
Genome-Wide Association Study of Starch Properties in Local Thai Rice
 , ,
, ,
Abstract
:1. Introduction
2. Results
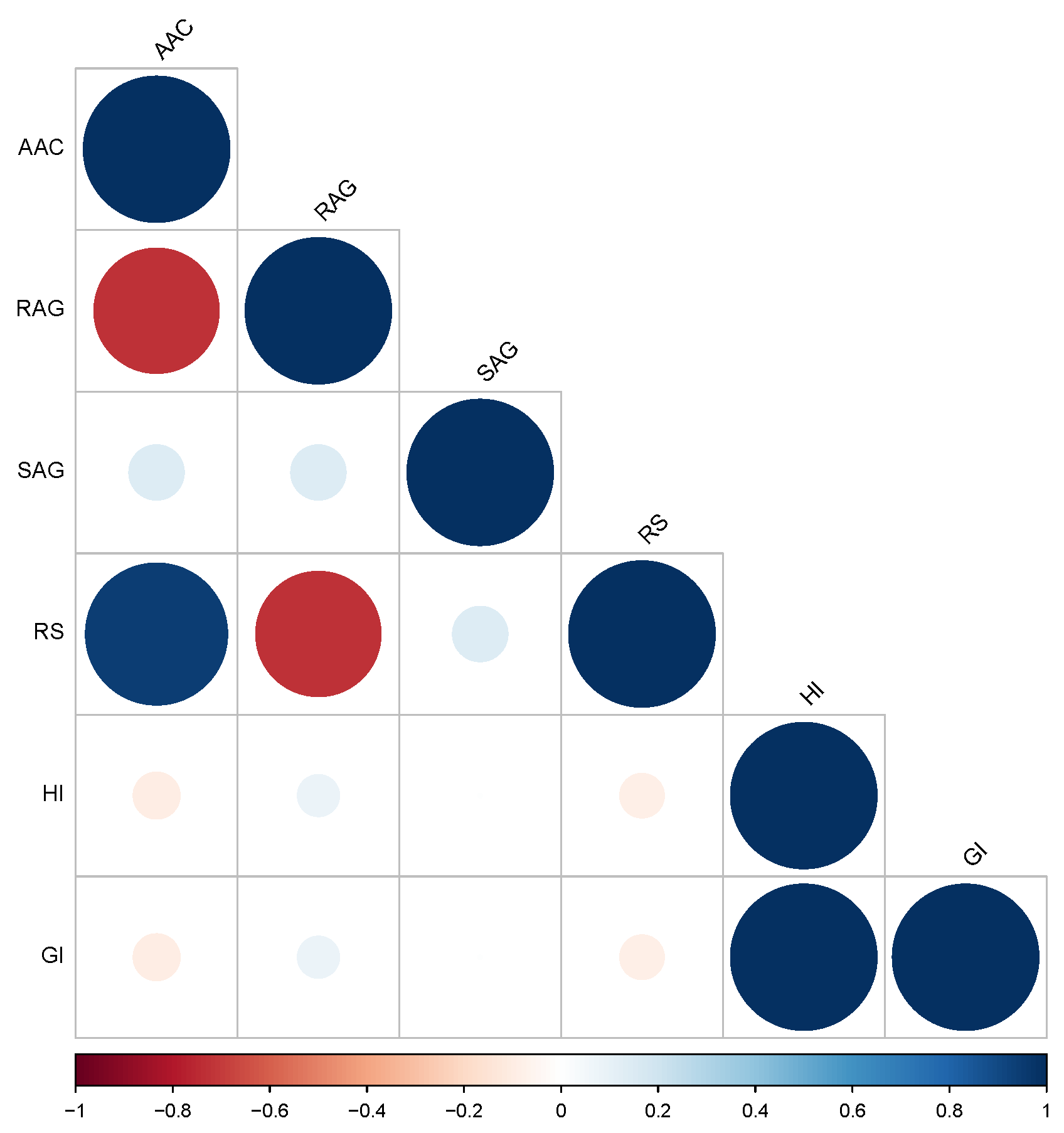
2.1. Evaluation of Phenotypic Parameters
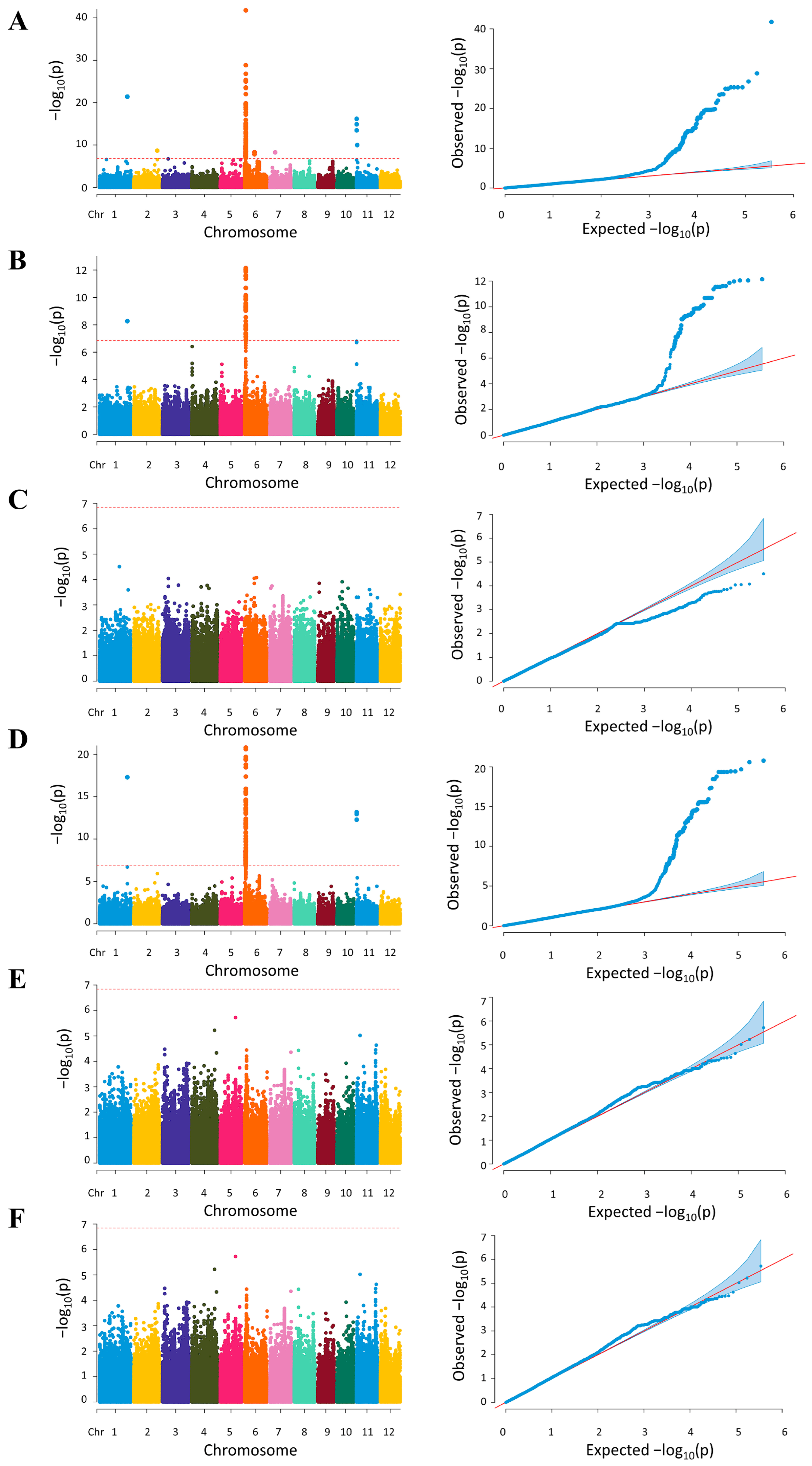
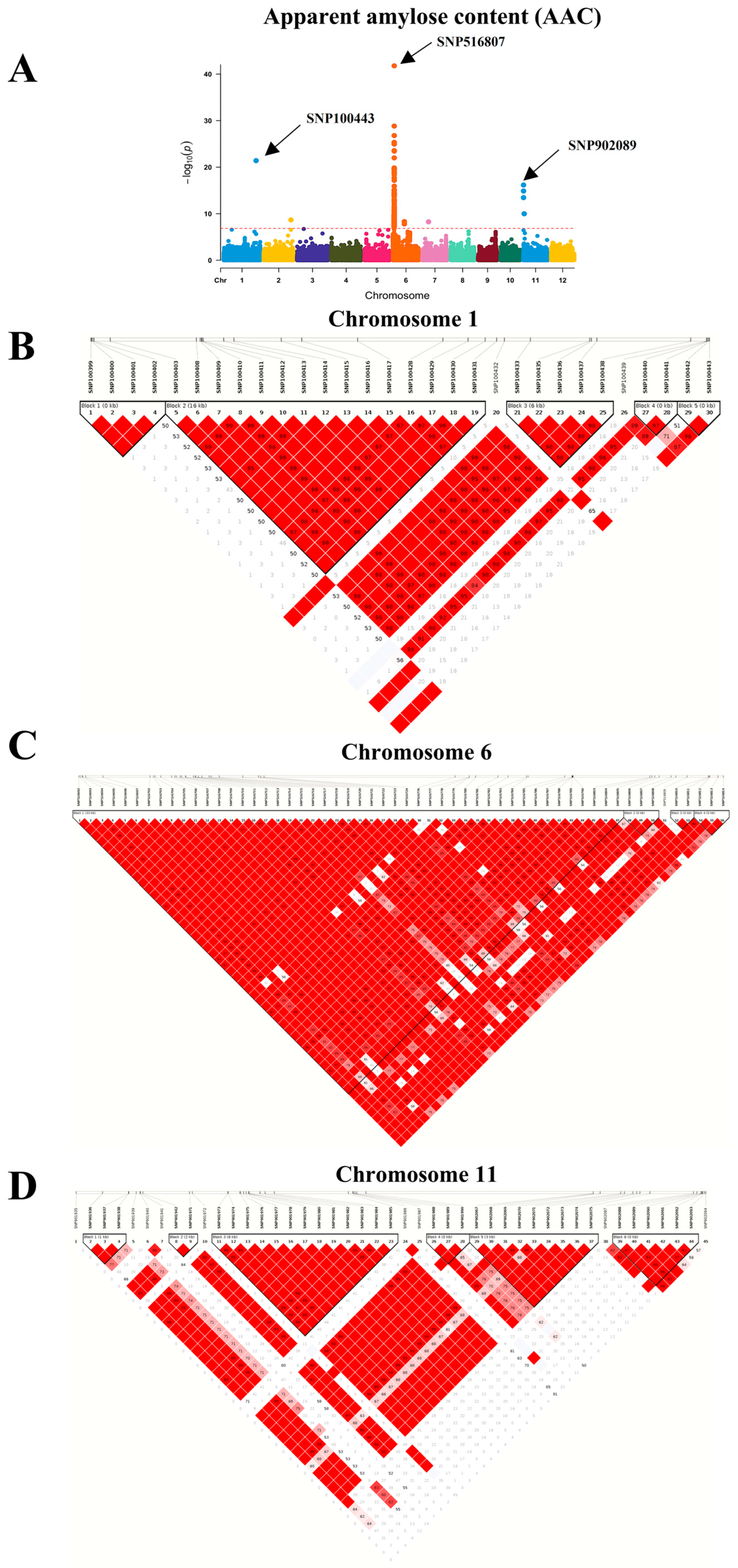
2.2. Genome Wide Association of Starch Traits in Thai Rice Populations
2.2.1. Association Mapping
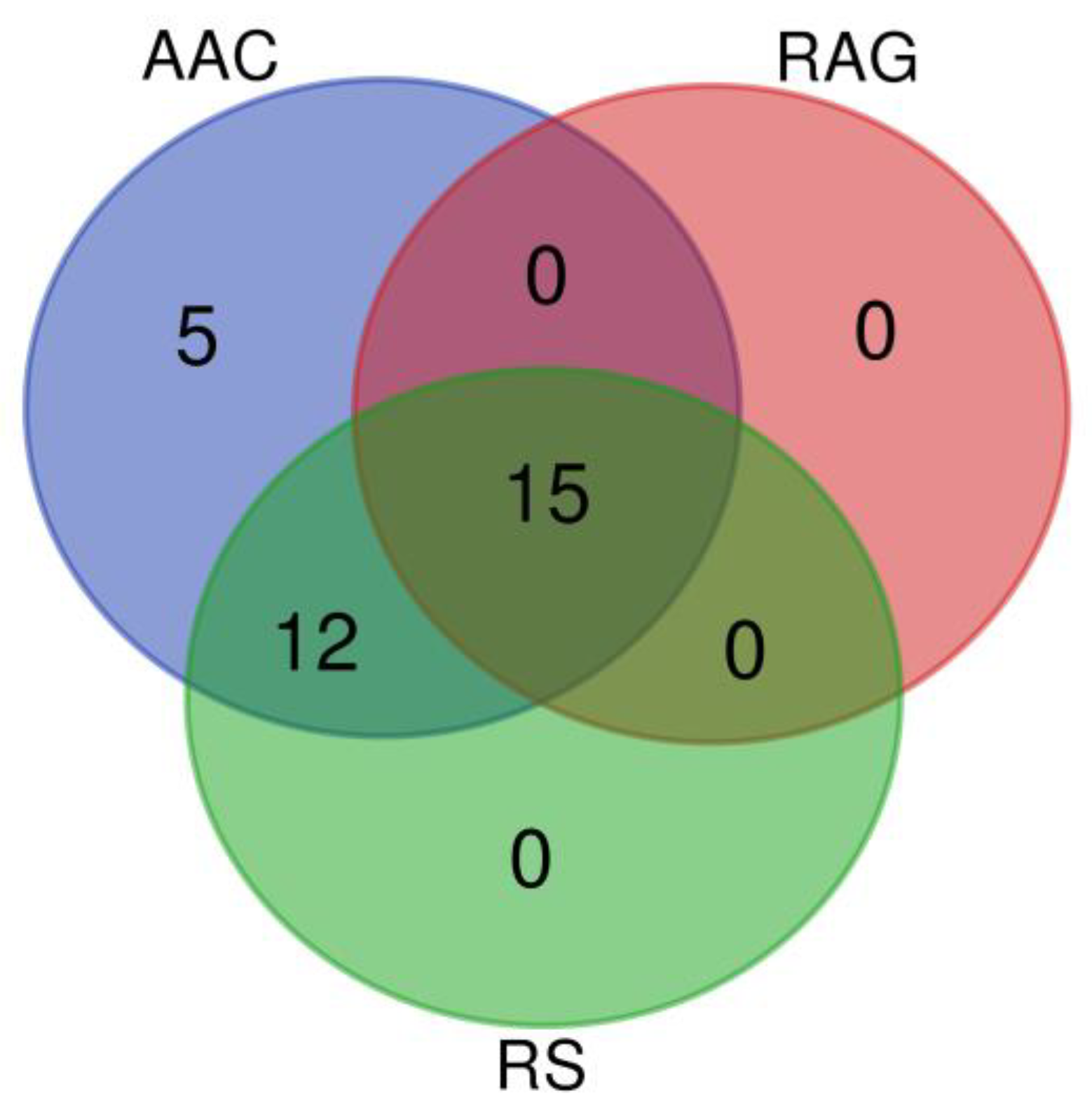
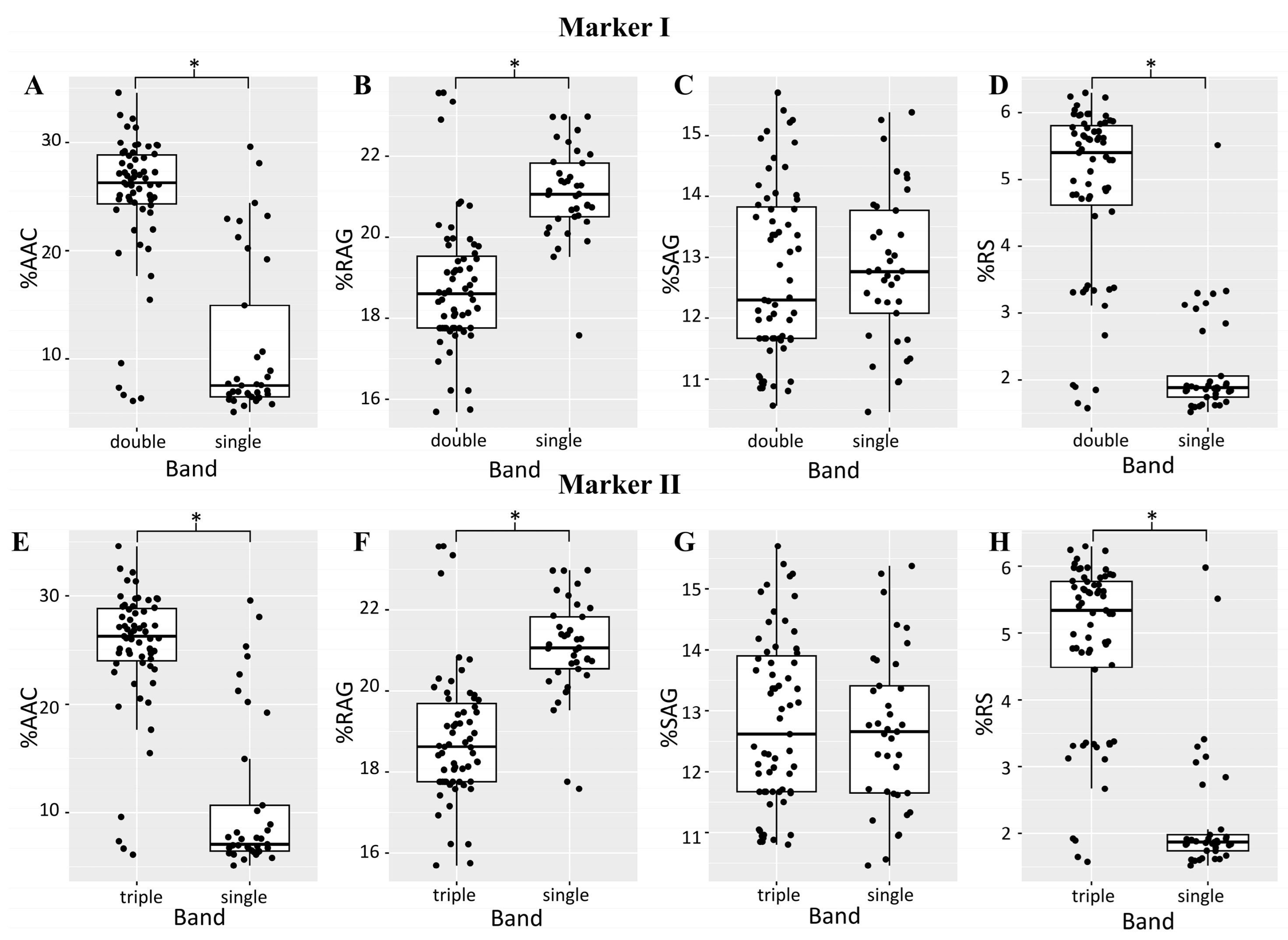
2.2.2. Molecular Marker Analysis
3. Discussion
3.1. Evaluation of Rice Phenotypes
3.2. Association Mapping and Molecular Marker Analysis
4. Materials and Methods
4.1. Plant Materials and DNA Extraction
4.2. Evaluation of Phenotypic Parameters
4.2.1. Apparent Amylose Content (AAC)
4.2.2. Digestible Starch Measurement
4.2.3. Glycemic Index
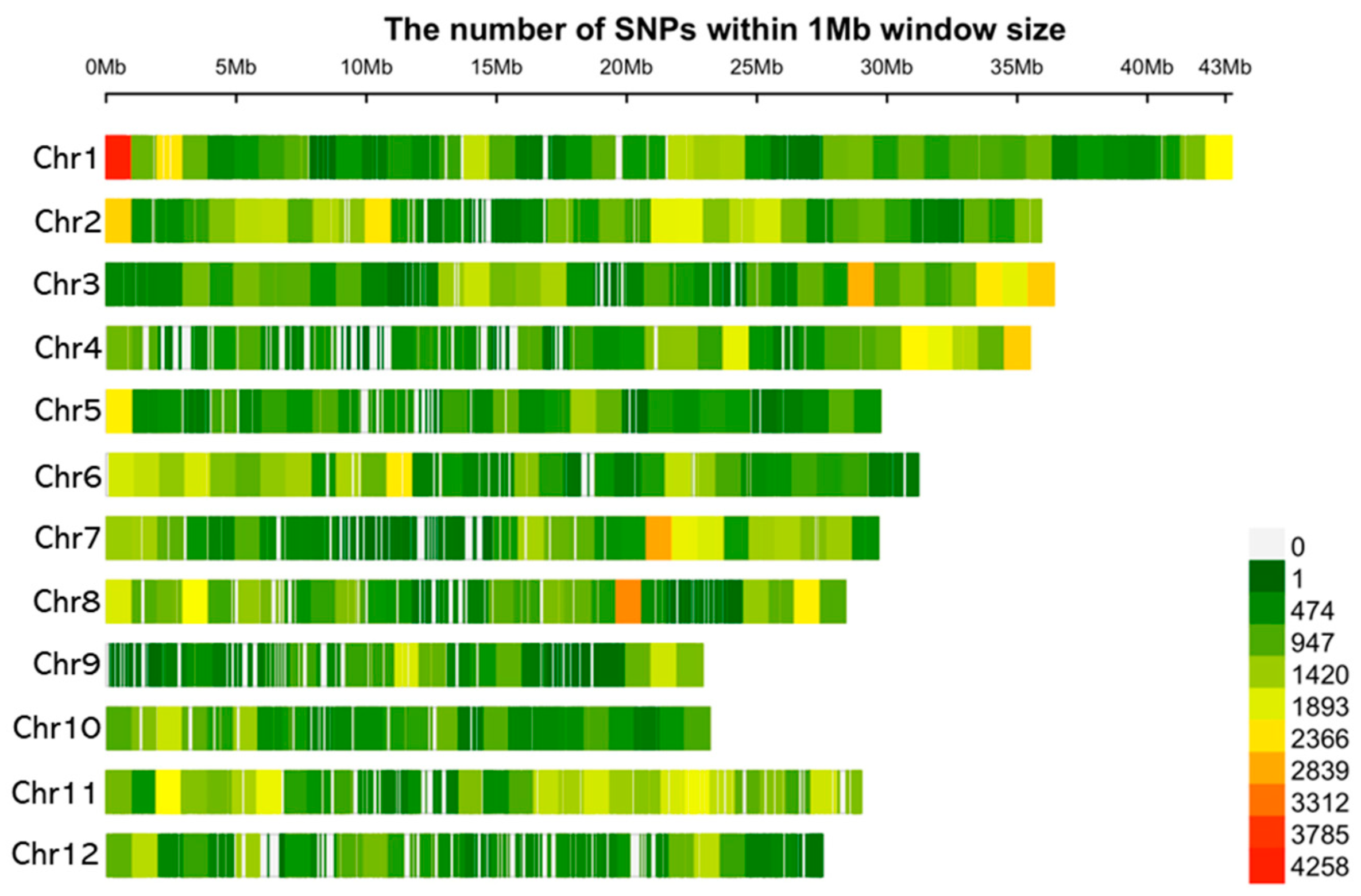
4.3. Analysis of Genome-Wide Association and Molecular Markers
4.3.1. Association Mapping and Candidate Gene Analysis
4.3.2. Molecular Marker Analysis
4.4. Statistical Analysis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, R.; Karuranga, S.; Malanda, B.; Saeedi, P.; Basit, A.; Besançon, S.; Bommer, C.; Esteghamati, A.; Ogurtsova, K.; Zhang, P.; et al. Global and regional estimates and projections of diabetes-related health expenditure: Results from the international diabetes federation diabetes atlas, 9th edition. Diabetes Res. Clin. Pract. 2020, 162, 108072. [Google Scholar] [CrossRef] [PubMed]
- Keenan, M.J.; Zhou, J.; Hegsted, M.; Pelkman, C.; Durham, H.A.; Coulon, D.B.; Martin, R.J. Role of resistant starch in improving gut health, adiposity, and insulin resistance. Adv. Nutr. 2015, 6, 198–205. [Google Scholar] [CrossRef]
- Nishida, C.; Uauy, R.; Kumanyika, S.; Shetty, P. The joint WHO/FAO expert consultation on diet, nutrition and the prevention of chronic diseases: Process, product and policy implications. Public Health Nutr. 2004, 7, 245–250. [Google Scholar] [CrossRef]
