
Diurnal Regulation of Leaf Photosynthesis Is Related to Leaf-Age-Dependent Changes in Assimilate Accumulation in Camellia oleifera

Abstract
:1. Introduction
2. Results
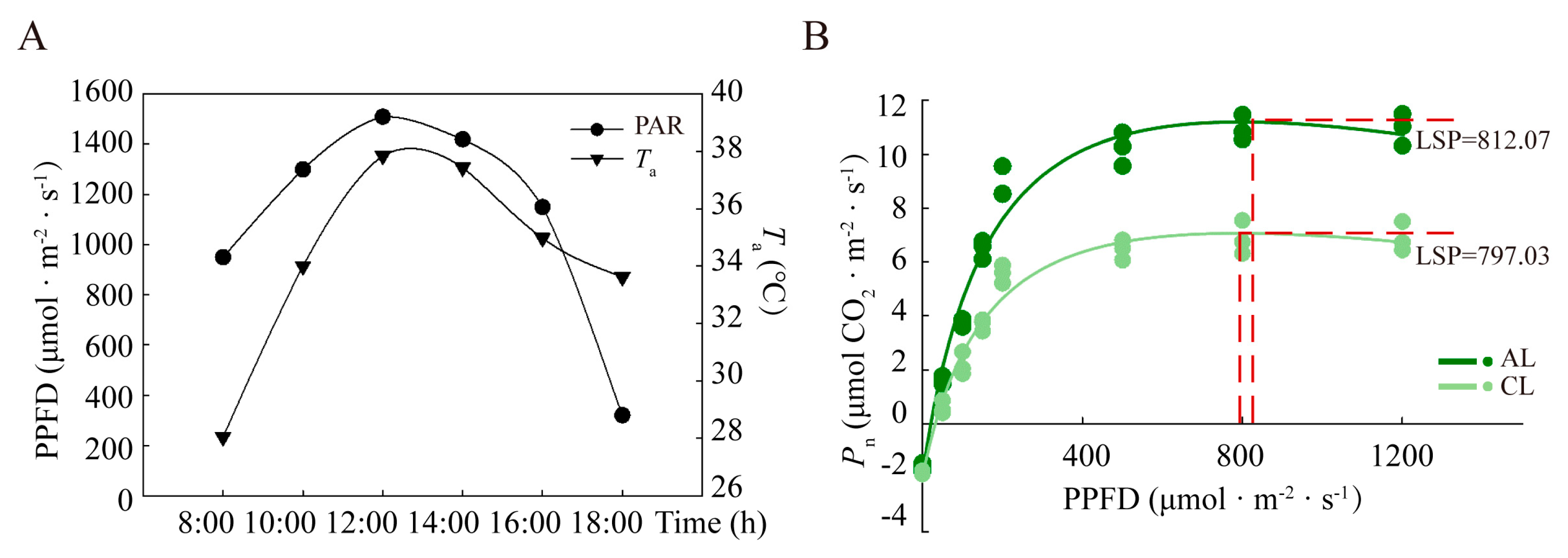
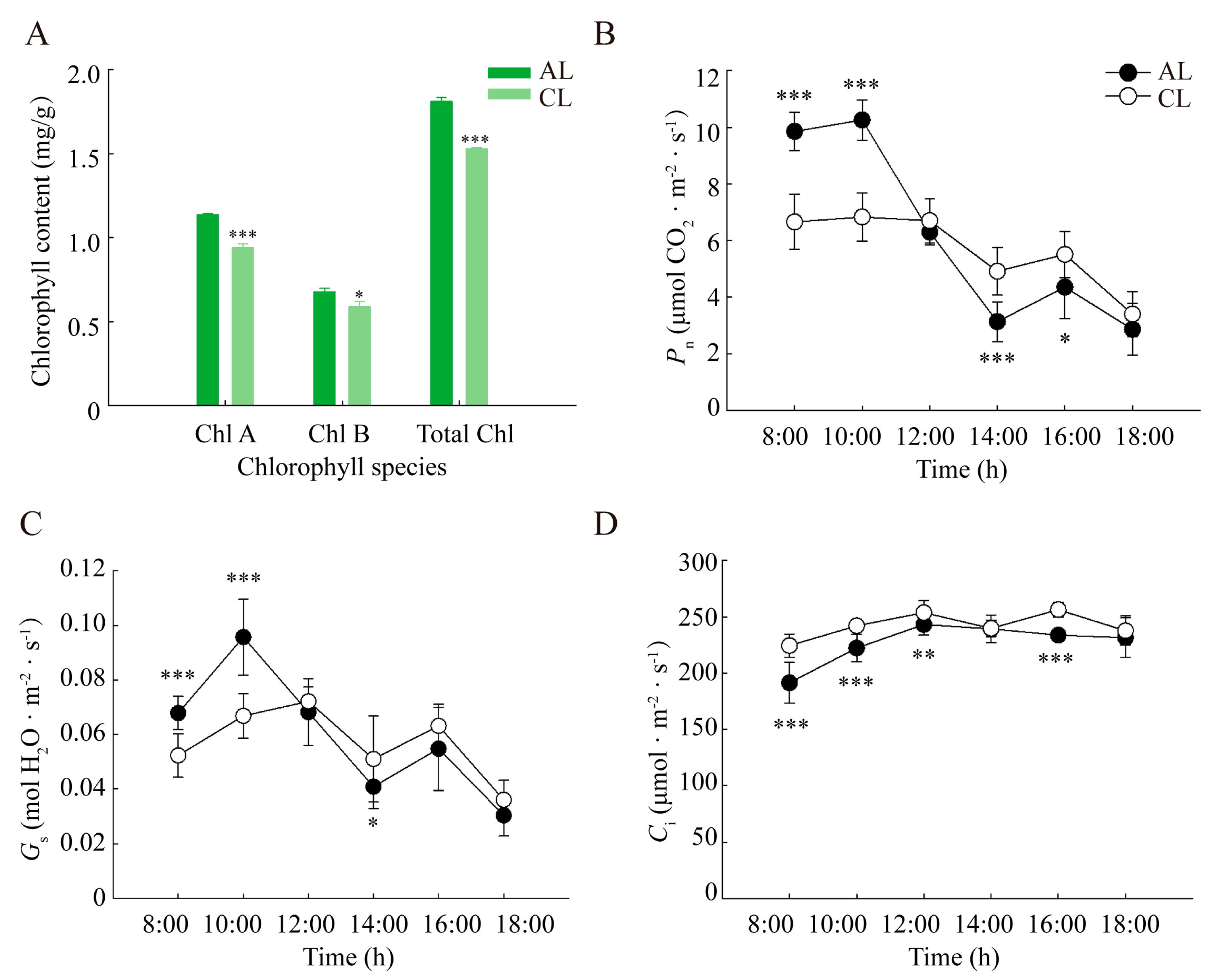
2.1. Diurnal Variation in the Gas Exchange of Leaves of Different Leaf Ages in Camellia oleifera
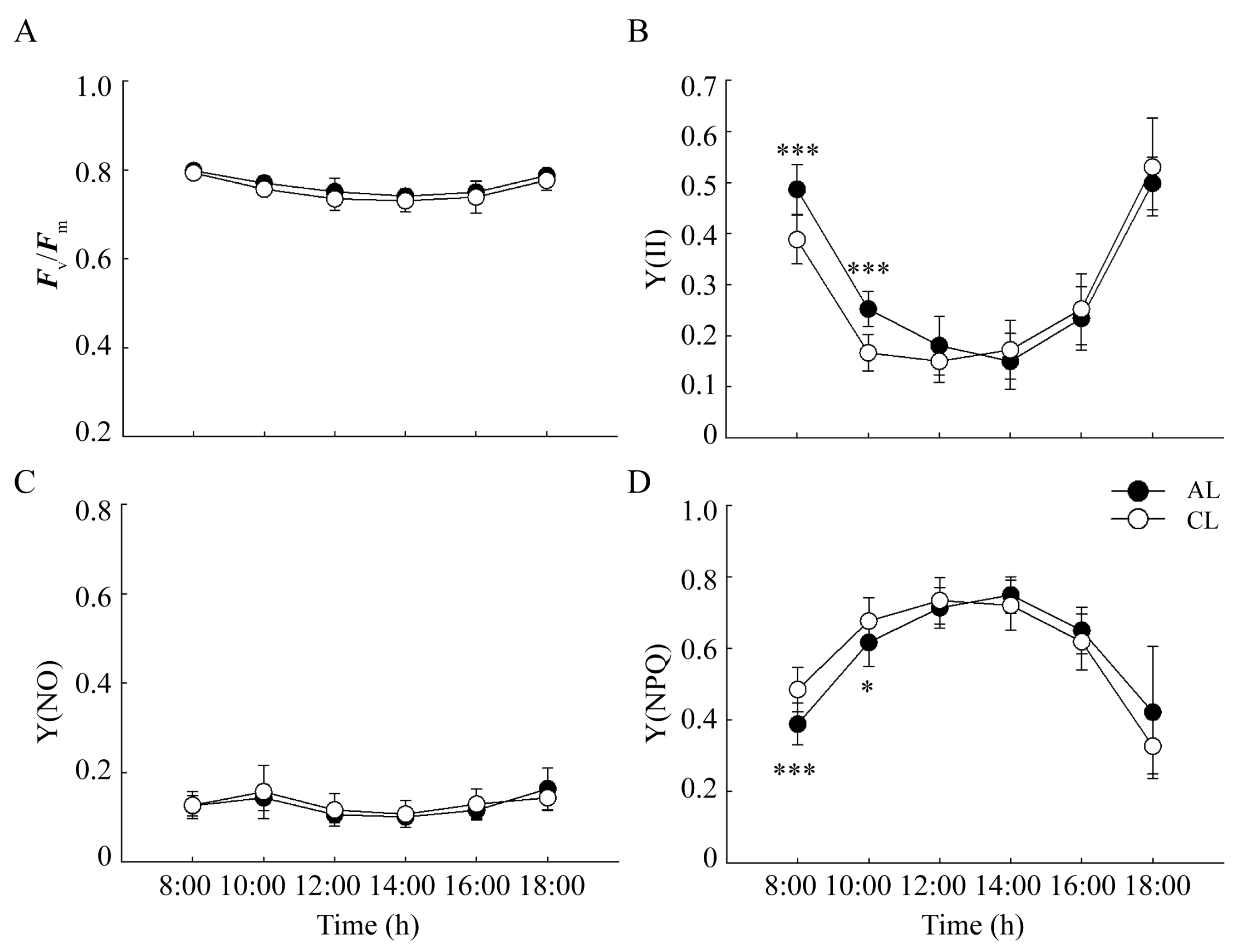
2.2. Changes in the Chlorophyll Fluorescence Parameters of Leaves of Different Leaf Ages in Camellia oleifera
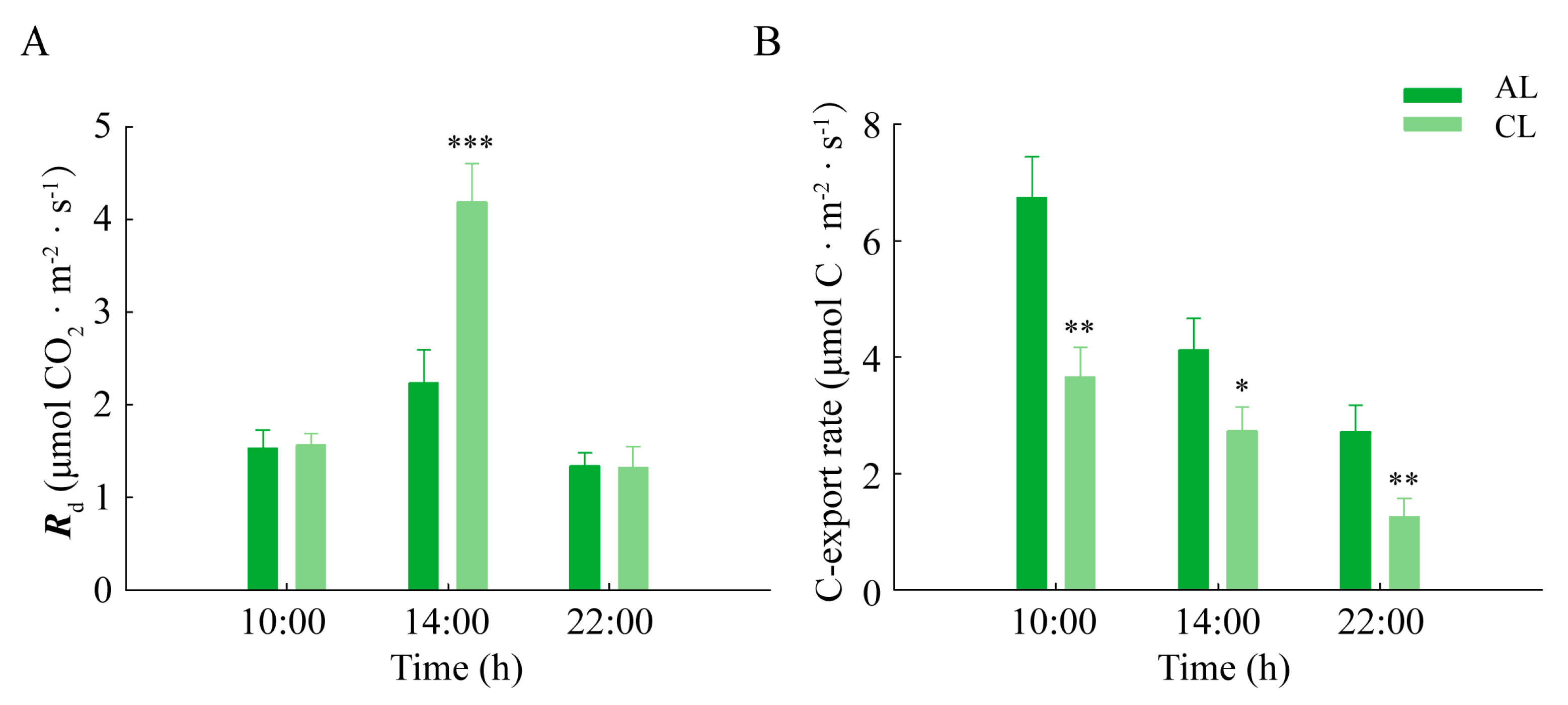
2.3. Dark Respiration and the Carbon Export Rate of Leaves of Different Leaf Ages in Camellia oleifera
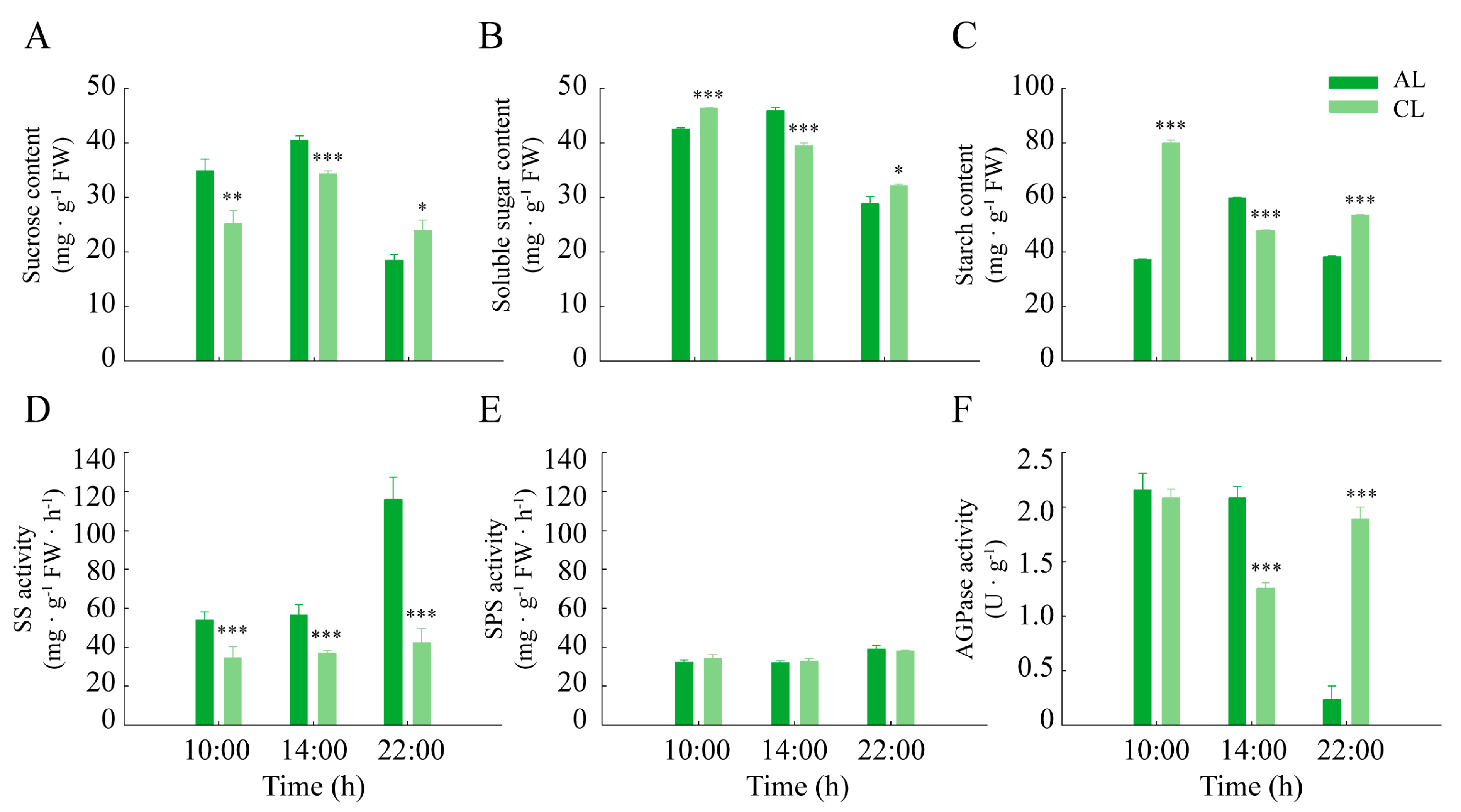
2.4. Assimilate Metabolism Differences of Leaves of Different Leaf Ages in Camellia oleifera
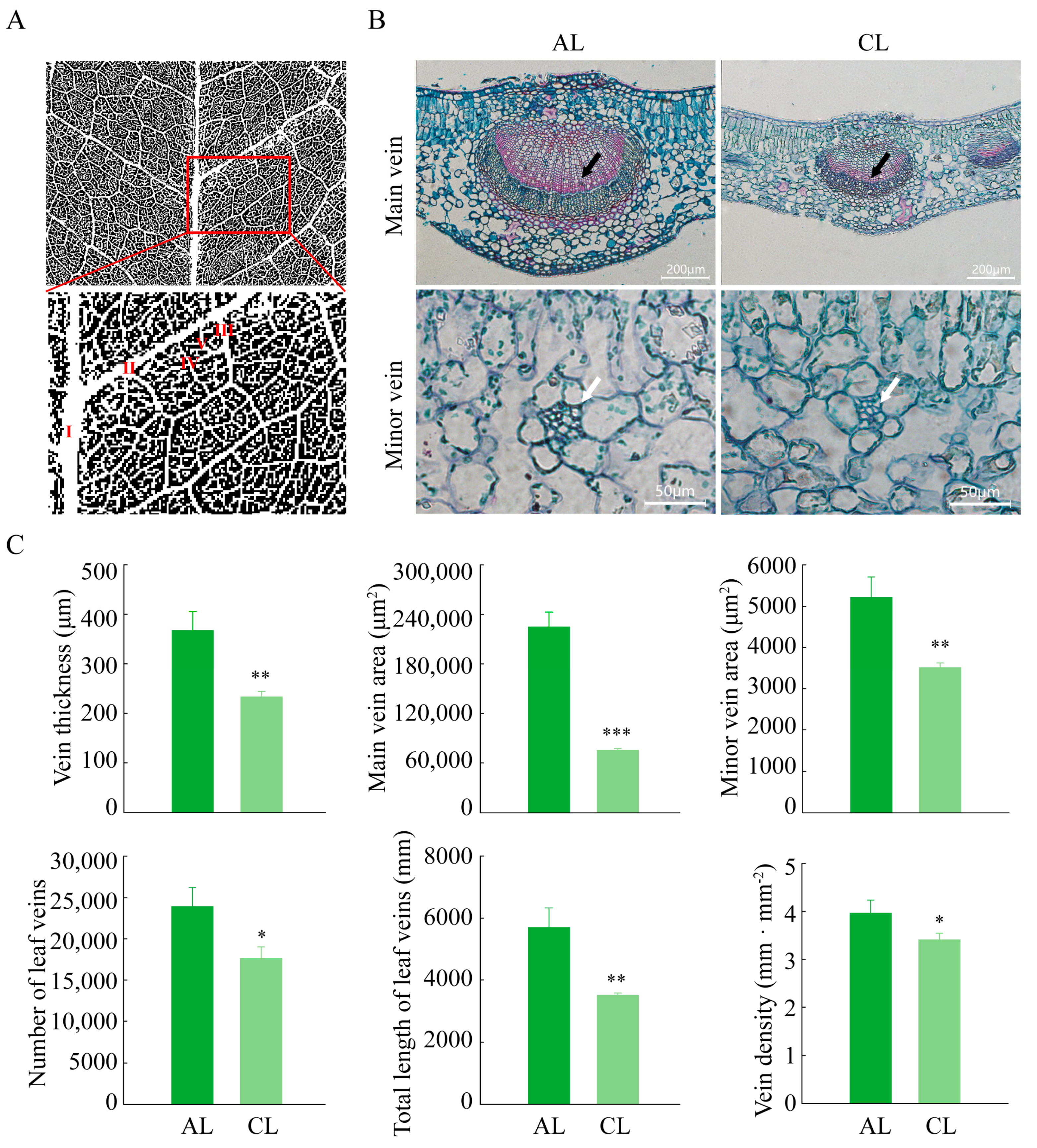
2.5. Structural Differences of Leaves of Different Leaf Ages in Camellia oleifera
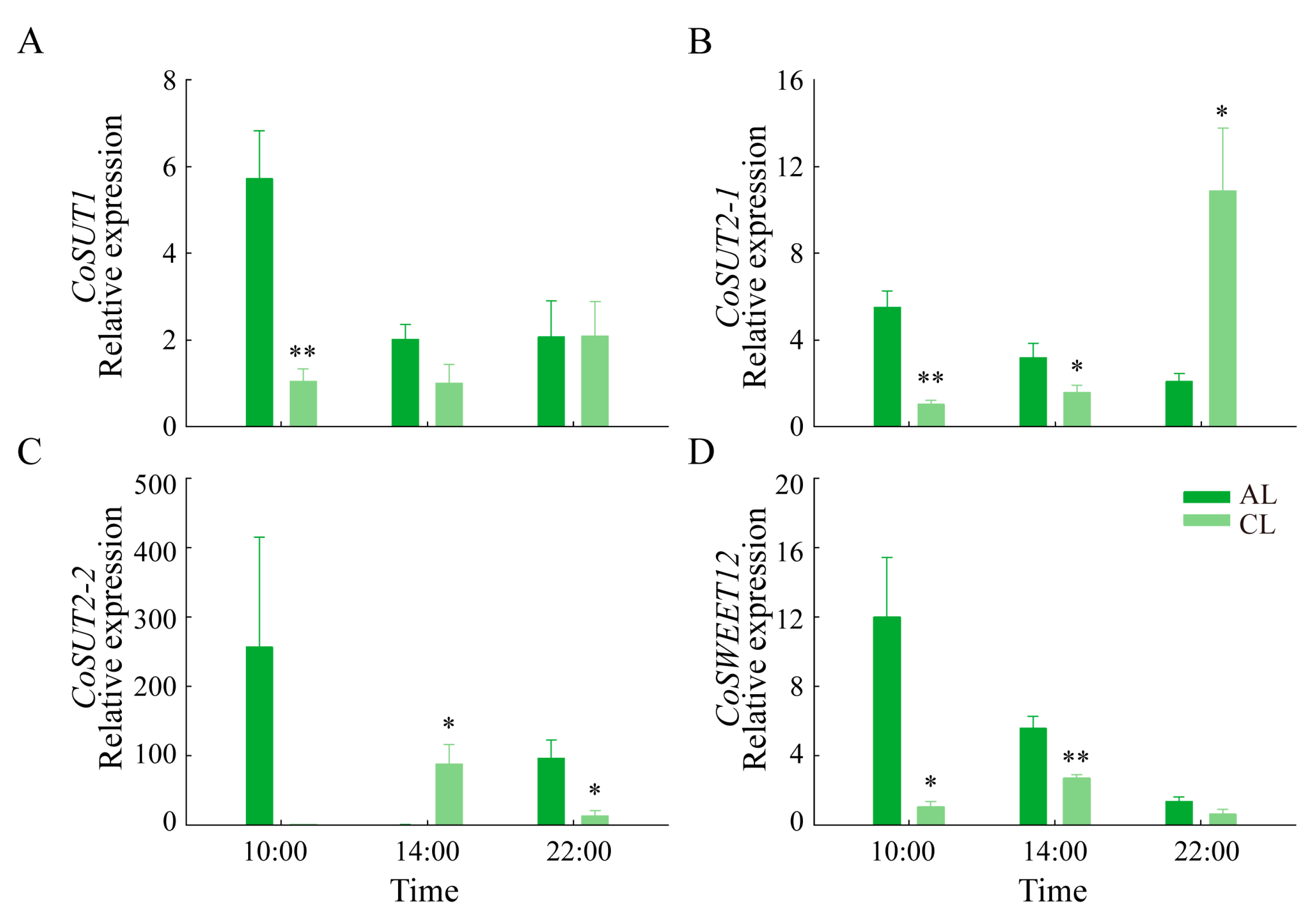
2.6. Gene Expression Differences in Leaves of Different Leaf Ages in Camellia oleifera
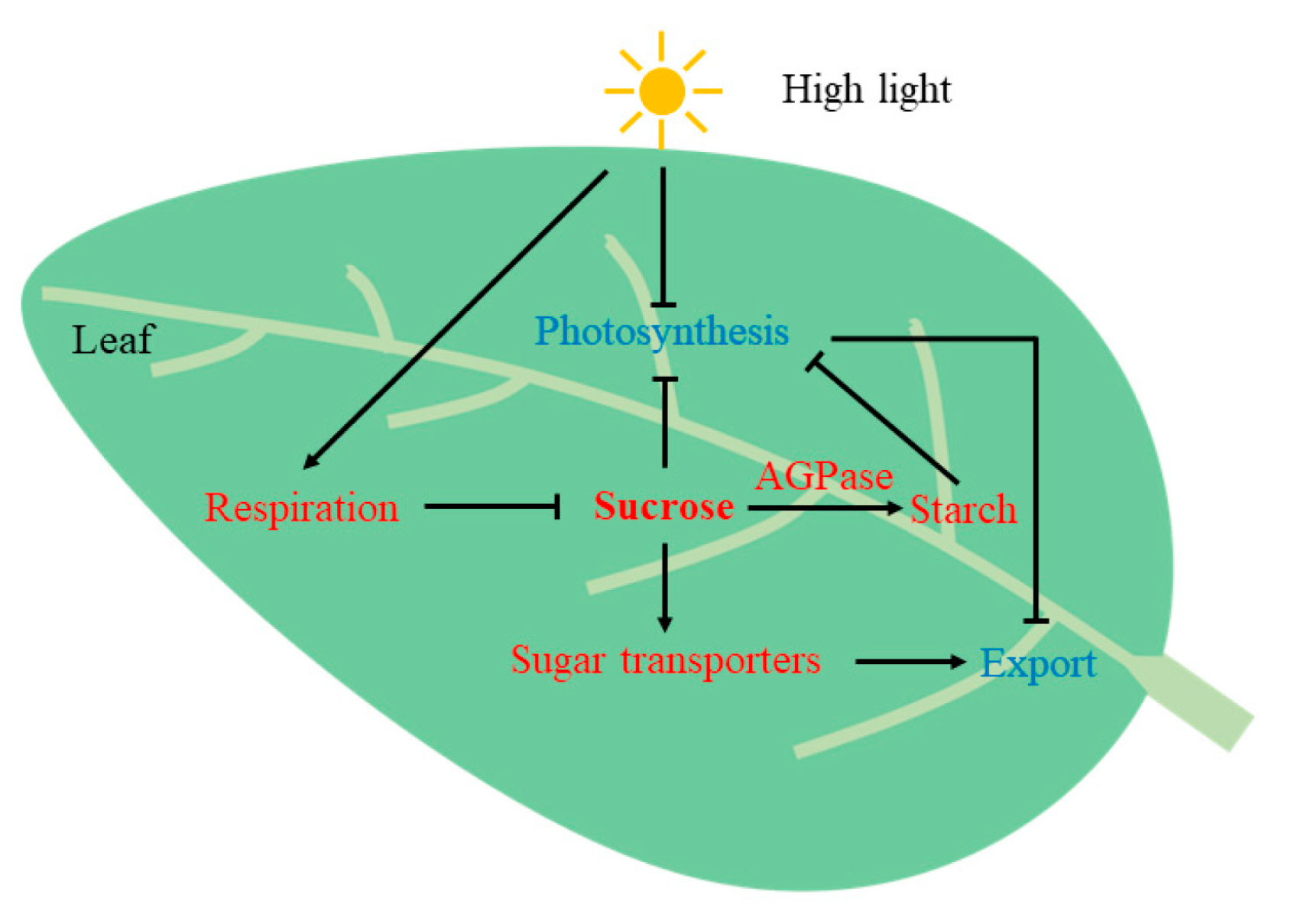
3. Discussion
3.1. Analysis of the Differences in the Photosynthetic ‘Lunch Break’ of Leaves of Different Leaf Ages
3.2. Regulation of the Assimilate Transport in Leaves of Different Leaf Ages
4. Materials and Methods
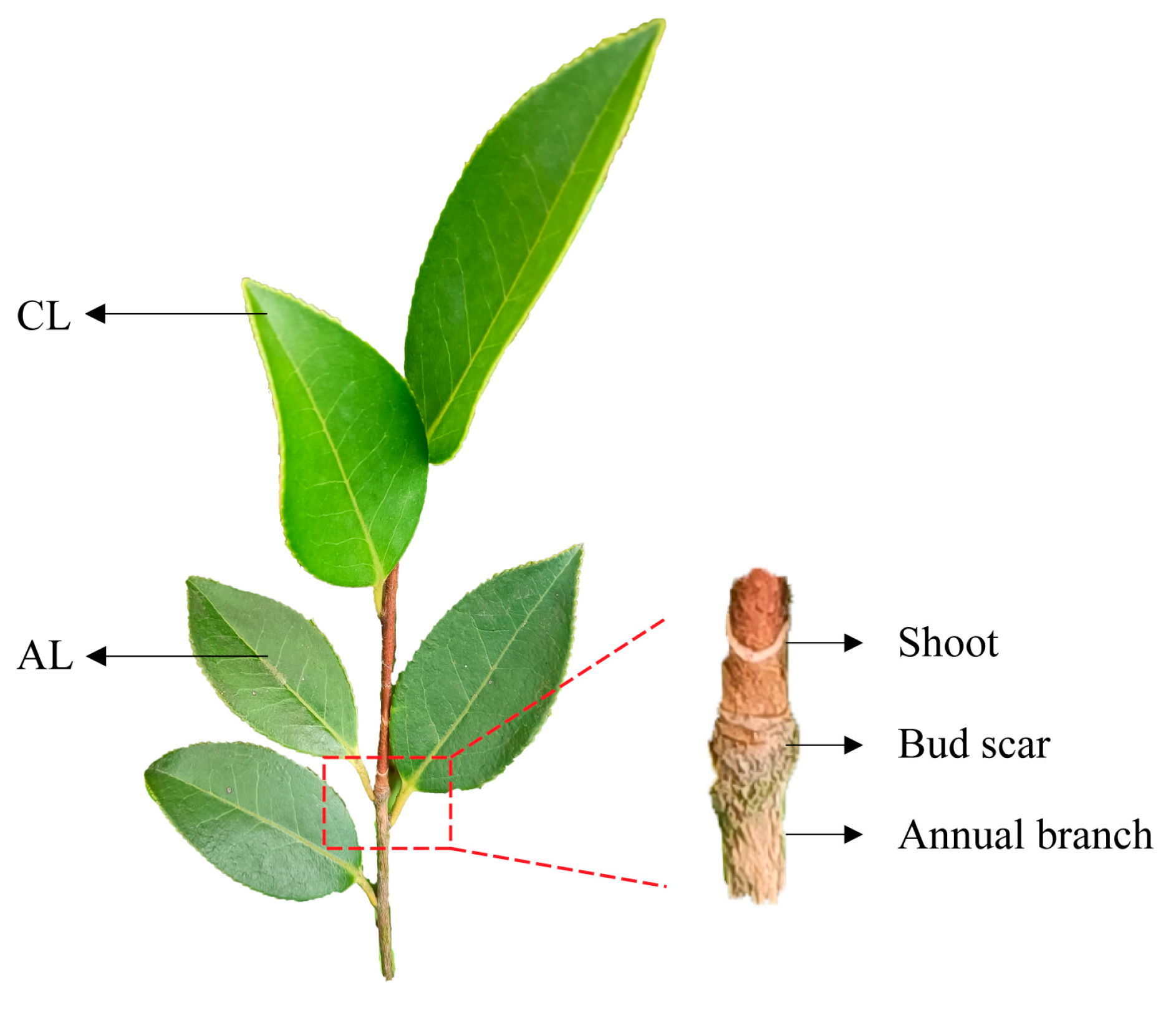
4.1. Plant Materials
4.2. Determination of Chlorophyll Content
4.3. Determination of Diurnal Gas Exchange Parameters and the Carbon Export Rate
4.4. Determination of Diurnal Chlorophyll Fluorescence Parameters
4.5. Determination of Soluble Sugar and Starch Content and Related Metabolic Enzyme Activity
4.6. Extraction of Leaf Venation Information and Observation of Leaf Structure
4.7. Analysis of Gene Expression Levels
