

An Efficient Transformation System for Fast Production of VcCHS Transgenic Blueberry Callus and Its Expressional Analysis

1
State Key Laboratory of Efficient Production of Forest Resources, Key Laboratory of Forest Silviculture and Conservation of the Ministry of Education, The College of Forestry, Beijing Forestry University, Beijing 100083, China
2
College of Life and Health, Dalian University, Dalian 116000, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Plants 2023, 12(16), 2905; https://doi.org/10.3390/plants12162905
Submission received: 17 July 2023
/
Revised: 6 August 2023
/
Accepted: 7 August 2023
/
Published: 9 August 2023
(This article belongs to the Special Issue Advances in Blueberry Research)
Abstract
:The Agrobacterium tumefaciens-mediated transformation for blueberries remains less efficient than is desirable. A new leaf callus regeneration and genetic transformation system was investigated in blueberries in this study. The leaf explants of cv. ‘Legacy’ and ‘Northland’ were used to establish the stable callus induction system when placed on the woody plant medium (WPM) supplemented with 1.0 mg·L−1 2, 4-D, 0.4 mg·L−1 6-BA for 30 d; then, the callus was sub-cultured in the proliferation medium supplemented with 1.5 mg·L−1 2, 4-D, 0.4 mg·L−1 6-BA in the darkness at 25 °C every 30 days. The co-cultivation of callus with A. tumefaciens was operated on WPM plus 100 μM acetosyringone for 4 days; then, the transferred callus was grown in WPM supplemented with 1.5 mg·L−1 2,4-D, 0.4 mg·L−1 6-BA, 50 mg·L−1 hygromycin, and 200 mg·L−1 cefotaxime. The VcCHS transgenic blueberry callus with both GFP signal and Hyg resistance was obtained from the transformed callus of cv. ‘Northland’. The rate of GFP signal detected in the transformed callus was as high as 49.02%, which was consistent with the PCR assay. Collectively, this study provides a highly efficient genetic transformation system in blueberry callus and a powerful approach for the molecular breeding of blueberries.
1. Introduction
Highbush blueberry (Vaccinium corymbosum L.), an important economically woody plant species known for its fruits, contains high amounts of antioxidants, anthocyanins, and various nutrients that have significant health benefits for the human body, which has driven industrial development around the world [1,2,3,4,5].
The process of breeding blueberries remains limited, due to high heterozygosity and the difficulty of regaining special traits from parent plants, as traditional methods are time-consuming and inefficient, although blueberry genetic improvement by genetic transformation methods is suitable [6,7,8,9]. The Agrobacterium tumefaciens-mediated transformation method has been extensively used in plant transformation [10]. It provides a powerful approach to increase breeding efficiency and improves the economic traits of blueberries, including fruit quality, yields, nutrient content, stress resistance, and the flowering phase [11,12,13,14,15,16,17,18]. To date, adventitious shoot regeneration systems and A. tumefaciens-mediated transformation methods of leaf explants have been reported for several blueberry cultivars. Some blueberry transgenic lines have shown the ability to survive low temperatures or to flower precociously or persistently [13,17]. High concentrations of common cytokines zeatin (ZT) and 6-(γ,γ-Dimethylallylamino) purine (2-ip) can induce shoot regeneration from leaf explants of cv. ‘Aurora’, ‘Brigitta’, ‘Elliott’, ‘Bluecrop’, and ‘Legacy’ [19]. In addition, thidiazuron (TDZ) and a-naphthaleneacetic (NAA) are commonly used to induce adventitious bud regeneration from blueberry leaves [20]. The shoot regeneration medium supplemented with modified WPM (reported by Rowland and Ogden in 1992), 1.0 mg·L−1 TDZ, and 0.5 mg·L−1 NAA was regarded as the best combination for leaf explants of most cultivars of highbush blueberry [19]. The combination of ZT and IBA has been demonstrated to effectively induce buds from callus derived from certain blueberry cultivars, particularly cv. ‘Red Button’, with the highest rates of callus induction and browning being 90.3% and 33.3%, respectively, which are influenced by genotype [21]. This phenomenon has also been observed in other research that used different types or concentrations of plant growth regulators [19,20]. Moreover, 83.33% of callus showed green, fluorescent signals, while only 12.82% efficiency was achieved in obtaining transgenic buds from the induced callus [21]. Despite the combination of different plant growth regulators for shoot regeneration systems and subsequent transformation techniques, such as A. tumefaciens-mediated transformation methods, being carried out in several blueberry cultivars, all these transformations involve inducing transgenic buds from leaf discs, which is time-consuming and requires several years for plant cultivation before conducting research on fruit quality and flowering [17,19,21]. Furthermore, these methods have not yet proven to be highly efficient approaches to transform genes of interest, as reported transgenic plant acquisition rates range between 5–12.82%, influenced by factors such as genotype, co-cultivation time, A. tumefaciens strain and concentration, explant age, and regeneration system [19,21,22]. Additionally, blueberries, being woody plants, require a long period to stably bear fruit, which limits further research on molecular mechanisms of fruit development, fruit ripening, anthocyanin accumulation, abiotic stress, and other basic research.
The callus induction and transformation system has been widely applied to fruit trees such as citrus, apple, chestnut, pear, etc., due to the advantages of time saving, fast growth, and high efficiency [23,24,25,26]. Common explants used for callus induction include mature seeds, immature embryos, leaves, stem segments, anther, etc. [27,28,29,30]. Frequently, plant growth regulators, such as 2, 4-D, 6-BA, TDZ, NAA, etc., are added to induction mediums [31,32,33]. At present, the mechanism of VcMYB4a in abiotic stress verified in blueberry callus only refers to apple callus transformation methods [16]. However, the callus regeneration and transformation system has not been reported in blueberries. Therefore, it is necessary to establish a highly efficient induction and transformation system for blueberry callus.
Anthocyanin is an important secondary metabolite in the flavonoid family which can attract pollinators and protect plants from visible light and UV-B radiation [34,35]. It is abundant in blueberries, which have antioxidant activities and protective effects on the cardiovascular system. The chalcone synthetase (CHS) gene provides a prerequisite for anthocyanin synthesis and is an indispensable substance in the synthesis process [36]. Moreover, chalcone is upstream in the flavonoid synthesis pathway, which can be induced by UV-B and low temperature, and the catalytic reaction of chalcone synthetase is an important rate-limiting step of the flavonoid synthesis pathway [34,37]. CHS is a key enzyme gene in the anthocyanin biosynthesis pathway. It has been reported that high light (HL) stress, causing red coloration of leaves, can enhance CHS gene expression levels in CHS-overexpression lines to improve plant adaptability to HL [38].
In this study, we established a stable callus regeneration system for leaf explants of blueberry. Then, we also concentrated on several crucial factors, including hygromycin sensitivity, co-cultivation time, and Agrobacterium concentration, that have been reported to significantly influence A. tumefaciens-mediated transformation efficiency. Finally, we summarized a highly efficient transformation method for leaf callus of blueberry. The new method for blueberries is reliable, improves the transformation efficiency by up to 49.02%, and supports for gene functional research and molecular breeding.
