
Biochemical and Structural Diversification of C4 Photosynthesis in Tribe Zoysieae (Poaceae)
 ,
,
Abstract
:1. Introduction
2. Results
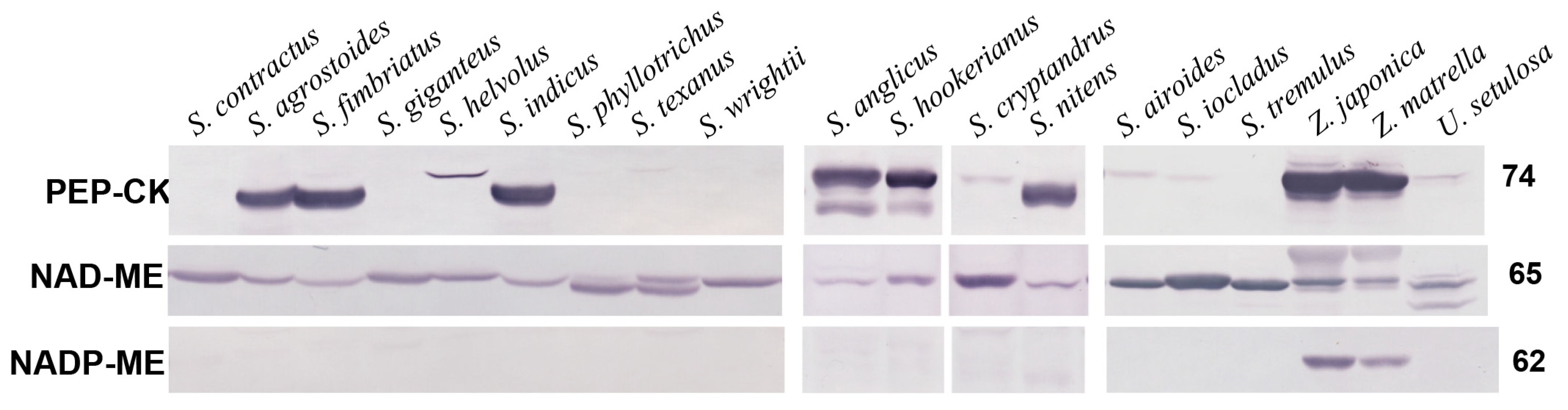
2.1. Western Blotting
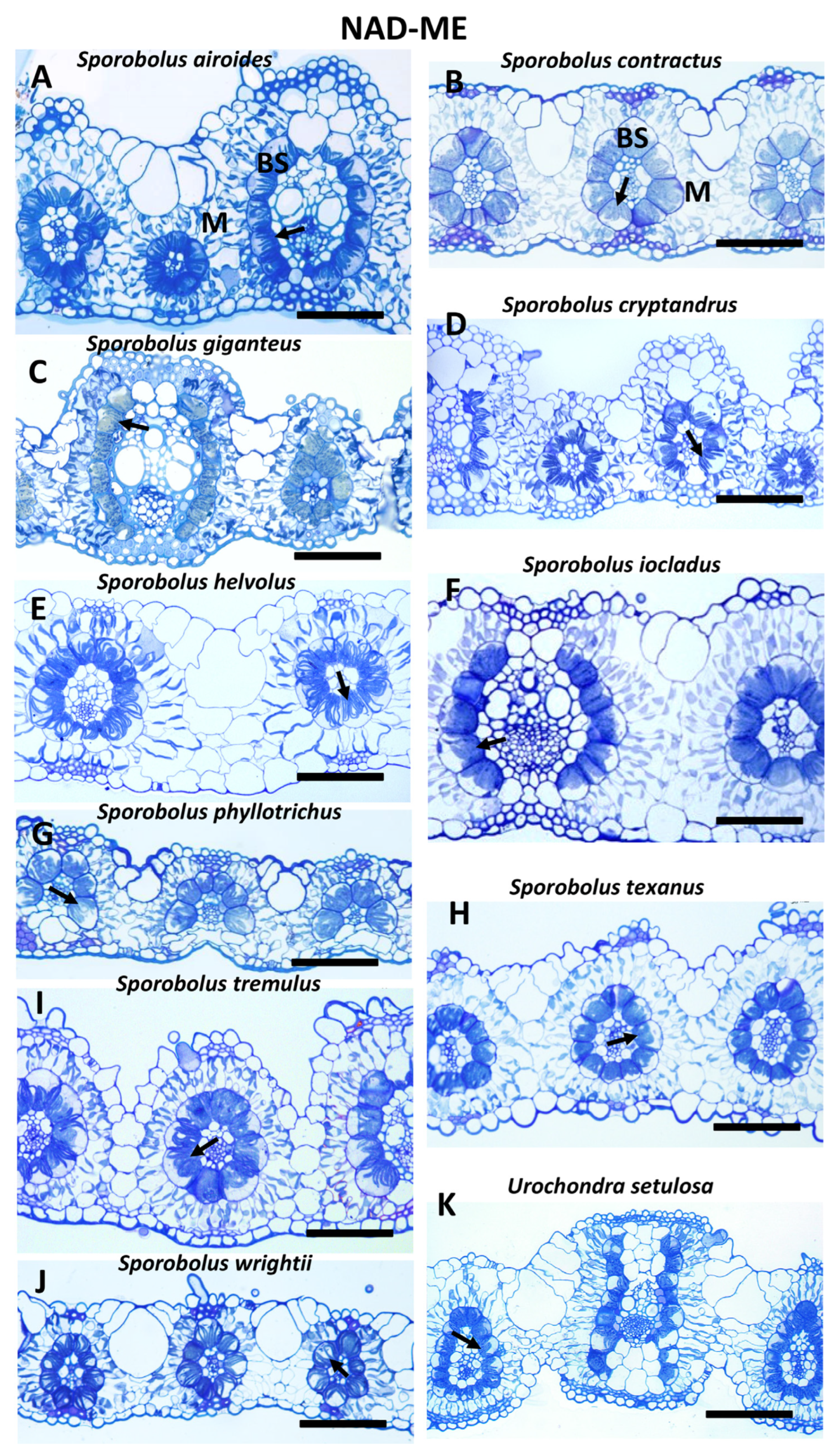
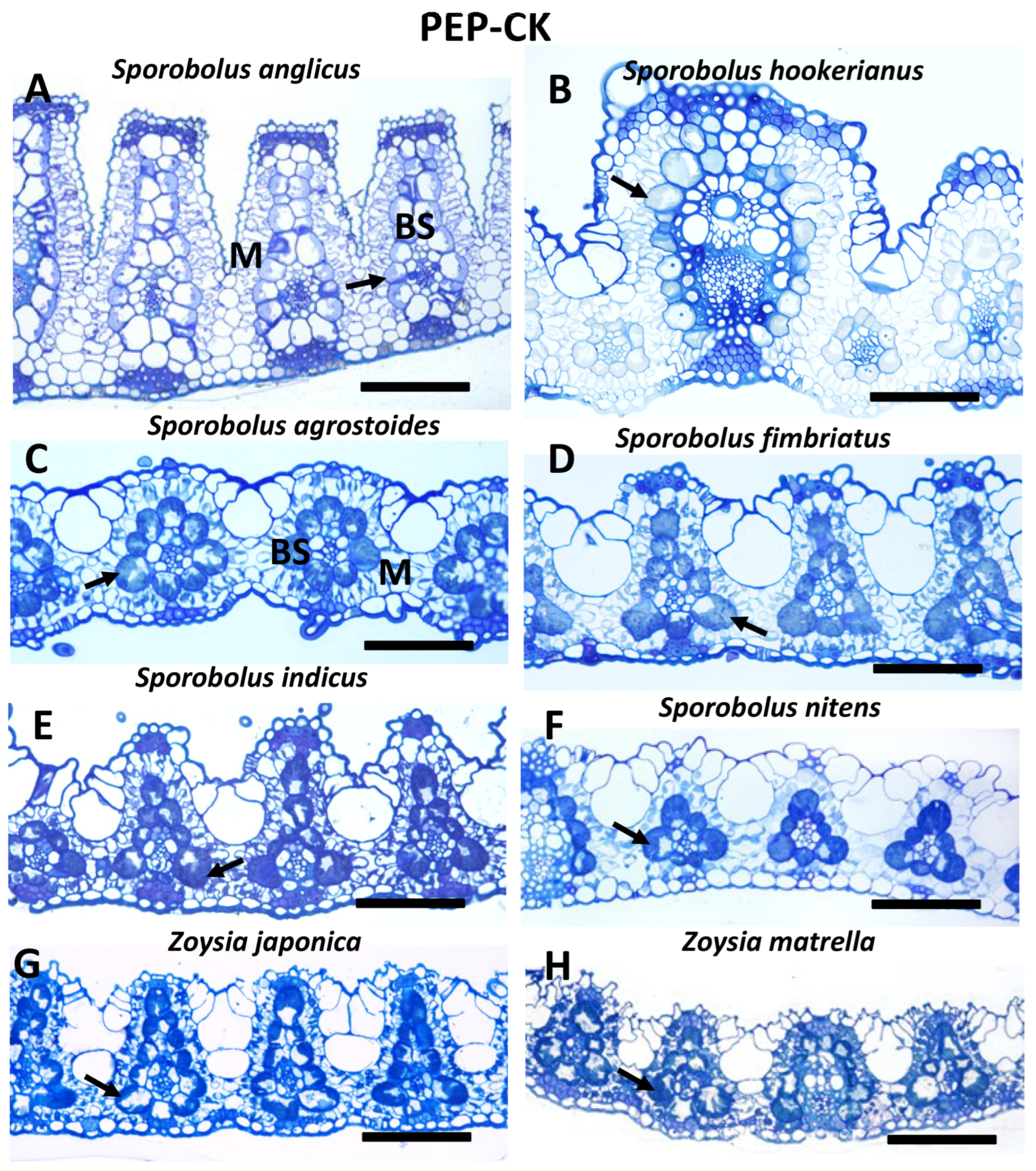
2.2. Light Microscopy
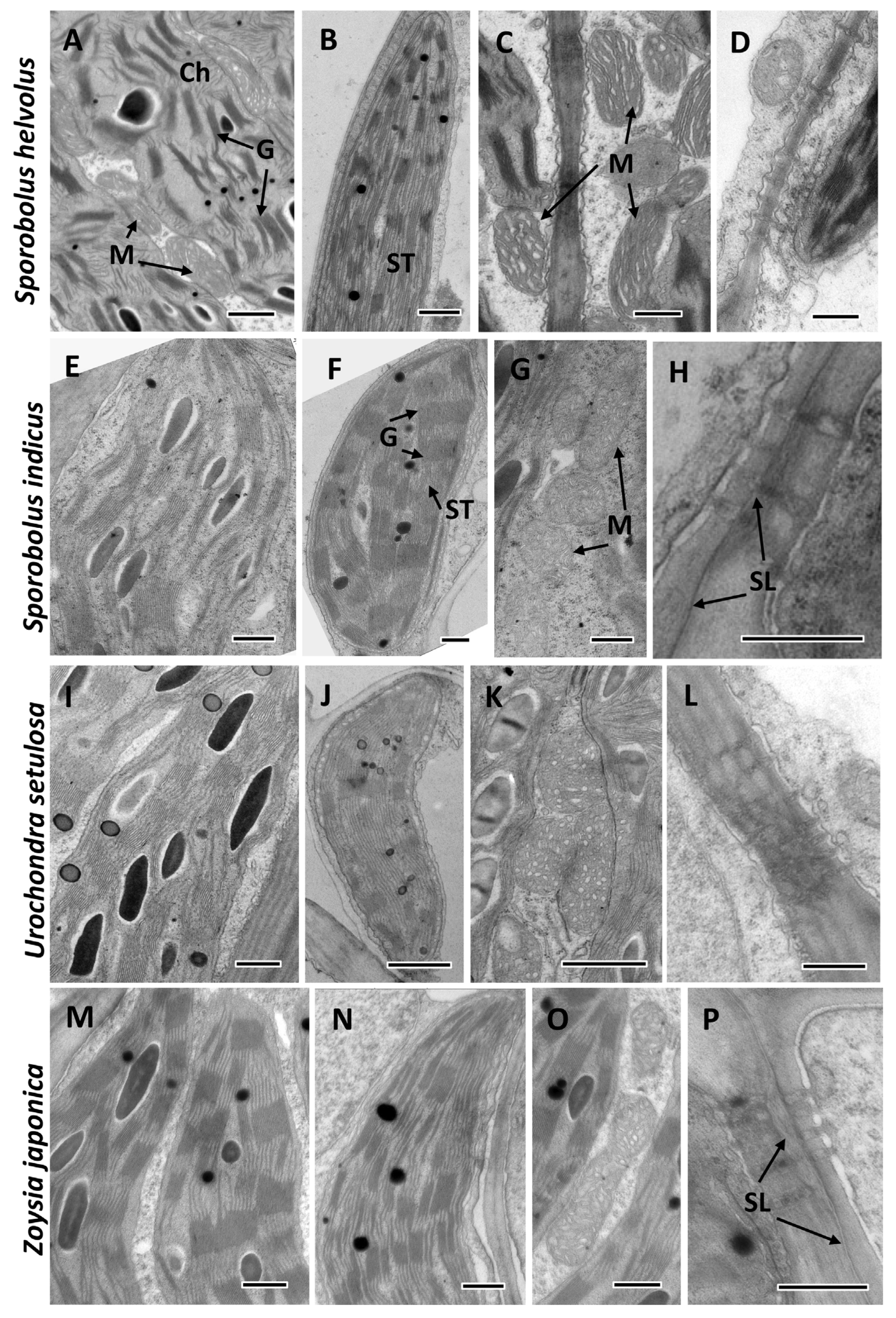
2.3. Transmission Electron Microscopy
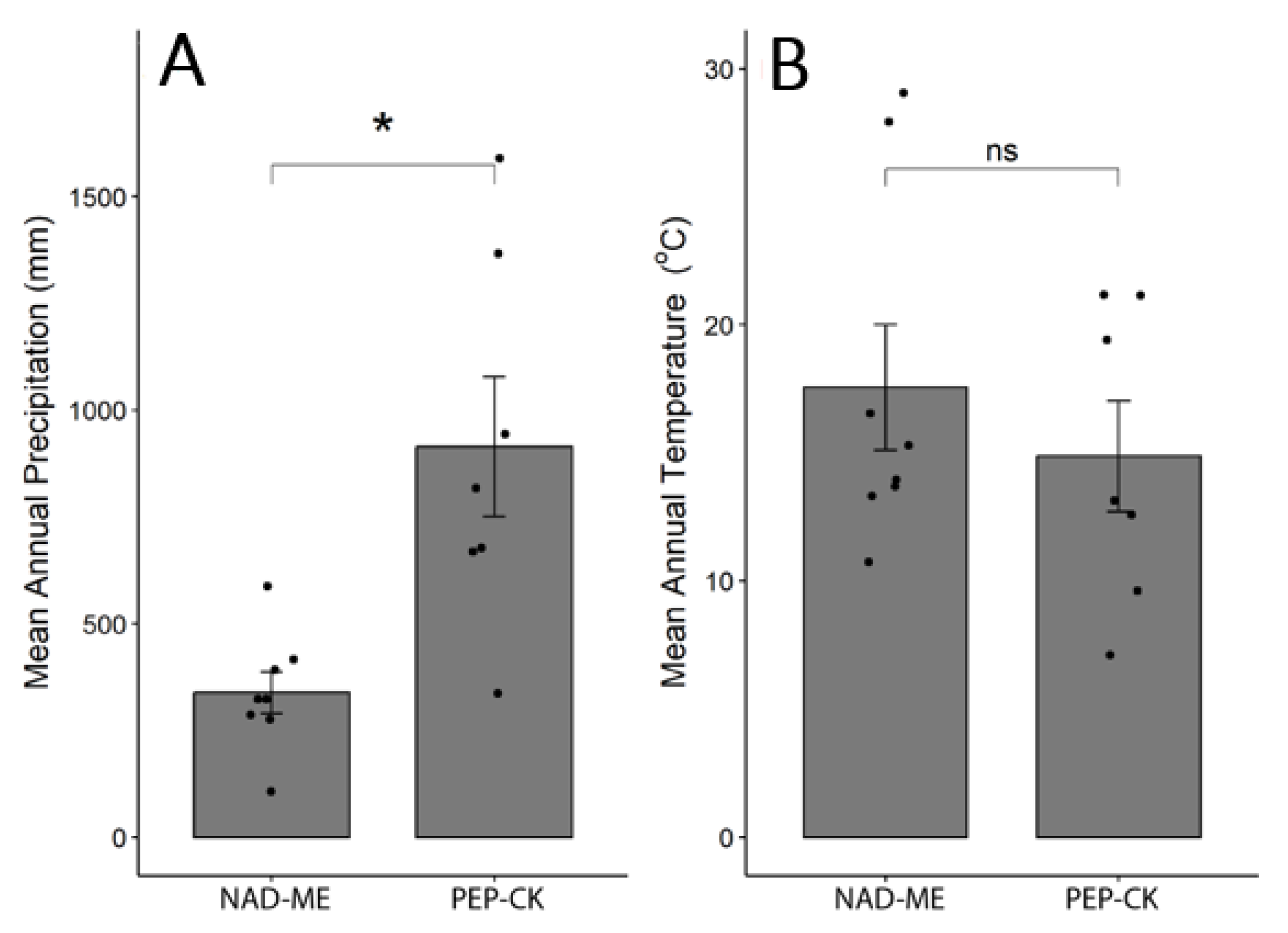
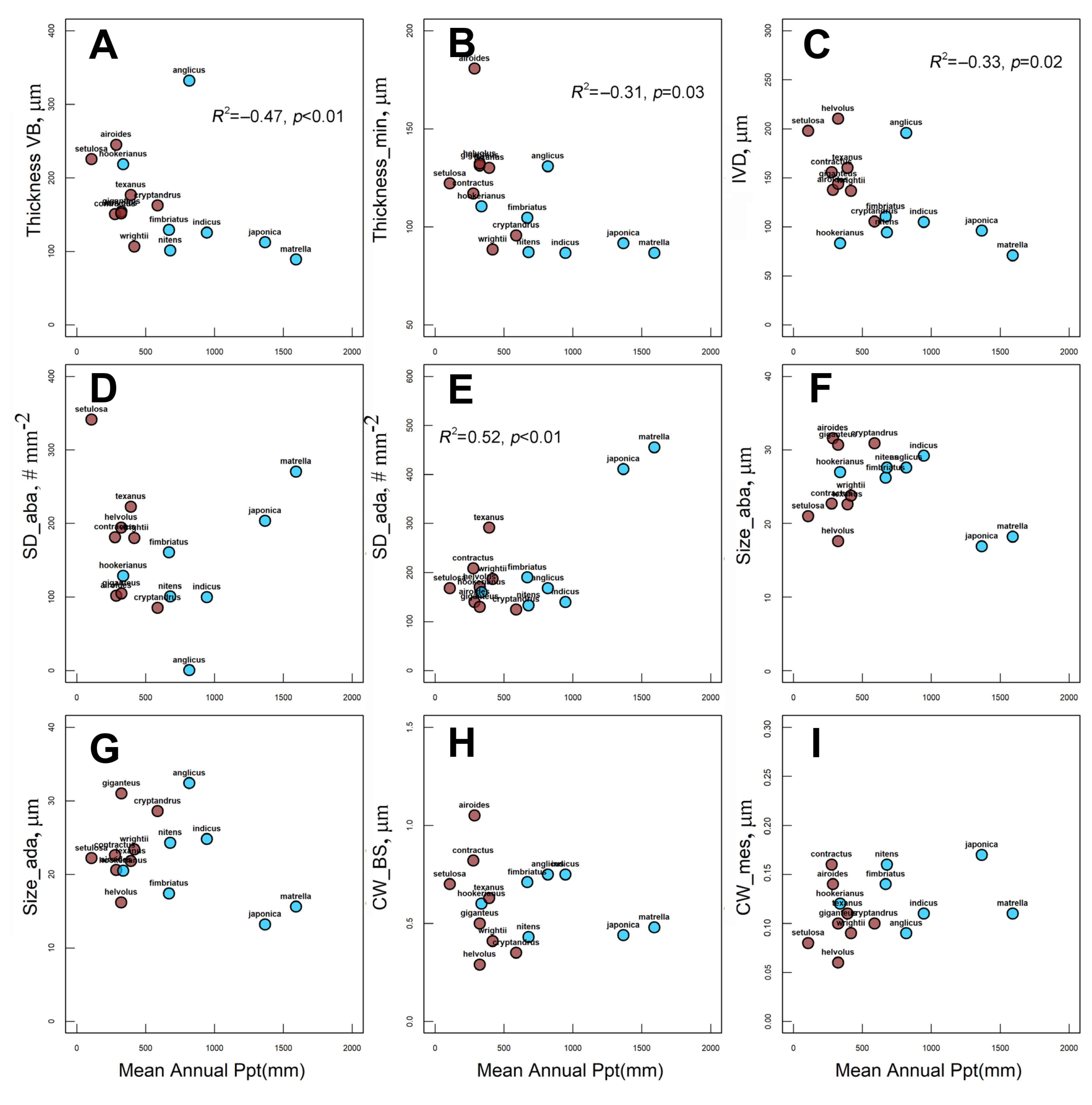
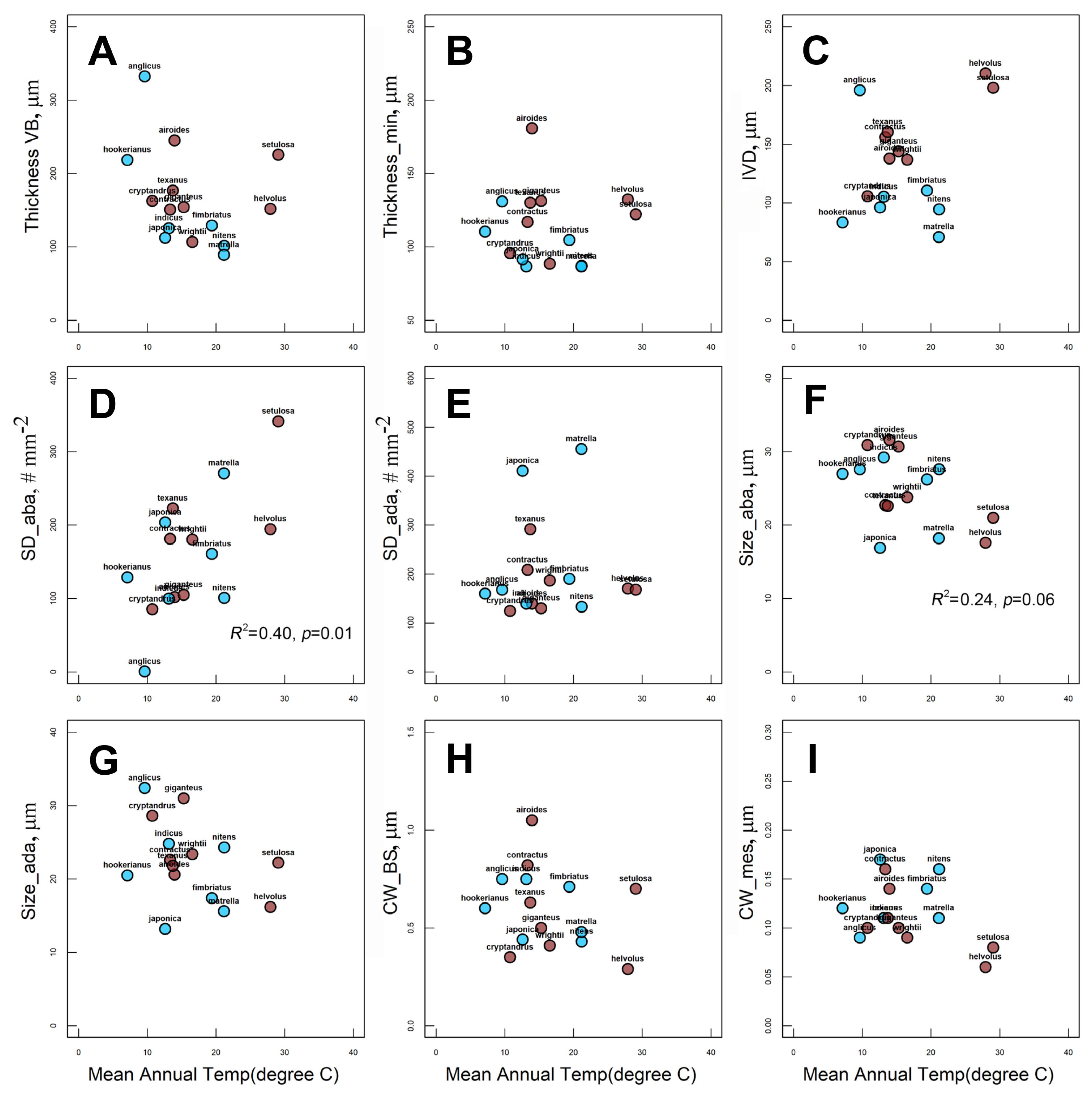
2.4. Effect of Habitat MAP and MAT on Species Distribution and Anatomical Traits
3. Discussion
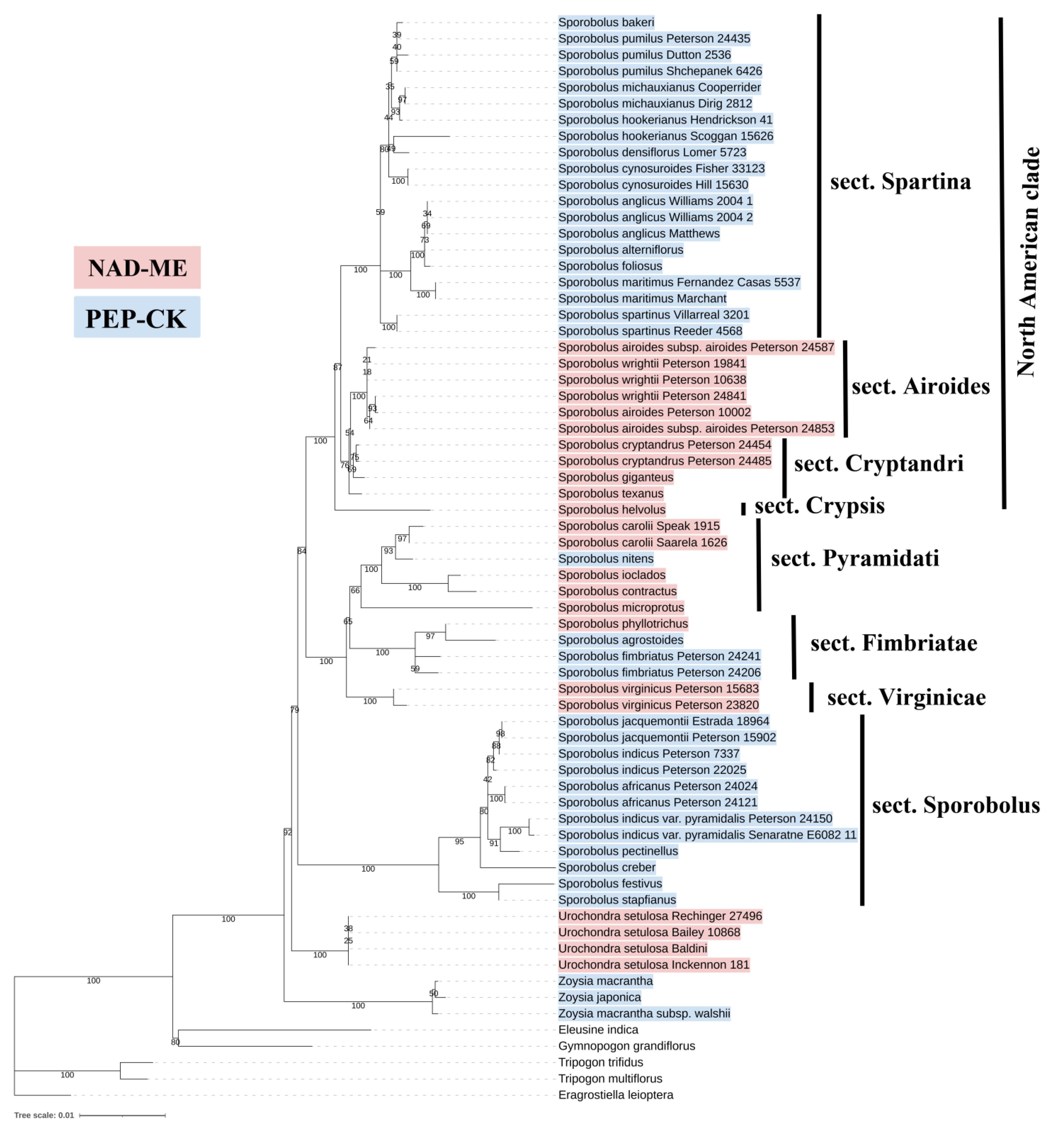
3.1. Diversity in C4 Subtypes in Zoysieae
3.2. Evolutionary Scenario of C4 Diversification
3.3. Habitat Effect on Zoysieae Diversification
4. Materials and Methods
4.1. Plant Growth Conditions
4.2. Light and Scan and Transmission Electron Microscopy
4.3. Western Blot Analysis
4.4. Phylogenetic Tree Reconstruction
4.5. Habitat Mean Annual Precipitation and Mean Annual Temperature
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hatch, M.D. C4 photosynthesis: A unique blend of modified biochemistry, anatomy and ultrastructure. Biochim. Biophys. Acta 1987, 895, 81–106. [Google Scholar] [CrossRef]
- Edwards, G.E.; Walker, D.A. C3, C4: Mechanisms, and Cellular and Environmental Regulation, of Photosynthesis; Blackwell Scientific Publications: Oxford, UK, 1983; p. 542. [Google Scholar]
- Edwards, G.E.; Voznesenskaya, E.V. C4 photosynthesis: Kranz forms and single-cell C4 in terrestrial plants. In C4 Photosynthesis and Related CO2 Concentrating Mechanisms; Raghavendra, A.S., Sage, R.F., Eds.; Advances in Photosynthesis Research; Springer: Dordrecht, The Netherlands, 2011; pp. 29–61. [Google Scholar]
- Gutierrez, M.; Gracen, V.E.; Edwards, G.E. Biochemical and cytological relationships in C4 plants. Planta 1974, 119, 279–300. [Google Scholar] [CrossRef] [PubMed]
- Hatch, M.D.; Kagawa, T.; Craig, S. Subdivision of C4-pathway species based on differing C4 acid decarboxylating systems and ultrastructural features. Aust. J. Plant Physiol. 1975, 2, 111–128. [Google Scholar] [CrossRef]
- Sage, R.F.; Li, M.; Monson, R.K. The taxonomic distribution of C4 photosynthesis. In C4 Plant Biology; Sage, R.F., Monson, R.K., Eds.; Academic Press: New York, NY, USA, 1999; pp. 551–584. [Google Scholar]
