
Urea Coated with Polyaspartic Acid-Chitosan Increases Foxtail Millet (Setaria italica L. Beauv.) Grain Yield by Improving Nitrogen Metabolism

Abstract
:1. Introduction
2. Results
2.1. Grain Yield and Yield Components
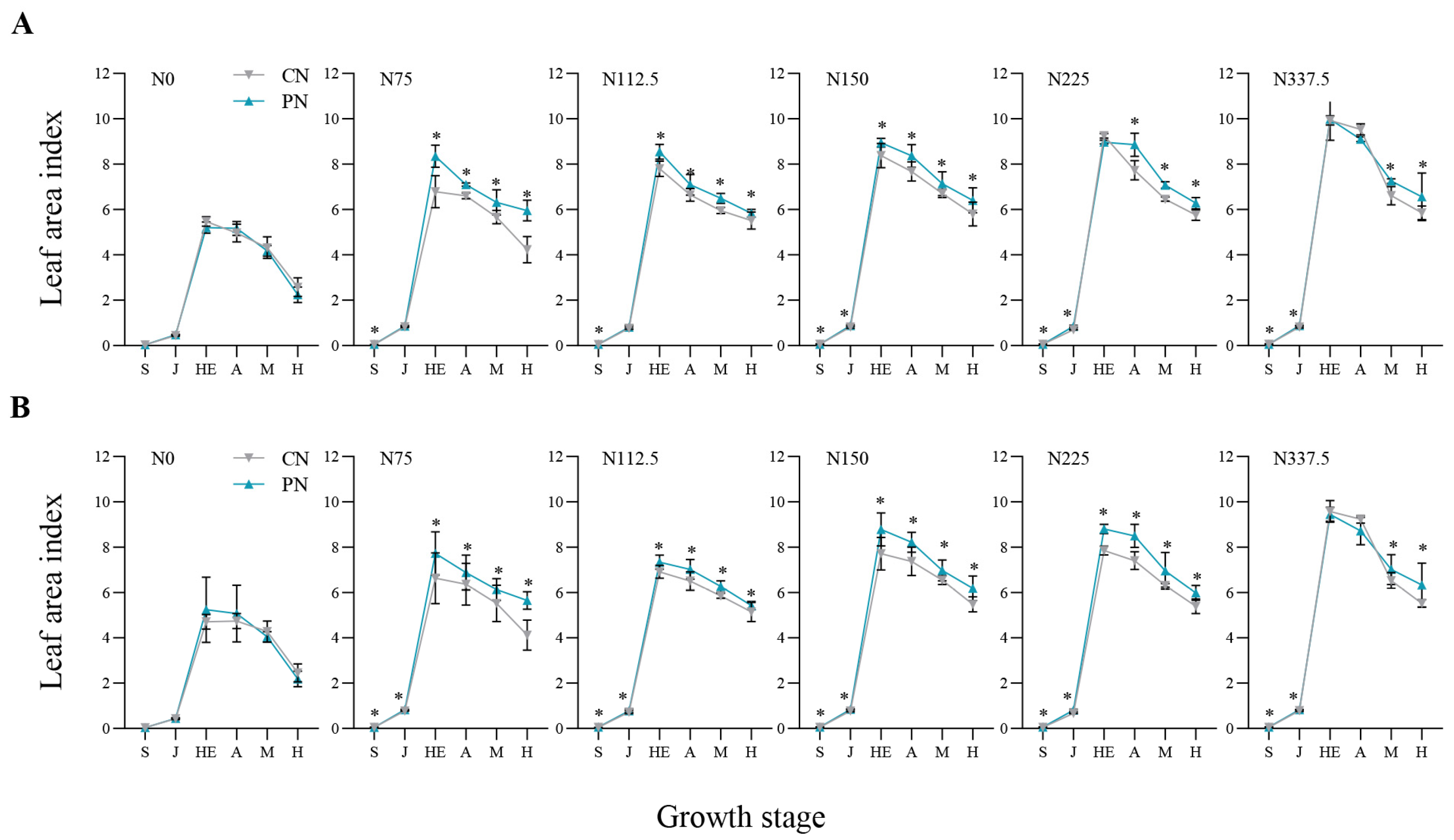
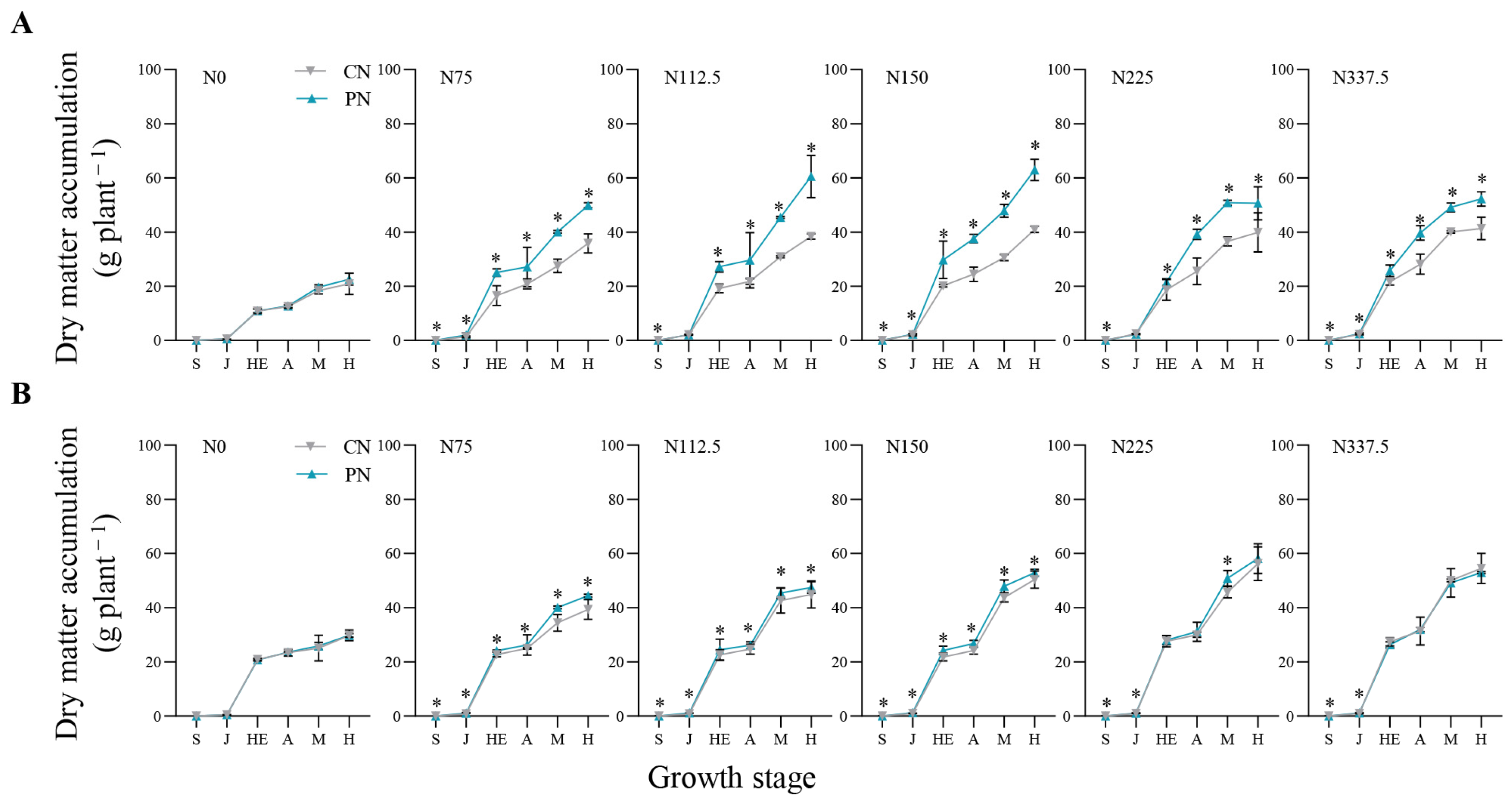
2.2. Leaf Area Index and Dry Matter Accumulation
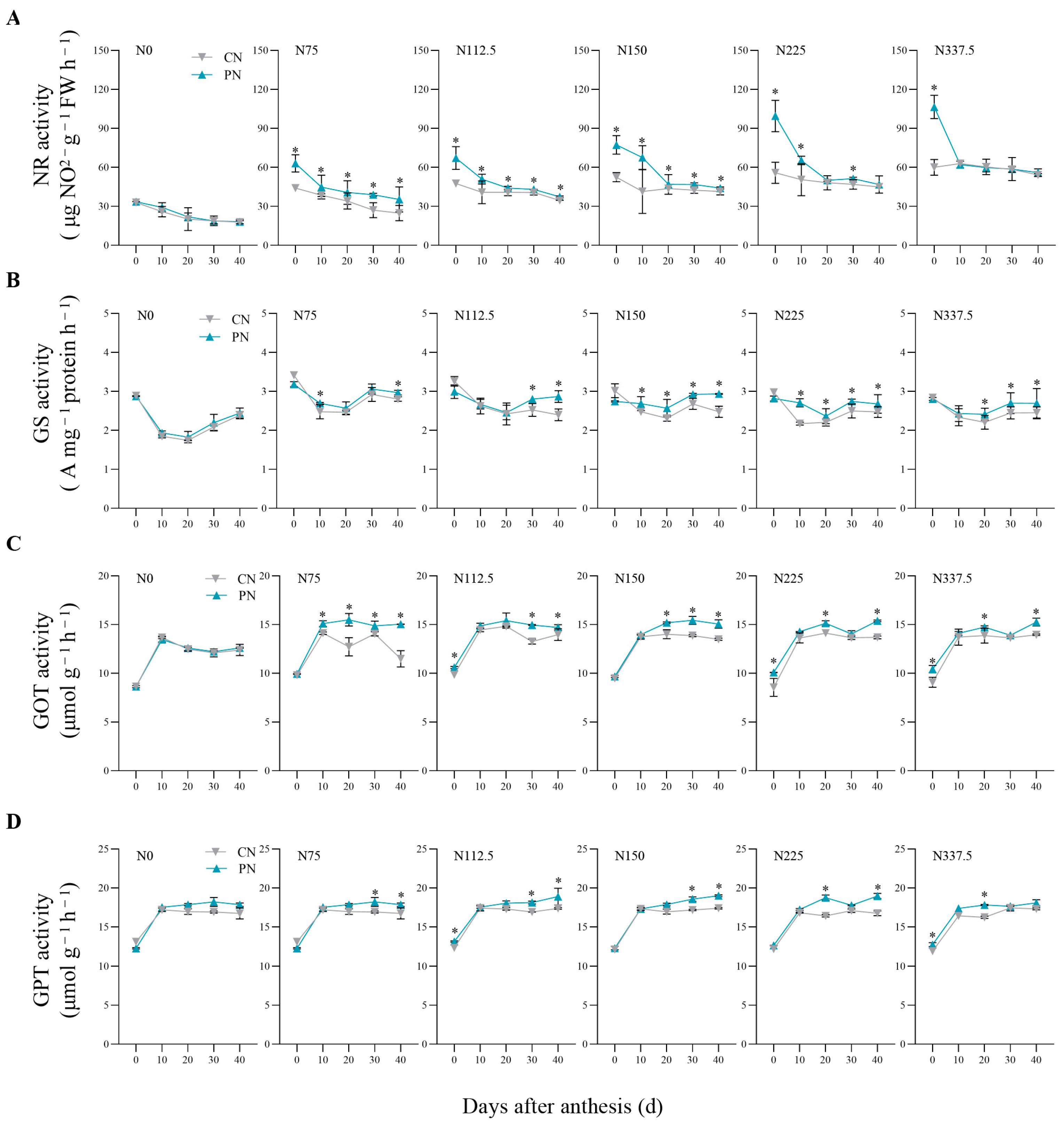
2.3. Activities of NR, GS, GOT, and GPT
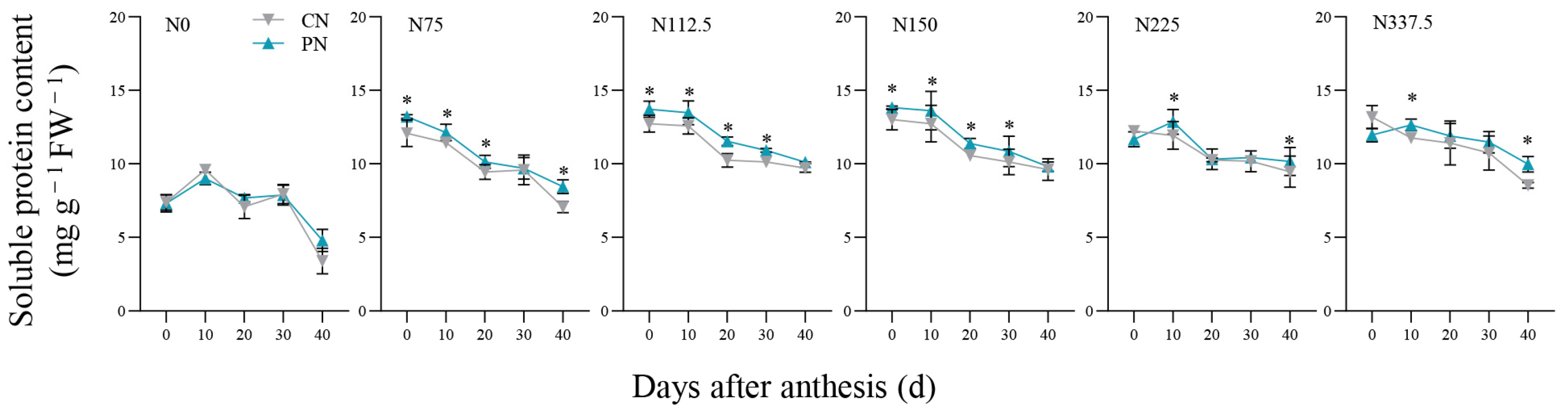
2.4. Soluble Protein Content
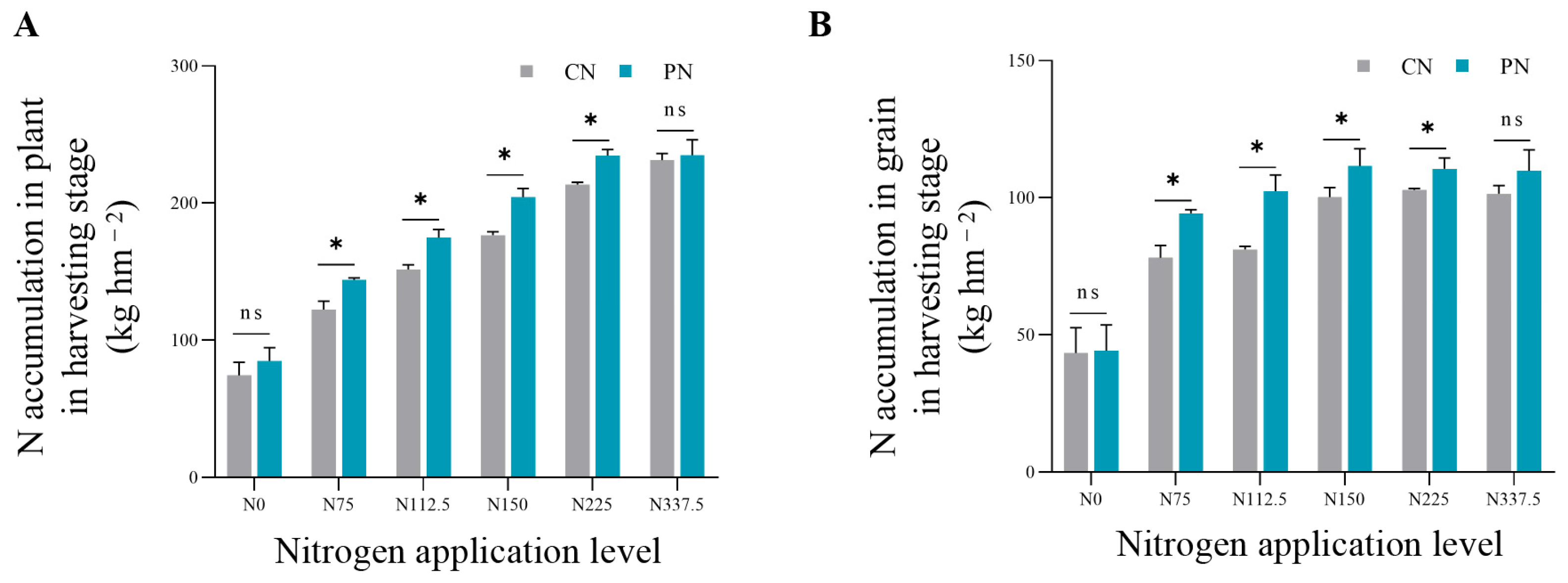
2.5. Total Nitrogen Accumulation in the Plant and Grain
2.6. Nitrogen Use Efficiency
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Experimental Design
4.2. Sampling and Measurement
4.2.1. Yield and Yield Components
4.2.2. Leaf Area Index and Dry Matter Accumulation
4.2.3. Determination of Enzyme Activities Related to Nitrogen Metabolism
4.2.4. Net Assimilation Rate
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hou, Y.L.; Zhang, J.P.; Liu, K.B.; Wu, N.Q.; Li, Y.M.; Zhou, K.S.; Ye, M.L.; Zhang, T.Y.; Zhang, H.J.; Yang, X.Y.; et al. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10,000 years ago. Proc. Natl. Acad. Sci. USA 2009, 106, 7367–7372. [Google Scholar] [CrossRef]
- Diao, X.M. Production and genetic improvement of minor cereals in China. Crop J. 2017, 5, 103–114. [Google Scholar] [CrossRef]
- Consuelo, M.P.; Bienvenido, O.J.; Samuel, P.L.; Jovencio, M.A.; Kenneth, G.C. Effects of late nitrogen fertilizer application on head rice yield, protein content, and grain quality of rice. Cereal Chem. 1996, 73, 556–560. [Google Scholar] [CrossRef]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Oliveira, R.M.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Du, W.Y.; Zhang, Y.X.; Wan, Z.S.; Li, H.W.; Xia, H.Y.; Fan, S.J.; Kong, L.A. Metabolic analysis reveals temporal changes in C/N metabolites and senescence of flag leaf in wheat during grain filling. Acta Physiol. Plant 2021, 43, 104. [Google Scholar] [CrossRef]