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ainsworth, E.A.; Bush, D.R. Carbohydrate Export from the Leaf: A Highly Regulated Process and Target to Enhance Photosynthesis and Productivity. Plant Physiol. 2011, 155, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Tenhunen, J.D.; Pearcy, R.W.; Lange, O.L. Diurnal variations in leaf conductance and gas exchange in natural environments. In Stomatal Function; Zeiger, E., Farquhar, G.D., Cowan, I.R., Eds.; Stanford University Press: Stanford, CA, USA, 1987; pp. 323–351. [Google Scholar]
- Farquhar, G.D.; Sharkey, T.D. Stomatal Conductance and Photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Sage, R.F. Variation in the kcat of Rubisco in C3 and C4 plants and some implications for photosynthetic performance at high and low temperature. J. Exp. Bot. 2002, 53, 609–620. [Google Scholar] [CrossRef]
- Wullschleger, S.D. Biochemical Limitations to Carbon Assimilation in C3 Plants—A Retrospective Analysis of the A/Ci Curves from 109 Species. J. Exp. Bot. 1993, 44, 907–920. [Google Scholar] [CrossRef]
- Hu, M.J.; Guo, Y.P.; Shen, Y.G.; Guo, D.P.; Li, D.Y. Midday depression of photosynthesis and effects of mist spray in citrus. Ann. Appl. Biol. 2009, 154, 143–155. [Google Scholar] [CrossRef]
- Krause, G.H.; Winter, K.; Matsubara, S.; Krause, B.; Jahns, P.; Virgo, A.; Aranda, J.; Garcia, M. Photosynthesis, photoprotection, and growth of shade-tolerant tropical tree seedlings under full sunlight. Photosynth. Res. 2012, 113, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Maai, E.; Nishimura, K.; Takisawa, R.; Nakazaki, T. Light stress-induced chloroplast movement and midday depression of photosynthesis in sorghum leaves. Plant Prod. Sci. 2020, 23, 172–181. [Google Scholar] [CrossRef]
- Henry, C.; Watson-Lazowski, A.; Oszvald, M.; Griffiths, C.A.; Paul, M.; Furbank, R.T.; Ghannoum, O. Sugar sensing responses to low and high light in leaves of the C4 model grass Setaria viridis. J. Exp. Bot. 2020, 71, 1039–1052. [Google Scholar] [CrossRef]
