

Reproductive Ecology of the Invasive Alien Shrub Pyracantha angustifolia in the Grassland Biome, South Africa
 , ,
, ,
Abstract
:1. Introduction
2. Results
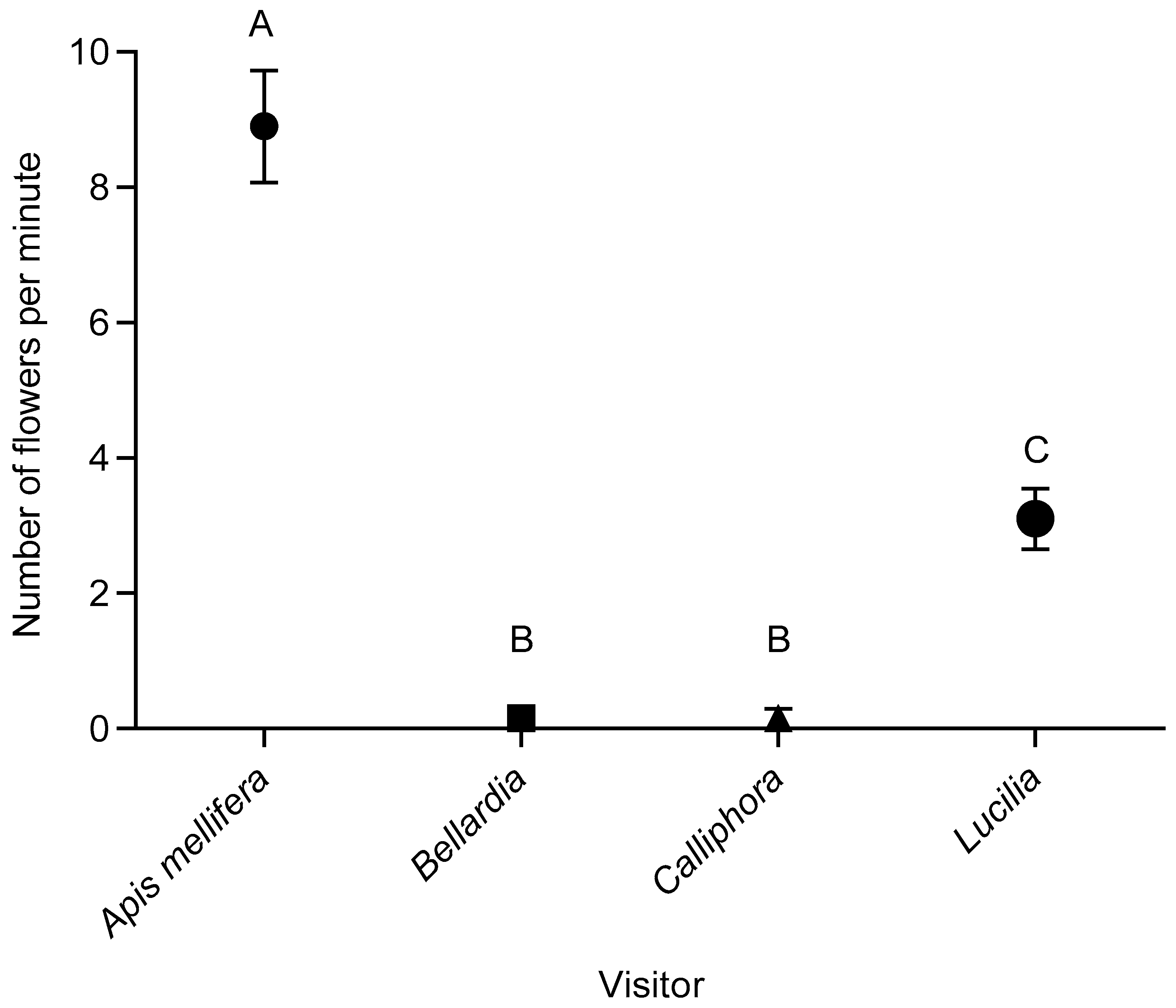
2.1. Visitation Frequencies of Floral Visitors
2.2. Pollen Loads
2.3. Pollinator Exclusions
2.4. Shrub Size Distributions
2.5. Fruit Estimates
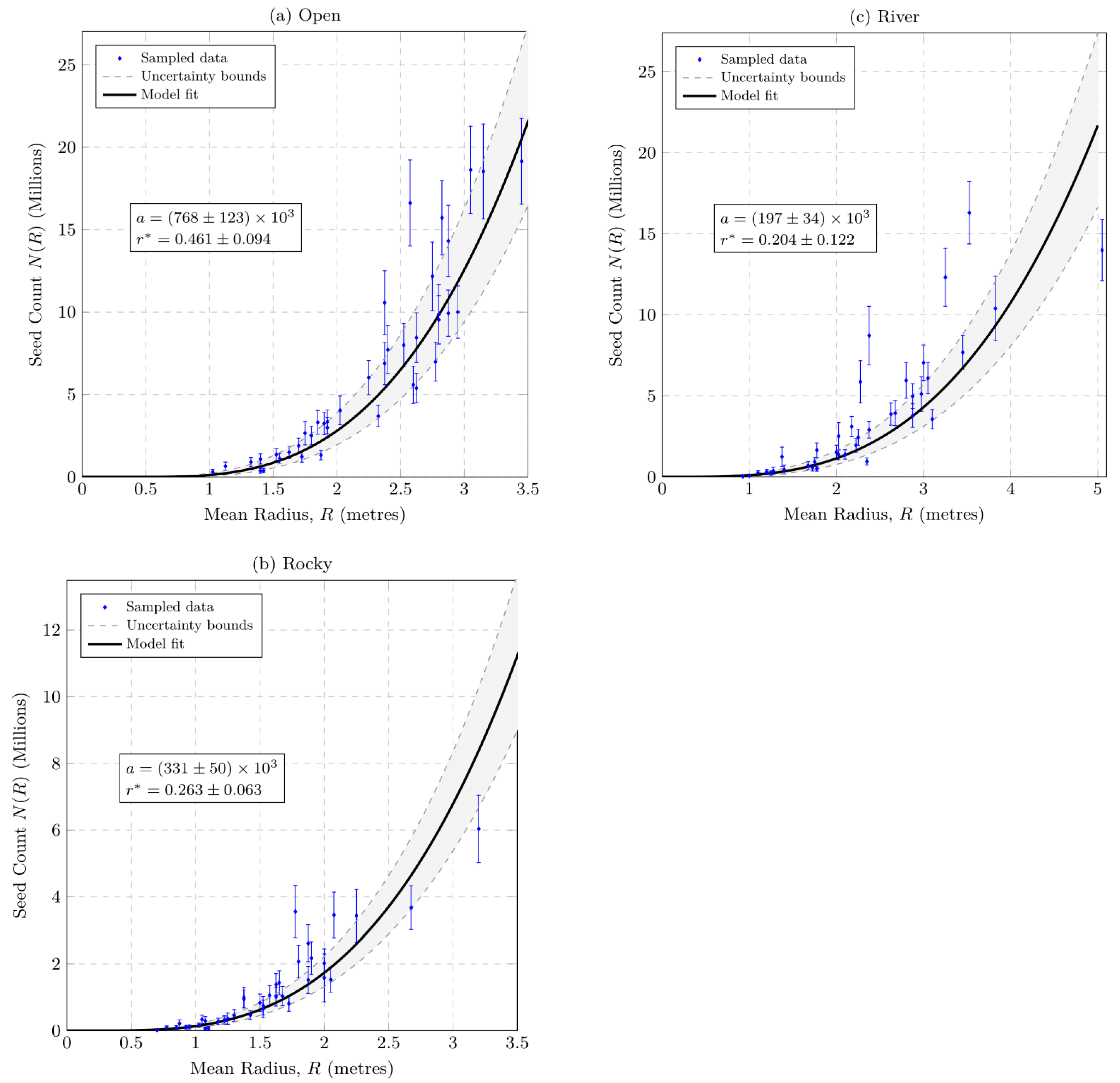
2.6. Pyracantha angustifolia Population Seed Counts
2.7. Seed Rain
2.8. Soil Seed Bank
2.9. Seed Viability and Longevity in the Soil
3. Discussion
4. Materials and Methods
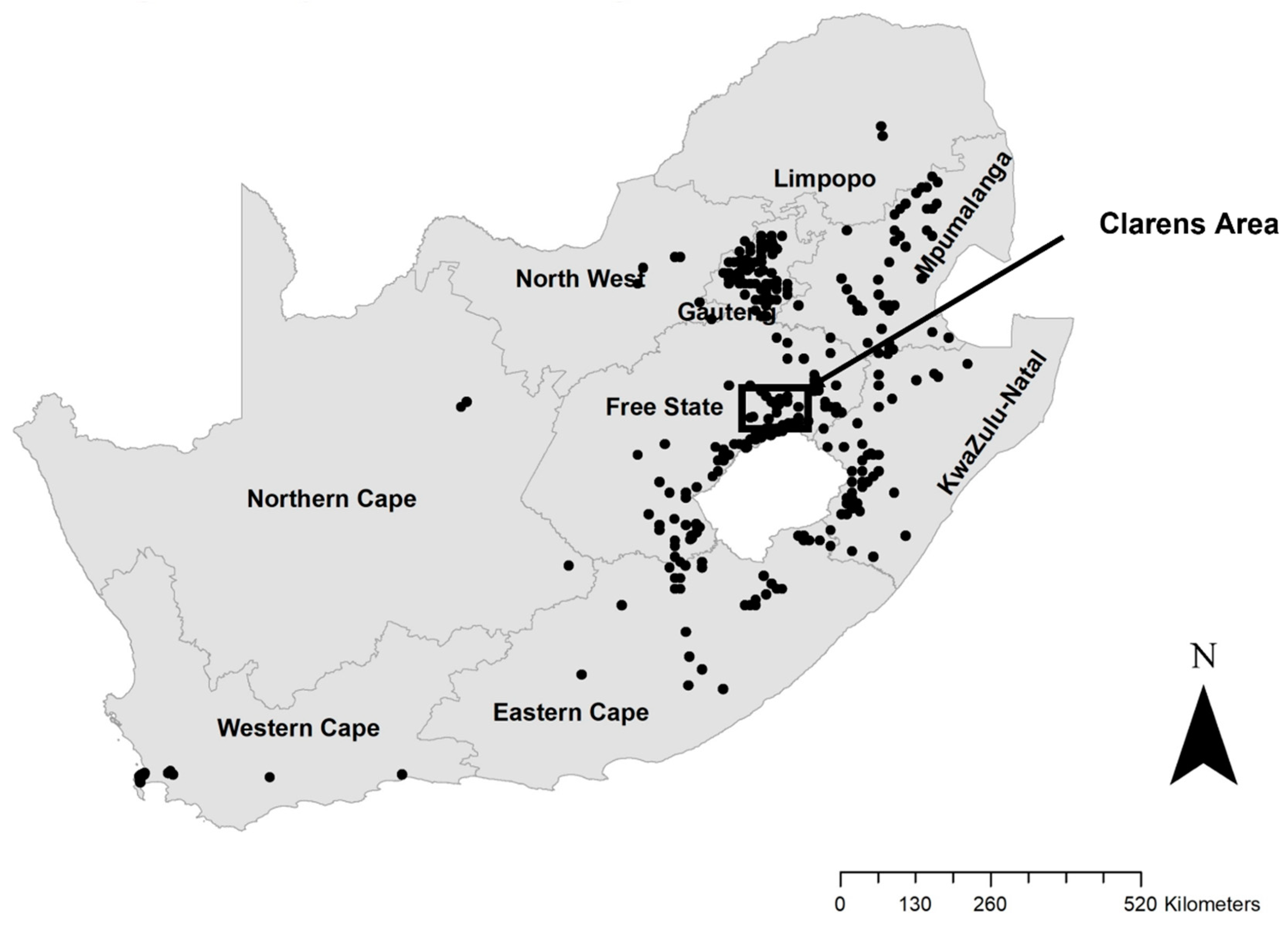
4.1. Study Area
4.2. Visitation Frequencies of Floral Visitors
4.3. Pollen Loads
4.4. Pollinator Exclusions
4.5. Fruit Estimates
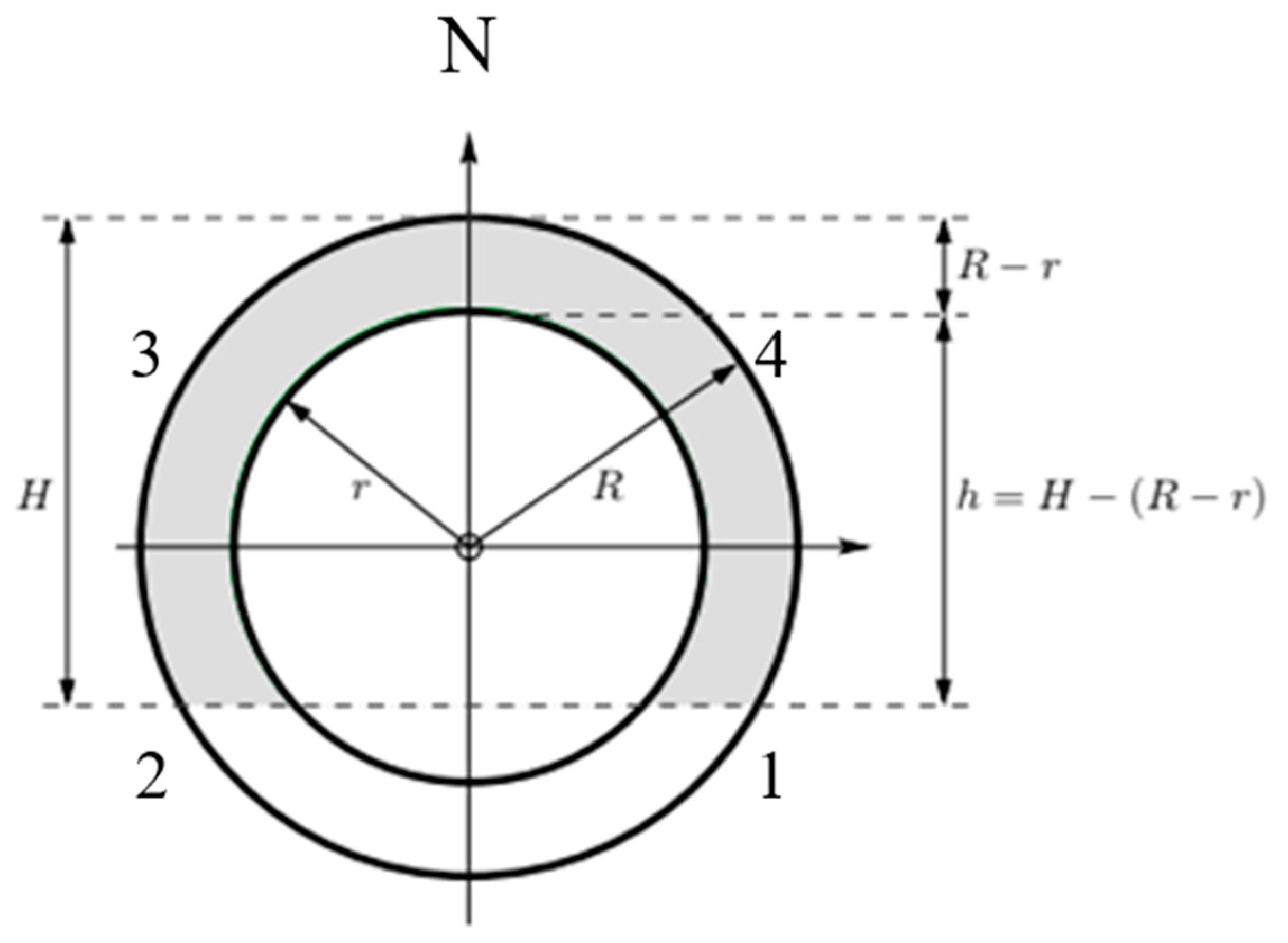
4.6. Seed Count Model
- (a)
- Determining seed counts.
- (b)
- Functional relationship.
4.7. Seed Rain
4.8. Soil Seed Bank
4.9. Seed Viability and Longevity in the Soil
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Connor, T.G.; van Wilgen, B.W. The impact of invasive alien plants on rangelands in South Africa. Biol. Invasions S. Afr. 2020, 14, 459–487. [Google Scholar]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kleunen, M.; Dawson, W.; Maurel, N. Characteristics of successful alien plants. Mol. Ecol. 2015, 24, 1954–1968. [Google Scholar] [CrossRef] [Green Version]