- Burnell, J.N.; Hatch, M.D. Photosynthesis in phosphoenolpyruvate carboxykinase-type C4 plants: Pathways of C4 acid decarboxylation in bundle sheath cells of Urochloa panicoides. Arch. Biochem. Biophys. 1988, 260, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Kanai, R.; Edwards, G. The biochemistry of C4 photosynthesis. In C4 Plant Biology. Physiological Ecology Series; Sage, R.F., Monson, R.K., Eds.; Academic Press: San Diego, CA, USA, 1999; pp. 49–87. [Google Scholar]
- Voznesenskaya, E.V.; Franceschi, V.R.; Chuong, S.D.X.; Edwards, G.E. Functional characterization of phosphoenolpyruvate carboxykinase type C4 leaf anatomy: Immuno, cytochemical and ultrastructural analyses. Ann. Bot. 2006, 98, 77–91. [Google Scholar] [CrossRef]
- Chapman, K.S.R.; Hatch, M.D. Asparate decarboxylation in bundle sheath cells of Zea mays and its possible contribution to C4 photosynthesis. Aust. J. Plant Physiol. 1981, 8, 237–248. [Google Scholar] [CrossRef]
- Wingler, A.; Walker, R.P.; Chen, Z.-H.; Leegood, R.C. Phosphoenolpyruvate carboxykinase is involved in the decarboxylation of aspartate in the bundle sheath of maize. Plant Physiol. 1999, 120, 539–545. [Google Scholar] [CrossRef]
- Koteyeva, N.K.; Voznesenskaya, E.V.; Edwards, G.E. An assessment of the capacity for phosphoenolpyruvate carboxykinase to contribute to C4 photosynthesis. Plant Sci. 2015, 235, 70–80. [Google Scholar] [CrossRef]
- Muhaidat, R.; McKown, A.D. Significant involvement of PEP-CK in carbon assimilation of C4 eudicots. Ann. Bot. 2013, 111, 577–589. [Google Scholar] [CrossRef]
- Sommer, M.; Bräutigam, A.; Weber, A.P.M. The dicotyledonous NAD malic enzyme C4 plant Cleome gynandra displays age-dependent plasticity of C4 decarboxylation biochemistry. Plant Biol. 2012, 14, 621–629. [Google Scholar] [CrossRef]
- Furbank, R.T. Evolution of the C4 photosynthetic mechanism: Are there really three C4 acid decarboxylation types? J. Exp. Bot. 2011, 62, 3103–3108. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, H.D.V.; Hattersley, P.W.; Stone, N.E. New structural/biochemical associations in leaf blades of C4 grasses (Poaceae). Aust. J. Plant Physiol. 1987, 14, 403–420. [Google Scholar] [CrossRef]