2. Results
2.1. Leaf Callus Induction and Proliferation of Blueberry cv. ‘Legacy’
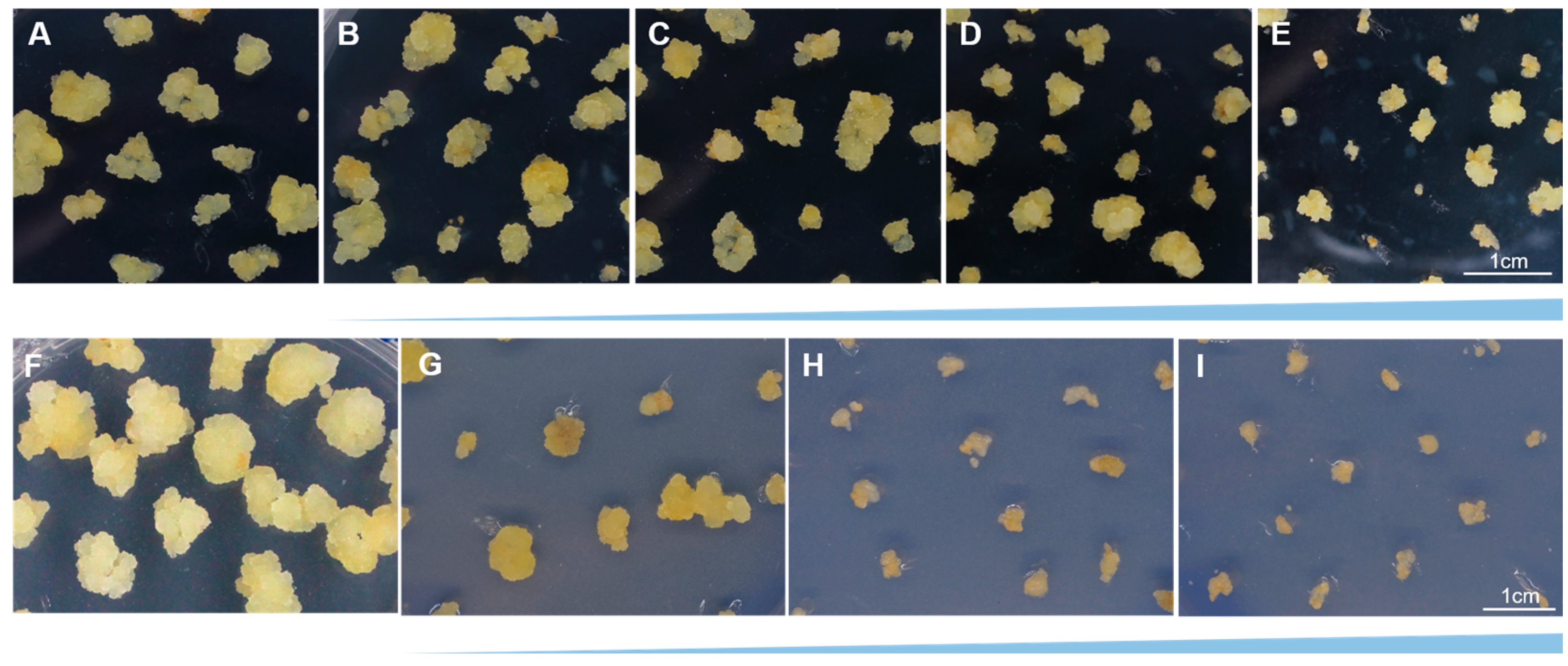
In order to find out the suitable callus induction medium for leaf explants of cv. ‘Legacy’, the medium was added with MS or WPM separately, containing different concentrations of 2, 4-D (Figure S1 and Table S1). The leaves on MS medium produce the green and close callus, while the leaves on WPM medium induce the yellow and soft callus. To obtain more callus, leaves were cultivated on WPM medium with different concentrations of 2, 4-D. We found that 1 or 2 mg·L−1 2, 4-D efficiently induced a more yellow and soft callus. But callus in this combination grew slowly, and only a little callus was obtained for two months. When the concentration of 2, 4-D increased to 4 mg·L−1, callus growth was limited, and the color became brown and hard to induce new callus.
Therefore, we added different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1) to the WPM medium containing different concentrations of 2, 4-D (1 or 4 mg·L−1) to obtain more healthy, yellow, soft callus from leaf explants (Figure 1A–H and Table S2). We found that leaf explants induced different conditions of callus in different combinations significantly after 30 days. When the WPM medium was supplemented with 1 mg·L−1 2, 4-D and 0.2 mg·L−1 6-BA, the callus was compact and a grayish yellow (Figure 1A). When the WPM medium supplemented with 1 mg·L−1 2, 4-D and 0.4 or 0.6 mg·L−1 6-BA, the callus was yellow, soft, loose, and grew rapidly, as we harvested callus after 30 days (Figure 1B,C). When the concentration of 2, 4-D increased to 4 mg·L−1 with the supplement of 6-BA, the callus became brown and grew some white tissue (Figure 1E–H). Therefore, the WPM medium added with 1 mg·L−1 2, 4-D and 0.4 mg·L−1 6-BA was the optimum callus induction medium for cv. ‘Legacy’ and was suitable for proliferating new callus in the darkness at 25 °C for 30 days (Figure 1B).
Since there was no significant difference in inducing callus proliferate between 1 and 2 mg·L−1 for the concentration of 2, 4-D, we took the average, 1.5 mg·L−1, as the suitable concentration for callus proliferation medium (Figure S2A,B). The callus proliferation medium was supplemented with 1.5 mg·L−1 2, 4-D and different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1) (Table S3). When the concentration of 6-BA in the subculture medium increased to 0.4 mg·L−1, the callus not only became yellow, soft, and loose, but also grew quickly, and we obtained a mass of loose and healthy callus after it was sub-cultured for 30 d. Therefore, the optimum proliferation medium for callus of cv. ‘Legacy’ was WPM medium supplemented with 1.5 mg·L−1 2, 4-D and 0.4 mg·L−1 6-BA in the darkness at 25 °C every 30 days (Figure S3).
2.2. Leaf Callus Induction and Proliferation of Blueberry cv. ‘Northland’
In order to further verify whether this system is suitable for other cultivars, we conducted a verification in cv. ‘Northland’. The leaf explants of cv. ‘Northland’ were cultivated in WPM + 1 mg·L−1 2,4-D + 0.4 mg·L−1 6-BA and induced yellow, soft callus (Figure 2A). After 40 days, the leaves of cv. ‘Northland’ which had induced callus were cut into small pieces, transferred in proliferation medium, and then produced new callus for 20~30 d (Figure 2B). Then, the callus was taken out from the small pieces of leaves and transferred to new proliferation medium and proliferated quickly (Figure 2C). The results indicated that the induction medium and proliferation medium selected in this study were also suitable for cv. ‘Northland’.
2.3. Selection of Cefotaxime and Hygromycin Concentration
The bacteriostatic agent type and concentration significantly inhibited growth of explants, which was necessary to screen optimum concentration of bacteriostatic agent to reduce its influence on plant tissue growth. The Cef was chosen as the bacteriostatic agent to limit the growth of A. tumefaciens, and Hyg was used to identify the transgenic callus. In order to select the optimum cefotaxime and hygromycin concentration, callus was transferred to the selection medium containing different concentrations of Cef and Hyg. The callus grew rapidly when it was cultivated in the control group (CK) without Cef and Hyg (Figure 3).
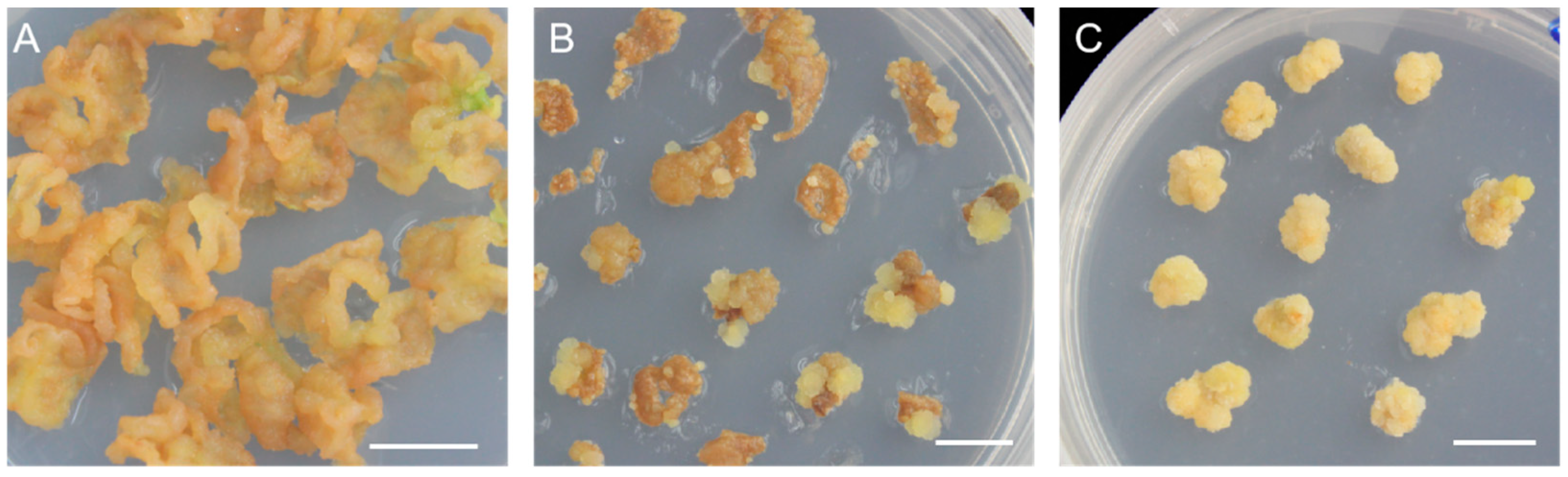
Here, we found that when the concentration of Cef was between 100 to 200 mg·L−1, callus growth was limited (Figure 3B,C). Until the concentration of Cef was between 300 to 400 mg·L−1, the callus could not grow up obviously (Figure 3D,E). In order to reduce the inhibition of callus growth, we chose 200 mg·L−1 as the optimum concentration of Cef, since the concentrations of 100~400 mg·L−1 inhibited bacteria growth effectively (Figure 3C).
