Tomato Plants Treated with Systemin Peptide Show Enhanced Levels of Direct and Indirect Defense Associated with Increased Expression of Defense-Related Genes

 ,
,
Abstract
:1. Introduction
2. Results
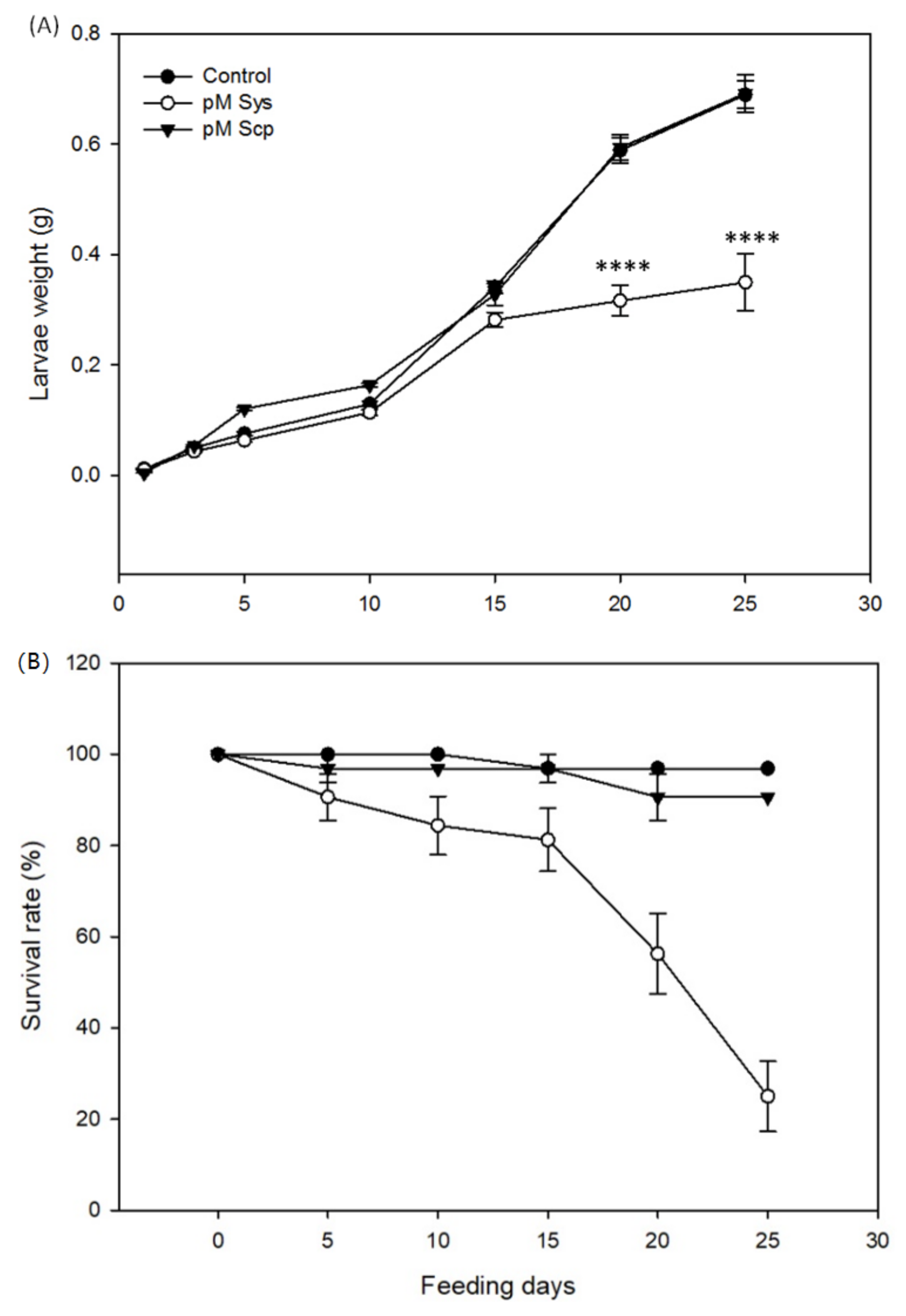
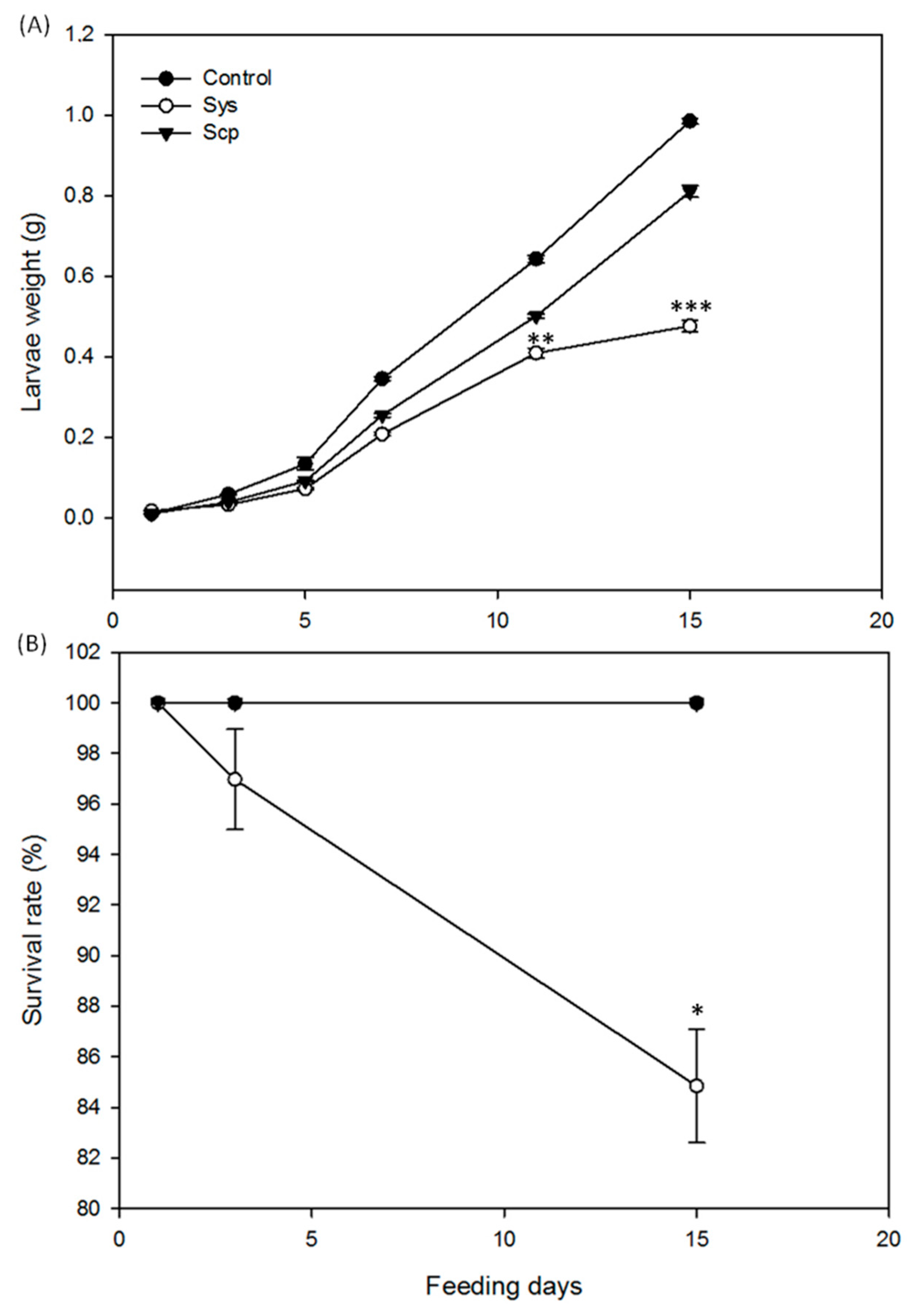
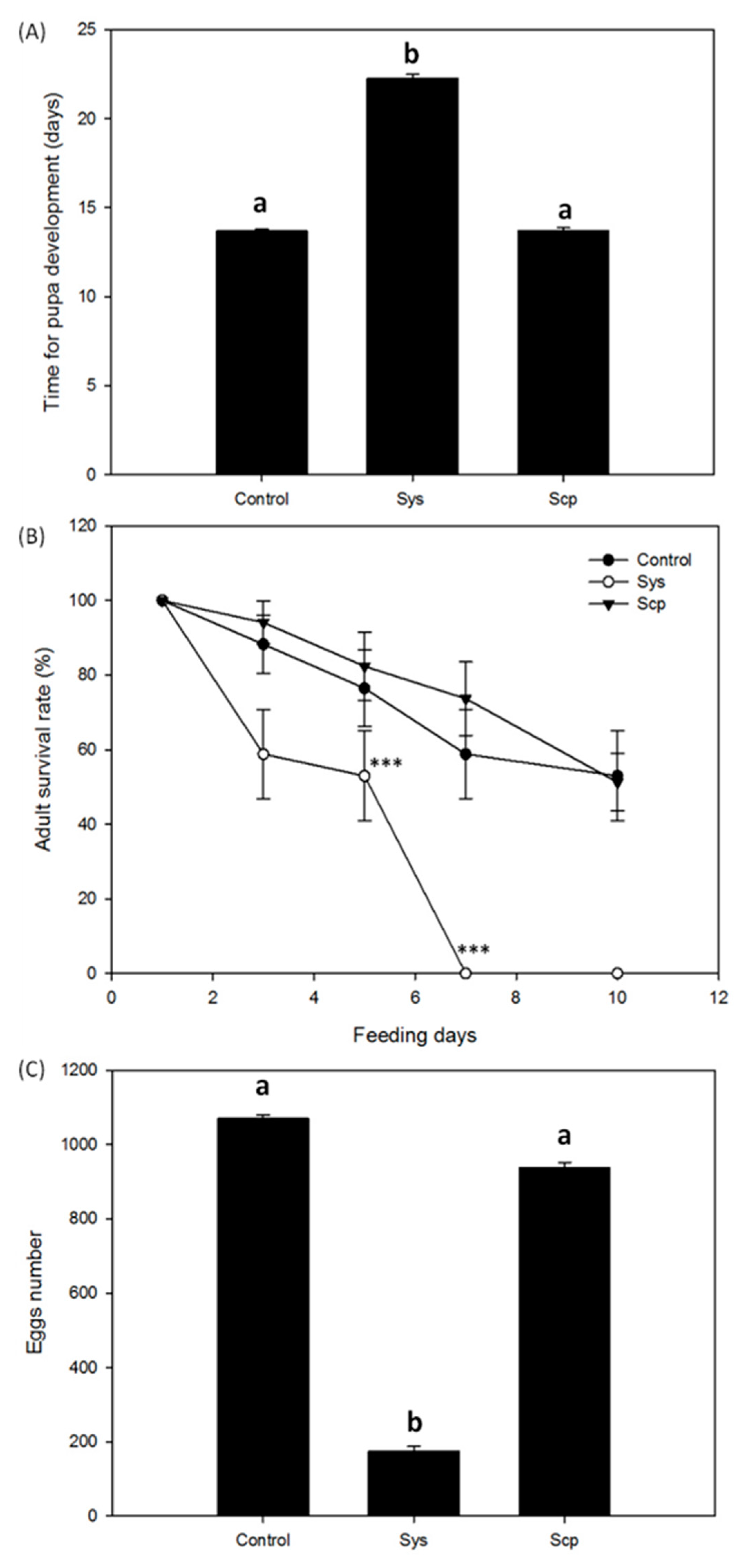
2.1. Sys Supply Promotes Direct Defenses against Spodoptera littoralis
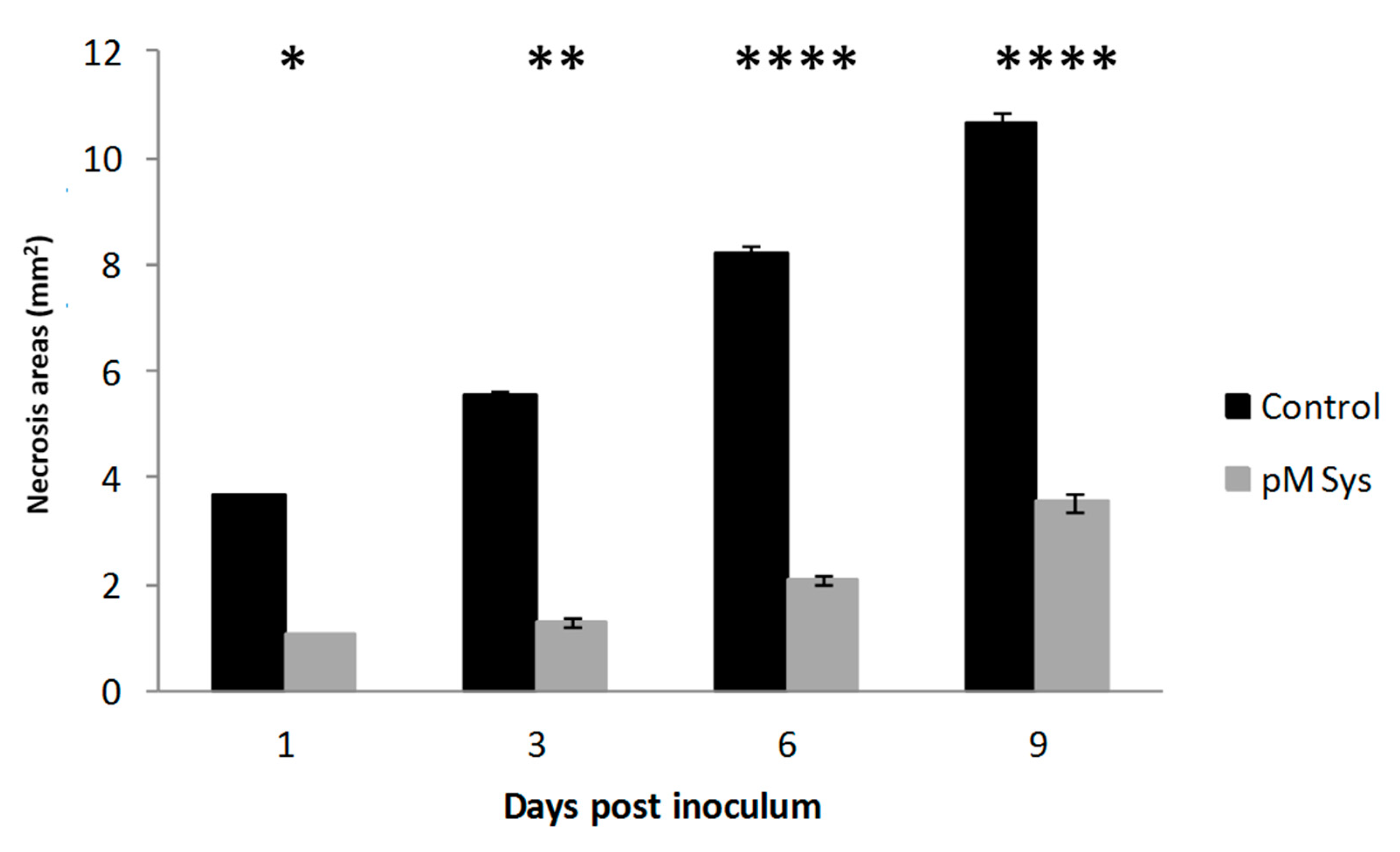
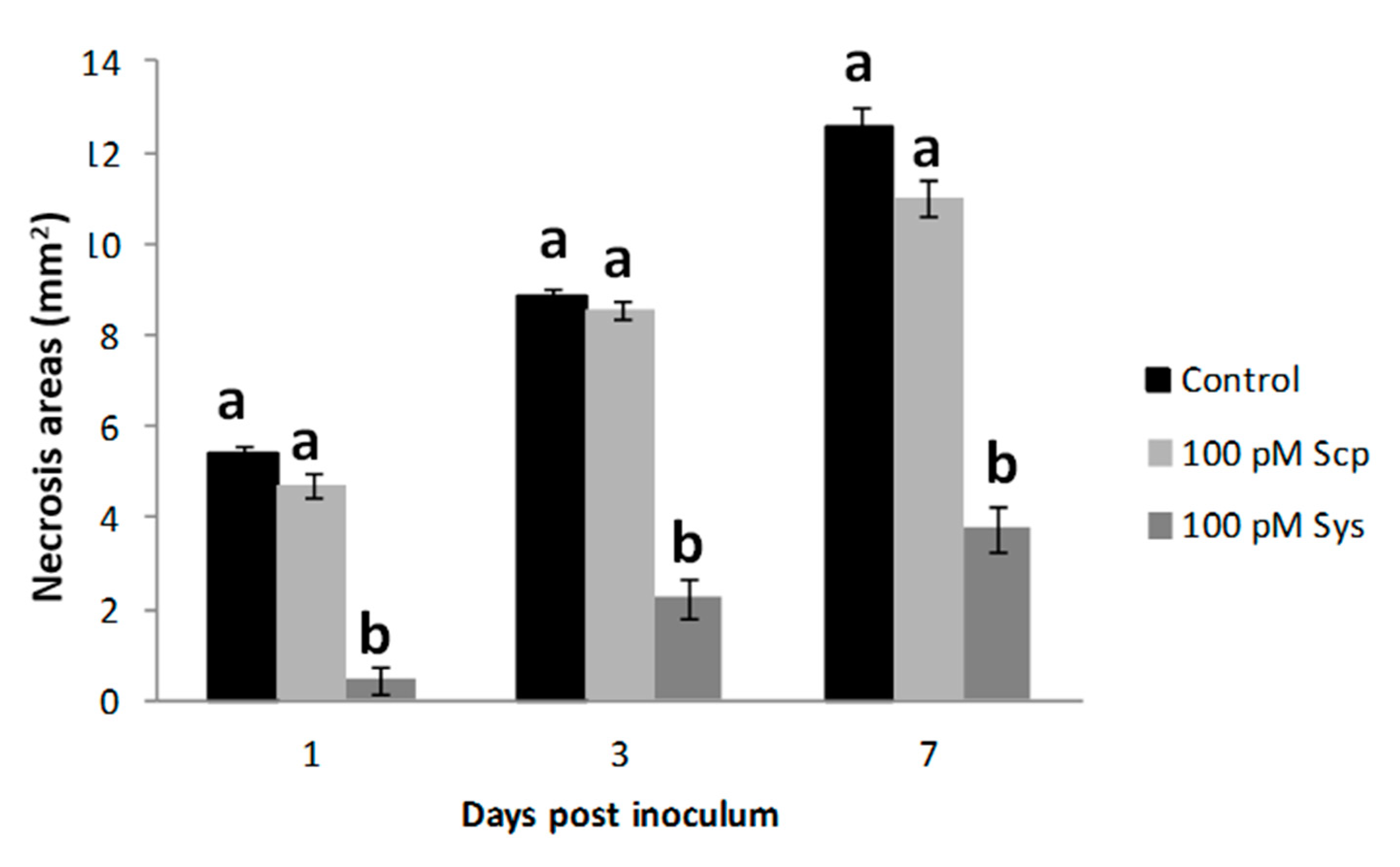
2.2. Sys Supply Enhances Plant Tolerance against Botrytis Cinerea
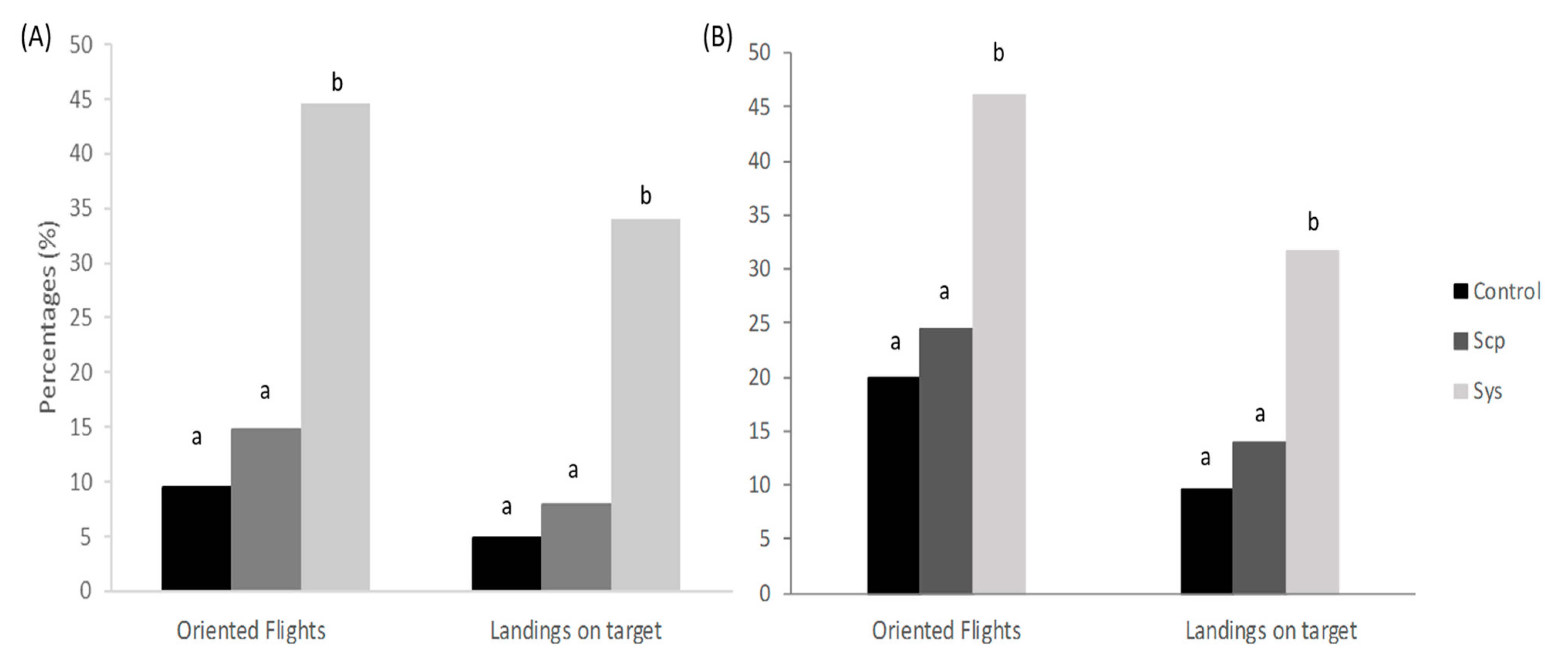
2.3. Sys Supply Promotes Indirect Defenses by Increasing the Emission of VOCs
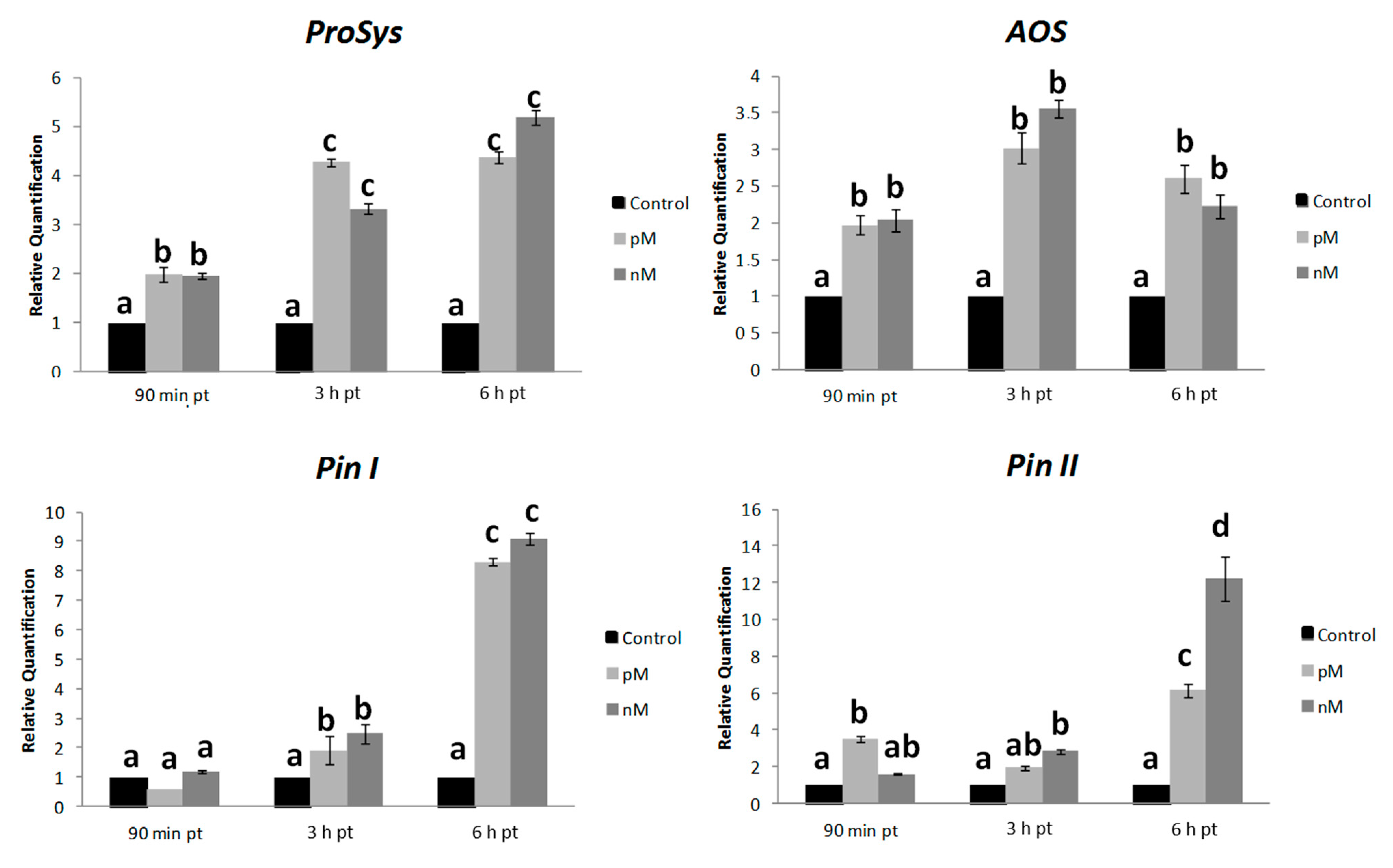
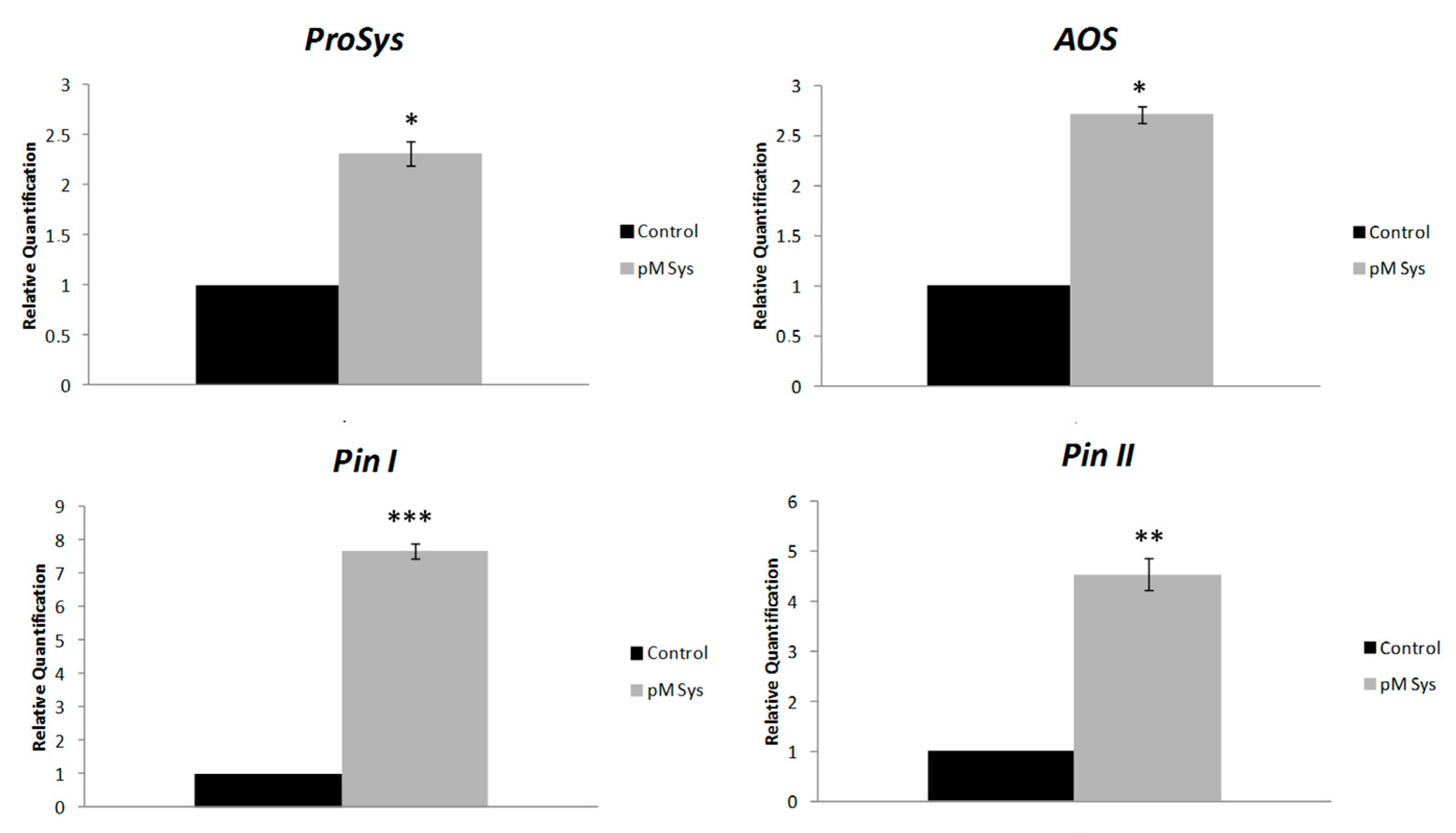
2.4. Systemin Supply on Leaves of Intact Plants Induce the Expression of Defense Genes
3. Materials and Methods
3.1. Plant Materials
3.2. Bioassay with Spodoptera Littoralis
3.3. Bioassay with Botrytis Cinerea
3.4. Aphidius Ervi Flight Behavior
3.5. Volatile Organic Compounds (VOCs) Collection and Analysis
3.6. Calibration of Methyl-Jasmonate Standard
3.7. Gene Expression Analysis
3.8. Statistical Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Ethical Statement
References
- Dayan, F.E.; Cantrell, C.L.; Duke, S.O. Natural products in crop protection. Bioorg. Med. Chem. 2009, 17, 4022–4034. [Google Scholar] [CrossRef] [PubMed]
- Pennacchio, F.; Giordana, B.; Rao, R. Applications of Parasitoid Virus and Venom Research in Agriculture. In Parasitoid Viruses, Symbionts and Pathogens; Beckage, N., Drezen, J., Eds.; Academic Press: London, UK, 2012; pp. 269–283. [Google Scholar]
- Xu, S.; Zhou, W.; Pottinger, S.; Baldwin, I.T. Herbivore associated elicitor-induced defenses are highly specific among closely related Nicotiana species. BMC Plant Biol. 2015, 15, 2. [Google Scholar] [CrossRef] [PubMed]
- Savatin, D.V.; Gramegna, G.; Modesti, V.; Cervone, F. Wounding in the plant tissue: The defense of a dangerous passage. Front. Plant Sci. 2014, 5, 470. [Google Scholar] [CrossRef] [PubMed]
- Albersheim, P.; Anderson, A.J. Proteins from plant cell walls inhibit polygalacturonases secreted by plant pathogens. Proc. Natl. Acad. Sci. USA 1971, 68, 1815–1819. [Google Scholar] [CrossRef]
- Chai, H.B.; Doke, N. Activation of the potential of potato leaf tissue to react hypersensitively to Phytophthora infestans by cystospore germination fluid and the enhancement of the potential by calcium ion. Physiol. Mol. Plant Pathol. 1987, 30, 27–37. [Google Scholar] [CrossRef]