- Hossein, A.; Huong, D.N.T.; Peter, J.L.; Idriss, B.; Hang, T.T. Poly(aspartic acid) in biomedical applications: From polymerization, modification, properties, degradation, and biocompatibility to applications. ACS Biomater. Sci. Eng. 2021, 7, 2083–2105. [Google Scholar] [CrossRef]
- Hu, M.M.; Dou, Q.H.; Cui, X.M.; Lou, Y.H.; Zhuge, Y.P. Polyaspartic acid mediates the absorption and translocation of mineral elements in tomato seedlings under combined copper and cadmium stress. J. Integr. Agric. 2019, 18, 1130–1137. [Google Scholar] [CrossRef]
- Yan, P.; Fang, M.Y.; Lu, L.; Ren, L.; Dong, X.R.; Dong, Z.Q. Effect of urea coated with polyaspartic acid on the yield and nitrogen use efficiency of sorghum (Sorghum bicolor (L.) Moench.). Plants 2022, 11, 1724. [Google Scholar] [CrossRef]
- Román-Doval, R.; Torres-Arellanes, S.P.; Tenorio-Barajas, A.Y.; Gómez-Sánchez, A.; Valencia-Lazcano, A.A. Chitosan: Properties and its application in agriculture in context of molecular weight. Polymers 2023, 15, 2876. [Google Scholar] [CrossRef]
- Howard, K.; Charles, H.T.; Molly, S.S. Chitosan implants in the rat spinal cord: Biocompatibility and biodegradation. J. Biomed. Mater. Res. Part A 2011, 97A, 395–404. [Google Scholar] [CrossRef]
- Mi, F.L.; Tan, Y.C.; Liang, H.F.; Sung, H.W. In vivo biocompatibility and degradability of a novel injectable-chitosan-based implant. Biomaterials 2002, 23, 181–191. [Google Scholar] [CrossRef]
- Shi, C.M.; Zhu, Y.; Ran, X.Z.; Wang, M.; Su, Y.P.; Cheng, T.M. Therapeutic potential of chitosan and its derivatives in regenerative medicine. J. Surg. Res. 2006, 133, 185–192. [Google Scholar] [CrossRef]
- Lin, X.Z.; Gong, X.C.; Ruan, Q.C.; Xu, W.; Zhang, C.J.; Zhao, K. Antimicrobial application of chitosan derivatives and their nanocomposites. Curr. Med. Chem. 2023, 30, 1736–1755. [Google Scholar] [CrossRef]