- Huang, X.; Qin, B.; Xia, S.; Su, Y.; Ku, W.; Chen, R.; Peng, K. A comparative study on the effects of strong light stress on the photosynthetic characteristics of the shade plant Camellia petelotii (Merr.) Sealy. Biologia 2022, 77, 3455–3468. [Google Scholar] [CrossRef]
- Gersony, J.T.; Hochberg, U.; Rockwell, F.E.; Park, M.; Gauthier, P.P.G.; Holbrook, N.M. Leaf Carbon Export and Nonstructural Carbohydrates in Relation to Diurnal Water Dynamics in Mature Oak Trees. Plant Physiol. 2020, 183, 1612–1621. [Google Scholar] [CrossRef]
- Mialet-Serra, I.; Clement-Vidal, A.; Roupsard, O.; Jourdan, C.; Dingkuhn, M. Whole-plant adjustments in coconut (Cocos nucifera) in response to sink-source imbalance. Tree Physiol. 2008, 28, 1199–1209. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J.; Foyer, C.H. Sink regulation of photosynthesis. J. Exp. Bot. 2001, 52, 1383–1400. [Google Scholar] [CrossRef] [PubMed]
- Von Schaewen, A.; Stitt, M.; Schmidt, R.; Sonnewald, U.; Willmitzer, L. Expression of a yeast-derived invertase in the cell wall of tobacco and Arabidopsis plants leads to accumulation of carbohydrate and inhibition of photosynthesis and strongly influences growth and phenotype of transgenic tobacco plants. EMBO J. 1990, 9, 3033–3044. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.J.; Muller, O.; Cohu, C.M.; Demmig-Adams, B.; Adams, W.W., 3rd. Quantification of Leaf Phloem Anatomical Features with Microscopy. Methods Mol. Biol. 2019, 2014, 55–72. [Google Scholar] [CrossRef]
- Braun, D.M. Phloem Loading and Unloading of Sucrose: What a Long, Strange Trip from Source to Sink. Annu. Rev. Plant Biol. 2022, 73, 553–584. [Google Scholar] [CrossRef]
- Turgeon, R. The Role of Phloem Loading Reconsidered. Plant Physiol. 2010, 152, 1817–1823. [Google Scholar] [CrossRef]
- Li, M.C.; Kong, G.Q.; Zhu, J.J. Vertical and leaf-age-related variations of nonstructural carbohydrates in two alpine timberline species, southeastern Tibetan Plateau. J. For. Res. 2009, 14, 229–235. [Google Scholar] [CrossRef]
