
Population Growth of Fall Armyworm, Spodoptera frugiperda Fed on Cereal and Pulse Host Plants Cultivated in Yunnan Province, China

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Development Time and Adult Longevity of S. frugiperda
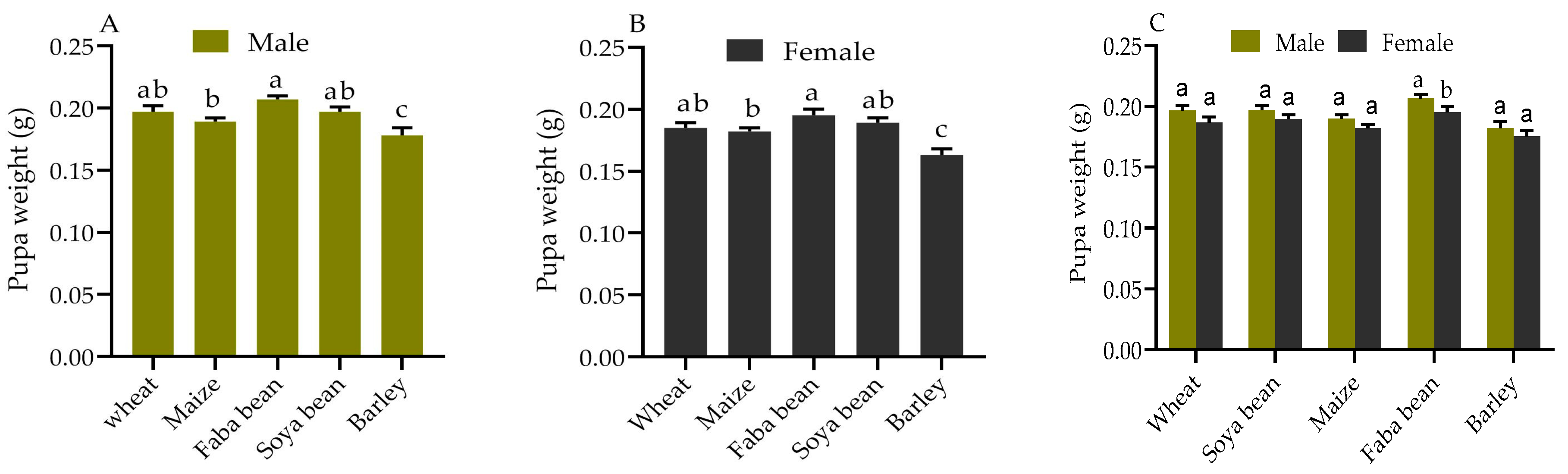
2.2. Pupae Weight
2.3. Population Parameters of S. frugiperda