- Milbau, A.; Stout, J.C. Factors associated with alien plants transitioning from casual, to naturalized, to invasive. Conserv. Biol. 2008, 22, 308–317. [Google Scholar] [CrossRef]
- Tandon, R.; Shivanna, K.R.; Koul, M. (Eds.) Reproductive Ecology of Flowering Plants: Patterns and Processes; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Chen, X.; Liu, W.; Pennings, S.C.; Zhang, Y. Plasticity and selection drive hump-shaped latitudinal patterns of flowering phenology in an invasive intertidal plant. Ecology 2021, 102, e03311. [Google Scholar] [CrossRef] [PubMed]
- Martin-Albarracin, V.L.; Amico, G.C. Plant origin and fruit traits shape fruit removal patterns by native birds in invaded plant communities. Biol. Invasions 2021, 23, 857–870. [Google Scholar] [CrossRef]
- Chikowore, G.; Steenhuisen, S.L.; Mutamiswa, R.; Martin, G.D.; Chidawanyika, F. Integration of invasive tree, black locust, into agro-ecological flower visitor networks induces competition for pollination services. Arthropod-Plant Interact. 2021, 15, 787–796. [Google Scholar] [CrossRef]
- Parra-Tabla, V.; Arceo-Gómez, G. Impacts of plant invasions in native plant–pollinator networks. New Phytol. 2021, 230, 2117–2128. [Google Scholar] [CrossRef]