- Pearce, G.; Strydom, D.; Johnson, S.; Ryan, C.A. A polypeptide from tomato leaves induces wound-inducible proteinase inhibitor proteins. Science 1991, 253, 895–897. [Google Scholar] [CrossRef]
- Chen, K.; Kirber, M.T.; Xiao, H.; Yang, Y.; Keaney, J.F. Regulation of ROS signal transduction by NADPH oxidase 4 localization. J. Cell Biol. 2008, 181, 1129–1139. [Google Scholar] [CrossRef] [Green Version]
- Bergey, D.R.; Orozco-Cárdenas, M.; De Moura, D.S.; Ryan, C.A. A wound- and systemin-inducible polygalacturonase in tomato leaves. Proc. Natl. Acad. Sci. USA 1999, 96, 1756–1760. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.A.; Dangl, J.L.; Jones, J.D. Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. [Google Scholar] [CrossRef]
- Huffaker, A.; Pearce, G.; Ryan, C.A. An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proc. Natl. Acad. Sci. USA 2006, 103, 10098–10103. [Google Scholar] [CrossRef]
- Albert, M. Peptides as triggers of plant defense. J. Exp. Bot. 2013, 64, 5269–5279. [Google Scholar] [CrossRef] [PubMed]
- Czyzewicz, N.; Yue, K.; Beeckman, T.; Smet, I.D. Message in a bottle: Small signalling peptide outputs during growth and development. J. Exp. Bot. 2013, 64, 5281–5296. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Barona, G.; Ryan, C.A.; Pearce, G. GmPep914, an eight-amino acid peptide isolated from soybean leaves, activates defense-related genes. Plant Physiol. 2011, 156, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.A. The systemin signalling pathway: Differential activation of plant defensive genes. Biochim. Biophys. Acta 2000, 1477, 112–121. [Google Scholar] [CrossRef]
- Scheer, J.M.; Ryan, C.A. The systemin receptor SR160 from Lycopersicon peruvianum is a member of the LRR receptor kinase family. Proc. Natl. Acad. Sci. USA 2002, 99, 9585–9590. [Google Scholar] [CrossRef]
- Wang, L.; Einig, E.; Almeida-Trapp, M.; Albert, M.; Fliegmann, J.; Mithöfer, A.; Kalbacher, H.; Felix, G. The systemin receptor SYR1 enhances resistance of tomato against herbivorous insects. Nat. Plants 2018, 4, 152–156. [Google Scholar] [CrossRef]
- Schilmiller, A.L.; Howe, G. Systemic signaling in the wound response. Curr. Opin. Plant Biol. 2005, 8, 369–377. [Google Scholar] [CrossRef]