- Li, Q.; Liu, X.; Zhang, C.; Jiang, L.; Jiang, M.; Zhong, M.; Fan, X.; Gu, M.; Liu, Q. Rice soluble starch synthase I: Allelic variation, expression, function, and interaction with Waxy. Front. Plant Sci. 2018, 9, 1591. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. Rice in Human Nutrition; FAO Food and Nutrition Series, No.26; The International Rice Research Institute (IRRI): Los Baños, Philippines, 1993; pp. 43–44. [Google Scholar]
- Juliano, B.O. Rice starch properties and grain quality. J. Jpn. Soc. Starch Sci. 1992, 39, 11–21. [Google Scholar] [CrossRef]
- Englyst, H.; Kingman, S.; Cummings, J. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46 (Suppl. 2), S33–S50. [Google Scholar] [PubMed]
- Englyst, H.N.; Veenstra, J.; Hudson, G.J. Measurement of rapidly available glucose (RAG) in plant foods: A potential in vitro predictor of the glycaemic response. Br. J. Nutr. 1996, 75, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Xu, F.; Park, Y.; Zhou, X.; He, Q. Genome-wide association study of the resistant starch content in rice grains. Starch 2017, 69, 1600343. [Google Scholar] [CrossRef]
- Kumar, A.; Panda, P.A.; Lal, M.K.; Ngangkham, U.; Sahu, C.; Soren, K.R.; Subudhi, H.N.; Samantaray, S.; Sharma, S. Addition of pulses, cooking oils, and vegetables enhances resistant starch and lowers the glycemic index of rice (Oryza sativa L.). Starch Stärke 2020, 72, 1900081. [Google Scholar] [CrossRef]
- Kaur, B.; Ranawana, V.; Henry, J. The glycemic index of rice and rice products: A review and table of GI values. Crit. Rev. Food Sci. Nutr. 2016, 56, 215–236. [Google Scholar] [CrossRef]
- Qu, J.; Xu, S.; Zhang, Z.; Chen, G.; Zhong, Y.; Liu, L.; Zhang, R.; Xue, J.; Guo, D. Evolutionary, structural and expression analysis of core genes involved in starch synthesis. Sci. Rep. 2018, 8, 12736. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, M.J.; Chantler, S.E.; Okita, T.W. N- and C-terminal peptide sequences are essential for enzyme assembly, allosteric, and/or catalytic properties of ADP-glucose pyrophosphorylase. Plant J. 1998, 14, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Siedlecka, A.; Ciereszko, I.; Mellerowicz, E.; Martz, F.; Chen, J.; Kleczkowski, L.A. The small subunit ADP-glucose pyrophosphorylase (ApS) promoter mediates