- Sperry, J.H.; O’Hearn, D.; Drake, D.R.; Hruska, A.M.; Case, S.B.; Vizentin-Bugoni, J.; Arnett, C.; Chambers, T.; Tarwater, C.E. Fruit and seed traits of native and invasive plant species in Hawaii: Implications for seed dispersal by non-native birds. Biol. Invasions 2021, 23, 1819–1835. [Google Scholar] [CrossRef]
- Yannelli, F.A.; Karrer, G.; Hall, R.; Kollmann, J.; Heger, T. Seed density is more effective than multi-trait limiting similarity in controlling grassland resistance against plant invasions in mesocosms. Appl. Veg. Sci. 2018, 21, 411–418. [Google Scholar] [CrossRef]
- Molefe, K.L.; Tedder, M.J.; Thabethe, V.; Rushworth, I.; Downs, C.T. Role of native avian frugivores in germination facilitation and potential dispersal of invasive American bramble (Rubus cuneifolius) in South Africa. Biol. Invasions 2020, 22, 1109–1120. [Google Scholar] [CrossRef]
- Henderson, L. Invasive Alien Plants in South Africa. In Plant Protection Research Institute Handbook No. 21; Agricultural Research Council: Pretoria, South Africa, 2020. [Google Scholar]
- Chikowore, G.; Mutamiswa, R.; Sutton, G.F.; Chidawanyika, F.; Martin, G.D. Reduction of grazing capacity in high-elevation rangelands after Black locust invasion in South Africa. Rangel. Ecol. Manag. 2021, 76, 109–117. [Google Scholar] [CrossRef]