- Sun, J.; Jiang, H.; Li, C. Systemin/Jasmonate-Mediated Systemic Defense Signaling in Tomato. Mol. Plant 2011, 4, 607–615. [Google Scholar] [CrossRef]
- Beloshistov, R.E.; Dreizler, K.; Galiullina, R.A.; Tuzhikov, A.I.; Serebryakova, M.V.; Reichardt, S.; Shaw, J.; Taliansky, M.E.; Pfannstiel, J.; Chichkova, N.V.; et al. Phytaspase-mediated precursor processing and maturation of the wound hormone systemin. New Phytol. 2017, 218, 1167–1178. [Google Scholar] [CrossRef]
- Scheer, J.M.; Pearce, G.; Ryan, C.A. Generation of systemin signaling in tobacco by transformation with the tomato systemin receptor kinase gene. Proc. Natl. Acad. Sci. USA 2003, 100, 10114–10117. [Google Scholar] [CrossRef] [Green Version]
- Pearce, G. Systemin, hydroxyproline-rich systemin and the induction of protease inhibitors. Curr. Protein Pept. Sci. 2011, 12, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, R.; krishna Koramutla, M.; Negi, M.; Pearce, G.; Ryan, C.A. Hydroxyproline-rich glycopeptide signals in potato elicit signalling associated with defense against insects and pathogens. Plant Sci. 2013, 207, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Narváez-Vásquez, J.; Orozco-Cárdenas, M.L.; Ryan, C.A. Systemic wound signaling in tomato leaves is cooperatively regulated by systemin and hydroxyproline-rich glycopeptide signals. Plant Mol. Biol. 2007, 65, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Pearce, G.; Moura, D.S.; Stratmann, J.; Ryan, C.A. Production of multiple plant hormones from a single polyprotein precursor. Nature 2001, 411, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.A.; Carroll, M.J.; LeClere, S.; Phipps, S.M.; Meredith, J.; Chourey, P.S.; Alborn, H.T.; Teal, P.E. Fragments of ATP synthase mediate plant perception of insect attack. Proc. Natl. Acad. Sci. USA 2006, 103, 8894–8899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huffaker, A.; Dafoe, N.J.; Schmelz, E.A. ZmPep1, an ortholog of Arabidopsis elicitor peptide 1, regulates maize innate immunity and enhances disease resistance. Plant Physiol. 2011, 155, 1325–1338. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Pearce, G.; Ryan, C.A. The cell surface leucine-rich repeat receptor for AtPep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc. Natl. Acad. Sci. USA 2006, 103, 10104–10109. [Google Scholar] [CrossRef]
- McGurl, B.; Pearce, G.; Orozco-Cardenas, M.; Ryan, C.A. Structure, expression, and antisense inhibition of the systemin precursor gene. Science 1992, 255, 1570–1573. [Google Scholar] [CrossRef]