okadaic acid-sensitive uidA expression in starch-synthesizing tissues and cells in Arabidopsis. Planta 2003, 217, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yang, C.; Zhu, J.; Zhang, L.; Bai, Y.; Li, E.; Gilbert, R.G. Competition between granule bound starch synthase and starch branching enzyme in starch biosynthesis. Rice 2019, 12, 96. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Feng, B.; Xu, Z.; Sestili, F.; Zhao, G.; Xiang, C.; Lafiandra, D.; Wang, T. Identification and characterization of granule bound starch synthase I (GBSSI) gene of tartary buckwheat (Fagopyrum tataricum Gaertn.). Gene 2014, 534, 229–235. [Google Scholar] [CrossRef]
- Tetlow, I.J.; Emes, M.J. A review of starch-branching enzymes and their role in amylopectin biosynthesis. IUBMB Life 2014, 66, 546–558. [Google Scholar] [CrossRef]
- Biselli, C.; Cavalluzzo, D.; Perrini, R.; Gianinetti, A.; Bagnaresi, P.; Urso, S.; Orasen, G.; Desiderio, F.; Lupotto, E.; Cattivelli, L.; et al. Improvement of marker-based predictability of apparent amylose content in japonica rice through GBSSI allele mining. Rice 2014, 7, 1. [Google Scholar] [CrossRef]
- Dobo, M.; Ayres, N.; Walker, G.; Park, W.D. Polymorphism in the GBSS gene affects amylose content in US and European rice germplasm. J. Cereal Sci. 2010, 52, 450–456. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, J.; Chen, S.; Fan, X.; Li, Q.; Lu, Y.; Wang, M.; Yu, H.; Yi, C.; Tang, S.; et al. Wxlv, the ancestral allele of rice Waxy gene. Mol. Plant. 2019, 12, 1157–1166. [Google Scholar] [CrossRef]
- Kong, X.; Chen, Y.; Zhu, P.; Sui, Z.; Corke, H.; Bao, J. Relationships among genetic, structural, and functional properties of rice starch. J. Agric. Food Chem. 2015, 63, 6241–6248. [Google Scholar] [CrossRef]
- Anacleto, R.; Badoni, S.; Parween, S.; Butardo, V.M., Jr.; Misra, G.; Cuevas, R.P.; Kuhlmann, M.; Trinidad, T.P.; Mallillin, A.C.; Acuin, C.; et al. Integrating a genome-wide association study with a large-scale transcriptome analysis to predict genetic regions influencing the glycaemic index and texture in rice. Plant Biotechnol. J. 2019, 17, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Biselli, C.; Volante, A.; Desiderio, F.; Tondelli, A.; Gianinetti, A.; Finocchiaro, F.; Taddei, F.; Gazza, L.; Sgrulletta, D.; Cattivelli, L.; et al. GWAS for starch-related parameters in Japonica rice (Oryza sativa L.). Plants 2019, 8, 292. [Google Scholar] [CrossRef]
- Fitzgerald, M.A.; Rahman, S.; Resurreccion, A.P.; Concepcion, J.; Daygon, V.D.; Dipti, S.S.; Kabir, K.A.; Klingner, B.; Morell, M.K.; Bird, A.R. Identification of a major genetic determinant of glycaemic index in rice. Rice 2011, 4, 66–74. [Google Scholar] [CrossRef]
- Jiang, C.; Rashid, M.; Zhang, Y.; Zhao, Y.; Pan, Y. Genome wide association study on development and evolution of glutinous rice. BMC Genom. Data 2022, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Butardo, V.M., Jr.; Anacleto, R.; Parween, S.; Samson, I.; de Guzman, K.; Alhambra, C.M.; Misra, G.; Sreenivasulu, N. Systems genetics identifies a novel regulatory domain of amylose synthesis. Plant Physiol. 2017, 173, 887–906. [Google Scholar] [CrossRef] [PubMed]
- Misra, G.; Badoni, S.; Domingo, C.J.; Cuevas, R.P.O.; Llorente, C.; Mbanjo, E.G.N.; Sreenivasulu, N. Deciphering the genetic architecture of cooked rice texture. Front. Plant Sci. 2018, 9, 1405. [Google Scholar] [CrossRef]
- Deng, Z.; Liu, Y.; Gong, C.; Chen, B.; Wang, T. Waxy is an important factor for grain fissure resistance and head rice yield as revealed by a genome-wide association study. J. Exp. Bot. 2022, 73, 6942–6954. [Google Scholar] [CrossRef] [PubMed]
- Quero, G.; Gutiérrez, L.; Monteverde, E.; Blanco, P.; Pérez de Vida, F.; Rosas, J.; Fernández, S.; Garaycochea, S.; McCouch, S.; Berberian, N.; et al. Genome-wide association study using historical breeding populations discovers genomic regions involved in high-quality rice. Plant Genome 2018, 11. [Google Scholar] [CrossRef]
