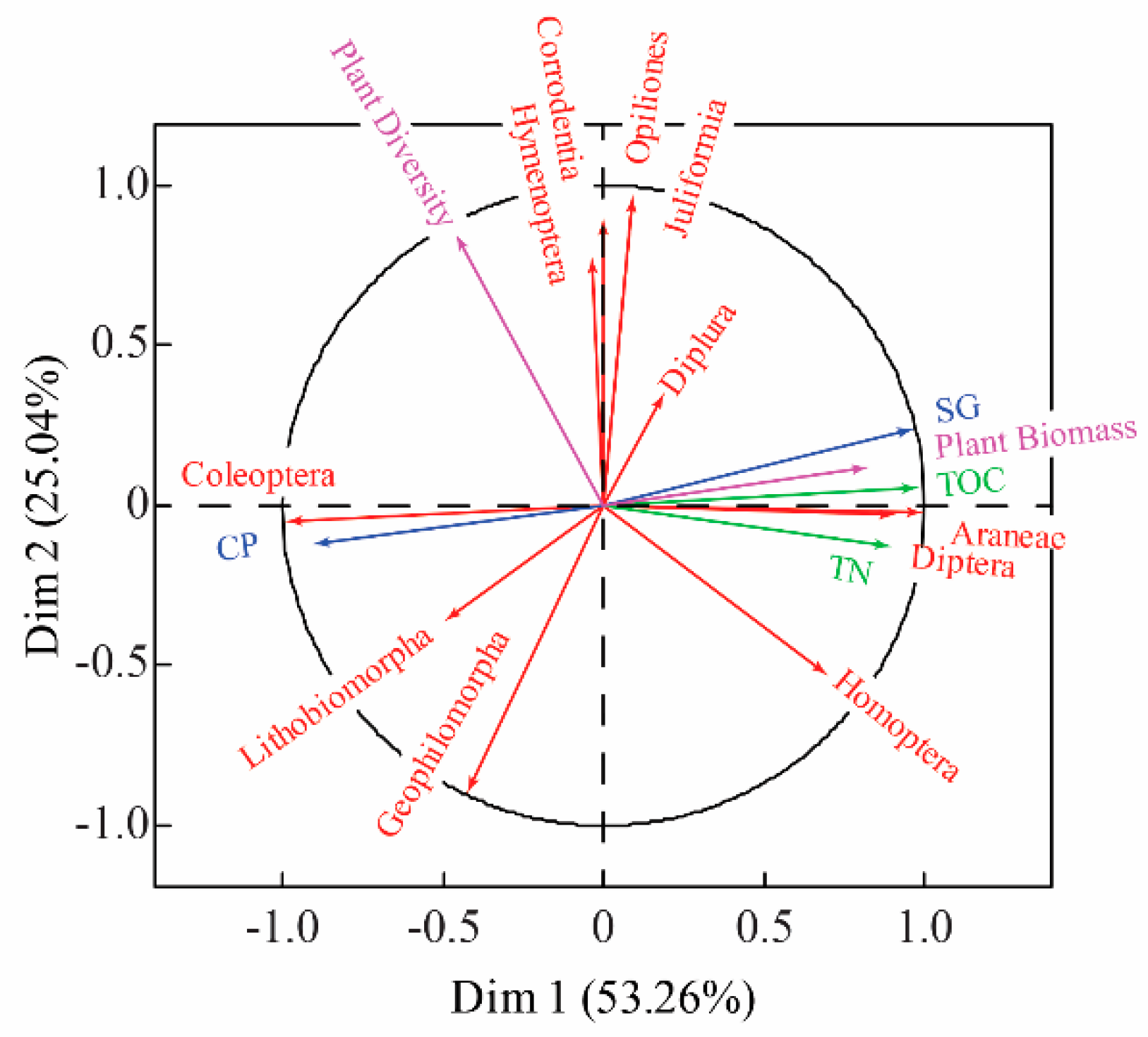
2.3. Correlation of Soil Macro-Arthropod Communities to Vegetation Heterogeneity
3. Discussion
3.1. Distribution Patterns of Soil Macro-Arthropods
3.2. Structure of the Soil Macro-Arthropod Communities
3.3. Effects of Vegetation Heterogeneity on Soil Macro-Arthropods
4. Materials and Methods
4.1. Site Descriptions
4.2. Sampling Design
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- MacArthur, R.H. Geographical Ecology: Patterns in the Distribution of Species; Harper & Row: New York, NY, USA, 1972. [Google Scholar]
- Mcclain, C.R.; Barry, J.P. Habitat heterogeneity, disturbance, and productivity work in concert to regulate biodiversity in deep submarine canyons. Ecology 2010, 91, 964–976. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef] [PubMed]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal species diversity driven by habitat heterogeneity/diversity: The importance of keystone structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef]
- Yin, X.; Song, B.; Dong, W.; Xin, W.; Wangm, Y. A review on the ecogeography of soil fauna in China. J. Geogr. Sci. 2010, 20, 333–346. [Google Scholar] [CrossRef]
- Joly, F.X.; Coq, S.; Coulis, M.; Nahmani, J.; Hättenschwiler, S. Litter conversion into detritivore faeces reshuffles the quality control over C and N dynamics during decomposition. Funct. Ecol. 2018, 32, 2605–2614. [Google Scholar] [CrossRef]
- Coulis, M.; Hättenschwiler, S.; Coq, S.; David, J.F. Leaf litter consumption by macroarthropods and burial of their faeces enhance decomposition in a mediterranean ecosystem. Ecosystems 2016, 19, 1104–1115. [Google Scholar] [CrossRef]
- Joly, F.X.; Coulis, M.; Gérard, A.; Fromin, N.; Hättenschwiler, S. Litter-type specific microbial responses to the transformation of leaf litter into millipede feces. Soil Biol. Biochem. 2015, 86, 17–23. [Google Scholar] [CrossRef]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Mundepi, A.; Phillips, C.A.; Callaham, M.A. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Kaneko, N.; Salamanca, E. Mixed leaf litter effects on decomposition rates and soil microarthropod communities in an oak-pine stand in Japan. Ecol. Res. 2010, 14, 131–138. [Google Scholar] [CrossRef]
- Gutiérrez-López, M.; Jesús, J.B.; Trigo, D.; Fernández, R.; Novo, M.; Díaz-Cosín, D.J. Relationships among spatial distribution of soil microarthropods, earthworm species and soil properties. Pedobiologia 2010, 53, 381–389. [Google Scholar]
- Shiels, A.B.; Sanford, R.L. Soil nutrient differences between two krummholz-form tree species and adjacent alpine tundra. Geoderma 2001, 102, 205–217. [Google Scholar] [CrossRef]
- Wipf, S.; Stoeckli, V.; Bebi, P. Winter climate change in alpine tundra: Plant responses to changes in snow depth and snowmelt timing. Clim. Chang. 2009, 94, 105–121. [Google Scholar] [CrossRef]