- Hattersley, P.; Watson, L. Diversification of photosynthesis. In Grass Evolution and Domestication; Chapman, G., Ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 1992; pp. 38–116. [Google Scholar]
- Hattersley, P.W. C4 photosynthetic pathway variation in grasses (Poaceae): Its significance for arid and semi-arid lands. In Desertified Grasslands: Their Biology and Management; Chapman, G., Ed.; Academic Press: London, UK, 1992; pp. 181–212. [Google Scholar]
- Ingram, A.L. Evolution of leaf blade anatomy in Eragrostis (Poaceae). Syst. Bot. 2010, 35, 755–765. [Google Scholar] [CrossRef]
- Sage, R.F. A portrait of the C4 photosynthetic family on the 50th anniversary of its discovery: Species number, evolutionary lineages, and Hall of Fame. J. Exp. Bot. 2016, 67, 4039–4056. [Google Scholar] [CrossRef] [PubMed]
- Grass Phylogeny Working Group II. New grass phylogeny resolves deep evolutionary relationships and discovers C4 origins. New Phytol. 2012, 193, 304–312. [Google Scholar] [CrossRef]
- Christin, P.-A. Traces of strong selective pressures in the genomes of C4 grasses. J. Exp. Bot. 2017, 68, 103–105. [Google Scholar] [CrossRef]
- Bianconi, M.E.; Hackel, J.; Vorontsova, M.S.; Alberti, A.; Arthan, W.; Burke, S.V.; Duvall, M.R.; Kellogg, E.A.; Lavergne, S.; McKain, M.R.; et al. Continued adaptation of C4 photosynthesis after an initial burst of changes in the Andropogoneae grasses. Syst. Biol. 2019, 69, 445–461. [Google Scholar] [CrossRef]
- Soreng, R.J.; Peterson, P.M.; Romaschenko, K.; Davidse, G.; Zuloaga, F.O.; Judziewicz, E.J.; Filgueiras, T.S.; Davis, J.I.; Morrone, O. A worldwide phylogenetic classification of the Poaceae (Gramineae). J. Syst. Evol. 2015, 53, 117–137. [Google Scholar] [CrossRef]
- Maricle, B.R.; Koteyeva, N.K.; Voznesenskaya, E.V.; Thomasson, J.R.; Edwards, G.E. Diversity in leaf anatomy, and stomatal distribution and conductance, between salt marsh and freshwater species in the C4 genus Spartina (Poaceae). New Phytol. 2009, 184, 216–233. [Google Scholar] [CrossRef]
- Schmidt, M.; König, K.; Müller, J.; Brunken, U.; Zizka, G. Modelling the distribution of photosynthetic types of grasses in Sahelian Burkina Faso with high-resolution satellite data. Ecotropica 2011, 17, 53–63. [Google Scholar]
- Vecchia, F.D.; El Asmar, T.; Calamassi, R.; Rascio, N.; Vazzana, C. Morphological and ultrastructural aspects of dehydration and rehydration in leaves of Sporobolus stapfianus. Plant Growth Regul. 1998, 24, 219–228. [Google Scholar] [CrossRef]