- Coppola, M.; Corrado, G.; Coppola, V.; Cascone, P.; Martinelli, R.; Digilio, M.C.; Pennacchio, F.; Rao, R. Prosystemin Overexpression in Tomato Enhances Resistance to Different Biotic Stresses by Activating Genes of Multiple Signaling Pathways. Plant Mol. Biol. Rep. 2015, 33, 1270–1285. [Google Scholar] [CrossRef]
- Ahmad, F.H.; Wu, X.; Stintzi, A.; Schaller, A.; Schulze, W.X. The systemin signaling cascade as derived from time course analyses of the systemin-responsive phosphoproteome. Mol. Cell Proteom. 2019, 18, 1526–1542. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, P.; Zhao, J.; Jiang, H.; Wang, H.; Zhu, Y.; Botella, M.A.; Samaj, J.; Li, C.; Lin, J. Expression of tomato prosystemin gene in Arabidopsis reveals systemic translocation of its mRNA and confers necrotrophic fungal resistance. New Phytol. 2018, 217, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wilkerson, C.G.; Kuchar, J.A.; Phinney, B.S.; Howe, G.A. Jasmonate-inducible plant enzymes degrade essential amino acids in the herbivore midgut. Proc. Natl. Acad. Sci. USA 2005, 102, 19237–19242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, C.A.; Pearce, G. Systemin: A polypeptide signal for plant defensive genes. Annu. Rev. Cell Dev. Biol. 1998, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- McGurl, B.; Orozco-Cárdenas, M.; Pearce, G.; Ryan, C.A. Overexpression of the prosystemin gene in transgenic tomato plants generates a systemic signal that constitutively induces proteinase inhibitor synthesis. Proc. Natl. Acad. Sci. USA 1994, 91, 9799–9802. [Google Scholar] [CrossRef]
- Corrado, G.; Sasso, R.; Pasquariello, M.; Iodice, L.; Carretta, A.; Cascone, P.; Ariati, L.; Digilio, M.C.; Guerrieri, E.; Rao, R. Systemin regulates both systemic and volatile signaling in tomato plants. J. Chem. Ecol. 2007, 33, 669–681. [Google Scholar] [CrossRef]
- Degenhardt, D.C.; Refi-Hind, S.; Stratmann, J.W.; Lincoln, D.E. Systemin and jasmonic acid regulate constitutive and herbivore-induced systemic volatile emissions in tomato, Solanum lycopersicum. Phytochemistry 2010, 71, 2024–2037. [Google Scholar] [CrossRef]
- Orsini, F.; Cascone, P.; De Pascale, S.; Barbieri, G.; Corrado, G.; Rao, R.; Maggio, A. Systemin-dependent salinity tolerance in tomato: Evidence of specific convergence of abiotic and biotic stress responses. Physiol. Plant 2010, 138, 10–21. [Google Scholar] [CrossRef]