- Ahmed, H.H.A.; Mohamed, R.A.N.; Hesham, A.A.; Amira, F.E. Effect of preharvest chitosan foliar application on growth, yield and chemical composition of Washington navel orange trees grown in two different regions. Afr. J. Biochem. Res. 2016, 10, 59–69. [Google Scholar] [CrossRef]
- Phothi, R.; Theerakarunwong, C.D. Effect of chitosan on physiology, photosynthesis and biomass of rice (Oryza sativa L.) under elevated ozone. Aust. J. Crop Sci. 2017, 11, 624–630. [Google Scholar] [CrossRef]
- Wu, L.; Liu, M.Z. Preparation and properties of chitosan-coated NPK compound fertilizer with controlled-release and water-retention. Carbohydr. Polym. 2008, 72, 240–247. [Google Scholar] [CrossRef]
- Wu, W.; Zhao, J. Advances on plants’ nitrogen assimilation and utilization. Chin. Agric. Sci. Bull. 2010, 26, 75–78. [Google Scholar]
- Liu, M.; Ma, Z.Q.; Liang, Q.; Zhang, Y.; Yang, Y.A.; Hou, H.P.; Wu, X.D.; Ge, J.Z. Spring wheat–summer maize annual crop system grain yield and nitrogen utilization response to nitrogen application rate in the thermal–resource–limited region of the North China Plain. Agronomy 2023, 13, 155. [Google Scholar] [CrossRef]
- Berry, P.M.; Sterling, M.; Spink, J.H.; Baker, C.J.; Sylvester-Bradley, R.; Mooney, S.J.; Tams, A.R.; Ennos, A.R. Understanding and reducing lodging in cereals. Adv. Agron. 2004, 84, 217–271. [Google Scholar] [CrossRef]
- Zhang, A.Y.; Guo, E.H.; Wang, J.; Fan, H.P.; Li, Y.H.; Wang, L.X.; Wang, X.Q.; Cheng, L.P. Effect of nitrogen application rate on agronomic, photosynthetic characteristics and yield of spring foxtail millet. Sci. Agric. Sin. 2015, 48, 2939–2951. [Google Scholar] [CrossRef]
- Deng, F.; Wang, L.; Mei, X.F.; Li, S.X.; Pu, S.L.; Li, Q.P.; Ren, W.J. Polyaspartic acid (PASP)-urea and optimised nitrogen management increase the grain nitrogen concentration of rice. Sci. Rep. 2019, 9, 313. [Google Scholar] [CrossRef]