- Walker, R.P.; Acheson, R.M.; Tecsi, L.I.; Leegood, R.C. Phosphoenolpyruvate carboxykinase in C4 plants: Its role and regulation. Aust. J. Plant Physiol. 1997, 24, 459–468. [Google Scholar] [CrossRef]
- Kemp, P.R.; Cunningham, G.L.; Adams, H.P. Specialization of mesophyll morphology in relation to C4 photosynthesis in the Poaceae. Am. J. Bot. 1983, 70, 349–354. [Google Scholar] [CrossRef]
- Carmo-Silva, A.E.; Bernardes da Silva, A.; Keys, A.J.; Parry, M.A.J.; Arrabaça, M.C. The activities of PEP carboxylase and the C4 acid decarboxylases are little changed by drought stress in three C4 grasses of different subtypes. Photosynth. Res. 2008, 97, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Ueno, O. Immunogold localization of photosynthetic enzymes in leaves of various C4 plants, with particular reference to pyruvate orthophosphate dikinase. J. Exp. Bot. 1998, 49, 1637–1646. [Google Scholar] [CrossRef]
- Buchmann, N.; Brooks, J.R.; Rapp, K.D.; Ehleringer, J.R. Carbon isotope composition of C4 grasses is influenced by light and water supply. Plant Cell Environ. 1996, 19, 392–402. [Google Scholar] [CrossRef]
- Peterson, P.M.; Romaschenko, K.; Arrieta, Y.H.; Saarela, J.M. A molecular phylogeny and new subgeneric classification of Sporobolus (Poaceae: Chloridoideae: Sporobolinae). Taxon 2014, 63, 1212–1243. [Google Scholar] [CrossRef]
- Peterson, P.; Romaschenko, K.; Arrieta, Y.; Saarela, J. A molecular phylogeny of the subtribe Sporobolinae and a classification of the subfamily Chloridoideae (Poaceae). Mem. N. Y. Bot. Gard. 2017, 188, 127–151. [Google Scholar] [CrossRef]
- Renvoize, S.A. A survey of leaf-blade anatomy in grasses IV. Eragrostideae. Kew Bull. 1983, 38, 469–478. [Google Scholar] [CrossRef]
- Christin, P.-A.; Petitpierre, B.; Salamin, N.; Büchi, L.; Besnard, G. Evolution of C4 phosphoenolpyruvate carboxykinase in grasses, from genotype to phenotype. Mol. Biol. Evol. 2009, 26, 357–365. [Google Scholar] [CrossRef]
- Prendergast, H.D.V.; Hattersley, P.W. Australian C4 grasses (Poaceae): Leaf blade anatomical features in relation to C4 acid decarboxylation types. Aust. J. Bot. 1987, 35, 355–382. [Google Scholar] [CrossRef]