- Tolbert, W.W.; Tolbert, V.R.; Ambrose, R.E. Distribution, abundance, and biomass of Colorado alpine tundra arthropods. Arct. Alp. Res. 1977, 9, 221–234. [Google Scholar] [CrossRef]
- Sigmund, H.; Kari, K. Effect of simulated environmental change on alpine soil arthropods. Glob. Chang. Biol. 2010, 15, 2972–2980. [Google Scholar]
- Toyota, A.; Tayasu, I.; Fujimaki, R.; Kaneko, N.; Uchida, M.; Shibata, Y.; Hiura, T. Effects of vegetation switch and subsequent change in soil invertebrate composition on soil carbon accumulation patterns, revealed by radiocarbon concentrations. Radiocarbon 2010, 52, 1471–1486. [Google Scholar] [CrossRef]
- de Araújo, A.S.F.; Eisenhauer, N.; Nunes, L.A.P.L.; Leite, L.F.C.; Cesarz, S. Soil surface-active fauna in degraded and restored lands of Northeast Brazil. Land Degrad. Dev. 2015, 26, 1–8. [Google Scholar]
- García-Palacios, P.; Maestre, F.T.; Kattge, J.; Wall, D.H. Climate and litter quality differently modulate the effects of soil fauna on litter decomposition across biomes. Ecol. Lett. 2013, 16, 1045–1053. [Google Scholar] [Green Version]
- Sauvadet, M.; Chauvat, M.; Brunet, N.; Bertrand, I. Can changes in litter quality drive soil fauna structure and functions? Soil Biol. Biochem. 2017, 107, 94–103. [Google Scholar] [CrossRef]
- Blott, S.J.; Pye, K. Particle size distribution analysis of sand-sized particles by laser diffraction: An experimental investigation of instrument sensitivity and the effects of particle shape. Sedimentology 2010, 53, 671–685. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, Z.; Malhi, S.S.; Vera, C.L.; Zhang, Y.; Wang, J. Particle-size effects on soil temperature, evaporation, water use efficiency and watermelon yield in fields mulched with gravel and sand in semi-arid Loess Plateau of northwest China. Agr. Water Manag. 2010, 97, 917–923. [Google Scholar]
- Riutta, T.; Slade, E.M.; Bebber, D.P.; Taylor, M.E.; Malhi, Y.; Riordan, P.; Macdonald, D.W.; Morecroft, M.D. Experimental evidence for the interacting effects of forest edge, moisture and soil macrofauna on leaf litter decomposition. Soil Biol. Biochem. 2012, 49, 124–131. [Google Scholar] [CrossRef]
- Ma, C.; Yin, X. Responses of soil invertebrates to different forest types in the Changbai Mountains of China. J. For. Res.-JPN 2019, 24, 153–161. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, Z.; Feng, J. Geographical and Ecological Security of the Changbai Mountains; Northeast Normal University Press: Changchun, China, 2015; pp. 258–275. [Google Scholar]
- Haimi, J.; Laamanen, J.; Penttinen, R.; Räty, M.; Koponen, S.; Kellomäki, S.; Niemelä, P. Impacts of elevated CO2 and temperature on the soil fauna of boreal forests. Appl. Soil Ecol. 2005, 30, 104–112. [Google Scholar] [CrossRef]