When the concentration of Hyg was 30 mg·L−1, callus growth was limited and became yellowish white and loose. When the concentration of Hyg increased to 50 mg·L−1, callus growth was limited obviously, and it gradually browned and died. Therefore, we chose 50 mg·L−1 Hyg as the optimum concentration of Hyg to select for the Hyg-resistant callus (Figure 3H).
2.4. Agrobacterium-Mediated Transformation of Callus
In order to establish an Agrobacterium-mediated stable transformation for cv. ‘Northland’, we used EHA105 containing the binary vector pMDC85 to infect the callus, and nine different treatment schemes were designed for bacteria liquid concentration (value of OD600) infection time and co-cultivation time through the L9 (33) orthogonal table to select the best combination for the transformation system (Table 1).
The highest rate of GFP signal detected in the transformed callus was 49.02%, which was observed at OD600 = 0.7 after 10 min infection and 4 days’ co-cultivation (Table 1). According to the value of quadratic sum and significance (p < 0.05), the influence of different factors on the genetic transformation efficiency of the callus of cv. ‘Northland’ were sorted by size as infection time > bacteria liquid concentration > co-cultivation time through variance analysis. Infection time influenced transformation efficiency significantly, as bacteria liquid concentration and co-cultivation time had no significant effect, which indicated that infection time is the main influencing factor for transformation efficiency (Table 2). Compared with the mean value, the longer the infection time, the lower the efficiency of callus transformation. When the infection time increased to 30 min, the transformation efficiency was 7.15%, which was the lowest of all treatments (Table 3). Therefore, the optimum transformation treatment was observed at OD600 = 0.7 for bacteria liquid concentration after 10 min infection and 4 days’ co-cultivation, according to the mean value and the rate of GFP signal detected in the transformed callus.
2.5. Molecular Identification and GFP Detection for Transgenic Callus
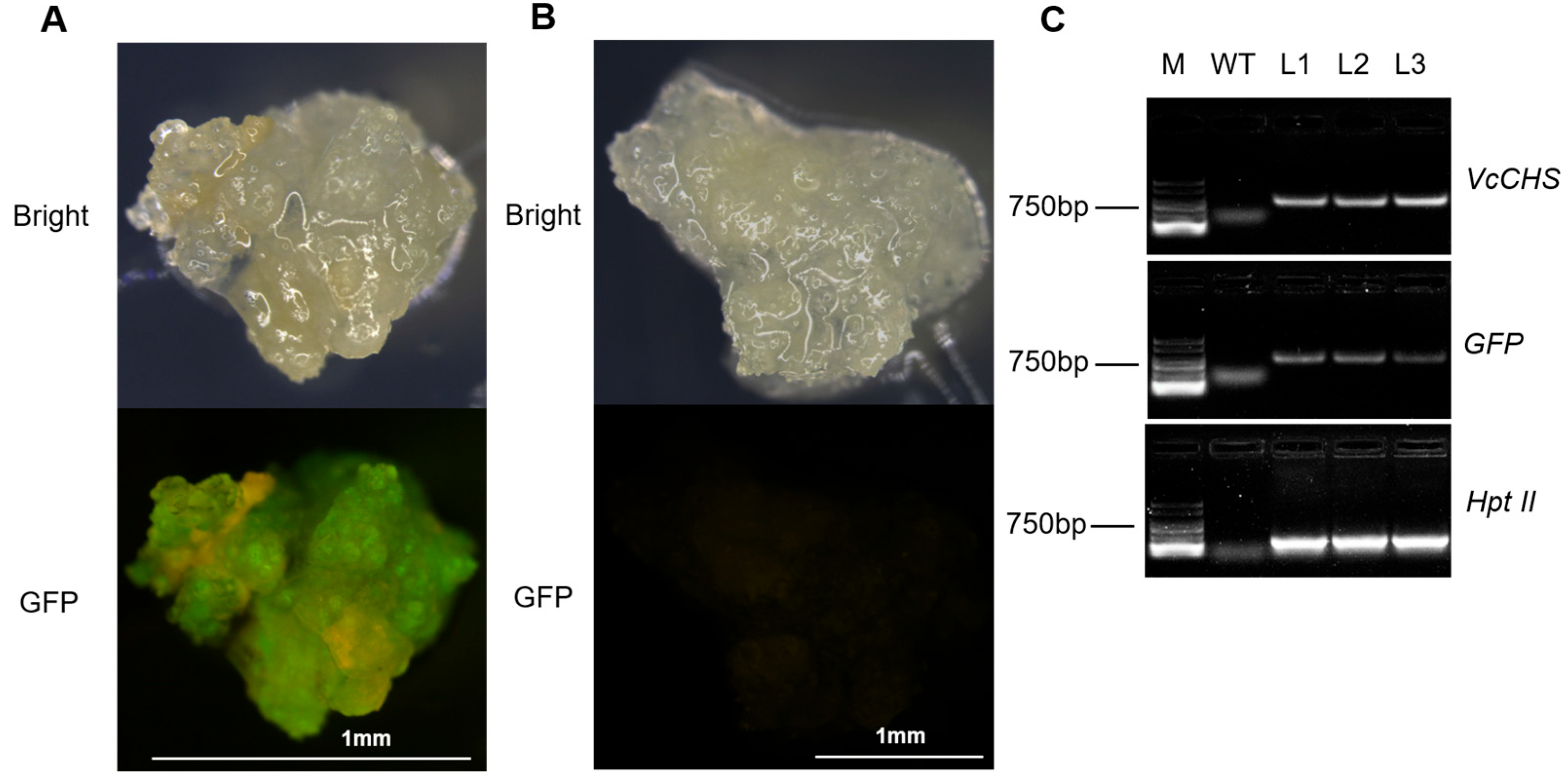
The wild-type callus (WT, non-transformed callus) and transgenic callus was identified by stereo fluorescence microscope. Obvious green fluorescence was observed in the transgenic callus, while the WT showed no green fluorescence (Figure 4A,B). In order to further identify the resistant callus by molecular techniques, callus containing GFP signal was selected to detect exogenous VcCHS and gene insertion by PCR (Figure 4C). Clear bands could be observed in the transgenic callus. The semi-quantitative detection of both callus showed that no bands were observed in the WT callus, while the expression level of VcCHS genes in the transgenic callus was 6 times that of the WT callus (Figure 5B,C).