- Edwards, G.E.; Kanai, R.; Black, C.C. Phosphoenolpyruvate carboxykinase in leaves of certain plants which fix CO2 by the C4-dicarboxylic acid cycle of photosynthesis. Biochem. Biophys. Res. Commun. 1971, 45, 278–285. [Google Scholar] [CrossRef]
- Koteyeva, N.K.; Voznesenskaya, E.V.; Berim, A.; Gang, D.R.; Edwards, G.E. Structural diversity in salt excreting glands and salinity tolerance in Oryza coarctata, Sporobolus anglicus and Urochondra setulosa. Planta 2023, 257, 9. [Google Scholar] [CrossRef] [PubMed]
- Christin, P.-A.; Freckleton, R.P.; Osborne, C.P. Can phylogenetics identify C4 origins and reversals? Trends Ecol. Evol. 2010, 25, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.P.; Trevanion, S.J.; Leegood, R.C. Phosphoenolpyruvate carboxykinase from higher-plants: Purification from cucumber and evidence of rapid proteolytic cleavage in extracts from a range of plant-tissues. Planta 1995, 196, 58–63. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Cerling, T.E.; Helliker, B.R. C4 photosynthesis, atmospheric CO2, and climate. Oecologia 1997, 112, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Osborne, C.P.; Freckleton, R.P. Ecological selection pressures for C4 photosynthesis in the grasses. Proc. R. Soc. B Biol. Sci. 2009, 276, 1753–1760. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.F. The evolution of C4 photosynthesis. New Phytol. 2004, 161, 341–370. [Google Scholar] [CrossRef]
- Cabido, M.; Pons, E.; Cantero, J.J.; Lewis, J.P.; Anton, A. Photosynthetic pathway variation among C4 grasses along a precipitation gradient in Argentina. J. Biogeogr. 2008, 35, 131–140. [Google Scholar] [CrossRef]
- Taub, D.R. Climate and the U.S. distribution of C4 grass subfamilies and decarboxylation variants of C4 photosynthesis. Am. J. Bot. 2000, 87, 1211–1215. [Google Scholar] [CrossRef]
- Vogel, J.C.; Fuls, A.; Danin, A. Geographical and environmental distribution of C3 and C4 grasses in the Sinai, Negev, and Judean deserts. Oecologia 1986, 70, 258–265. [Google Scholar] [CrossRef]
- Liu, H.; Osborne, C.P. Water relations traits of C4 grasses depend on phylogenetic lineage, photosynthetic pathway, and habitat water availability. J. Exp. Bot. 2014, 66, 761–773. [Google Scholar] [CrossRef]