- Cai, J.; Jiang, D.; Wollenweber, B.; Dai, T.; Cao, W. Effects of nitrogen application rate on dry matter redistribution, grain yield, nitrogen use efficiency and photosynthesis in malting barley. Acta Agric. Scand. Sect. B 2012, 62, 410–419. [Google Scholar] [CrossRef]
- Liu, Q.Y.; Li, M.; Ji, X.; Liu, J.; Wang, F.J.; Wei, Y.F. Characteristics of grain yield, dry matter production and nitrogen uptake and transport of rice varieties with different grain protein content. Agronomy 2022, 12, 2866. [Google Scholar] [CrossRef]
- Xu, G.H.; Fan, X.R.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- Ma, Z.B.; Dong, X.R.; Fang, M.Y.; Wang, Q.; Yan, P.; Wang, Q.Y.; Lu, L.; Dong, Z.Q. Effects of basic application of chlorocholine chloride combined with nitrogen fertilizer on nitrogen use of summer maize in North China Plain. Chin. J. Appl. Ecol. 2021, 32, 931–941. [Google Scholar] [CrossRef]
- Sylvie, F.M.; Céline, M.; Akira, S.; Marie, H.V.; Bertrand, H.; Christine, H.F. Glutamine and α-ketoglutarate are metabolite signals involved in nitrate reductase gene transcription in untransformed and transformed tobacco plants deficient in ferredoxin-glutamine-α-ketoglutarate aminotransferase. Planta 2001, 213, 265–271. [Google Scholar] [CrossRef]
- Kenis, J.D.; Trippi, V.S. Involvement of oxidation, proteolysis and reductant availability in the regulation of in vivo nitrate reductase in attached oat leaves during growth and senescence. Plant Cell Physiol. 1987, 28, 1307–1312. [Google Scholar] [CrossRef]
- Bernard, S.M.; Habash, D.Z. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. New Phytol. 2009, 182, 608–620. [Google Scholar] [CrossRef]
- Liang, C.G.; Chen, L.P.; Wang, Y.; Liu, J.; Xu, G.L.; Li, T. High temperature at grain-filling stage affects nitrogen metabolism enzyme activities in grains and grain nutritional quality in rice. Rice Sci. 2011, 18, 210–216. [Google Scholar] [CrossRef]