2.6. Phenotype Analysis of Transgenic Callus
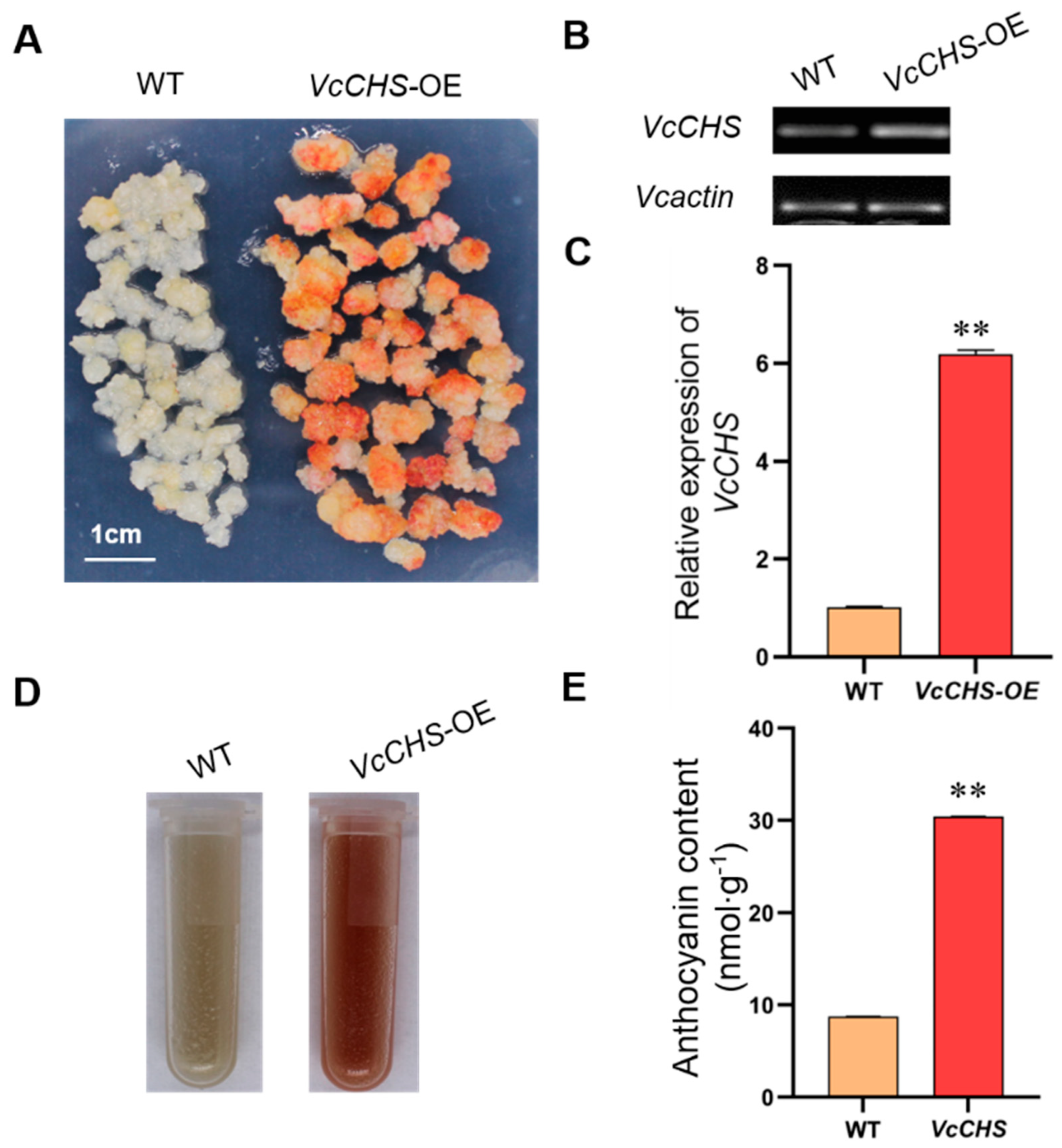
The WT callus and VcCHS transgenic callus grown for 20 days was transferred in continuous low temperatures containing white and UV light. The transgenic callus exposed to light showed a large area of coloring and red on day 5, while the WT callus showed only a small amount of dot coloring (Figure 5A,D). The analysis of anthocyanin content showed that there was a significant difference in anthocyanin content between the WT callus and overexpressed VcCHS transgenic callus, indicating that overexpressed VcCHS transgenic callus could significantly increase anthocyanin accumulation in callus of cv. ‘Northland’ (Figure 5E).
3. Discussion
Callus induction efficiency was significantly correlated with plant genotype, combination and concentration of plant growth regulators, light intensity, and culture medium [39,40,41,42]. In other plants, many studies have shown that different genotypes of the same species have significant effects on callus induction efficiency, and that even different parts of the same genotype plant have different induction efficiency [43]. In this study, the callus induction system constructed by the cv. ‘Legacy’, including culture medium, plant growth regulators, and culture condition, was found to be suitable for cv. ‘Northland’, indicating that these two plant growth regulators may have a certain universality in inducing callus of northern highbush blueberry. The leaf explants were also wounded before transformation because it was easier to induce the callus after injury [44]. In addition, reports of callus induction in other plants also showed that 2, 4-D could induce callus from different types of explants, and 6-BA had a significant effect on accelerating callus generation and propagation [45]. This experiment also verified the research that the culture medium we obtained had better effects on callus induction for two blueberry cultivars. Some plants had reported that the suitable culture medium for one genotype does not work well for others [31,46]. Therefore, whether this callus induction system is suitable for other genotypes needs to be proved by further experiments.
It was reported that the Agrobacterium strain and concentration, genotype, explant type, and co-cultivation time significantly influence transformation efficiency [47,48,49,50]. There are different kinds of antibiotics to distinguish transgenic material from explants, such as kanamycin (Km) and hygromycin (Hyg), which were frequently applied to A. tumefaciens-mediated transformation methods for some woody plants [51,52]. The tolerance concentration of wild-type materials adapted to antibiotics was the key factor to screen, since the growth of wild-type plant materials was inhibited at a certain critical concentration [53]. Therefore, screening antibiotics could effectively identify transformed plant materials and improve the screening efficiency of positive transgenic materials. The infection time and co-cultivation were also key factors. If the infection time was short, A. tumefaciens could not fully contact plant materials; if the infection time was too long, the growth of plant materials was inhibited or even caused explants to die. If the culture time was too short, the infection effect of A. tumefaciens was affected and the positive rate of the material was reduced; on the contrary, if the culture time was too long, the overgrowth of A. tumefaciens would pollute the growth of plant materials, which was difficult to clean and even led to the death of plant materials [54]. In this study, Hyg was chosen as the screening antibiotic and Cef as a bacteriostatic agent. The tolerance of cv. ‘Northland’ callus was assessed, and the concentration at which callus growth was significantly inhibited was selected as the screening concentration. When determining the concentration of Cef for screening, we aimed to select a treatment with low concentration and strong bacteriostatic effect, in order to minimize its impact on callus proliferation. Some studies have suggested that yellow callus is more suitable for genetic transformation [55]. In line with this, we obtained light yellow callus that exhibited loose structure and rapid proliferation, similar to other species used for genetic transformation. The transformation efficiency reached 49.02%, indicating significant improvement in the A. tumefaciens-mediated transformation method developed for cv. ‘Northland’ callus. Furthermore, the rate of GFP signal detected in transformed callus correlated with PCR detection results, confirming stability, reliability, and high efficiency of the transformation system in this study. Transgenic callus was successfully generated, and RT-qPCR analysis revealed a significant six-fold increase in VcCHS expression compared to wild-type.
There is no doubt that it is valuable for the transformation system of plants to obtain regenerated plantlets. However, it is also worth noting that direct plantlet generation from callus can be challenging, especially for fruit trees and woody plants, because of the high heterozygosity. On the other hand, this process requires specific hormones, conditions, and a lot of time to cultivate the transgenic plants to harvest. Compared to the transgenic seedlings that require a long period of time, A. tumefaciens-mediated callus transformation could be rapidly and directly carried out for gene function verification and phenotype study. For example, callus transformation system is commonly utilized in coloration assays to reveal the molecular mechanisms underlying anthocyanin biosynthesis, abiotic stresses, and fruit quality in apples and grapevines [56,57,58,59,60,61,62,63]. Therefore, it had unique advantages and accelerated the research on agronomic characteristics of woody plants or fruit trees. In addition, it can also be used for the synthesis of bioactive secondary metabolites through transgenic callus instead of generating transgenic plants [64]. It has been reported that high added-value plant compounds were obtained through transgenic callus [65,66]. This will provide a reference for mass synthesis of bioactive secondary metabolites that have medicinal and health values in the future.
4. Materials and Methods
4.1. Plant Materials and Callus Induction
Stem segments of highbush blueberry cv. ‘Legacy’ and ‘Northland’ were cultured at 25 °C for 30 days in order to collect the explants to obtain tissue culture seedlings. Leaf explants cultivated on the shoot stock culture medium for 45–50 d were used. To select the suitable callus induction medium for leaf explants, leaves of cv. ‘Legacy’ were wounded transversely with a scalpel and placed on woody plant medium (WPM) supplemented with 30 g·L−1 sucrose, 6.4 g·L−1 agar, and different concentrations of 2,4-D (1, 2, 4 mg·L−1) and 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1); then, the explants were cultured at 25 °C in the darkness. Callus growth condition and color were recorded after 35 days. The callus was transferred onto the callus proliferation medium supplemented with WPM, 30 g·L−1 sucrose,6.4 g·L−1 agar, 1.5 mg·L−1 2,4-D, and different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1), and cultured at 25 °C in the darkness. The callus growth condition and color were recorded after 35 days. The pH of all the medium was kept 5.2~5.5 before adding agar and autoclaving at 121 °C for 20 min. The callus was sub-cultured on the fresh proliferation medium every 4 weeks. After two subcultures in the darkness, the callus which were yellow and loose were chosen for the transformation experiment.
4.2. Selection of Antibiotics Concentration
In order to screen the transgenic callus and inhibit the growth of A. tumefaciens, wild-type callus (WT, non-transformed callus) of cv. ‘Northland’ was cultivated in selection medium containing different concentrations of Hyg (0, 30, 50, 80 mg·L−1) and Cef (0, 100, 200, 300, 400 mg·L−1) to observe the tolerance value in the darkness at 25 °C. Fifty callus clumps was used for each treatment and repeated 3 times. The callus growth condition and survival rate were recorded after 30 days.