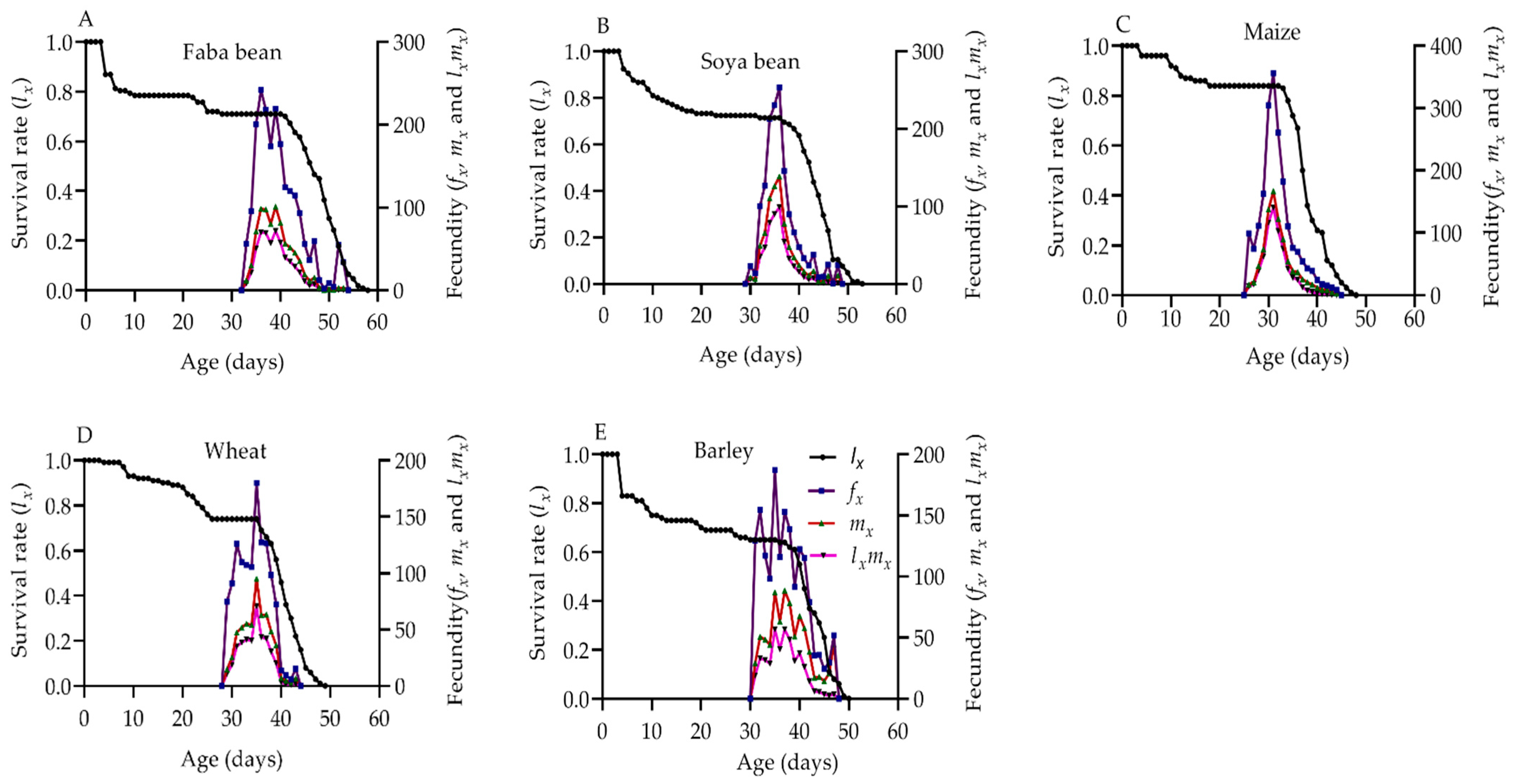
2.4. Reproduction Parameters of S. frugiperda
2.5. Population Parameters of S. frugiperda
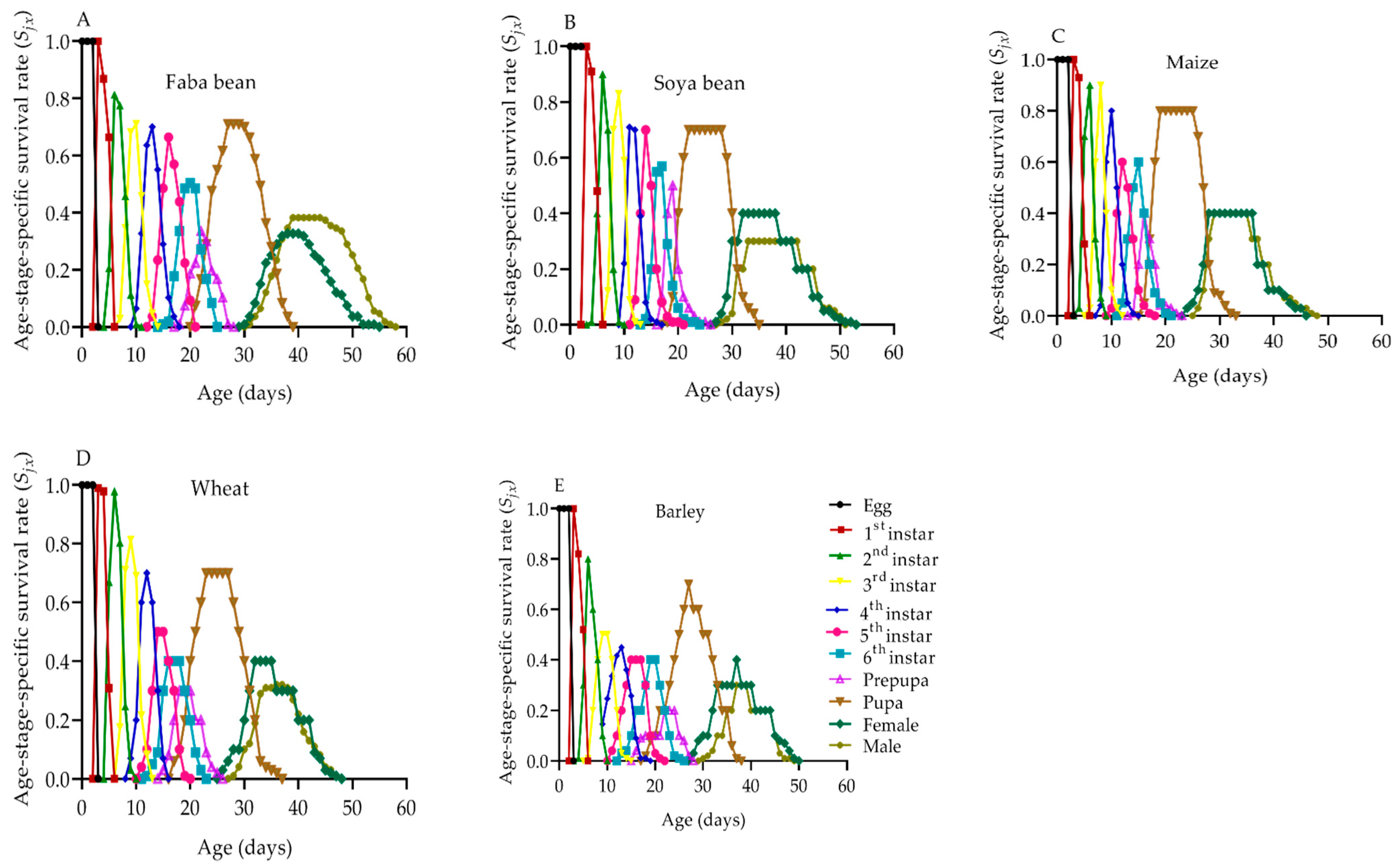
