
The Effect of Combined Drought and Temperature Stress on the Physiological Status of Calcareous Grassland Species as Potential Candidates for Urban Green Infrastructure
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. The Effect of Drought and Temperature Stress on Growth, Gas Exchange, and Pigment Content
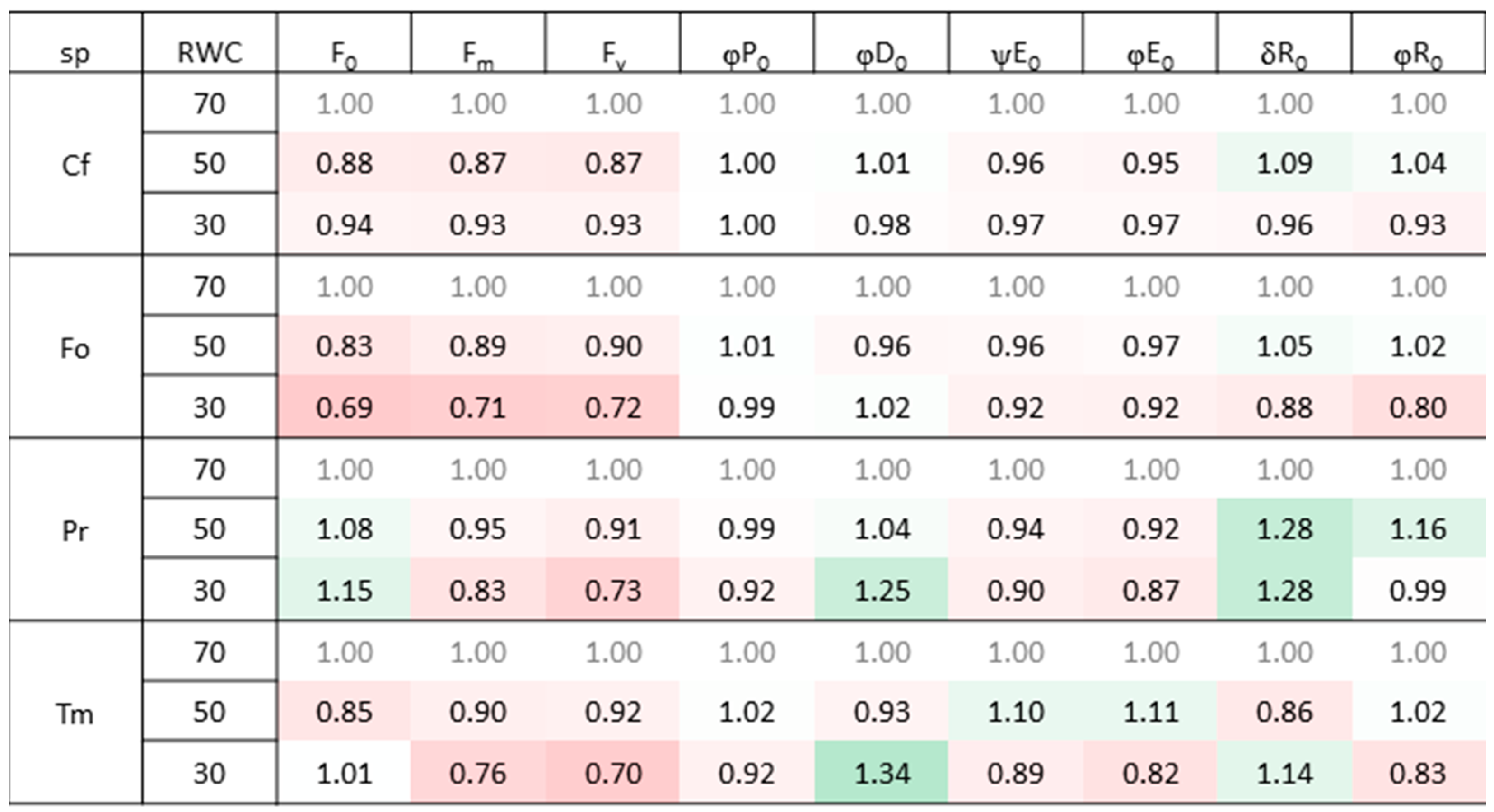
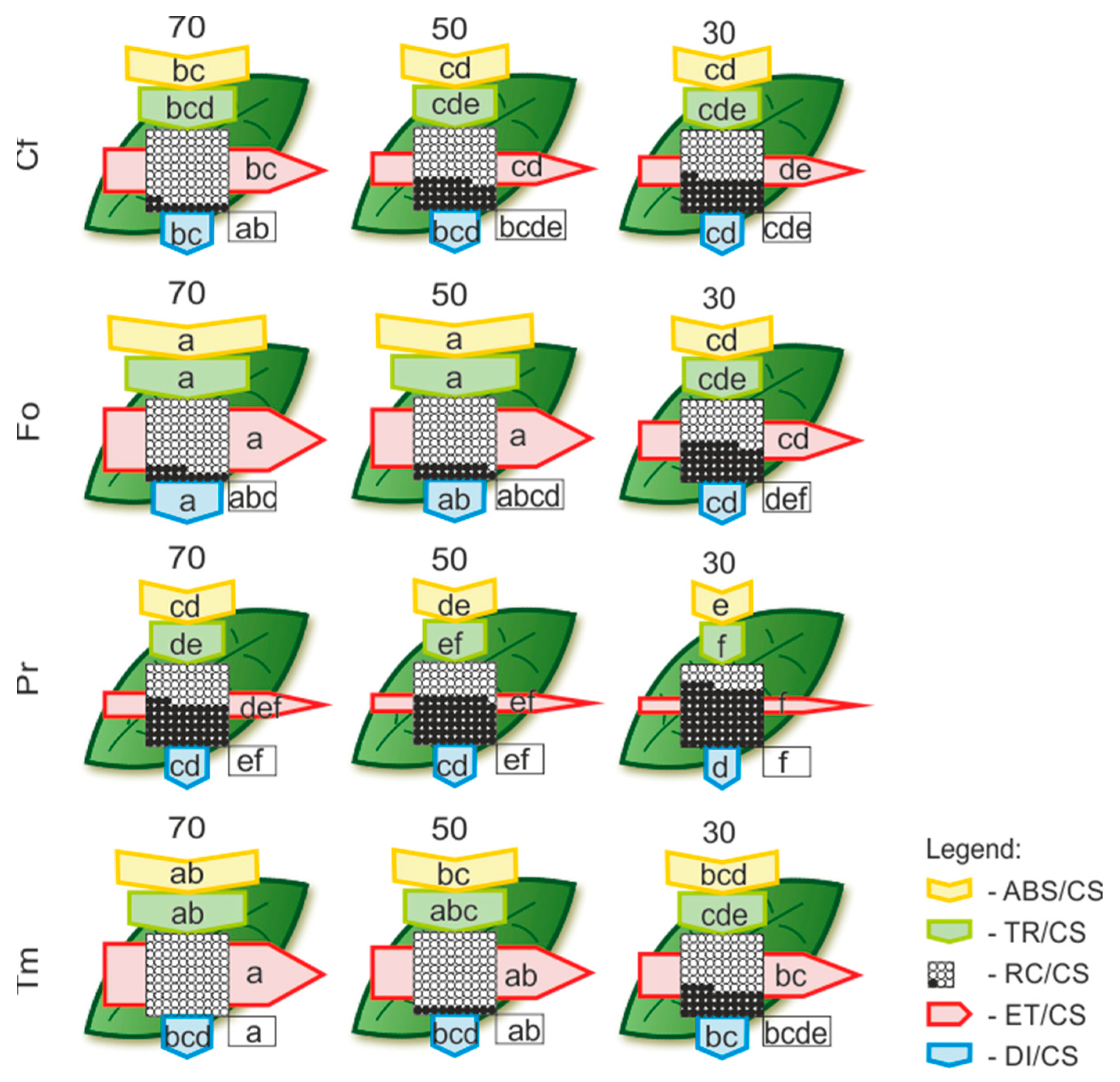
2.2. The Impact of Drought and Temperature Stress on Photosynthetic Apparatus
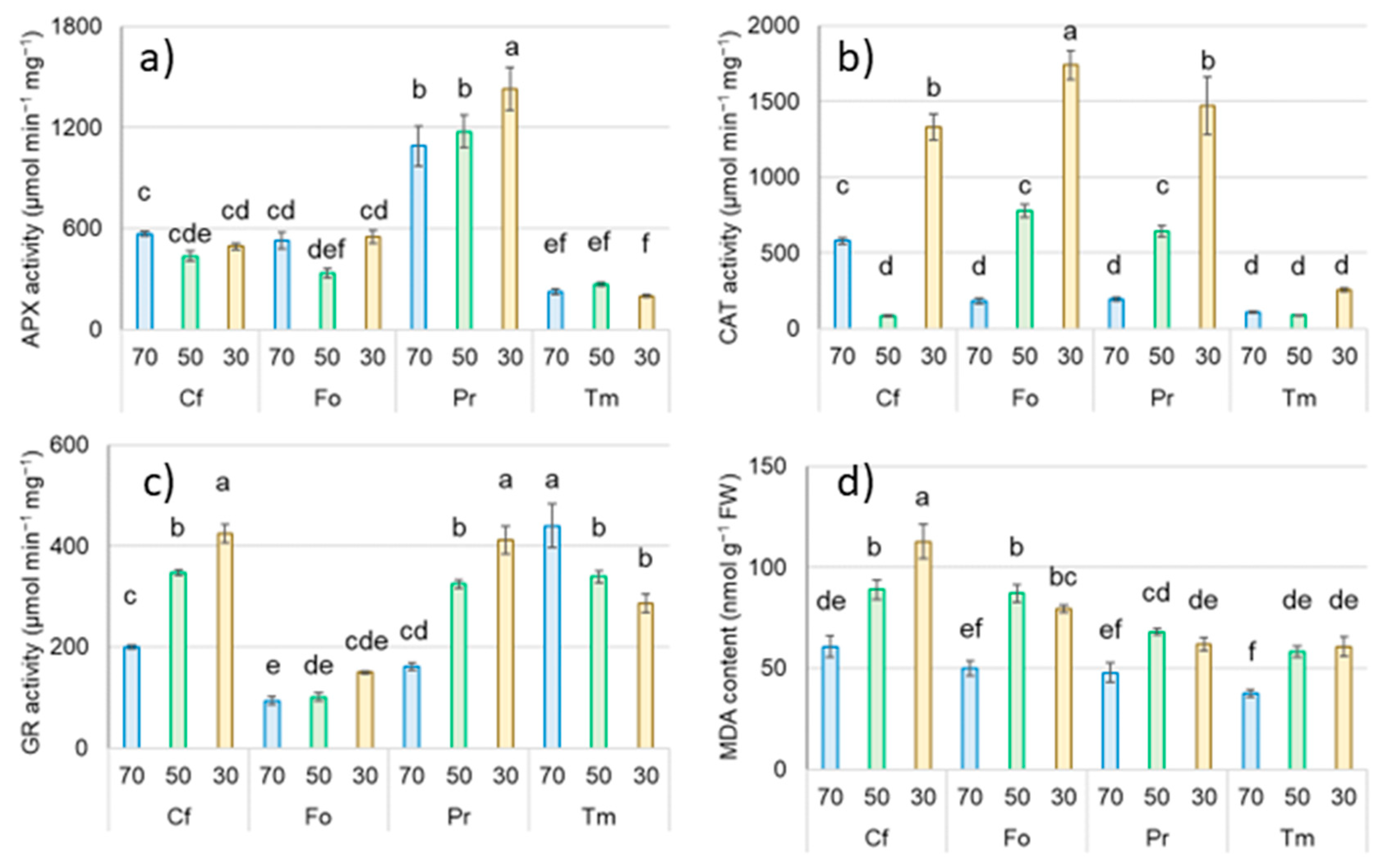
2.3. The Effect of Drought and Temperature Stress on Enzyme Activity and MDA Content
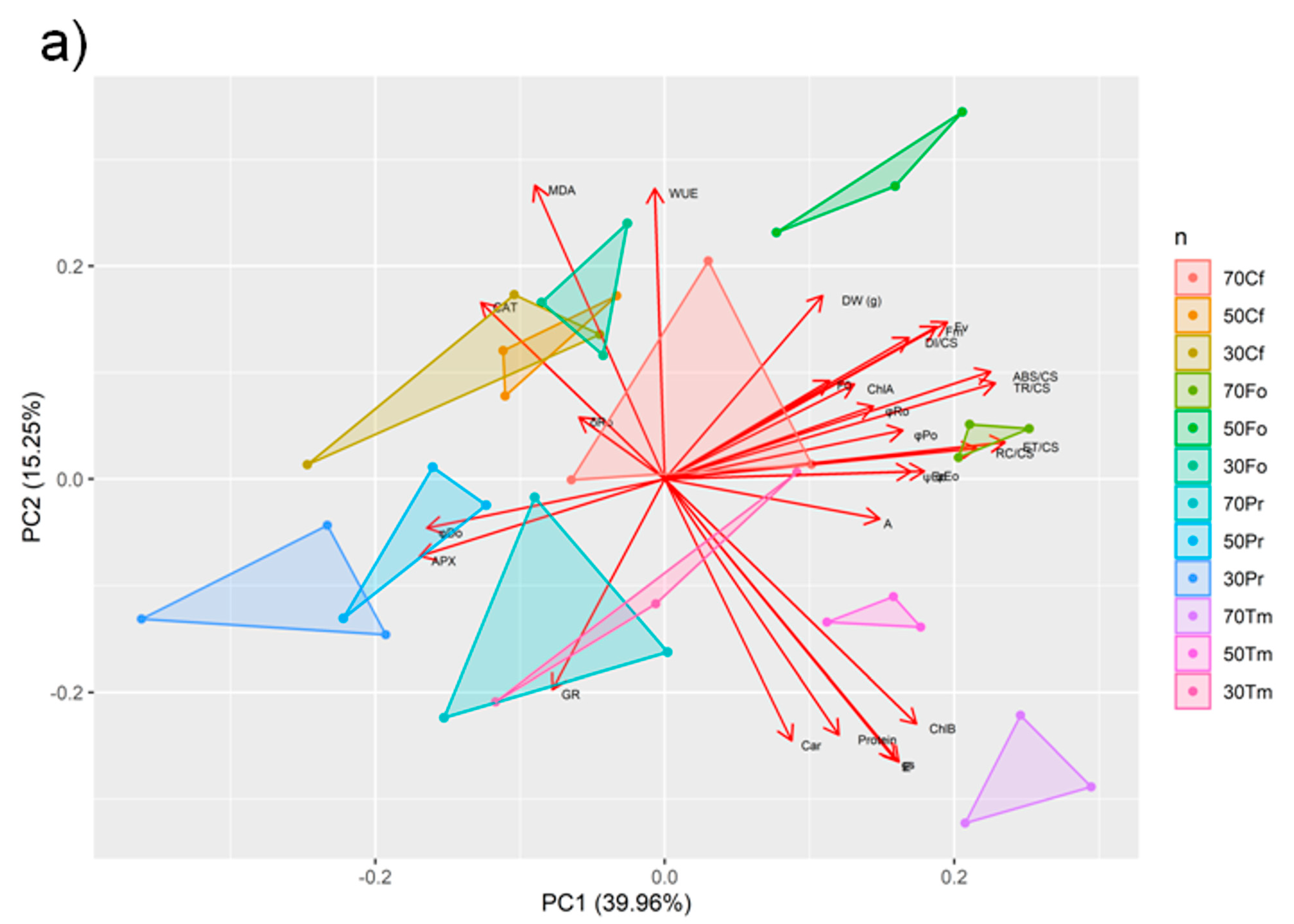
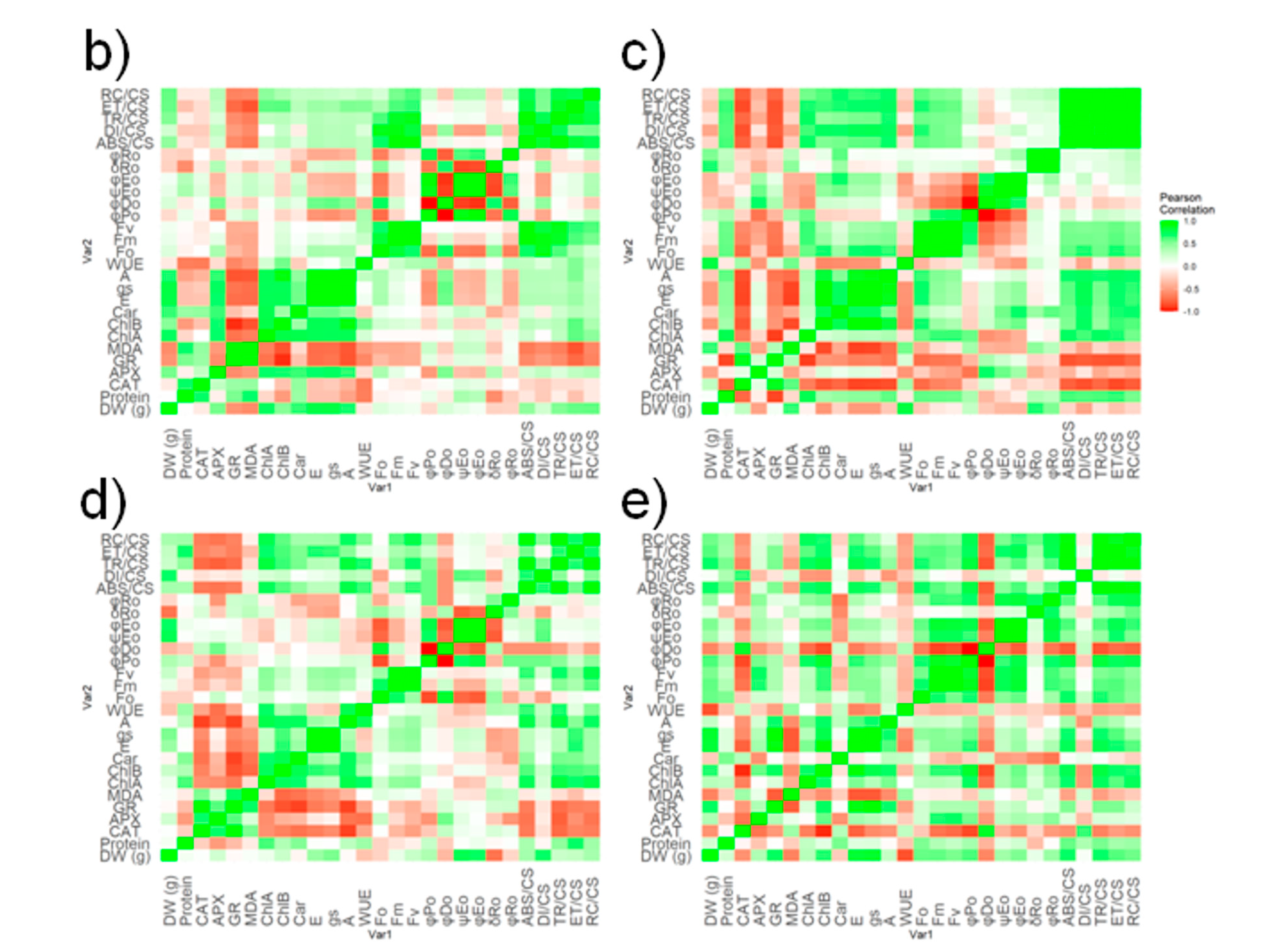
2.4. Principal Component Analysis and Corelations
3. Discussion
4. Materials and Methods
4.1. Experiment Design
4.2. Soil Substrate Physicochemical Parameters
4.3. Plant In Vivo Measurements
4.4. Plant Enzymatic Activity
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, N.H.; Tan, C.L.; Kolokotsa, D.D.; Takebayashi, H. Greenery as a mitigation and adaptation strategy to urban heat. Nat. Rev. Earth Environ. 2021, 2, 166–181. [Google Scholar] [CrossRef]
- Sharma, R.; Malaviya, P. Management of stormwater pollution using green infrastructure: The role of rain gardens. Wiley Interdiscip. Rev. Water 2021, 8, e1507. [Google Scholar] [CrossRef]
- Kerishnan, P.B.; Maruthaveeran, S.; Maulan, S. Investigating the usability pattern and constraints of pocket parks in Kuala Lumpur, Malaysia. Urban For. Urban Green. 2020, 50, 126647. [Google Scholar] [CrossRef]