4.3. Plasmid Construction
The binary vector pMDC85 with kanamycin resistance gene, hygromycin resistance gene, and green fluorescent protein gene (GFP) was used in this study. The genomic fragment of VcCHS was cloned with gene-cloning primers through blueberry genomic DNA. The PCR products for VcCHS were constructed into the binary vector pMDC85, and the promoter was 35S, while the expression vectors pMDC85-VcCHS were transformed into A. tumefaciens strain EHA105.
4.4. Agrobacterium-Mediated Transformation for Blueberry Callus
The A. tumefaciens strain EHA105 was cultivated overnight in YEB liquid medium supplemented with 50 mg·L−1 rifampicin and 50 mg·L−1 kanamycin at 120 rpm at 28 °C, until OD600 was 0.8~1.0. Bacteria liquid was transferred into a 50 mL tube to centrifuge at 5000 rpm for 10 min and then resuspended in WPM liquid medium with 20 g·L−1 sucrose and 100μM Acetosyringone (AS). The callus was transferred into different bacteria liquid concentrations (OD600 = 0.4, 0.7, 1.0) and infected with A. tumefaciens for 10, 20, or 30 min in the darkness, then operated in shake cultivation in 120 rpm at 28 °C. After being dried through filter paper, callus was co-cultivated on regeneration medium extra containing 100 μM AS with A. tumefaciens for 2, 3, or 4 days in the darkness. Then, callus was transferred onto the selection medium that regeneration medium supplemented with 50 mg·L−1 hygromycin and 200 mg·L−1 cefotaxime (Cef), and the selection medium were updated every 20 days. After 50 days, the resistant callus produced a new callus and continued cultivating on the selection medium, and the unsuccessfully infected callus became brown and stopped growing.
4.5. Molecular Identification of Transgenic Callus
The transgenic callus was identified by stereo fluorescence microscope with a GFP filter and observed in 450~490 nm.
The total genomic DNA of transgenic callus was isolated by the method of CTAB. The DNA of WT and binary vector pMDC85 were the control group for polymerase chain reaction (PCR). The primers corresponding to a 963 bp fragment of the coding region of VcCHS were 5′-ATGGGAATCATTCCAGAGTCTCCTCT-3′ and 5′-CTAGATCTCCCCATCCCAGAGCTCCT-3′. The PCR reaction conditions were 94 °C for 3 min, 30 cycles of 98 °C for 10 s, 60 °C for 20 s, and 72 °C for 70 s, and then 72 °C for 5 min. The products of PCR were detected in 1% agarose gel. The qRT-PCR was carried out in ABI-StepOnePlu system, the reaction conditions were followed as PowerUpTM AYBRTM Green Master Mix in ABI. The relative transcriptional expression levels of each gene were calculated by 2−ΔΔCt. The primers of VcCHS-forward were 5′-TCAACCAACGCAACGATTCCAG-3′ and VcCHS-reverse were 5′-ACAATGCTCCGACTGGTAAACG-3′. The primers of VcActin-forward were 5′-ACACGGGGAGGTGTGACAA-3′ and VcActin-reverse were 5′-CCTCCAATGGATCCTCGTTA-3′.
4.6. Anthocyanin Content Detection
The positive transgenic callus and WT callus was cultivated in a manual climatic box at 14 °C under continuous white light (light intensity of 10,000 lx) containing UV-B. After 5 days, the color of callus was recorded, and the anthocyanin of callus was extracted by using 1% (v/v) HCl-alcohol. The procedures were as follows: 0.5 g callus was ground into powder with liquid nitrogen and transferred into a 2 mL tube, adding anthocyanin extract liquid (95% alcohol + 1.5 mol·L−1 HCl) at 4 °C for 24 h in the darkness. The samples were centrifuged at 5000 rpm for 5 min and obtained supernatant, detected to have a spectrophotometric quantification of 530, 620, and 650 nm. Anthocyanin content = OD λ/ξ λ × V/M × 106, OD λ(anthocyanin density) = (A530 − A620) − 0.1×(A650 − A620), ξ λ(extinction coefficient of anthocyanin at 530 nm) = 4.62 × 104, V = extraction volume, M = callus weight. The experiments were repeated three times independently [60].
4.7. Statistical Analysis
Data for callus induction and stable transformation were analyzed for significance by analysis of Duncan’s test in ANOVA through SPSS 19.0. The figures were made by SigamPlot 2018.
Callus induction efficiency = the number of leaf explants generates callus/Total leaf explants per dish ×100%. The rate of GFP signal detected in the transformed callus (%) = the number of callus containing GFP signal/The number of transformed callus ×100%.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12162905/s1, Figure S1: Leaf growth status of cv. ‘Legacy’ in WPM or MS containing different combinations of 2,4-D; Figure S2: Callus growth status of cv. ‘Legacy’ in WPM containing different combinations of 2,4-D; Figure S3: The growth status of callus of cv. ‘Legacy’ in WPM + 1.5 mg−1 2,4-D + 0.4 mg−1 6-BA medium for 30 days; Table S1: Effects of different mediums and 2,4-D concentrations on callus induction for cv. ‘Legacy’; Table S2: Effects of different plant growth regulator combinations on callus induction for leaves of cv. ‘Legacy’; Table S3: Effects of different concentrations of 6-BA on the subculture of callus of cv. ‘Legacy’.
Author Contributions
Conceptualization, L.Z.; methodology, X.Q. and J.H.; software, X.Q. and J.H.; validation, X.Q. and H.S.; formal analysis, X.Q.; investigation, X.Q., H.S. and Y.C.; resources, G.X.; data curation, X.Q. and J.H.; writing—original draft preparation, X.Q.; writing—review and editing, L.Z. and Y.C.; visualization, L.Z. and Y.C.; supervision, L.Z.; project administration, L.Z. and Y.C.; funding acquisition, L.Z. and Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the National Natural Science Foundation of China (grant number 31872942 to L.Z.), Natural Science Foundation of Beijing (grant number 6222041 to L.Z.) and China Postdoctoral Science Foundation (grant number 2021T140060 to Y.C.).
Data Availability Statement
All relevant data can be found within the manuscript and its Supplementary material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Silva, S.; Costa, E.M.; Veiga, M.; Morais, R.M.; Calhau, C.; Pintado, M. Health promoting properties of blueberries: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Herniter, I.A.; Kim, Y.; Wang, Y.; Havill, J.S.; Johnson-Cicalese, J.; Muehlbauer, G.J.; Iorizzo, M.; Vorsa, N. Trait Mapping of Phenolic Acids in an Interspecific (Vaccinium corymbosum var. caesariense× V. darrowii) Diploid Blueberry Population. Plants 2023, 12, 1346. [Google Scholar] [CrossRef] [PubMed]
- Woolf, E.K.; Terwoord, J.D.; Litwin, N.S.; Vazquez, A.R.; Lee, S.Y.; Ghanem, N.; Michell, K.A.; Smith, B.T.; Grabos, L.E.; Ketelhut, N.B.; et al. Daily blueberry consumption for 12 weeks improves endothelial function in postmenopausal women with above-normal blood pressure through reductions in oxidative stress: A randomized controlled trial. Food Funct. 2023, 14, 2621–2641. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-Y.; Chen, H.; Camp, M.-J.; Ehlenfeldt, M.-K. Flavonoid constituents and their contribution to antioxidant activity in cultivars and hybrids of rabbiteye blueberry (Vaccinium ashei Reade). Food Chem. 2012, 132, 855–864. [Google Scholar] [CrossRef]
- Yang, H.; Wu, Y.-Q.; Zhang, C.-H.; Wu, W.L.; Lyu, L.-F.; Li, W.-L. Growth and physiological characteristics of four blueberry cultivars under different high soil pH treatments. Environ. Exp. Bot. 2022, 197, 104842. [Google Scholar] [CrossRef]
- Cabezas, D.; de Bem Oliveira, I.; Acker, M.; Lyrene, P.; Munoz, P.R. Evaluating Wild Germplasm Introgression into Autotetraploid Blueberry. Agronomy 2021, 11, 614. [Google Scholar] [CrossRef]