- Zimmerman, R.C.; Kohrs, D.G.; Steller, D.L.; Alberte, R.S. Carbon Partitioning in Eelgrass (Regulation by Photosynthesis and the Response to Daily Light-Dark Cycles). Plant Physiol. 1995, 108, 1665–1671. [Google Scholar] [CrossRef]
- Campa, C.; Urban, L.; Mondolot, L.; Fabre, D.; Roques, S.; Lizzi, Y.; Aarrouf, J.; Doulbeau, S.; Breitler, J.-C.; Letrez, C.; et al. Juvenile Coffee Leaves Acclimated to Low Light Are Unable to Cope with a Moderate Light Increase. Front. Plant Sci. 2017, 8, 1126. [Google Scholar] [CrossRef]
- Zhuang, R. Camellia Oleifera in China, 2nd ed.; China Forestry Press: Beijing, China, 2008; p. 3. [Google Scholar]
- Conde, C.; Delrot, S.; Geros, H. Physiological, biochemical and molecular changes occurring during olive development and ripening. J. Plant Physiol. 2008, 165, 1545–1562. [Google Scholar] [CrossRef]
- Yang, S.; Liang, K.; Sun, Y.; Zhang, J.; Cao, Y.; Zhou, J.; Wang, A.; Zhang, L. Evidence of the predominance of passive symplastic phloem loading and sugar transport with leaf ageing in Camellia oleifera. Hortic. Plant J. 2023, in press. [Google Scholar] [CrossRef]
- Yang, S.W.; Liang, K.H.; Wang, A.B.; Zhang, M.; Qiu, J.M.; Zhang, L.Y. Physiological Characterization and Transcriptome Analysis of Camellia oleifera Abel. during Leaf Senescence. Forests 2020, 11, 812. [Google Scholar] [CrossRef]
- Galmés, J.; Ribas-Carbó, M.; Medrano, H.; Flexas, J. Rubisco activity in Mediterranean species is regulated by the chloroplastic CO2 concentration under water stress. J. Exp. Bot. 2011, 62, 653–665. [Google Scholar] [CrossRef]
- Han, L.J.; Fan, D.Y.; Wang, X.P.; Xu, C.Y.; Xia, X.L.; Chow, W.S. The Protective Role of Non-Photochemical Quenching in PSII Photo-Susceptibility: A Case Study in the Field. Plant Cell Physiol. 2023, 64, 43–54. [Google Scholar] [CrossRef]