- Calfapietra, C.; Peñuelas, J.; Niinemets, Ü. Urban plant physiology: Adaptation-mitigation strategies under permanent stress. Trends Plant Sci. 2015, 20, 72–75. [Google Scholar] [CrossRef]
- Shirgir, E.; Kheyroddin, R.; Behzadfar, M. Developing Strategic Principles of Intervention in Urban Green Infrastructure to Create and Enhance Climate Resilience in Cities—Case Study: Yousef Abad in Tehran. J. Clim. Chang. 2019, 5, 61–73. [Google Scholar] [CrossRef]
- Heim, A.; Xie, G.; Lundholm, J. Functional and Phylogenetic Characteristics of Vegetation: Effects on Constructed Green Infrastructure. In Urban Services to Ecosystems, 1st ed.; Catalano, C., Andreucci, M.B., Guarino, R., Bretzel, F., Leone, M., Pasta, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; Volume 17, pp. 61–83. [Google Scholar] [CrossRef]
- Manso, M.; Teotónio, I.; Silva, C.M.; Cruz, C.O. Green roof and green wall benefits and costs: A review of the quantitative evidence. Renew. Sustain. Energy Rev. 2021, 135, 110111. [Google Scholar] [CrossRef]
- Lampinen, J.; García-Antúnez, O.; Olafsson, A.S.; Kavanagh, K.C.; Gulsrud, N.M.; Raymond, C.M. Envisioning carbon-smart and just urban green infrastructure. Urban For. Urban Green. 2022, 75, 127682. [Google Scholar] [CrossRef]
- Radhakrishnan, M.; Kenzhegulova, I.; Eloffy, M.G.; Ibrahim, W.A.; Zevenbergen, C.; Pathirana, A. Development of context specific sustainability criteria for selection of plant species for green urban infrastructure: The case of Singapore. Sustain. Prod. Consum. 2019, 20, 316–325. [Google Scholar] [CrossRef]
- Farrell, C.; Szota, C.; Williams, N.S.; Arndt, S.K. High water users can be drought tolerant: Using physiological traits for green roof plant selection. Plant Soil 2013, 372, 177–193. [Google Scholar] [CrossRef]