- Oliveira, I.D.B.; Amadeu, R.R.; Ferrão, L.F.V.; Muñoz, P.R. Optimizing whole-genomic prediction for autotetraploid blueberry breeding. Heredity 2020, 125, 437–448. [Google Scholar] [CrossRef]
- Kulkarni, K.P.; Vorsa, N.; Natarajan, P.; Elavarthi, S.; Iorizzo, M.; Reddy, U.K.; Melmaiee, K. Admixture Analysis Using Genotyping-by-Sequencing Reveals Genetic Relatedness and Parental Lineage Distribution in Highbush Blueberry Genotypes and Cross Derivatives. Int. J. Mol. Sci. 2020, 22, 163. [Google Scholar] [CrossRef]
- Sater, H.M.; Bizzio, L.N.; Tieman, D.M.; Muñoz, P.D. A Review of the Fruit Volatiles Found in Blueberry and Other Vaccinium Species. J. Agric. Food Chem. 2020, 68, 5777–5786. [Google Scholar] [CrossRef]
- Altpeter, F.; Springer, N.M.; E Bartley, L.; Blechl, A.E.; Brutnell, T.P.; Citovsky, V.; Conrad, L.J.; Gelvin, S.B.; Jackson, D.P.; Kausch, A.P.; et al. Advancing Crop Transformation in the Era of Genome Editing. Plant Cell 2016, 28, 1510–1520. [Google Scholar] [CrossRef] [Green Version]
- Aliu, E.; Lee, K.; Wang, K. CRISPR RNA-guided integrase enables high-efficiency targeted genome engineering in Agrobacterium tumefaciens. Plant Biotechnol. J. 2022, 20, 1916–1927. [Google Scholar] [CrossRef]
- Jiang, D.; Zhu, W.; Wang, Y.; Sun, C.; Zhang, K.-Q.; Yang, J. Molecular tools for functional genomics in filamentous fungi: Recent advances and new strategies. Biotechnol. Adv. 2013, 31, 1562–1574. [Google Scholar] [CrossRef]
- Walworth, A.E.; Rowland, L.J.; Polashock, J.J.; Hancock, J.F.; Song, G.-Q. Overexpression of a blueberry-derived CBF gene enhances cold tolerance in a southern highbush blueberry cultivar. Mol. Breed. 2012, 30, 1313–1323. [Google Scholar] [CrossRef]
- Song, G.-Q.; Roggers, R.; Sink, K.; Particka, M.; Zandstra, B. Production of herbicide-resistant highbush blueberry ‘legacy’ by Agrobacterium-mediated transformation of the bar gene. Acta Hortic. 2007, 738, 397–407. [Google Scholar] [CrossRef]
- Li, X.-Y.; Hou, Y.-M.; Xie, X.; Li, H.-X.; Li, X.-D.; Zhu, Y.; Zhai, L.-L.; Zhang, C.Y.; Bian, S.-M. A blueberry MIR156a–SPL12 module coordinates the accumulation of chlorophylls and anthocyanins during fruit ripening. J. Exp. Bot. 2020, 71, 5976–5989. [Google Scholar] [CrossRef]
- Zhang, C.-Y.; Liu, H.-C.; Zhang, X.-S.; Guo, Q.-X.; Bian, S.-M.; Wang, J.-Y.; Zhai, L.-L. VcMYB4a, an R2R3-MYB transcription factor from Vaccinium corymbosum, negatively regulates salt, drought, and temperature stress. Gene 2020, 757, 144935. [Google Scholar] [CrossRef]
- Gao, X.; Walworth, A.E.; Mackie, C.; Song, G.-Q. Overexpression of blueberry Flowering locus T is associated with changes in the expression of phytohormone-related genes in blueberry plants. Hortic. Res. 2016, 3, 16053. [Google Scholar] [CrossRef]
- Zhao, X.; Zhan, L.-P.; Zou, X.-Z. Improvement of cold tolerance of the half-high bush Northland blueberry by transformation with the LEA gene from Tamarix androssowii. Plant Growth Regul. 2010, 63, 13–22. [Google Scholar] [CrossRef]
- Song, G.-Q.; Sink, K.C. Agrobacterium tumefaciens-mediated transformation of blueberry (Vaccinium corymbosum L.). Plant Cell Rep. 2004, 23, 475–484. [Google Scholar] [CrossRef]
- Song, G.-Q. Blueberry (Vaccinium corymbosum L.). Agrobacterium Protocols. In Methods in Molecular Biology; Springer: New York, NY, USA, 2014; Volume 1224, pp. 121–131. [Google Scholar] [CrossRef]
- Wang, G.-D.; Liu, M.-M.; Liu, G.-Z.; Bao, Z.L.; Ma, F.F. Establishment and optimization of agrobacterium-mediated transformation in blueberry (Vaccinium species). Sci. Hortic. 2022, 304, 111258. [Google Scholar] [CrossRef]
- Cao, X.; Liu, Q.; Rowland, L.J.; Hammerschlag, F.A. GUS expression in blueberry (Vaccinium spp.): Factors influencing Agrobacterium -mediated gene transfer efficiency. Plant Cell Rep. 1998, 18, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Gao, E.-L.; Zhao, Y.-J.; Guo, W.-W.; Wang, P.-W. Optimized plasmids for callus transformation and functional studies of autophagy in citrus. Plant Cell Tissue Organ Cult. (PCTOC) 2022, 152, 507–516. [Google Scholar] [CrossRef]
- An, X.-H.; Tian, Y.; Chen, K.-Q.; Wang, X.-F.; Hao, Y.-J. The apple WD40 protein MdTTG1 interacts with bHLH but not MYB proteins to regulate anthocyanin accumulation. J. Plant Physiol. 2012, 169, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.-L.; Zhou, W.; Yan, J.-D.; Gao, Y.-R.; Li, X.-W.; Sun, J.-C.; Fang, K.-F.; Zhang, Q.; Xing, Y.; Qin, L.; et al. Agrobacterium-mediated genetic transformation of Chinese chestnut (Castanea mollissima Blume). Plant Cell Tissue Organ Cult. (PCTOC) 2019, 140, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-H.; Zhou, F.-L.; Liu, J.-L.; Liu, W.-Q.; Zhang, S.-L.; Li, D.-L.; Song, J.-K.; Wang, R.; Yang, Y.-J. Establishment of efficient callus genetic transformation system for Pyrus armeniacaefolia. Sci. Hortic. 2021, 289, 110429. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Wang, P.-F.; Li, R.; Hyden, B.; An, X.-Y.; Jing, R.-Y.; Zhao, X.-L.; Zhang, Y.; Qiao, H.-Y.; Han, Y.; et al. Cellular and metabolic characteristics of peach anther-derived callus. Sci. Hortic. 2023, 311, 111796. [Google Scholar] [CrossRef]
- Saika, H.; Toki, S. Mature seed-derived callus of the model indica rice variety Kasalath is highly competent in Agrobacterium-mediated transformation. Plant Cell Rep. 2010, 29, 1351–1364. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-F.; Karioti, A.; Rohr, D.; Bilia, A.R.; Efferth, T. Production of rosmarinic acid and salvianolic acid B from callus culture of Salvia miltiorrhiza with cytotoxicity towards acute lymphoblastic leukemia cells. Food Chem. 2016, 201, 292–297. [Google Scholar] [CrossRef]
- Rakshana, P.; Valarmathi, R.; Raveendran, M. Optimization of tissue culture protocol for rapid regeneration of traditional therapeutic rice genotype ‘Kavuni’. Electron. J. Plant Breed. 2019, 10, 334. [Google Scholar] [CrossRef]
- Long, Y.; Yang, Y.; Pan, G.-T.; Shen, Y.-U. New Insights Into Tissue Culture Plant-Regeneration Mechanisms. Front. Plant Sci. 2022, 13, 926752. [Google Scholar] [CrossRef]
- Zeng, S.-J.; Wang, J.; Wu, K.-L.; da Silva, J.A.T.; Zhang, J.-X.; Duan, J. In vitro propagation of Paphiopedilum hangianum Perner & Gruss. Sci. Hortic. 2013, 151, 147–156. [Google Scholar] [CrossRef]
- Zhu, X.-T.; Li, X.-Q.; Ding, W.-J.; Jin, S.-H.; Wang, Y. Callus induction and plant regeneration from leaves of peony. Hortic. Environ. Biotechnol. 2018, 59, 575–582. [Google Scholar] [CrossRef]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Jiang, M.-M.; Ren, L.; Lian, H.-L.; Liu, Y.; Chen, H.-Y. Novel insight into the mechanism underlying light-controlled anthocyanin accumulation in eggplant (Solanum melongena L.). Plant Sci. 2016, 249, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Richter, A.S.; Nägele, T.; Grimm, B.; Kaufmann, K.; Schroda, M.; Leister, D.; Kleine, T. Retrograde signaling in plants: A critical review focusing on the GUN pathway and beyond. Plant Commun. 2023, 4, 100511. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Zheng, X.-T.; Sun, B.-Y.; Peng, C.-L.; Chow, W.S. Over-expression of the CHS gene enhances resistance of Arabidopsis leaves to high light. Environ. Exp. Bot. 2018, 154, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.-Z.; Xu, C.-Y.; Xu, K.; Hu, Y.-X. Lateral organ boundaries domain transcription factors direct callus formation in Arabidopsis regeneration. Cell Res. 2012, 22, 1169–1180. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Cao, H.-F.; Zhang, Q.-Q.; Wang, H.-Z.; Xin, W.; Xu, E.-J.; Zhang, S.-Q.; Yu, R.-X.; Yu, D.-X.; Hu, Y.-X. Control of auxin-induced callus formation by bZIP59–LBD complex in Arabidopsis regeneration. Nat. Plants 2018, 4, 108–115. [Google Scholar] [CrossRef]