- Wang, Y.C.; Gu, W.R.; Ye, L.F.; Sun, Y.; Li, L.J.; Zhang, H.; Li, J.; Wei, S. Physiological mechanisms of delaying leaf senescence in maize treated with compound mixtures of DCPTA and CCC. J. Northeast Agric. Univ. 2015, 22, 1–15. [Google Scholar] [CrossRef]
- Lu, L.; Dong, Z.Q.; Dong, X.R.; Jiao, L.; Li, G.Y.; Gao, J. Effects of ethylene-chlormequat-potassium on leaf nitrogen assimilation after anthesis and early senescence under different planting densities. Acta Agrono. Sin. 2015, 41, 1870–1879. [Google Scholar] [CrossRef]
- Liu, K.; Zhou, S.Q.; Li, S.Y.; Wang, J.; Wang, W.L.; Zhang, W.Y.; Zhang, H.; Gu, J.F.; Yang, J.C.; Liu, L.J. Differences and mechanisms of post-anthesis dry matter accumulation in rice varieties with different yield levels. Crop Environ. 2022, 1, 262–272. [Google Scholar] [CrossRef]
- Ning, H.F.; Liu, Z.H.; Wang, Q.S.; Lin, Z.M.; Chen, G.J.; Li, G.H.; Wang, S.H.; Ding, Y.F. Effect of nitrogen fertilizer application on grain phytic acid and protein concentrations in japonica rice and its variations with genotypes. J. Cereal. Sci. 2009, 50, 49–55. [Google Scholar] [CrossRef]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities for improvement in the nitrogen utilization of crop. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Y.; Tang, H.H.; Li, G.Y.; Dong, H.; Dong, X.R.; Xu, Y.L.; Dong, Z.Q. Polyaspartic acid improves maize (Zea mays L.) seedling nitrogen assimilation mainly by enhancing nitrate reductase activity. Agronomy 2018, 8, 188. [Google Scholar] [CrossRef]
- El-Tanahy, A.M.M.; Mahmoud, A.R.; Abde-Mouty, M.M.; Ali, A.H. Effect of chitosan doses and nitrogen sources on the growth, yield and seed quality of cowpea. Aust. J. Basic Appl. Sci. 2012, 6, 115–121. [Google Scholar]