- Hummel, I.; Pantin, F.; Sulpice, R.; Piques, M.; Rolland, G.; Dauzat, M.; Christophe, A.; Pervent, M.; Bouteille, M.; Stitt, M.; et al. Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: An integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol. 2010, 154, 357–372. [Google Scholar] [CrossRef]
- Al-Yasi, H.; Attia, H.; Alamer, K.; Hassan, F.; Ali, E.; Elshazly, S.; Siddique, K.H.M.; Hessini, K. Impact of drought on growth, photosynthesis, osmotic adjustment, and cell wall elasticity in Damask rose. Plant Physiol. Biochem. 2020, 150, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Bury, M.; Rusinowski, S.; Sitko, K.; Krzyżak, J.; Kitczak, T.; Możdżer, E.; Siwek, H.; Włodarczyk, M.; Zieleźnik-Rusinowska, P.; Szada-Borzyszkowska, A.; et al. Physiological status and biomass yield of Sida hermaphrodita (L.) Rusby cultivated on two distinct marginal lands in Southern and Northern Poland. Ind. Crop Prod. 2021, 167, 113502. [Google Scholar] [CrossRef]
- Dai, H.; Wei, S.; Pogrzeba, M.; Rusinowski, S.; Krzyżak, J.; Jia, G. Exogenous jasmonic acid decreased Cu accumulation by alfalfa and improved its photosynthetic pigments and antioxidant system. Ecotoxicol. Environ. Saf. 2020, 190, 110176. [Google Scholar] [CrossRef]
- Landi, S.; Hausman, J.F.; Guerriero, G.; Esposito, S. Poaceae vs. abiotic stress: Focus on drought and salt stress, recent insights and perspectives. Front. Plant Sci. 2017, 8, 1214. [Google Scholar] [CrossRef]
- Lundholm, J.T. Green roofs and facades: A habitat template approach. Urban Habitats 2006, 4, 87–101. [Google Scholar]