- Jiang, Y.; Yin, X.; Wang, F. Composition and spatial distribution of soil mesofauna along an elevation gradient on the North Slope of the Changbai Mountains, China. Pedosphere 2015, 25, 811–824. [Google Scholar] [CrossRef]
- Ma, C.; Yin, X.; Kou, X.; Wang, Z.; Li, X.; Jiang, Y.; Wang, H.; Bernard, E.C. Effects of soil fauna on cellulose and lignin decomposition of plant litter in the Changbai Mountain, China. Environ. Entomol. 2019, 48, 592–602. [Google Scholar] [CrossRef]
- Staley, J.T.; Hodgson, C.J.; Mortimer, S.R.; Morecroft, M.D.; Masters, G.J.; Brown, V.K.; Taylor, M.E. Effects of summer rainfall manipulations on the abundance and vertical distribution of herbivorous soil macro-invertebrates. Eur. J. Soil Biol. 2007, 43, 189–198. [Google Scholar] [CrossRef]
- Brown, J.; Scholtz, C.H.; Janeau, J.L.; Grellier, S.; Podwojewski, P. Dung beetles (Coleoptera: Scarabaeidae) can improve soil hydrological properties. Appl. Soil Ecol. 2010, 46, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Ismail, S.M.; Ozawa, K. Improvement of crop yield, soil moisture distribution and water use efficiency in sandy soils by clay application. Appl. Clays Sci. 2007, 37, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S. Dependence of unfrozen water content in unsaturated frozen clay soil on initial soil moisture content. Soil Sci. Plant Nutr. 2004, 50, 603–606. [Google Scholar] [CrossRef] [Green Version]
- Cramer, M.J.; Willig, M.R. Habitat heterogeneity, species diversity and null models. Oikos 2005, 108, 209–218. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Habitat selection and population interactions: The search for mechanism. Am. Nat. 1991, 137, S5–S28. [Google Scholar] [CrossRef]
- García-Tejero, S.; Taboada, Á. Microhabitat heterogeneity promotes soil fertility and ground-dwelling arthropod diversity in Mediterranean wood-pastures. Agr. Ecosyst. Environ. 2016, 233, 192–201. [Google Scholar]
- Yin, W. Pictorial Keys to Soil Animals of China; Science Press: Beijing, China, 1998; pp. 15–358. [Google Scholar]
- Bao, S. Soil Agro-Chemistrical Analysis; China Agricultural Press: Beijing, China, 2000; pp. 25–151. [Google Scholar]
- Day, P.R. Particle fractionation and particle size analysis. In Methods of Soil Analysis, Part 1, Physical and Mineralogical Methods, 1st ed.; Black, C.A., Ed.; ASA: Madison, WI, USA, 1965; pp. 545–567. [Google Scholar]
- Chen, H.B. VennDiagram: Generate High-Resolution Venn and Euler Plots. R Package Version 1. 6.17. Available online: https://CRAN.R-project.org/package=VennDiagram (accessed on 16 April 2016).
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 50. Available online: http://purl.oclc.org/estimates (accessed on 1 December 2018).