- Fravolini, A.; Williams, D.G.; Thompson, T.L. Carbon isotope discrimination and bundle sheath leakiness in three C4 subtypes grown under variable nitrogen, water and atmospheric CO2 supply. J. Exp. Bot. 2002, 53, 2261–2269. [Google Scholar] [CrossRef] [PubMed]
- Carmo-Silva, A.E.; Soares, A.S.; Marques da Silva, J.; Bernardes da Silva, A.; Keys, A.J.; Arrabaça, M.C. Photosynthetic responses of three C4 grasses of different metabolic subtypes to water deficit. Funct. Plant Biol. 2007, 34, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Pathare, V.S.; Sonawane, B.V.; Koteyeva, N.; Cousins, A.B. C4 grasses adapted to low precipitation habitats show traits related to greater mesophyll conductance and lower leaf hydraulic conductance. Plant Cell Environ. 2020, 43, 1897–1910. [Google Scholar] [CrossRef]
- Sack, L.; Holbrook, N.M. Leaf hydraulics. Annu. Rev. Plant Biol. 2006, 57, 361–381. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Clemente-Moreno, M.J.; Bota, J.; Brodribb, T.J.; Gago, J.; Mizokami, Y.; Nadal, M.; Perera-Castro, A.V.; Roig-Oliver, M.; Sugiura, D.; et al. Cell wall thickness and composition are involved in photosynthetic limitation. J. Exp. Bot. 2021, 72, 3971–3986. [Google Scholar] [CrossRef] [PubMed]
- Pathare, V.S.; Panahabadi, R.; Sonawane, B.V.; Apalla, A.J.; Koteyeva, N.; Bartley, L.E.; Cousins, A.B. Altered cell wall hydroxycinnamate composition impacts leaf- and canopy-level CO2 uptake and water use in rice. Plant Physiol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Danila, F.R.; Thakur, V.; Chatterjee, J.; Bala, S.; Coe, R.A.; Acebron, K.; Furbank, R.T.; von Caemmerer, S.; Quick, W.P. Bundle sheath suberisation is required for C4 photosynthesis in a Setaria viridis mutant. Commun. Biol. 2021, 4, 254. [Google Scholar] [CrossRef] [PubMed]
- Pathare, V.S.; Koteyeva, N.; Cousins, A.B. Increased adaxial stomatal density is associated with greater mesophyll surface area exposed to intercellular air spaces and mesophyll conductance in diverse C4 grasses. New Phytol. 2020, 225, 169–182. [Google Scholar] [CrossRef]
- Redmann, R.E. Adaptation of grasses to water stress-leaf rolling and stomate distribution. Ann. Mo. Bot. Gard. 1985, 72, 833–842. [Google Scholar] [CrossRef]
- Taylor, S.H.; Franks, P.J.; Hulme, S.P.; Spriggs, E.; Christin, P.A.; Edwards, E.J.; Woodward, F.I.; Osborne, C.P. Photosynthetic pathway and ecological adaptation explain stomatal trait diversity amongst grasses. New Phytol. 2012, 193, 387–396. [Google Scholar] [CrossRef]