- Mazalla, L.; Diekmann, M.; Duprè, C. Microclimate shapes vegetation response to drought in calcareous grasslands. Appl. Veg. Sci. 2022, 25, e12672. [Google Scholar] [CrossRef]
- Lehmair, T.A.; Pagel, E.; Poschlod, P.; Reisch, C. Surrounding landscape structures, rather than habitat age, drive genetic variation of typical calcareous grassland plant species. Landsc. Ecol. 2020, 35, 2881–2893. [Google Scholar] [CrossRef]
- Joosten, H.; Clarke, D. Wise Use of Mires and Peatlands—Background and Principles Including a Framework for Decision Making, 1st ed.; International Mire Conservation Group and International Peat Society: Devon, UK, 2002. [Google Scholar]
- Lamers, L.P.; Vile, M.A.; Grootjans, A.P.; Acreman, M.C.; van Diggelen, R.; Evans, M.G.; Richardson, C.J.; Rochefort, L.; Kooijman, A.M.; Roelofs, J.G.M.; et al. Ecological restoration of rich fens in Europe and North America: From trial and error to an evidence-based approach. Biol. Rev. 2015, 90, 182–203. [Google Scholar] [CrossRef] [PubMed]
- Hájek, M.; Horsáková, V.; Hájková, P.; Coufal, R.; Dítě, D.; Němec, T.; Horsák, M. Habitat extremity and conservation management stabilise endangered calcareous fens in a changing world. Sci. Total Environ. 2020, 719, 134693. [Google Scholar] [CrossRef]
- Van Mechelen, C.; Dutoit, T.; Hermy, M. Adapting green roof irrigation practices for a sustainable future: A review. Sustain. Cities Soc. 2015, 19, 74–90. [Google Scholar] [CrossRef]
- Kaluarachchi, Y. Potential advantages in combining smart and green infrastructure over silo approaches for future cities. Front. Eng. Manag. 2021, 8, 98–108. [Google Scholar] [CrossRef]