- Carbutt, C. The emerging invasive alien plants of the Drakensberg Alpine Centre, southern Africa. Bothalia 2012, 42, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Yapi, T.S.; O’Farrell, P.J.; Dziba, L.E.; Esler, K.J. Alien tree invasion into a South African montane grassland ecosystem: Impact of Acacia species on rangeland condition and livestock carrying capacity. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2018, 14, 105–116. [Google Scholar] [CrossRef]
- Humphrey, L.; Fraser, G.; Martin, G. The economic implications of Robinia pseudoacacia L. (black locust) on agricultural production in South Africa. Agrekon 2019, 58, 216–228. [Google Scholar] [CrossRef]
- Richardson, D.M.; Foxcroft, L.C.; Latombe, G.; Le Maitre, D.C.; Rouget, M.; Wilson, J.R. The biogeography of South African terrestrial plant invasions. In Biological Invasions in South Africa; Springer: Berlin/Heidelberg, Germany, 2020; pp. 67–96. [Google Scholar]
- Kobisi, K.; Seleteng-Kose, L.; Moteetee, A. Invasive alien plants occurring in Lesotho: Their ethnobotany potential risks distribution and origin. Bothalia Afr. Biodivers. Conserv. 2019, 49, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hyde, M.A.; Wursten, B.T.; Ballings, P.; Coates Palgrave, M. Flora of Zimbabwe: Species Information: Records of Pyracantha angustifolia. 2022. Available online: https://www.zimbabweflora.co.zw/speciesdata/species-display.php?species_id=125340 (accessed on 27 January 2022).
- Urcelay, C.; Longo, S.; Geml, J.; Tecco, P.A. Can arbuscular mycorrhizal fungi from non-invaded montane ecosystems facilitate the growth of alien trees? Mycorrhiza 2019, 29, 39–49. [Google Scholar] [CrossRef]
- Moreschi, E.G.; Funes, G.; Zeballos, S.R.; Tecco, P.A. Post-burning germination responses of woody invaders in a fire-prone ecosystem. Austral Ecol. 2019, 44, 1163–1173. [Google Scholar] [CrossRef]
- Guix, J.C. The role of alien plants in the composition of fruit-eating bird assemblages in Brazilian urban ecosystems. Orsis Org. I Sist. 2007, 22, 87–104. [Google Scholar]
- De Villalobos, A.E.; Vázquez, D.P.; Martin, J.L. Soil disturbance, vegetation cover and the establishment of the exotic shrub Pyracantha coccinea in southern France. Biol. Invasions 2010, 12, 1023–1029. [Google Scholar] [CrossRef]
- GBIF. Global Biodiversity Information Facility [WWW Document]. 2019. Available online: https://www.gbif.org/species/2988222 (accessed on 6 May 2019).