- Corrado, G.; Agrelli, D.; Rocco, M.; Basile, B.; Marra, M.; Rao, R. Systemin-inducible defense against pests is costly in tomato. Biol. Plant 2011, 552, 305–311. [Google Scholar] [CrossRef]
- Coppola, M.; Cascone, P.; Madonna, V.; Di Lelio, I.; Esposito, F.; Avitabile, C.; Romanelli, A.; Guerrieri, E.; Vitiello, A.; Pennacchio, F.; et al. Plant-to-plant communication triggered by systemin primes anti-herbivore resistance in tomato. Sci. Rep. 2017, 7, 15522. [Google Scholar] [CrossRef]
- Romanelli, A.; Moggio, L.; Montella, R.C.; Campiglia, P.; Iannaccone, M.; Capuano, F.; Pedone, C.; Capparelli, R. Peptides from Royal Jelly: Studies on the antimicrobial activity of jelleins, jelleins analogs and synergy with temporins. J. Pept. Sci. 2011, 17, 348–352. [Google Scholar] [CrossRef]
- Czyzewicz, N.; Stes, E.; De Smet, I. Tips and Tricks for Exogenous Application of Synthetic Post-translationally Modified Peptides to Plants. In Plant Hormones: Methods and Protocols; Kleine-Vehn, J., Sauer, M., Eds.; Humana Press: New York, NY, USA, 2017; pp. 19–28. [Google Scholar]
- Sannino, L.; Espinosa, B.; Balbiani, A. Lepidotteri Delle Ortive e del Tabacco; Calderini Edagricole: Bologna, Italy, 2001; p. 323. [Google Scholar]
- Guerrieri, E.; Pennacchio, F.; Tremblay, E. Flight behaviour of the aphid parasitoid Aphidius ervi (Hymenoptera: Braconidae) in response to plant and host volatiles. Eur. J. Entomol. 1993, 90, 415–421. [Google Scholar]
- Corrado, G.; Alagna, F.; Rocco, M.; Renzone, G.; Varricchio, P.; Coppola, V.; Coppola, M.; Garonna, A.; Baldoni, L.; Scaloni, A.; et al. Molecular interactions between the olive and the fruit fly Bactroceraoleae. BMC Plant Biol. 2012, 12, 86. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Marum, L.; Miguel, A.; Ricardo, C.P.; Miguel, C. Reference Gene Selection for Quantitative Real-time PCR Normalization in Quercussuber. PLoS ONE 2012, 7, e35113. [Google Scholar]
- Müller, O.A.; Grau, J.; Thieme, S.; Prochaska, S.; Adlung, N.; Sorgatz, A.; Bonas, U. Genome-Wide Identification and Validation of Reference Genes in Infected Tomato Leaves for Quantitative RT-PCR Analyses. PLoS ONE 2015, 10, e0136499. [Google Scholar] [CrossRef] [PubMed]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman and Co.: New York, NY, USA, 1995; p. 915. [Google Scholar]