- Nitoslawski, S.A.; Galle, N.J.; Van Den Bosch, C.K.; Steenberg, J.W. Smarter ecosystems for smarter cities? A review of trends, technologies, and turning points for smart urban forestry. Sustain. Cities Soc. 2019, 51, 101770. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Rayner, J.P.; Farrell, C.; Raynor, K.J.; Murphy, S.M.; Williams, N.S. Plant establishment on a green roof under extreme hot and dry conditions: The importance of leaf succulence in plant selection. Urban For. Urban Green. 2016, 15, 6–14. [Google Scholar] [CrossRef]
- Kinlocka, N.L.; Schindler, B.Y.; Gurevitch, J. Biological invasions in the context of green roofs. Isr. J. Ecol. Evol. 2016, 62, 32–43. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef]
- Volk, M.; Niklaus, P.A.; Körner, C. Soil moisture effects determine CO2 responses of grassland species. Oecologia 2000, 125, 380–388. [Google Scholar] [CrossRef]
- Getter, K.L.; Rowe, D.B. Selecting Plants for Extensive Green Roofs in the United States; Michigan State University Extension: East Lansing, MI, USA, 2008. [Google Scholar]
- Wang, J.; Burgess, P.; Bonos, S.A.; Meyer, W.A.; Huang, B. Differential physiological responses and genetic variations in fine fescue species for heat and drought stress. J. Am. Soc. Hortic. Sci. 2017, 142, 367–375. [Google Scholar] [CrossRef]
- Khoshkholghsima, N.A.; Rohollahi, I. Evaluating biochemical response of some selected perennial grasses under drought stress in Iran. Hortic. Environ. Biotechnol. 2015, 56, 383–390. [Google Scholar] [CrossRef]