- Cadic, A. Pyracantha breeding program in France: First results. In III International Workshop on Fire Blight; ISHS: Bordeaux, France, 1983; Volume 151, pp. 307–314. [Google Scholar]
- Andreu, J.; Vilà, M. Native plant community response to alien plant invasion and removal. Manag. Biol. Invasions 2011, 2, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Brändle, M.; Brandl, R. The time lag between introduction and escape from cultivation of alien woody plant species decreases with polyploidization. Web Ecol. 2012, 12, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chari, L.D.; Martin, G.D.; Steenhuisen, S.L.; Adams, L.D.; Clark, V.R. Biology of Invasive Plants 1. Pyracantha angustifolia (Franch.) CK Schneid. Invasive Plant Sci. Manag. 2020, 13, 120–142. [Google Scholar] [CrossRef]
- Department of Environmental Affairs. National Environmental Management: Biodiversity Act 2004 (Act No. 10 of 2004) Alien and Invasive Species Regulations, 2014; Government Gazette Volume 590, No. 37885; Department of Environmental Affairs: Pretoria, South Africa, 2014.
- Henderson, L. Invasive alien woody plants of Natal and the north-eastern Orange Free State. Bothalia 1989, 19, 237–261. [Google Scholar] [CrossRef] [Green Version]
- Henderson, L. Invasive alien woody plants of the southern and southwestern Cape region, South Africa. Bothalia 1998, 28, 91–112. [Google Scholar] [CrossRef] [Green Version]
- Giorgis, M.A.; Tecco, P.A.; Cingolani, A.M.; Renison, D.; Marcora, P.; Paiaro, V. Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina. Biol. Invasions 2011, 13, 1423–1434. [Google Scholar] [CrossRef]
- Canavan, K.; Canavan, S.; Clark, V.R.; Gwate, O.; Richardson, D.M.; Sutton, G.F.; Martin, G.D. The Alien Plants That Threaten South Africa’s Mountain Ecosystems. Land 2021, 10, 1393. [Google Scholar] [CrossRef]
- Plenderleith, F.A.; Irrazabal, V.A.; Burslem, D.F.; Travis, J.M.; Powell, P.A. Predicting spatially heterogeneous invasive spread: Pyracantha angustifolia invading a dry Andean valley in northern Argentina. Biol. Invasions 2022, 24, 2201–2216. [Google Scholar] [CrossRef]
- Tecco, P.A.; Pais-Bosch, A.I.; Funes, G.; Marcora, P.I.; Zeballos, S.R.; Cabido, M.; Urcelay, C. Mountain invasions on the way: Are there climatic constraints for the expansion of alien woody species along an elevation gradient in Argentina? J. Plant Ecol. 2016, 9, 380–392. [Google Scholar] [CrossRef] [Green Version]
- Adams, L.D.; Martin, G.D.; Downs, C.T.; Clark, V.R.; Thabethe, V.; Raji, I.A.; Steenhuisen, S.L. Seed dispersal by frugivores and germination of the invasive alien shrub Pyracantha angustifolia (Franch.) CK Schneid. in Free State Province, South Africa. Biol. Invasions 2022, 24, 2809–2819. [Google Scholar] [CrossRef]
- Giantomasi, A.; Tecco, P.A.; Funes, G.; Gurvich, D.E.; Cabido, M. Canopy effects of the invasive shrub Pyracantha angustifolia on seed bank composition, richness and density in a montane shrubland (Córdoba, Argentina). Austral Ecol. 2008, 33, 68–77. [Google Scholar] [CrossRef]
- Tecco, P.A.; Diaz, S.; Gurvich, D.E.; Perez-Harguindeguy, N.; Cabido, M.; Bertone, G.A. Facilitation and interference underlying the association between the woody invaders Pyracantha angustifolia and Ligustrum lucidum. Appl. Veg. Sci. 2007, 10, 211–218. [Google Scholar] [CrossRef]
- Tapella, M.P.; Marcora, P.I.; Tecco, P.A. Reciprocal interactions between a non-native shrub and the dominant native trees of a high mountain woodland: Who benefits? Biol. Invasions 2020, 23, 53–67. [Google Scholar] [CrossRef]
- Semenya, S.S.; Tshisikhawe, M.P.; Potgieter, M.T. Invasive alien plant species: A case study of their use in the Thulamela Local Municipality, Limpopo Province, South Africa. Sci. Res. Essays 2012, 7, 2363–2369. [Google Scholar]
- Vergara-Tabares, D.L.; Badini, J.; Peluc, S.I. Fruiting phenology as a “triggering attribute” of invasion process: Do invasive species take advantage of seed dispersal service provided by native birds? Biol. Invasions 2016, 18, 677–687. [Google Scholar] [CrossRef]