- Jiang, C.D.; Gao, H.Y.; Zou, Q.; Jiang, G.; Li, L. The co-operation of leaf orientation, photorespiration and thermal dissipation alleviate photoinhibition in young leaves of soybean plants. Acta Ecol. Sin. 2005, 25, 319–325. [Google Scholar]
- Jeannette, E.; Reyss, A.; Grégory, N.; Gantet, P.; Prioul, J.L. Carbohydrate metabolism in a heat-girdled maize source leaf. Plant Cell Environ. 2000, 23, 61–69. [Google Scholar] [CrossRef]
- Azcon-Bieto, J. Inhibition of photosynthesis by carbohydrates in wheat leaves. Plant Physiol. 1983, 73, 681–686. [Google Scholar] [CrossRef]
- Sugiura, D.; Betsuyaku, E.; Terashima, I. Interspecific differences in how sink-source imbalance causes photosynthetic downregulation among three legume species. Ann. Bot. 2019, 123, 715–726. [Google Scholar] [CrossRef]
- Araya, T.; Noguchi, K.; Terashima, I. Effects of carbohydrate accumulation on photosynthesis differ between sink and source leaves of Phaseolus vulgaris L. Plant Cell Physiol. 2006, 47, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Huang, S.R.; Ai, Y.; Zhang, E.Z.; Wang, X.C.; Du, J.B.; Yang, W.Y. Comparative study on the different responses of maize photosynthesis to systemic regulation under light heterogeneity. Plant Sci. 2020, 301, 110666. [Google Scholar] [CrossRef]
- Wu, Q.J.; Chen, Z.D.; Sun, W.J.; Deng, T.T.; Chen, M.J. De novo Sequencing of the Leaf Transcriptome Reveals Complex Light-Responsive Regulatory Networks in Camellia sinensis cv. Baijiguan. Front. Plant Sci. 2016, 7, 332. [Google Scholar] [CrossRef] [PubMed]
- Mitamura, M.; Yamamura, Y.; Nakano, T. Large-scale canopy opening causes decreased photosynthesis in the saplings of shade-tolerant conifer, Abies veitchii. Tree Physiol. 2009, 29, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.V.; Machado, E.C.; Habermann, G.; Santos, M.G.; Oliveira, R.F. Seasonal effects on the relationship between photosynthesis and leaf carbohydrates in orange trees. Funct. Plant Biol. 2012, 39, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Moscatello, S.; Proietti, S.; Augusti, A.; Scartazza, A.; Walker, R.P.; Famiani, F.; Battistelli, A. Late summer photosynthesis and storage carbohydrates in walnut (Juglans regia L.): Feed-back and feed-forward effects. Plant Physiol. Biochem. 