- Vahdati, N.; Tehranifar, A.; Kazemi, F. Assessing chilling and drought tolerance of different plant genera on extensive green roofs in an arid climate region in Iran. J. Environ. Manag. 2017, 192, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Seyedabadi, M.R.; Karrabi, M.; Nabati, J. Investigating green roofs’ CO2 sequestration with cold- and drought-tolerant plants (a short- and long-term carbon footprint view). Environ. Sci. Pollut. Res. 2022, 29, 14121–14130. [Google Scholar] [CrossRef] [PubMed]
- Van der Kolk, H.J.; van den Berg, P.; Korthals, G.; Bezemer, T.M. Shading enhances plant species richness and diversity on an extensive green roof. Urban Ecosyst. 2020, 23, 935–943. [Google Scholar] [CrossRef]
- Matsuoka, T.; Tsuchiya, K.; Yamada, S.; Lundholm, J.; Okuro, T. The effects of growth form on the impact of companion planting of nectar-producing plant species with Sedum album for extensive green roofs. Urban For. Urban Green. 2020, 56, 126875. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Ma, X. Different response on drought tolerance and post-drought recovery between the small-leafed and the large-leafed white clover (Trifolium repens L.) associated with antioxidative enzyme protection and lignin metabolism. Acta Physiol. Plant. 2013, 35, 213–222. [Google Scholar] [CrossRef]
- Reynolds-Henne, C.E.; Langenegger, A.; Mani, J.; Schenk, N.; Zumsteg, A.; Feller, U. Interactions between temperature, drought and stomatal opening in legumes. Environ. Exp. Bot. 2010, 68, 37–43. [Google Scholar] [CrossRef]
- PN-ISO 11265:1997; Soil Quality—Determination of the Specific Electrical Conductivity. PKN: Warsaw, Poland, 1997.