- Vergara-Tabares, D.L.; Toledo, M.; García, E.; Peluc, S.I. Aliens will provide: Avian responses to a new temporal resource offered by ornithocorous exotic shrubs. Oecologia 2018, 188, 173–182. [Google Scholar] [CrossRef]
- Vergara-Tabares, D.L.; Blendinger, P.G.; Tello, A.; Peluc, S.I.; Tecco, P.A. Fleshy-fruited invasive shrubs indirectly increase native tree seed dispersal. Oikos 2022, 2022. [Google Scholar] [CrossRef]
- Williams, P.A.; Karl, B.J.; Bannister, P.; Lee, W.G. Small mammals as potential seed dispersers in New Zealand. Austral Ecol. 2000, 25, 523–532. [Google Scholar] [CrossRef]
- Veldtman, R. Are managed pollinators ultimately linked to the pollination ecosystem service paradigm? S. Afr. J. Sci. 2018, 114, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Stout, J.C.; Tiedeken, E.J. Direct interactions between invasive plants and native pollinators: Evidence, impacts and approaches. Funct. Ecol. 2017, 31, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Padrón, B.; Bartomeus, I.; Traveset, A. Consequences of plant invasions on compartmentalization and species’ roles in plant–pollinator networks. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140773. [Google Scholar] [CrossRef] [Green Version]
- Frost, C.M.; Allen, W.J.; Courchamp, F.; Jeschke, J.M.; Saul, W.C.; Wardle, D.A. Using network theory to understand and predict biological invasions. Trends Ecol. Evol. 2019, 34, 831–843. [Google Scholar] [CrossRef]
- Hickman, J.C. (Ed.) The Jepson Manual: Higher Plants of California; 3rd Printing with Corrections; University of California Press: Berkeley, CA, USA, 1993; 972p. [Google Scholar]
- Gioria, M.; Pyšek, P. Early bird catches the worm: Germination as a critical step in plant invasion. Biol. Invasions 2017, 19, 1055–1080. [Google Scholar] [CrossRef] [Green Version]
- Aslan, C.; Rejmanek, M. Native fruit traits may mediate dispersal competition between native and non-native plants. NeoBiota 2012, 12, 1–24. [Google Scholar] [CrossRef]
- Klinerová, T.; Tasevová, K.; Dostál, P. Large generative and vegetative reproduction independently increases global success of perennial plants from Central Europe. J. Biogeogr. 2018, 45, 1550–1559. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Heger, T. (Eds.) Invasion Biology: Hypotheses and Evidence; CABI: Wallingford, UK, 2018. [Google Scholar]
- Milton, S.J.; Wilson, J.R.U.; Richardson, D.M.; Seymour, C.L.; Dean, W.R.J.; Iponga, D.M.; Procheş, Ş. Invasive alien plants infiltrate bird-mediated shrub nucleation processes in arid savanna. J. Ecol. 2007, 95, 648–661. [Google Scholar] [CrossRef]
- White, E.; Vivian-Smith, G. Contagious dispersal of seeds of synchronously fruiting species beneath invasive and native fleshy-fruited trees. Austral Ecol. 2011, 36, 195–202. [Google Scholar] [CrossRef]
- Wilson, B.G.; Witkowski, E.T.F. Seed banks, bark thickness and change in age and size structure (1978–1999) of the African savanna tree, Burkea africana. Plant Ecol. 2003, 167, 151–162. [Google Scholar] [CrossRef]
- Gioria, M.; Carta, A.; Baskin, C.C.; Dawson, W.; Essl, F.; Kreft, H.; Pergl, J.; van Kleunen, M.; Weigelt, P.; Winter, M.; et al. Persistent soil seed banks promote naturalisation and invasiveness in flowering plants. Ecol. Lett. 2021, 24, 1655–1667. [Google Scholar] [CrossRef] [PubMed]
- Marks, P.L. On the origin of the field plants of the northeastern United States. Am. Nat. 1983, 122, 210–228. [Google Scholar] [CrossRef]
- Downey, P.O.; Richardson, D.M. Alien plant invasions and native plant extinctions: A six-threshold framework. AoB Plants 2016, 8, plw047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucina, L.; Rutherford, M.C. The Vegetation of South Africa, Lesotho and Swaziland; South African National Biodiversity Institute: Pretoria, South Africa, 2006.