- Hoshino, Y.; Miyashita, T.; Thomas, T.D. In vitro culture of endosperm and its application in plant breeding: Approaches to polyploidy breeding. Sci. Hortic. 2011, 130, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Farhadi, N.; Panahandeh, J.; Azar, A.M.; Salte, S.A. Effects of explant type, growth regulators and light intensity on callus induction and plant regeneration in four ecotypes of Persian shallot (Allium hirtifolium). Sci. Hortic. 2017, 218, 80–86. [Google Scholar] [CrossRef]
- Xia, Y.-F.; Cao, Y.; Ren, Y.-Y.; Ling, A.-Y.; Du, K.; Li, Y.; Yang, J.; Kang, X.-Y. Effect of a suitable treatment period on the genetic transformation efficiency of the plant leaf disc method. Plant Methods 2023, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Iwase, A.; Harashima, H.; Ikeuchi, M.; Rymen, B.; Ohnuma, M.; Komaki, S.; Morohashi, K.; Kurata, T.; Nakata, M.; Ohme-Takagi, M.; et al. WIND1 Promotes Shoot Regeneration through Transcriptional Activation of ENHANCER OF SHOOT REGENERATION1 in Arabidopsis. Plant Cell 2016, 29, 54–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.-X.; Zhang, Z.-G.; Zhang, Q.; Lang, Z.-H.; Sun, X.-H. Scarabaeid Larvae- and Herbicide-Resistant Transgenic Perennial Ryegrass (Lolium perenne L.) Obtained by Agrobacterium tumefaciens-Mediated Transformation of cry8Ca2, cry8Ga and bar Genes. J. Integr. Agric. 2012, 11, 53–61. [Google Scholar] [CrossRef]
- Hiei, Y.; Komari, T. Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed. Nat. Protoc. 2008, 3, 824–834. [Google Scholar] [CrossRef]
- Tie, W.-W.; Zhou, F.; Wang, L.; Xie, W.-B.; Chen, H.; Li, X.-H.; Lin, Y.-J. Reasons for lower transformation efficiency in indica rice using Agrobacterium tumefaciens-mediated transformation: Lessons from transformation assays and genome-wide expression profiling. Plant Mol. Biol. 2011, 78, 1–18. [Google Scholar] [CrossRef]
- Zhao, Y.-Y.; Yang, D.-X.; Liu, Y.-M.; Han, F.-Q.; Li, Z.-S. A highly efficient genetic transformation system for broccoli and subcellular localization. Front. Plant Sci. 2023, 14, 1091588. [Google Scholar] [CrossRef]
- Hiei, Y.; Ishida, Y.; Komari, T. Progress of cereal transformation technology mediated by Agrobacterium tumefaciens. Front. Plant Sci. 2014, 5, 628. [Google Scholar] [CrossRef] [Green Version]
- Dai, H.-Y.; Li, W.-R.; Han, G.-F.; Yang, Y.; Ma, Y.; Li, H.; Zhang, Z.-H. Development of a seedling clone with high regeneration capacity and susceptibility to Agrobacterium in apple. Sci. Hortic. 2013, 164, 202–208. [Google Scholar] [CrossRef]
- Wang, X.-P.; Chen, S.-S.; Zhang, H.-N.; Luo, P.; Zhou, F.-P.; Zeng, B.-S.; Xu, J.-M.; Fan, C.-J. Agrobacterium-mediated genetic transformation of the most widely cultivated superior clone Eucalyptus urophylla × Eucalyptus grandis DH32-29 in Southern China. Front. Plant Sci. 2022, 13, 1011245. [Google Scholar] [CrossRef]
- Shan, Q.-W.; Zhang, Y.; Chen, K.-L.; Zhang, K.; Gao, C.-X. Creation of fragrant rice by targeted knockout of the OsBADH2 gene using TALEN technology. Plant Biotechnol. J. 2015, 13, 791–800. [Google Scholar] [CrossRef]
- Zhang, J.-X.; Wu, J.-P.; Hao, X.-Y.; Xie, Y.-P.; Lv, K.; Xu, W.-R. Establishment of a stable grape immature zygotic embryo-based genetic transformation system. Sci. Hortic. 2023, 316, 112009. [Google Scholar] [CrossRef]
- Sutradhar, M.; Mandal, N. Reasons and riddance of Agrobacterium tumefaciens overgrowth in plant transformation. Transgenic Res. 2023, 32, 33–52. [Google Scholar] [CrossRef]
- Ribas, A.F.; Dechamp, E.; Champion, A.; Bertrand, B.; Combes, M.-C.; Verdeil, J.-L.; Lapeyre, F.; Lashermes, P.; Etienne, H. Agrobacterium-mediated genetic transformation of Coffea arabica (L.) is greatly enhanced by using established embryogenic callus cultures. BMC Plant Biol. 2011, 11, 92. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-F.; An, J.-P.; Liu, X.; Su, L.; You, C.-X.; Hao, Y.-J. The Nitrate-Responsive Protein MdBT2 Regulates Anthocyanin Biosynthesis by Interacting with the MdMYB1 Transcription Factor. Plant Physiol. 2018, 178, 890–906. [Google Scholar] [CrossRef] [Green Version]
- An, J.-P.; Zhang, X.-W.; You, C.-X.; Bi, S.-Q.; Wang, X.-F.; Hao, Y.-J. MdWRKY40 promotes wounding-induced anthocyanin biosynthesis in association with MdMYB1 and undergoes MdBT2-mediated degradation. New Phytol. 2019, 224, 380–395. [Google Scholar] [CrossRef]
- Dong, Q.-L.; Zheng, W.-Q.; Duan, D.-Y.; Huang, D.; Wang, Q.; Liu, C.-H.; Li, C.; Gong, X.-Q.; Li, C.-Y.; Mao, K.; et al. MdWRKY30, a group IIa WRKY gene from apple, confers tolerance to salinity and osmotic stresses in transgenic apple callus and Arabidopsis seedlings. Plant Sci. 2020, 299, 110611. [Google Scholar] [CrossRef]
- Gao, M.; Zhao, H.-Y.; Zheng, L.-T.; Zhang, L.-H.; Peng, Y.-J.; Ma, W.-F.; Tian, R.; Yuan, Y.-Y.; Ma, F.-W.; Li, M.-J.; et al. Overexpression of apple Ma12, a mitochondrial pyrophosphatase pump gene, leads to malic acid accumulation and the upregulation of malate dehydrogenase in tomato and apple calli. Hortic. Res. 2022, 9, uhab053. [Google Scholar] [CrossRef]
- An, J.-P.; Qu, F.-J.; Yao, J.-F.; Wang, X.-N.; You, C.-X.; Wang, X.F.; Hao, Y.-J. The bZIP transcription factor MdHY5 regulates anthocyanin accumulation and nitrate assimilation in apple. Hortic. Res. 2017, 4, 17023. [Google Scholar] [CrossRef] [Green Version]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. VaCPK20, a calcium-dependent protein kinase gene of wild grapevine Vitis amurensis Rupr., mediates cold and drought stress tolerance. J. Plant Physiol. 2015, 185, 1–12. [Google Scholar] [CrossRef]
- Ananev, A.A.; Suprun, A.R.; Aleynova, O.A.; Nityagovsky, N.N.; Ogneva, Z.V.; Dubrovina, A.S.; Kiselev, K.V. Effect of VaMyb40 and VaMyb60 Overexpression on Stilbene Biosynthesis in Cell Cultures of Grapevine Vitis amurensis Rupr. Plants 2022, 11, 1916. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Shi, Y.; Wang, J.; Duan, C.-Q.; Yu, K.-J. Transcription factor VvibHLH93 negatively regulates proanthocyanidin biosynthesis in grapevine. Front. Plant Sci. 2022, 13, 1007895. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T. Biotechnology Applications of Plant Callus Cultures. Engineering 2019, 5, 50–59. [Google Scholar] [CrossRef]
- Martínez-Márquez, A.; Morante-Carriel, J.A.; Palazon, J.; Bru-Martínez, R. Rosa hybrida orcinol O-methyl transferase-mediated production of pterostilbene in metabolically engineered grapevine cell cultures. New Biotechnol. 2018, 42, 62–70. [Google Scholar] [CrossRef]
- Rivera, S.M.; Christou, P.; Canela-Garayoa, R. Identification of carotenoids using mass spectrometry. Mass Spectrom. Rev. 2014, 33, 353–372. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Leaf growth status of cv. ‘Legacy’ in callus induction medium containing different combinations of 2,4-D and 6-BA. (A–D) Leaf growth status in WPM containing 1 mg·L−1 2,4-D and different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1), respectively. (E–H) Leaf growth status in WPM containing 4 mg·L−1 2,4-D and different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1), respectively. The scale in the figure is 1 cm.