- Yang, X.; Xia, X.; Zeng, Y.; Nong, B.; Zhang, Z.; Wu, Y.; Xiong, F.; Zhang, Y.; Liang, H.; Deng, G.; et al. Identification of candidate genes for gelatinization temperature, gel consistency and pericarp color by GWAS in rice based on SLAF-sequencing. PLoS ONE 2018, 13, 196690. [Google Scholar] [CrossRef]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Chung, H.-J.; Liu, Q.; Lee, L.; Wei, D. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents. Food Hydrocoll. 2011, 25, 968–975. [Google Scholar] [CrossRef]
- Li, N.; Guo, Y.; Zhao, S.; Kong, J.; Qiao, D.; Lin, L.; Lin, Q.; Zhang, B. Amylose content and molecular-order stability synergistically affect the digestion rate of indica rice starches. Int. J. Biol. Macromol. 2020, 144, 373–379. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, M.; Fu, J.; Shen, Y.; Ding, Y.; Wu, D.; Shu, X.; Song, W. Identifying genes for resistant starch, slowly digestible starch, and rapidly digestible starch in rice using genome-wide association studies. Genes Genomics. 2020, 42, 1227–1238. [Google Scholar] [CrossRef]
- Chelliah, R.; Chandrashekar, S.; Saravanakumar, K.; Ramakrishnan, S.R.; Rubab, M.; Daliri, E.B.; Barathikannan, K.; Tyagi, A.; Kwame Ofosu, F.; Chen, X.; et al. Effect of rice processing towards lower rapidly available glucose (RAG) favors Idli, a South Indian fermented food suitable for diabetic patients. Nutrients 2019, 11, 1497. [Google Scholar] [CrossRef]
- Englyst, K.N.; Englyst, H.N.; Hudson, G.J.; Cole, T.J.; Cummings, J.H. Rapidly available glucose in foods: An in vitro measurement that reflects the glycemic response. Am. J. Clin. Nutr. 1999, 69, 448–454. [Google Scholar] [CrossRef]
- Chatterjee, M.; Berbezy, P.; Vyas, D.; Coates, S.; Barsby, T. Reduced expression of a protein homologous to glycogenin leads to reduction of starch content in Arabidopsis leaves. Plant Sci. 2005, 168, 501–509. [Google Scholar] [CrossRef]
- Willats, W.G.; Orfila, C.; Limberg, G.; Buchholt, H.C.; van Alebeek, G.J.; Voragen, A.G.; Marcus, S.E.; Christensen, T.M.; Mikkelsen, J.D.; Murray, B.S.; et al. Modulation of the degree and pattern of methylesterification of pectic homogalacturonan in plant cell walls. Implications for pectin methyl esterase action, matrix properties, and cell adhesion. J. Biol. Chem. 2001, 276, 19404–19413. [Google Scholar] [CrossRef] [PubMed]