- PNR-04032:1998; Soils and Mineral Soil Materials—Soil Sampling and Determination of Particle Size Distribution in Mineral Soil Material. PKN: Warsaw, Poland, 1998. (In Polish)
- ISO 13878:1998; Determination of Total Nitrogen Content by Dry Combustion (“Elemental Analysis”). ISO: Geneva, Switzerland, 1998.
- Egner, H.; Riehm, H.; Domingo, W.R. Investigations on the chemical soil analysis as a basis for assessing the soil nutrient status. II: Chemical extraction methods for phosphorus and potassium determination. K. Lantbr. Ann. 1960, 26, 199–215. [Google Scholar]
- Bremner, J.M. Chapter 37 Nitrogen total. In Methods of Soil Analysis: Part 3 Chemical Methods, 5th ed.; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Soltanpour, P.N., Tabatabai, M.A., Johnston, C.T., Sumner, M.E., Eds.; The American Society of Agronomy, Soil Science Society of America: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar] [CrossRef]
- PN-EN 14582:2017; Characterisation of Waste—Halogen and Sulphur Content—Oxygen Incineration in Closed Systems and Methods of Determination. PKN: Wasaw, Poland, 2017. (In English)
- Rusinowski, S.; Krzyżak, J.; Clifton-Brown, J.; Jensen, E.; Mos, M.; Webster, R.; Sitko, K.; Pogrzeba, M. New Miscanthus Hybrids Cultivated at a Polish Metal-Contaminated Site Demonstrate High Stomatal Regulation and Reduced Shoot Pb and Cd Concentrations. Environ. Pollut. 2019, 252, 1377–1387. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. Oxidative stress and antioxidative systems: Recipes for successful data collection and interpretation. Plant Cell Environ. 2016, 39, 1140–1160. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Barnes, J.D.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A.W. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Vimala, T.; Poonghuzhali, T.V. Estimation of pigments from seaweeds by using acetone and DMSO. Int. J. Sci. Res. 2015, 4, 1850–1854. Available online: https://www.ijsr.net/get_abstract.php?paper_id=SUB159224 (accessed on 1 April 2023).
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sp | RWC | Shoot Dry Weight (g) | Protein Content (mg g−1 FW) | Chl A Concentration (mg g−1 FW) | Chl B Concentration (mg g−1 FW) | Carotenoids Concentration (mg g−1 FW) | Transpiration (mmol m−2 s−1) | Stomatal Conductance (mmol m−2 s−1) | Photosynthesis Rate (µmol CO2 m−2 s−1) | Water Use Efficiency |
|---|---|---|---|---|---|---|---|---|---|---|
| Cf | 70 | 4.99 ± 1.34 cd | 7.9 ± 0.4 c | 1.78 ± 0.17 bc | 0.44 ± 0.04 e | 1.11 ± 0.12 de | 1.04 ± 0.28 bcd | 0.15 ± 0.05 bcd | 11.72 ± 3.04 a | 89.6 ± 15.5 a |
| 50 | 3.47 ± 0.21 d | 7.6 ± 0.9 c | 1.01 ± 0.12 e | 0.11 ± 0.01 fg | 0.63 ± 0.03 f | 0.57 ± 0.16 ef | 0.07 ± 0.02 cd | 5.23 ± 1.06 cd | 83.2 ± 13.1 a | |
| 30 | 3.41 ± 0.29 d | 11.2 ± 1.1 c | 1.14 ± 0.12 de | 0.01 ± 0.01 g | 0.99 ± 0.12 e | 0.38 ± 0.05 f | 0.04 ± 0.00 d | 1.92 ± 0.37 d | 43.5 ± 4.9 bcd | |