- South African Environmental Observation Network. South African Risk and Vulnerability Atlas [WWW Document]. 2022. Available online: http://sarva2.dirisa.org/atlas/weather-and-climate (accessed on 6 May 2022).
- Martin, G.D. Prospects for the biological control of Northern Temperate Weeds in South Africa. Afr. Entomol. 2021, 29, 791–808. [Google Scholar] [CrossRef]
- eFlora. Pyracantha angustifolia (Franchet) C. K. Schneider. Flora China 2021, 9. Available online: http://www.efloras.org (accessed on 28 January 2021).
- Egolf, D.R.; Andrick, A.O. A Checklist of Pyracantha Cultivars. In U.S. National Arboretum Contribution; U.S. Department of Agriculture—Agricultural Research Service: Beltsville, MD, USA, 1995; 91p. [Google Scholar]
- Motulsky, H.J. Analyzing Data with GraphPad Prism 1999; GraphPad Software Inc.: San Diego, CA, USA, 1999. [Google Scholar]
- Beattie, A.J. A technique for the study of insect-borne pollen. Pan-Pac. Entomol. 1971, 47, 82. [Google Scholar]
- Geerts, S.; Botha, P.W.; Visser, V.; Richardson, D.M.; Wilson, J.R.U. Montpellier broom (Genista monspessulana) and Spanish broom (Spartium junceum) in South Africa: An assessment of invasiveness and options for management. S. Afr. J. Bot. 2013, 87, 134–145. [Google Scholar] [CrossRef] [Green Version]
- Holmes, P.M. Depth distribution and composition of seed-banks in alien-invaded and uninvaded fynbos vegetation. Austral Ecol. 2002, 27, 110–120. [Google Scholar] [CrossRef]
- International Seed Testing Association (ISTA). Seed testing procedure. Seed Sci. Technol. 1993, 21, 106–120. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seed Density (Million Seeds/m2) | Mean Shrub Radius (m) | Notes | |
|---|---|---|---|
| River | 3.022 | 2.63 | High seed number. Larger shrubs. |
| Open | 1.738 | 2.23 | Seeds more dispersed, large and small shrubs. |
| Rocky | 1.653 | 1.78 | Seeds more dispersed. Smaller shrubs. |
| P. angustifolia (n = 44) | L. sericea (n = 2) | R. rubiginosa (n = 6) | Fence (n = 1) | F Value | p-Value | |
|---|---|---|---|---|---|---|
| Total seed rain | 56,766 ± 7735 | 71 ± 55 | 2310 ± 2244 | 7 | 6.1 | 0.0054 |
| Seed rain from whole fruits | 55,395 ± 7490 | 61 ± 61 | 2248 ± 3195 | 0 | 12.8 | 0.0001 |
| Loose seed rain | 1371 ± 331 | 10 ± 6 | 62 ± 50 | 7 | 12.5 | 0.0329 |
| Seed Survival (%) | |||
|---|---|---|---|
| Months in the Soil | 1 | 3 | 6 |
| Site 1 | 98.0 ± 0.0 | 94.3 ± 0.7 | 0 |
| Site 2 | 98.0 ± 0.0 | 90.6 ± 1.4 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adams, L.D.; Giovannoni, D.; Clark, V.R.; Steenhuisen, S.-L.; Martin, G.D. Reproductive Ecology of the Invasive Alien Shrub Pyracantha angustifolia in the Grassland Biome, South Africa. Plants 2023, 12, 1308. https://doi.org/10.3390/plants12061308
Adams LD, Giovannoni D, Clark VR, Steenhuisen S-L, Martin GD. Reproductive Ecology of the Invasive Alien Shrub Pyracantha angustifolia in the Grassland Biome, South Africa. Plants. 2023; 12(6):1308. https://doi.org/10.3390/plants12061308
Chicago/Turabian StyleAdams, Lehlohonolo D., Dino Giovannoni, Vincent R. Clark, Sandy-Lynn Steenhuisen, and Grant D. Martin. 2023. "Reproductive Ecology of the Invasive Alien Shrub Pyracantha angustifolia in the Grassland Biome, South Africa" Plants 12, no. 6: 1308. https://doi.org/10.3390/plants12061308