- Ayres, N.M.; McClung, A.M.; Larkin, P.D.; Bligh, H.F.J.; Jones, C.A.; Park, W.D. Microsatellites and a single-nucleotide polymorphism differentiate apparent amylose classes in an extended pedigree of US rice germ plasm. Theor. Appl. Genet. 1997, 94, 773–781. [Google Scholar] [CrossRef]
- Sharma, R.; Cao, P.; Jung, K.H.; Sharma, M.K.; Ronald, P.C. Construction of a rice glycoside hydrolase phylogenomic database and identification of targets for biofuel research. Front. Plant Sci. 2013, 4, 330. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Kumar, R.; Tripathi, A.K.; Gupta, B.K.; Pareek, A.; Singla-Pareek, S.L. Genome-wide investigation and expression analysis of sodium/calcium exchanger gene family in rice and Arabidopsis. Rice 2015, 8, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, Z.; Wei, J.; Zhao, Z.; Sun, D.; Cui, S. A Na+/Ca2+ exchanger-like protein (AtNCL) involved in salt stress in Arabidopsis. J. Biol. Chem. 2012, 287, 44062–44070. [Google Scholar] [CrossRef] [PubMed]
- Juliano, B.O. A simplified assay for milled-rice amylose. Cereal Sci. Today 1971, 16, 334–360. [Google Scholar]
- Kumar, A.; Sahoo, U.; Baisakha, B.; Okpani, O.A.; Ngangkham, U.; Parameswaran, C.; Basak, N.; Kumar, G.; Sharma, S.G. Resistant starch could be decisive in determining the glycemic index of rice cultivars. J. Cereal Sci. 2018, 79, 348–353. [Google Scholar] [CrossRef]
- Goni, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Locus | Position | p-Value | Description | Parameter | Publication |
|---|---|---|---|---|---|---|
| 1 | LOC_Os01g65810 | 38218116 | 4.04 × 10−22 | expressed protein | AAC, RAG, RS | - |
| 2 | LOC_Os02g52950 | 32380686 | 2.16 × 10−9 | expressed protein | AAC | - |
| 6 | LOC_Os06g03960 | 1602299 | 7.05 × 10−9 | expressed protein | AAC, RS | [25] |
| 6 | LOC_Os06g03970 | 1611971 | 3.55 × 10−9 | receptor-like protein kinase 5 precursor | AAC, RS | [22,25] |
| 6 | LOC_Os06g03980 | 1616444 | 7.82 × 10−10 | expressed protein | AAC, RS | [22,25] |
| 6 | LOC_Os06g03990 | 1629652 | 1.96 × 10−10 | aminotransferase, classes I and II, domain-containing protein | AAC, RS | [22,25] |