| Fo | 70 | 8.16 ± 1.04 bc | 9.6 ± 0.5 c | 3.07 ± 0.35 a | 1.05 ± 0.08 c | 1.45 ± 0.13 abc | 1.41 ± 0.01 b | 0.26 ± 0.01 b | 11.98 ± 1.26 a | 46.6 ± 4.6 bcd |
| 50 | 13.56 ± 1.59 a | 9.4 ± 0.6 c | 3.27 ± 0.20 a | 0.19 ± 0.01 f | 1.19 ± 0.09 cde | 0.74 ± 0.03 cdef | 0.10 ± 0.01 cd | 7.73 ± 0.39 bc | 77.5 ± 2.7 ab | |
| 30 | 10.82 ± 2.32 ab | 8.0 ± 0.2 c | 2.32 ± 0.21 b | 0.10 ± 0.00 fg | 1.03 ± 0.08 de | 0.57 ± 0.01 ef | 0.06 ± 0.00 d | 4.19 ± 1.19 cd | 64.8 ± 15.3 abc | |
| Pr | 70 | 5.00 ± 1.85 cd | 10.3 ± 1.3 c | 2.33 ± 0.29 b | 0.71 ± 0.07 d | 1.70 ± 0.10 a | 1.12 ± 0.24 bc | 0.18 ± 0.06 bc | 7.32 ± 1.07 bc | 58.8 ± 30.1 abc |
| 50 | 4.20 ± 0.89 d | 8.6 ± 1.1 c | 1.79 ± 0.10 bc | 0.14 ± 0.03 fg | 1.13 ± 0.12 de | 0.82 ± 0.10 cde | 0.12 ± 0.02 cd | 4.63 ± 0.34 cd | 41.7 ± 6.3 bcd | |
| 30 | 4.16 ± 1.70 d | 9.6 ± 1.002 c | 1.67 ± 0.12 cd | 0.20 ± 0.00 f | 1.14 ± 0.07 cde | 0.66 ± 0.07 def | 0.08 ± 0.01 cd | 1.73 ± 0.13 d | 21.9 ± 1.4 d | |
| Tm | 70 | 6.38 ± 0.55 cd | 38.6 ± 4.1 a | 2.04 ± 0.18 bc | 1.53 ± 0.08 a | 1.65 ± 0.07 a | 1.81 ± 0.10 a | 0.48 ± 0.10 a | 9.27 ± 1.23 ab | 19.9 ± 2.1 d |
| 50 | 5.13 ± 0.82 cd | 28.1 ± 1.6 b | 1.94 ± 0.12 bc | 1.31 ± 0.06 b | 1.32 ± 0.17 bcd | 1.41 ± 0.11 b | 0.25 ± 0.02 b | 7.21 ± 2.11 bc | 31.3 ± 12.7 cd | |
| 30 | 4.26 ± 0.18 d | 32.4 ± 4.5b | 1.59 ± 0.05 cd | 0.10 ± 0.00 fg | 1.63 ± 0.13 ab | 0.90 ± 0.08 cde | 0.13 ± 0.01 cd | 4.41 ± 0.73 cd | 33.7 ± 2.5 cd |
| pHH2O | 8.46 ± 0.04 |
| pHKCl | 7.83 ± 0.01 |
| EC [µS cm−1] | 75.75 ± 3.27 |
| Ntotal [% d.w] | 0.13 ± 0.00 |
| P [mg kg−1 d.w] | 318.8 ± 4.4 |
| K [ mg kg−1 d.w] | 1123 ± 55 |
| Ca [ mg kg−1 d.w] | 58,330 ± 1510 |
| Mg [ mg kg−1 d.w] | 30,860 ± 840 |
| P2O5 available [mg 100 g −1 d.w] | 0.15 ± 0.02 |
| K2O available [mg 100 g −1 d.w] | 13.4 ± 0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyżak, J.; Rusinowski, S.; Sitko, K.; Szada-Borzyszkowska, A.; Borgulat, J.; Stec, R.; Hanslin, H.M.; Pogrzeba, M. The Effect of Combined Drought and Temperature Stress on the Physiological Status of Calcareous Grassland Species as Potential Candidates for Urban Green Infrastructure. Plants 2023, 12, 2003. https://doi.org/10.3390/plants12102003
Krzyżak J, Rusinowski S, Sitko K, Szada-Borzyszkowska A, Borgulat J, Stec R, Hanslin HM, Pogrzeba M. The Effect of Combined Drought and Temperature Stress on the Physiological Status of Calcareous Grassland Species as Potential Candidates for Urban Green Infrastructure. Plants. 2023; 12(10):2003. https://doi.org/10.3390/plants12102003
Chicago/Turabian StyleKrzyżak, Jacek, Szymon Rusinowski, Krzysztof Sitko, Alicja Szada-Borzyszkowska, Jacek Borgulat, Radosław Stec, Hans Martin Hanslin, and Marta Pogrzeba. 2023. "The Effect of Combined Drought and Temperature Stress on the Physiological Status of Calcareous Grassland Species as Potential Candidates for Urban Green Infrastructure" Plants 12, no. 10: 2003. https://doi.org/10.3390/plants12102003