2017, 118, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Lanoue, J.; Leonardos, E.D.; Grodzinski, B. Effects of Light Quality and Intensity on Diurnal Patterns and Rates of Photo-Assimilate Translocation and Transpiration in Tomato Leaves. Front. Plant Sci. 2018, 9, 756. [Google Scholar] [CrossRef]
- De Swaef, T.; Driever, S.M.; Van Meulebroek, L.; Vanhaecke, L.; Marcelis, L.F.M.; Steppe, K. Understanding the effect of carbon status on stem diameter variations. Ann. Bot. 2013, 111, 31–46. [Google Scholar] [CrossRef]
- Ho, L.C. The Regulation of Carbon Transport and the Carbon Balance of Mature Tomato Leaves. Ann. Bot. 1978, 42, 155–164. [Google Scholar] [CrossRef]
- Nakai, H.; Yasutake, D.; Kimura, K.; Kengo, I.; Hidaka, K.; Eguchi, T.; Hirota, T.; Okayasu, T.; Ozaki, Y.; Kitano, M. Dynamics of carbon export from leaves as translocation affected by the coordination of carbohydrate availability in field strawberry. Environ. Exp. Bot. 2022, 196, 104806. [Google Scholar] [CrossRef]
- Liu, X.B.; Xu, B.; Gu, W.J.; Yin, Y.C.; Wang, H.C. Plant leaf veins coupling feature representation and measurement method based on DeepLabV3+. Front. Plant Sci. 2022, 13, 20. [Google Scholar] [CrossRef]
- Amiard, V.; Mueh, K.E.; Demmig-Adams, B.; Ebbert, V.; Turgeon, R.; Adams, W.W. Anatomical and photosynthetic acclimation to the light environment in species with differing mechanisms of phloem loading. Proc. Natl. Acad. Sci. USA 2005, 102, 12968–12973. [Google Scholar] [CrossRef]
- Ni, J.P.; Li, J.M.; Zhu, R.X.; Zhang, M.Y.; Qi, K.J.; Zhang, S.L.; Wu, J. Overexpression of sugar transporter gene PbSWEET4 of pear causes sugar reduce and early senescence in leaves. Gene 2020, 743, 144582. [Google Scholar] [CrossRef]