- Long, J.J.; Berry, J.O. Tissue-specific and light-mediated expression of the C4 photosynthetic NAD-dependent malic enzyme of amaranth mitochondria. Plant Physiol. 1996, 112, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Maurino, V.G.; Drincovich, M.F.; Andreo, C.S. NADP-malic enzyme isoforms in maize leaves. Biochem. Mol. Biol. Int. 1996, 38, 239–250. [Google Scholar] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Bartley, L.E.; Peck, M.L.; Kim, S.-R.; Ebert, B.; Manisseri, C.; Chiniquy, D.M.; Sykes, R.; Gao, L.; Rautengarten, C.; Vega-Sánchez, M.E.; et al. Overexpression of a BAHD acyltransferase, OsAt10, alters rice cell wall hydroxycinnamic acid content and saccharification. Plant Physiol. 2013, 161, 1615–1633. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Hijmans, R.; van Etten, J. Geographic analysis and modeling with raster data. R Package Version 2012, 2, 1–25. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band Intensity in % to Reference Species * | Biochemical Subtype | BS Chloroplast Position | BS CW Suberin | Leaf Thickness **, µm | Interveinal Distance, µm | BS Mitochondria Size, Short Axes, µm | |||
|---|---|---|---|---|---|---|---|---|---|
| PEP-CK | NAD-ME | NADP-ME | |||||||
| S. airoides | 7 ± 3.3 d | 100 b | 0 | NAD-ME | cp | - | 245.20 ± 3.23 b | 137.86 ± 2.57 c | 0.61 ± 0.02 bc |
| S. contractus | 0 | 96 ± 6.9 b | 0 | NAD-ME | cp | - | 150.95 ± 1.87 d | 156.00 ± 2.79 bc | 0.59 ± 0.03 bc |
| S. cryptandrus | 3 ± 2.3 d | 93 ± 5.9 b | 0 | NAD-ME | cp | - | 162.58 ± 6.29 cd | 105.70 ± 2.83 d | 0.59 ± 0.03 bc |
| S. giganteus | 0 | 90 ± 7.5 bc | 0 | NAD-ME | cp | - | 154.4 ± 3.24 d | 143.9 ± 3.08 c | 0.56 ± 0.03 c |
| S. helvolus | 13 ± 2.3 d | 75 ± 6.0 c | 0 | NAD-ME | cp | - | 151.92 ± 8.04 cd | 210.43 ± 7.13 a | 0.68 ± 0.05 ab |
| S. ioclados | 5 ± 3.2 d | 137 ± 33.4 ab | 0 | NAD-ME | cp | - | 222.41 ± 5.00 b | 213.08 ± 9.71 a | 0.71 ± 0.03 ab |
| S. phyllotrichus | 0 | 122 ± 8.6 a | 0 | NAD-ME | cp | - | 101.54 ± 5.55 fg | 135.97 ± 3.07 c | 0.50 ± 0.03 c |
| S. texanus | 0 | 138 ± 7.3 a | 0 | NAD-ME | cp | - | 176.93 ± 6.42 c | 160.39 ± 6.11 b | 0.42 ± 0.02 d |
| S. tremulus | 0 | 91 ± 13.8 bc | 0 | NAD-ME | cp | - | 173.50 ± 3.97 c | 165.55 ± 4.15 b | 0.82 ± 0.04 a |