- Zhang, X.Q.; Li, K.C.; Xing, R.E.; Liu, S.; Li, P.C. Metabolite profiling of wheat seedlings induced by chitosan: Revelation of the enhanced carbon and nitrogen metabolism. Front. Plant Sci. 2017, 8, 2017. [Google Scholar] [CrossRef] [PubMed]
- Mondal, M.M.A.; Malek, M.A.; Puteh, A.B.; Ismail, M.R.; Ashrafuzzaman, M.; Naher, L. Effect of foliar application of chitosan on growth and yield in okra. Aust. J. Crop Sci. 2012, 6, 918–921. [Google Scholar]
- Wu, L.H.; Jiang, S.H.; Tao, Q.N. Colorimetric determination of plant transaminase (GOT and GPT) activity and its application. Chin. J. Soil. Sci. 1998, 29, 41–43. [Google Scholar] [CrossRef]
- Marion, M.B. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kenneth, G.C.; Achim, D.; Daniel, T.W. Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 2002, 31, 132–140. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Nitrogen Application Level | Treatment | 1000-Grain Weight (g) | ×104 Ears ha−1 | Grains per Ear | Yield (kg ha−1) |
|---|---|---|---|---|---|---|
| 2020 | N0 | CN | 2.8 ± 0.1 a | 42.9 ± 2.8 c | 3259.6 ± 294.1 f | 2841.9 ± 266.1 e |
| PN | 2.9 ± 0.0 a | 44.1 ± 1.4 c | 3179.9 ± 397.0 f | 2665.0 ± 576.4 e | ||
| N75 | CN | 3.0 ± 0.0 a | 48.4 ± 5.5 bc | 3762.9 ± 276.6 ef | 4819.2 ± 162.5 cd | |
| PN | 2.8 ± 0.2 a | 47.3 ± 1.4 bc | 5627.1 ± 483.3 abc | 5102.2 ± 41.8 bcd | ||
| N112.5 | CN | 2.9 ± 0.1 a | 48.9 ± 0.6 bc | 3704.0 ± 110.1 ef | 4708.3 ± 40.6 d | |
| PN | 2.7 ± 0.2 a | 54.2 ± 5.3 ab | 5970.8 ± 488.0 ab | 5449.6 ± 182.0 ab | ||
| N150 | CN | 2.8 ± 0.0 a | 54.3 ± 0.0 ab | 4261.0 ± 488.5 def | 5191.2 ± 319.8 bcd | |
| PN | 2.9 ± 0.0 a | 61.8 ± 0.0 a | 4655.5 ± 346.7 cde | 5807.4 ± 189.4 a | ||
| N225 | CN | 2.9 ± 0.0 a | 49.4 ± 0.4 bc | 4304.0 ± 28.8 def | 5322.4 ± 16.2 abc | |
| PN | 2.7 ± 0.0 a | 48.8 ± 1.1 bc | 4989.9 ± 134.5 bcd | 5573.2 ± 114.5 ab | ||
| N337.5 | CN | 2.9 ± 0.1 a | 44.4 ± 1.5 c | 5634.9 ± 224.4 abc | 5358.6 ± 90.6 abc | |
| PN | 2.8 ± 0.1 a | 47.3 ± 1.4 bc | 6178.4 ± 152.8 a | 5654.9 ± 225.7 ab | ||