- Ninan, A.S.; Grant, J.; Song, J.C.; Jameson, P.E. Expression of Genes Related to Sugar and Amino Acid Transport and Cytokinin Metabolism during Leaf Development and Senescence in Pisum sativum L. Plants 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.A.; Lemonnier, P.; Quebedeaux, J.C.; Montes, C.M.; Leakey, A.D.B.; Ainsworth, E.A. Similar photosynthetic response to elevated carbon dioxide concentration in species with different phloem loading strategies. Photosynth. Res. 2018, 137, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Gao, J. Experimental Guidance for Plant Physiology; Higher Education Press: Beijing, China, 2006; pp. 74–77, 105–107. [Google Scholar]
- Reich, P.B.; Stefanski, A.; Rich, R.L.; Sendall, K.M.; Wei, X.R.; Zhao, C.M.; Hou, J.H.; Montgomery, R.A.; Bermudez, R. Assessing the relevant time frame for temperature acclimation of leaf dark respiration: A test with 10 boreal and temperate species. Glob. Change Biol. 2021, 27, 2945–2958. [Google Scholar] [CrossRef]
- Ye, Z.P. A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa. Photosynthetica 2007, 45, 637–640. [Google Scholar] [CrossRef]
- Ye, Z.P. A review on modeling of responses of photosynthesis to light and CO2. Chin. J. Plant Ecol. 2010, 34, 727–740. [Google Scholar]
- Ye, Z.P.; Yu, Q. A coupled model of stomatal conductance and photosynthesis for winter wheat. Photosynthetica 2008, 46, 637–640. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of QA Redox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Zhang, Z.; Qu, W. Experimental Guidance for Plant Physiology, 3rd ed.; Higher Education Press: Beijing, China, 2003; pp. 128–129. [Google Scholar]
- Kulichikhin, K.; Mukherjee, S.; Ayele, B. Extraction and Assays of ADP-Glucose Pyrophosphorylase, Soluble Starch Synthase and Granule Bound Starch Synthase from Wheat (Triticum aestivum L.) Grains. Bio-Protocol 2016, 6, e1929. [Google Scholar] [CrossRef]