| S. wrightii | 0 | 90 ± 7.2 bc | 0 | NAD-ME | cp | - | 106.85 ± 3.70 f | 136.94 ± 4.51 c | 0.38 ± 0.01 d |
| U. setulosa | 10 ± 3.5 d | 73 ± 2.6 c | 0 | NAD-ME | cp | - | 225.60 ± 9.54 b | 197.97 ± 4.01 a | 0.69 ± 0.03 b |
| Average for NAD-ME species | 3.4 *** | 100.5 *** | 0 *** | 170.2 ns | 160.3 *** | 0.60 ns | |||
| S. anglicus | 100 b | 49 ± 9.3 cd | 0 | PEP-CK | cf | + | 312.38 ± 14.71 a | 182.73 ± 7.55 a | 0.66 ± 0.06 b |
| S. agrostoides | 59 ± 3.23 c | 53 ± 8.1 cd | 0 | PEP-CK | cf | + | 124.82 ± 2.91 e | 127.40 ± 3.61 c | 0.41 ± 0.02 d |
| S. fimbriatus | 58 ± 3.2 c | 24 ± 5.3 e | 0 | PEP-CK | cf | + | 129.36 ± 3.87 e | 110.56 ± 3.69 d | 0.41 ± 0.02 d |
| S. hookerianus | 111 ± 13.2 b | 59 ± 9.4 cd | 0 | PEP-CK | cf | + | 223.20 ± 6.33 b | 83.44 ± 1.80 e | 0.65 ± 0.05 b |
| S. indicus | 67 ± 5.5 c | 33 ± 10.2 de | 0 | PEP-CK | cf | + | 125.78 ± 3.44 e | 105.16 ± 3.22 d | 0.45 ± 0.01 d |
| S. nitens | 61 ± 6.4 c | 25 ± 5.3 e | 0 | PEP-CK | cf | + | 101.58 ± 4.14 fg | 94.57 ± 4.25 d | 0.53 ± 0.02 cd |
| Z. japonica | 196 ± 23.6 a | 94 ± 15.7 ab | 42 ± 5.7 a | PEP-CK | cf | + | 112.44 ± 2.89 ef | 96.24 ± 2.06 d | 0.42 ± 0.01 d |
| Z. matrella | 155 ± 31.2 a | 73 ± 2.3 c | 30 ± 4.1 a | PEP-CK | cf | + | 89.20 ± 2.32 g | 71.02 ± 1.32 f | 0.30 ± 0.01 e |
| Average for PEP-CK species | 100.9 | 51.3 | 36 | 163.8ns | 140.4 | 0.43 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koteyeva, N.K.; Voznesenskaya, E.V.; Pathare, V.S.; Borisenko, T.A.; Zhurbenko, P.M.; Morozov, G.A.; Edwards, G.E. Biochemical and Structural Diversification of C4 Photosynthesis in Tribe Zoysieae (Poaceae). Plants 2023, 12, 4049. https://doi.org/10.3390/plants12234049
Koteyeva NK, Voznesenskaya EV, Pathare VS, Borisenko TA, Zhurbenko PM, Morozov GA, Edwards GE. Biochemical and Structural Diversification of C4 Photosynthesis in Tribe Zoysieae (Poaceae). Plants. 2023; 12(23):4049. https://doi.org/10.3390/plants12234049
Chicago/Turabian StyleKoteyeva, Nuria K., Elena V. Voznesenskaya, Varsha S. Pathare, Tatyana A. Borisenko, Peter M. Zhurbenko, Grigory A. Morozov, and Gerald E. Edwards. 2023. "Biochemical and Structural Diversification of C4 Photosynthesis in Tribe Zoysieae (Poaceae)" Plants 12, no. 23: 4049. https://doi.org/10.3390/plants12234049