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna (Austria): R Foundation for Statistical Computing. Available online: https://www.R-project.org (accessed on 16 March 2016).
- Raivo, K. Pheatmap: Pretty Heatmaps. R Package Version 1.0.8. Available online: https://CRAN.R-project.org/package=pheatmap. (accessed on 4 January 2019).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.3-5. Available online: https://CRAN.R-project.org/package=vegan (accessed on 4 February 2019).
- Lamentowicz, M.; Lamentowicz, Ł.; van der Knaap, W.O.; Gabka, M.; Mitchell, E.A.D. Contrasting species-environment relationships in communities of Testate Amoebae, bryophytes and vascular plants along the Fen-bog gradient. Microb. Ecol. 2009, 59, 499–510. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J.; Valentin, D. Multiple factor analysis: Principal component analysis for multitable and multiblock data sets. WIREs Comput. Stat. 2013, 5, 149–179. [Google Scholar] [CrossRef]
- Rao, C.R. Use of Hellinger distance in graphical displays. In Multivariate Statistics and Matrices in Statistics; Tiit, E.M., Kollo, T., Niemi, H., Eds.; Brill Academic Publisher: Leiden, The Netherlands, 1995; pp. 143–161. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar]
- Grace, J.B. Structural Equation Modeling and Natural Systems; Cambridge University Press: New York, NY, USA, 2006. [Google Scholar]
- Birkhofer, K.; Diekoetter, T.; Boch, S.; Fischer, M.; Mueller, J.; Socher, S.; Wolters, V. Soil fauna feeding activity in temperate grassland soils increases with legume and grass species richness. Soil Biol. Biochem. 2011, 43, 2200–2207. [Google Scholar] [CrossRef]
- Schmidt, A.; John, K.; Auge, H.; Brandl, R.; Horgan, F.G.; Settele, J.; Zaitsev, A.S.; Wolters, V.; Schädler, M. Compensatory mechanisms of litter decomposition under alternating moisture regimes in tropical rice fields. Appl. Soil Ecol. 2016, 107, 79–90. [Google Scholar] [CrossRef]
- Fox, J.; Nie, Z.; Byrnes, J. Sem: Structural Equation Models. R Package Version 3.1-8. 2016. Available online: https://CRAN.R-project.org/package=sem (accessed on 25 February 2016).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitats | Location | Elevation (m) | Soil Organic Matter (g/kg) | Total N (g/kg) | Clay Particle (%) | Sand Grains (%) | Plant Diversity | Plant Biomass (g/m2) |
|---|---|---|---|---|---|---|---|---|
| Vaccinium uliginosum habitat | 42°24′7.18″ N 128°5′48.99″ E | 2250 | 27.23 | 9.74 | 6.09 | 39.94 | 0.91 | 1632 |
| Sanguisorba sitchensis habitat | 42°22′23.3″ N 128°5′53.0″ E | 2260 | 11.87 | 5.99 | 9.75 | 12.22 | 1.36 | 686.4 |
| Rhododendron aureum habitat | 42°23′56.9″ N 128°8′20.4″ E | 2253 | 20.33 | 8.78 | 9.68 | 23.41 | 1.14 | 496 |
| Deyeuxia angustifolia habitat | 42°24′51.8″ N 128°8′33.9″ E | 2245 | 21.11 | 7.72 | 7.46 | 33.12 | 1.71 | 1268.8 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, Y.; Wang, Z.; Ma, C.; He, H.; Xu, J.; Jin, Y.; Wang, H.; Zheng, X. Vegetation Heterogeneity Effects on Soil Macro-Arthropods in an Alpine Tundra of the Changbai Mountains, China. Plants 2019, 8, 418. https://doi.org/10.3390/plants8100418
Tao Y, Wang Z, Ma C, He H, Xu J, Jin Y, Wang H, Zheng X. Vegetation Heterogeneity Effects on Soil Macro-Arthropods in an Alpine Tundra of the Changbai Mountains, China. Plants. 2019; 8(10):418. https://doi.org/10.3390/plants8100418
Chicago/Turabian StyleTao, Yan, Zhongqiang Wang, Chen Ma, Hongshi He, Jiawei Xu, Yinghua Jin, Haixia Wang, and Xiaoxue Zheng. 2019. "Vegetation Heterogeneity Effects on Soil Macro-Arthropods in an Alpine Tundra of the Changbai Mountains, China" Plants 8, no. 10: 418. https://doi.org/10.3390/plants8100418