- Dıaz, J.; ten Have, A.; van Kan, J.A. The role of ethylene and wound signaling in resistance of tomato to Botrytis cinerea. Plant Physiol. 2002, 129, 1341–1351. [Google Scholar] [CrossRef]
- El Oirdi, M.; El Rahman, T.A.; Rigano, L.; El Hadrami, A.; Rodriguez, M.C.; Daayf, F.; Vojnov, A.; Bouarab, K. Botrytis cinerea manipulates the antagonistic effects between immune pathways to promote disease development in tomato. Plant Cell 2011, 23, 2405–2421. [Google Scholar] [CrossRef]
- Mucha, P.; Ruczynski, J.; Dobkowski, M.; Backtrog, E.; Rekowski, P. Capillary electrophoresis study of systemin peptides spreading in tomato plant. Electrophoresis 2019, 40, 336–342. [Google Scholar] [CrossRef]
- Chauvin, A.; Caldelari, D.; Wolfender, J.L.; Farmer, E.E. Four 13-lipoxygenases contribute to rapid jasmonate synthesis in wounded Arabidopsis thaliana leaves: A role for lipoxygenase 6 in responses to long-distance wound signals. New Phytol. 2013, 197, 566–575. [Google Scholar] [CrossRef]
- AbuQamar, S.; Chai, M.F.; Luo, H.; Song, F.; Mengiste, T. Tomato protein kinase 1b mediates signaling of plant responses to necrotrophic fungi and insect herbivory. Plant Cell 2008, 20, 1964–1983. [Google Scholar] [CrossRef]
- Xu, S.; Liao, C.J.; Jaiswal, N.; Lee, S.; Yun, D.J.; Lee, S.Y.; Garvey, M.; Kaplan, I.; Mengiste, T. Tomato PEPR1 ORTHOLOG RECEPTOR-LIKE KINASE1 Regulates Responses to Systemin, Necrotrophic Fungi, and Insect Herbivory. Plant Cell 2018, 30, 2214–2229. [Google Scholar] [CrossRef] [Green Version]
- de la Noval, B.; Pérez, E.; Martínez, B.; León, O.; Martínez-Gallardo, N.; Délano-Frier, J. Exogenous systemin has a contrasting effect on disease resistance in mycorrhizal tomato (Solanum lycopersicum) plants infected with necrotrophic or hemibiotrophic pathogens. Mycorrhiza 2007, 17, 449–460. [Google Scholar] [CrossRef]
- Pastor, V.; Sánchez-Bel, P.; Gamir, J.; Pozo, M.J.; Flors, V. Accurate and easy method for systemin quantification and examining metabolic changes under different endogenous levels. Plant Methods 2018, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Huffaker, A.; Bryan, A.C.; Tax, F.E.; Ryan, C.A. PEPR2 is a second receptor for the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis. Plant Cell 2010, 22, 508–522. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, C.; Robatzek, S.; Navarro, L.; Oakeley, E.J.; Jones, J.D.; Felix, G.; Boller, T. Bacterial disease resistance in Arabidopsis through flagellin perception. Nature 2004, 428, 764. [Google Scholar] [CrossRef] [PubMed]