| 6 | LOC_Os06g04010 | 1638628 | 1.85 × 10−10 | GAGA-binding protein | AAC, RS | [22,25,26] |
| 6 | LOC_Os06g04020 | 1641473 | 1.96 × 10−10 | histone H1 | AAC, RS | [22,25] |
| 6 | LOC_Os06g04030 | 1644780 | 1.96 × 10−10 | histone H3 | AAC, RS | [22,25] |
| 6 | LOC_Os06g04040 | 1650499 | 1.96 × 10−10 | WD domain, G-beta repeat domain-containing protein | AAC, RS | [22,25] |
| 6 | LOC_Os06g04070 | 1677364 | 2.97 × 10−9 | pyridoxal-dependent decarboxylase protein | AAC, RS | [22] |
| 6 | LOC_Os06g04080 | 1691152 | 7.83 × 10−10 | glycosyl hydrolases family 17, putative, expressed | AAC, RS | [22,26] |
| 6 | LOC_Os06g04090 | 1702532 | 8.59 × 10−14 | no apical meristem protein | AAC, RS | [22] |
| 6 | LOC_Os06g04130 | 1730335 | 1.32 × 10−15 | lung seven transmembrane domain-containing protein | AAC, RAG, RS | [22,25] |
| 6 | LOC_Os06g04140 | 1731629 | 2.30 × 10−11 | expressed protein | AAC, RAG, RS | [22] |
| 6 | LOC_Os06g04150 | 1733997 | 4.88 × 10−15 | magnesium-protoporphyrin O-methyltransferase | AAC, RAG, RS | [22,25] |
| 6 | LOC_Os06g04169 | 1740167 | 3.56 × 10−15 | hydrolase, alpha/beta fold family domain-containing protein | AAC, RAG, RS | [22,25,26,27] |
| 6 | LOC_Os06g04190 | 1755645 | 1.89 × 10−20 | rad1 | AAC, RAG, RS | [22,25] |
| 6 | LOC_Os06g04195 | 1761270 | 1.44 × 10−20 | expressed protein | AAC, RAG, RS | [25] |
| 6 | LOC_Os06g04200 | 1767006 | 1.69 × 10−42 | starch synthase | AAC, RAG, RS | [22,25,26,27,28,29,30] |
| 6 | LOC_Os06g04210 | 1776682 | 3.89 × 10−19 | expressed protein | AAC, RAG, RS | [28] |
| 6 | LOC_Os06g04220 | 1783512 | 4.81 × 10−26 | expressed protein | AAC, RAG, RS | [22,26,28] |
| 6 | LOC_Os06g04230 | 1784113 | 1.63 × 10−27 | expressed protein | AAC, RAG, RS | [22,25,28] |
| 6 | LOC_Os06g04240 | 1786344 | 2.83 × 10−24 | expressed protein | AAC, RAG, RS | [22,25,28] |
| 6 | LOC_Os06g04250 | 1791200 | 2.29 × 10−13 | phosphate-induced protein 1 conserved region domain-containing protein | AAC, RAG, RS | [22,28] |
| 6 | LOC_Os06g04270 | 1812542 | 9.66 × 10−12 | transketolase, chloroplast precursor | AAC, RAG, RS | [25] |