Figure 1.
Leaf growth status of cv. ‘Legacy’ in callus induction medium containing different combinations of 2,4-D and 6-BA. (A–D) Leaf growth status in WPM containing 1 mg·L−1 2,4-D and different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1), respectively. (E–H) Leaf growth status in WPM containing 4 mg·L−1 2,4-D and different concentrations of 6-BA (0.2, 0.4, 0.6, 0.8 mg·L−1), respectively. The scale in the figure is 1 cm.

Figure 2.
The callus growth process of cv. ‘Northland’. (A) Leaves of cv. ‘Northland’ were cultured in WPM + 1 mg−1 2,4-D + 0.4 mg−1 6-BA medium for 40 d. (B) The callus of cv. ‘Northland’ was removed from the leaves and cultured in WPM + 1.5 mg−1 2,4-D + 0.4 mg−1 6-BA for 20–30 d to produce yellowish spherical callus and grow rapidly. (C) The callus of cv. ‘Northland’ was cultivated in WPM + 1.5 mg−1 2,4-D + 0.4 mg−1 6-BA medium for 30 days. The scale in the figure is 1 cm.
Figure 2.
The callus growth process of cv. ‘Northland’. (A) Leaves of cv. ‘Northland’ were cultured in WPM + 1 mg−1 2,4-D + 0.4 mg−1 6-BA medium for 40 d. (B) The callus of cv. ‘Northland’ was removed from the leaves and cultured in WPM + 1.5 mg−1 2,4-D + 0.4 mg−1 6-BA for 20–30 d to produce yellowish spherical callus and grow rapidly. (C) The callus of cv. ‘Northland’ was cultivated in WPM + 1.5 mg−1 2,4-D + 0.4 mg−1 6-BA medium for 30 days. The scale in the figure is 1 cm.

Figure 3.
Callus of cv. ‘Northland’ growth status in different concentrations of cefotaxime and hygromycin. (A) CK. (B–E) Callus growth status in different concentrations of Cef (100, 200, 300, 400 mg·L−1). (F) CK. (G–I) Callus growth status in different concentrations of Hyg (30, 50, 80 mg·L−1). The scale in (A–D) is the same as in (E) and the scale in (F–H) is the same as in (I).
Figure 3.
Callus of cv. ‘Northland’ growth status in different concentrations of cefotaxime and hygromycin. (A) CK. (B–E) Callus growth status in different concentrations of Cef (100, 200, 300, 400 mg·L−1). (F) CK. (G–I) Callus growth status in different concentrations of Hyg (30, 50, 80 mg·L−1). The scale in (A–D) is the same as in (E) and the scale in (F–H) is the same as in (I).

Figure 4.
Identification of the GFP signal and PCR detection. (A) Green fluorescence was observed in the transformed callus. (B) Green fluorescence was not observed in the wild-type callus. (C) PCR detection of the VcCHS, GFP, and Hpt II gene, non-transgenic sample (WT), and 3 transgenic lines (L1–L3).
Figure 4.
Identification of the GFP signal and PCR detection. (A) Green fluorescence was observed in the transformed callus. (B) Green fluorescence was not observed in the wild-type callus. (C) PCR detection of the VcCHS, GFP, and Hpt II gene, non-transgenic sample (WT), and 3 transgenic lines (L1–L3).

Figure 5.
Phenotype analysis of the wild-type callus and VcCHS transgenic callus and RT-qPCR detection. (A) Red callus was observed in the transformed callus, while the WT callus showed only a small amount of dot coloring. (B) Analysis of VcCHS transcriptional levels in WT callus and transgenic lines by RT-PCR analysis. (C) Relative expression level of VcCHS gene in WT and transgenic callus (VcCHS−OE). ** indicated extremely significant difference. (D) Anthocyanin content of WT and VcCHS−OE. (E) Analysis of anthocyanin content of WT and VcCHS−OE. ** indicated extremely significant difference.
Figure 5.
Phenotype analysis of the wild-type callus and VcCHS transgenic callus and RT-qPCR detection. (A) Red callus was observed in the transformed callus, while the WT callus showed only a small amount of dot coloring. (B) Analysis of VcCHS transcriptional levels in WT callus and transgenic lines by RT-PCR analysis. (C) Relative expression level of VcCHS gene in WT and transgenic callus (VcCHS−OE). ** indicated extremely significant difference. (D) Anthocyanin content of WT and VcCHS−OE. (E) Analysis of anthocyanin content of WT and VcCHS−OE. ** indicated extremely significant difference.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Orthogonal experimental results of L9(33) for the transformation of cv. ‘Northland’ callus in different conditions.
Table 1.
Orthogonal experimental results of L9(33) for the transformation of cv. ‘Northland’ callus in different conditions.
| Influence Factors | Rate of Resistance (%) | Rate of GFP Signal (%) | ||
|---|---|---|---|---|
| OD600 | Infection Time (min) | Co-Cultivation Time (day) | ||
| 0.4 | 10 | 2 | 34.29 | 25.71 |
| 0.4 | 20 | 4 | 11.11 | 11.11 |
| 0.4 | 30 | 3 | 10.20 | 8.16 |
| 0.7 | 10 | 4 | 49.02 | 49.02 |
| 0.7 | 20 | 3 | 23.08 | 23.08 |
| 0.7 | 30 | 2 | 6.52 | 6.52 |
| 1.0 | 10 | 3 | 27.27 | 27.27 |
| 1.0 | 20 | 2 | 7.55 | 3.77 |
| 1.0 | 30 | 4 | 6.78 | 6.78 |
Table 2.
Variance analysis of factors affecting the efficiency of genetic transformation in cv. ‘Northland’ callus.
Table 2.
Variance analysis of factors affecting the efficiency of genetic transformation in cv. ‘Northland’ callus.
| Source of Variation | Quadratic Sum | Degree of Freedom | Mean Square | F | p |
|---|---|---|---|---|---|
| Error correction model | 1693.367 a | 6 | 282.228 | 12.688 | 0.075 |
| Intercept | 2895.157 | 1 | 2895.157 | 130.153 | 0.008 |
| A: OD600 | 316.395 | 2 | 158.198 | 7.112 | 0.123 |
| B: Infection time | 1206.674 | 2 | 603.337 | 27.123 | 0.036 * |
| C: Co-cultivation time | 170.299 | 2 | 85.149 | 3.828 | 0.207 |
| Error | 44.488 | 2 | 22.244 | ||
| Summation | 4633.013 | 8 |
Note: * indicates a significant difference (p < 0.05). a represented R square =0.974 (adjusted R square =0.898).
Table 3.
ANOVA analysis of infection time influence on transformation frequency for cv. ‘Northland’ callus.
Table 3.
ANOVA analysis of infection time influence on transformation frequency for cv. ‘Northland’ callus.
| Infection Time (min) | F | p | |||
|---|---|---|---|---|---|
| 10 | 20 | 30 | |||
| Transformation efficiency (%) | 34 ± 13.03 | 12.65 ± 9.75 | 7.15 ± 0.88 | 6.815 | 0.029 * |
Note: * indicates a significant difference (p < 0.05). The data was the mean value ± SD (n = 3).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qin, X.; Hu, J.; Xu, G.; Song, H.; Zhang, L.; Cao, Y. An Efficient Transformation System for Fast Production of VcCHS Transgenic Blueberry Callus and Its Expressional Analysis. Plants 2023, 12, 2905. https://doi.org/10.3390/plants12162905
AMA Style
Qin X, Hu J, Xu G, Song H, Zhang L, Cao Y. An Efficient Transformation System for Fast Production of VcCHS Transgenic Blueberry Callus and Its Expressional Analysis. Plants. 2023; 12(16):2905. https://doi.org/10.3390/plants12162905
Chicago/Turabian StyleQin, Xuejing, Jing Hu, Guohui Xu, Huifang Song, Lingyun Zhang, and Yibo Cao. 2023. "An Efficient Transformation System for Fast Production of VcCHS Transgenic Blueberry Callus and Its Expressional Analysis" Plants 12, no. 16: 2905. https://doi.org/10.3390/plants12162905
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.