- Holton, N.; Caño-Delgado, A.; Harrison, K.; Montoya, T.; Chory, J.; Bishop, G.J. Tomato BRASSINOSTEROID INSENSITIVE1 is required for systemin-induced root elongation in Solanum pimpinellifolium but is not essential for wound signaling. Plant Cell 2007, 19, 1709–1717. [Google Scholar] [CrossRef]
- Webster, B.; Bruce, T.; Dufour, S.; Birkemeyer, C.; Birkett, M.; Hardie, J.; Pickett, J. Identification of volatile compounds used in host location by the black bean aphid, Aphis fabae. J. Chem. Ecol. 2008, 34, 1153–1161. [Google Scholar] [CrossRef]
- Song, B.; Liang, Y.; Liu, S.; Zhang, L.; Tang, G.; Ma, T.; Yao, Y. Behavioral responses of Aphis citricola (Hemiptera: Aphididae) and its natural enemy Harmonia axyridis (Coleoptera: Coccinellidae) to non-host plant volatiles. Fla. Entomol. 2017, 100, 411–421. [Google Scholar] [CrossRef]
- Engelberth, J.; Alborn, H.T.; Schmelz, E.A.; Tumlinson, J.H. Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. USA 2004, 101, 1781–1785. [Google Scholar] [CrossRef] [Green Version]
- Sasso, R.; Iodice, L.; Woodcock, C.M.; Pickett, J.A.; Guerrieri, E. Electrophysiological and behavioural responses of Aphidius ervi (Hymenoptera: Braconidae) to tomato plant volatiles. Chemoecology 2009, 19, 195–201. [Google Scholar] [CrossRef]
- Constabel, C.P.; Yip, L.; Ryan, C.A. Prosystemin from potato, black nightshade, and bell pepper: Primary structure and biological activity of predicted systemin polypeptides. Plant Mol. Biol. 1998, 36, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Pearce, G.; Veyrat, N.; Erb, M.; Turlings, T.C.; Sartor, R.; Shen, Z.; Briggs, S.P.; Vaughan, M.M.; Alborn, H.T.; et al. Plant elicitor peptides are conserved signals regulating direct and indirect anti herbivore defense. Proc. Natl. Acad. Sci. USA 2013, 110, 5707–5712. [Google Scholar] [CrossRef] [PubMed]
- Trivilin, A.P.; Hartke, S.; Moraes, M.G. Components of different signalling pathways regulated by a new orthologue of AtPROPEP1 in tomato following infection by pathogens. Plant Pathol. 2014, 63, 1110–1118. [Google Scholar] [CrossRef]