| 2021 | N0 | CN | 2.8 ± 0.0 a | 31.6 ± 3.4 e | 2715.2 ± 69.1 f | 3394.5 ± 321.8 e |
| PN | 2.8 ± 0.0 ab | 33.3 ± 1.1 cde | 2738.0 ± 72.1 f | 3451.7 ± 92.8 e | ||
| N75 | CN | 2.6 ± 0.0 bc | 43.1 ± 1.8 ab | 3253.5 ± 14.9 e | 4436.0 ± 47.3 d | |
| PN | 2.6 ± 0.0 bc | 47.3 ± 3.7 a | 3924.5 ± 69.3 d | 4900.0 ± 42.4 bc | ||
| N112.5 | CN | 2.7 ± 0.0 abc | 43.3 ± 2.4 ab | 3228.6 ± 44.0 e | 4582.7 ± 157.5 cd | |
| PN | 2.7 ± 0.1 abc | 46.7 ± 3.1 a | 3836.6 ± 46.3 d | 5323.7 ± 45.7 b | ||
| N150 | CN | 2.7 ± 0.0 abc | 41.5 ± 2.0 abc | 3701.8 ± 56.0 d | 5204.2 ± 17.0 b | |
| PN | 2.7 ± 0.0 ab | 44.3 ± 0.5 ab | 4406.0 ± 109.5 c | 5878.5 ± 78.5 a | ||
| N225 | CN | 2.7 ± 0.0 abc | 37.5 ± 2.0 bcde | 3848.1 ± 126.6 d | 4728.6 ± 80.5 cd | |
| PN | 2.7 ± 0.0 abc | 40.7 ± 1.2 abcd | 4575.3 ± 86.4 c | 5221.8 ± 154.7 b | ||
| N337.5 | CN | 2.6 ± 0.0 c | 32.4 ± 2.7 de | 5946.0 ± 128.8 a | 4590.1 ± 171.4 d | |
| PN | 2.7 ± 0.0 abc | 33.3 ± 4.9 cde | 5452.3 ± 240.2 b | 5244.5 ± 216.3 b | ||
| ANOVA | ||||||
| Year | ** | ** | * | ** | ||
| N level | ns | ** | ** | ** | ||
| Treatment | ns | ns | ** | ** |
| Year | Nitrogen Application Level | Treatment | NPFP (kg kg−1) | NAE (kg kg−1) | NRE (%) | NPE (kg kg−1) |
|---|---|---|---|---|---|---|
| 2020 | N75 | CN | 64.3 ± 2.2 b | 26.4 ± 2.2 b | 64.0 ± 1.2 cd | 41.1 ± 0.9 a |
| PN | 68.0 ± 0.6 a | 32.5 ± 0.6 a | 78.7 ± 1.0 a | 41.2 ± 0.2 a | ||
| N112.5 | CN | 41.8 ± 0.4 d | 16.6 ± 0.4 d | 68.6 ± 1.7 b | 24.2 ± 0.3 d | |
| PN | 48.4 ± 1.6 c | 24.8 ± 1.6 b | 79.9 ± 0.7 a | 31.0 ± 0.8 b | ||
| N150 | CN | 34.6 ± 2.1 e | 15.7 ± 2.1 d | 68.2 ± 0.9 bc | 22.9 ± 0.3 d | |
| PN | 38.7 ± 1.3 d | 21.0 ± 1.3 c | 79.5 ± 2.5 a | 26.3 ± 0.8 c | ||
| N225 | CN | 23.7 ± 0.1 f | 11.0 ± 0.1 ef | 61.9 ± 0.4 d | 17.8 ± 0.0 f | |
| PN | 24.8 ± 0.5 f | 12.9 ± 0.5 de | 66.4 ± 1.2 bc | 19.4 ± 0.5 e | ||
| N337.5 | CN | 15.9 ± 0.3 g | 7.5 ± 0.3 f | 46.5 ± 0.8 e | 16.0 ± 0.3 g | |