- Price, C.A.; Symonova, O.; Mileyko, Y.; Hilley, T.; Weitz, J.S. Leaf Extraction and Analysis Framework Graphical User Interface: Segmenting and Analyzing the Structure of Leaf Veins and Areoles. Plant Physiol. 2011, 155, 236–245. [Google Scholar] [CrossRef]
- Ma, S.; Sun, L.L.; Sui, X.L.; Li, Y.X.; Chang, Y.; Fan, J.W.; Zhang, Z.X. Phloem loading in cucumber: Combined symplastic and apoplastic strategies. Plant J. 2019, 98, 391–404. [Google Scholar] [CrossRef] [PubMed]
- Yang, J. Improvement of traditional paraffin section preparation methods. J. Biol. 2006, 23, 45–46. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence |
|---|---|
| CoACTR3-F | GGTTGTAGTGGATGTTGGAGATGG |
| CoACTR3-R | AGCTGCTGGATGAAGAGAGTAAC |
| CoSUT1-F | ATGATAGTCCAACCCCTAGTCGG |
| CoSUT1-R | CCAGAACCCGACTACGAAAACC |
| CoSUT2-1-F | TGTGCGAATCTCAAGACCTG |
| CoSUT2-1-R | TGCCAACAATAGGTACCAC |
| CoSUT2-2-F | CAGATGGTGGTGTCAGTAGCAAG |
| CoSUT2-2-R | CAGCAATAGCCATGGGAACTTTG |
| CoSWEET12-F | CCAAAGAAAGAGAAGGGCAAGATCC |
| CoSWEET12-R | GCGTACATGATGATGGAGAAGATGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Zhang, L.; Wang, Q.; Liu, J.; Sun, Y. Diurnal Regulation of Leaf Photosynthesis Is Related to Leaf-Age-Dependent Changes in Assimilate Accumulation in Camellia oleifera. Plants 2023, 12, 2161. https://doi.org/10.3390/plants12112161
Zhang J, Zhang L, Wang Q, Liu J, Sun Y. Diurnal Regulation of Leaf Photosynthesis Is Related to Leaf-Age-Dependent Changes in Assimilate Accumulation in Camellia oleifera. Plants. 2023; 12(11):2161. https://doi.org/10.3390/plants12112161
Chicago/Turabian StyleZhang, Jinshun, Lingyun Zhang, Qi Wang, Jiali Liu, and Yongjiang Sun. 2023. "Diurnal Regulation of Leaf Photosynthesis Is Related to Leaf-Age-Dependent Changes in Assimilate Accumulation in Camellia oleifera" Plants 12, no. 11: 2161. https://doi.org/10.3390/plants12112161