- Lori, M.; Van Verk, M.C.; Hander, T.; Schatowitz, H.; Klauser, D.; Flury, P.; Gehring, C.A.; Boller, T.; Bartels, S. Evolutionary divergence of the plant elicitor peptides (Peps) and their receptors: Interfamily incompatibility of perception but compatibility of downstream signalling. J. Exp. Bot. 2015, 66, 5315–5325. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Betz, F.S. Messenger®: An Environmentally Sound Solution for Crop Production and Protection. ACS Symp. Ser. 2007, 957, 195–211. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (PPb) | |||||
|---|---|---|---|---|---|
| Name | Molecular Formula | Molecular Weight g/mol | Control | Sys | Scp |
| Benzaldehyde * | C7H6O | 106.124 | 1.54 × 106 ± 1.2 × 105 | 3.09 × 106 ± 2.8 × 105 | 1.81 × 106 ± 1.18 × 105 |
| Ethylbenzene, p-Xylene * | C8H10 | 106.168 | 1.53 × 106 ± 1.43 × 105 | 3.06 × 106 ± 2.7 × 105 | 1.81 × 106 ± 1.16 × 105 |
| β-Ocimene * | C10H16 | 136.238 | 1.01 × 106 ± 8.1 × 104 | 1.22 × 107 ± 1.46 × 105 | 1.19 × 106 ± 0.97 × 105 |
| α-pinene * | C10H16 | 136.238 | 1.01 × 106 ± 8.1 × 104 | 1.22 × 107 ± 1.46 × 105 | 1.19 × 106 ± 0.97 × 105 |
| limonene * | C10H16 | 136.238 | 1.01 × 106 ± 8.1 × 104 | 1.22 × 107 ± 1.46 × 105 | 1.19 × 106 ± 0.97 × 105 |
| Methyl Jasmonate * | C13H20O3 | 224.300 | 6.4 × 105 ± 4.2 × 104 | 1.16 × 106 ± 5.17 × 104 | 5.68 × 105 ± 4.02 × 104 |
| β-caryophyllene * | C15H24 | 204.357 | 1.75 × 105 ± 2.5 × 104 | 0.95 × 106 ± 7.6 × 104 | 1.16 × 105 ± 1.37 × 104 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coppola, M.; Lelio, I.D.; Romanelli, A.; Gualtieri, L.; Molisso, D.; Ruocco, M.; Avitabile, C.; Natale, R.; Cascone, P.; Guerrieri, E.; et al. Tomato Plants Treated with Systemin Peptide Show Enhanced Levels of Direct and Indirect Defense Associated with Increased Expression of Defense-Related Genes. Plants 2019, 8, 395. https://doi.org/10.3390/plants8100395
Coppola M, Lelio ID, Romanelli A, Gualtieri L, Molisso D, Ruocco M, Avitabile C, Natale R, Cascone P, Guerrieri E, et al. Tomato Plants Treated with Systemin Peptide Show Enhanced Levels of Direct and Indirect Defense Associated with Increased Expression of Defense-Related Genes. Plants. 2019; 8(10):395. https://doi.org/10.3390/plants8100395
Chicago/Turabian StyleCoppola, Mariangela, Ilaria Di Lelio, Alessandra Romanelli, Liberata Gualtieri, Donata Molisso, Michelina Ruocco, Concetta Avitabile, Roberto Natale, Pasquale Cascone, Emilio Guerrieri, and et al. 2019. "Tomato Plants Treated with Systemin Peptide Show Enhanced Levels of Direct and Indirect Defense Associated with Increased Expression of Defense-Related Genes" Plants 8, no. 10: 395. https://doi.org/10.3390/plants8100395