| PN | 16.8 ± 0.7 g | 8.9 ± 0.7 f | 44.4 ± 1.9 e | 19.9 ± 0.7 e | ||
| 2021 | N75 | CN | 59.2 ± 0.6 b | 13.9 ± 0.6 c | 54.4 ± 3.8 e | 25.6 ± 1.5 b |
| PN | 65.3 ± 0.6 a | 19.3 ± 0.6 a | 70.3 ± 6.6 bc | 27.7 ± 2.3 a | ||
| N112.5 | CN | 40.7 ± 0.3 d | 10.6 ± 0.3 e | 56.2 ± 11.2 e | 19.3 ± 2.4 d | |
| PN | 47.3 ± 0.4 c | 16.6 ± 0.4 b | 78.7 ± 0.5 ab | 21.1 ± 0.6 c | ||
| N150 | CN | 34.7 ± 0.1 f | 12.1 ± 0.1 d | 66.4 ± 2.8 cd | 18.2 ± 0.3 d | |
| PN | 39.2 ± 0.5 e | 16.2 ± 0.5 b | 84.9 ± 3.3 a | 19.1 ± 1.0 d | ||
| N225 | CN | 21.0 ± 0.4 h | 5.9 ± 0.4 g | 63.7 ± 8.1 cde | 9.4 ± 0.5 e | |
| PN | 23.2 ± 0.7 g | 7.9 ± 0.7 f | 80.0 ± 2.2 ab | 9.8 ± 0.8 e | ||
| N337.5 | CN | 13.6 ± 0.5 j | 3.5 ± 0.5 h | 59.9 ± 2.2 de | 5.9 ± 0.8 f | |
| PN | 15.5 ± 0.6 i | 5.3 ± 0.6 g | 76.5 ± 1.6 ab | 7.0 ± 0.9 f | ||
| ANOVA | ||||||
| Year | ns | ** | ns | ** | ||
| N level | ** | ** | ** | ** | ||
| Treatment | * | * | ns | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Wang, Q.; Zhang, W.; Gao, M.; Xv, Y.; Li, S.; Dong, H.; Chen, D.; Yan, P.; Dong, Z. Urea Coated with Polyaspartic Acid-Chitosan Increases Foxtail Millet (Setaria italica L. Beauv.) Grain Yield by Improving Nitrogen Metabolism. Plants 2024, 13, 415. https://doi.org/10.3390/plants13030415
Lu L, Wang Q, Zhang W, Gao M, Xv Y, Li S, Dong H, Chen D, Yan P, Dong Z. Urea Coated with Polyaspartic Acid-Chitosan Increases Foxtail Millet (Setaria italica L. Beauv.) Grain Yield by Improving Nitrogen Metabolism. Plants. 2024; 13(3):415. https://doi.org/10.3390/plants13030415
Chicago/Turabian StyleLu, Lin, Qi Wang, Wei Zhang, Ming Gao, Yanli Xv, Shujie Li, Haosheng Dong, Disu Chen, Peng Yan, and Zhiqiang Dong. 2024. "Urea Coated with Polyaspartic Acid-Chitosan Increases Foxtail Millet (Setaria italica L. Beauv.) Grain Yield by Improving Nitrogen Metabolism" Plants 13, no. 3: 415. https://doi.org/10.3390/plants13030415