| 6 | LOC_Os06g04280 | 1816427 | 8.21 × 10−12 | 3-phosphoshikimate 1-carboxyvinyltransferase, chloroplast precursor | AAC, RAG, RS | [22] |
| 6 | LOC_Os06g04300 | 1822237 | 3.86 × 10−9 | tRNA 2-phosphotransferase 1 | AAC | [22,25] |
| 6 | LOC_Os06g23530 | 13728922 | 4.94 × 10−9 | pre-mRNA-splicing factor ATP-dependent RNA helicase | AAC | - |
| 7 | LOC_Os07g12780 | 7314685 | 5.56 × 10−9 | cyclin | AAC | - |
| 11 | LOC_Os11g01580 | 334530 | 6.91 × 10−17 | sodium/calcium exchanger protein | AAC, RS | - |
| 11 | LOC_Os11g03130 | 1129093 | 1.04 × 10−10 | E2F-related protein | AAC | - |
| % AAC | % RAG | % SAG | % RS | |||||
|---|---|---|---|---|---|---|---|---|
| R2 | p-Value | R2 | p-Value | R2 | p-Value | R2 | p-Value | |
| Pop. I (Table S7) | ||||||||
| Marker I | 0.82 | 1.4 × 10−38 | 0.04 | 4.7 × 10−2 | 0.00 | 0.69 | 0.70 | 1.1 × 10−26 |
| Marker II | 0.85 | 2.8 × 10−42 | 0.06 | 1.6 × 10−2 | 0.00 | 0.96 | 0.81 | 1.3 × 10−16 |
| Pop. II (Table S8) | ||||||||
| Marker I | 0.50 | 2.9 × 10−16 | 0.36 | 2.9 × 10−11 | 0.00 | 0.71 | 0.57 | 7.0 × 10−20 |
| Marker II | 0.54 | 2.5 × 10−18 | 0.34 | 2.0 × 10−10 | 0.00 | 0.60 | 0.55 | 1.5 × 10−18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Praphasanobol, P.; Purnama, P.R.; Junbuathong, S.; Chotechuen, S.; Moung-Ngam, P.; Kasettranan, W.; Paliyavuth, C.; Comai, L.; Pongpanich, M.; Buaboocha, T.; et al. Genome-Wide Association Study of Starch Properties in Local Thai Rice. Plants 2023, 12, 3290. https://doi.org/10.3390/plants12183290
Praphasanobol P, Purnama PR, Junbuathong S, Chotechuen S, Moung-Ngam P, Kasettranan W, Paliyavuth C, Comai L, Pongpanich M, Buaboocha T, et al. Genome-Wide Association Study of Starch Properties in Local Thai Rice. Plants. 2023; 12(18):3290. https://doi.org/10.3390/plants12183290
Chicago/Turabian StylePraphasanobol, Parama, Putut Rakhmad Purnama, Supaporn Junbuathong, Somsong Chotechuen, Peerapon Moung-Ngam, Waraluk Kasettranan, Chanita Paliyavuth, Luca Comai, Monnat Pongpanich, Teerapong Buaboocha, and et al. 2023. "Genome-Wide Association Study of Starch Properties in Local Thai Rice" Plants 12, no. 18: 3290. https://doi.org/10.3390/plants12183290