2.6. Population Survival Rate and Fecundity of S. frugiperda
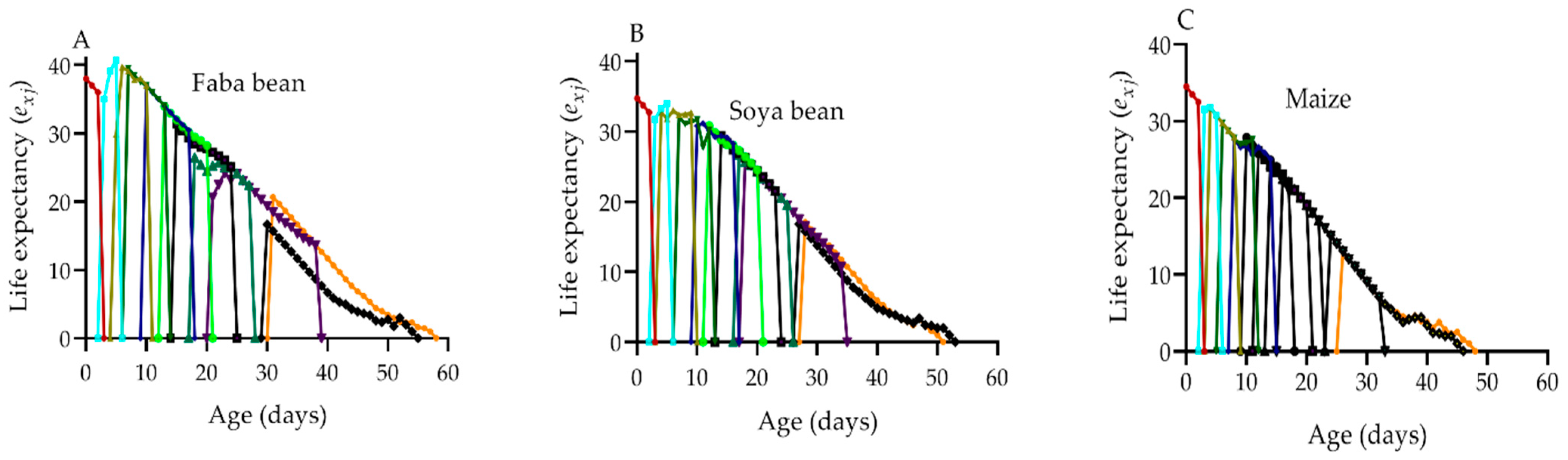
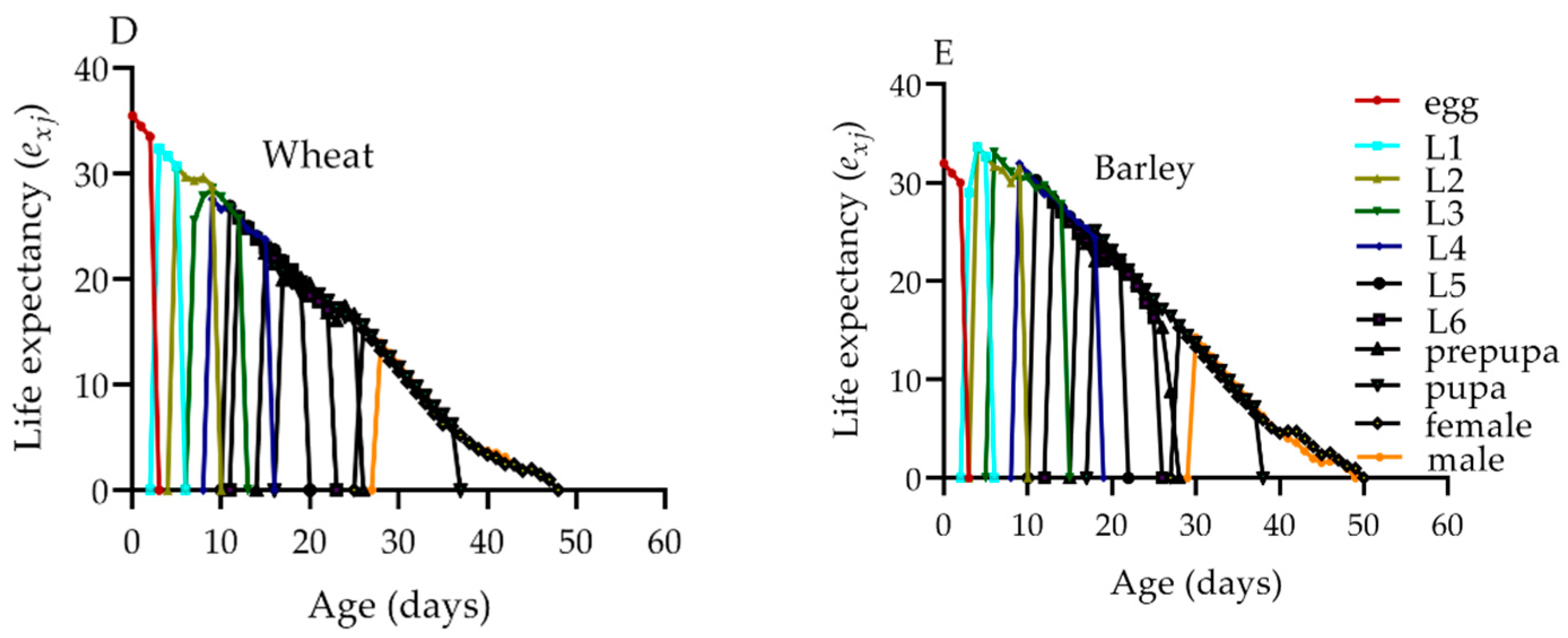
2.7. Life Expectancy of S. frugiperda
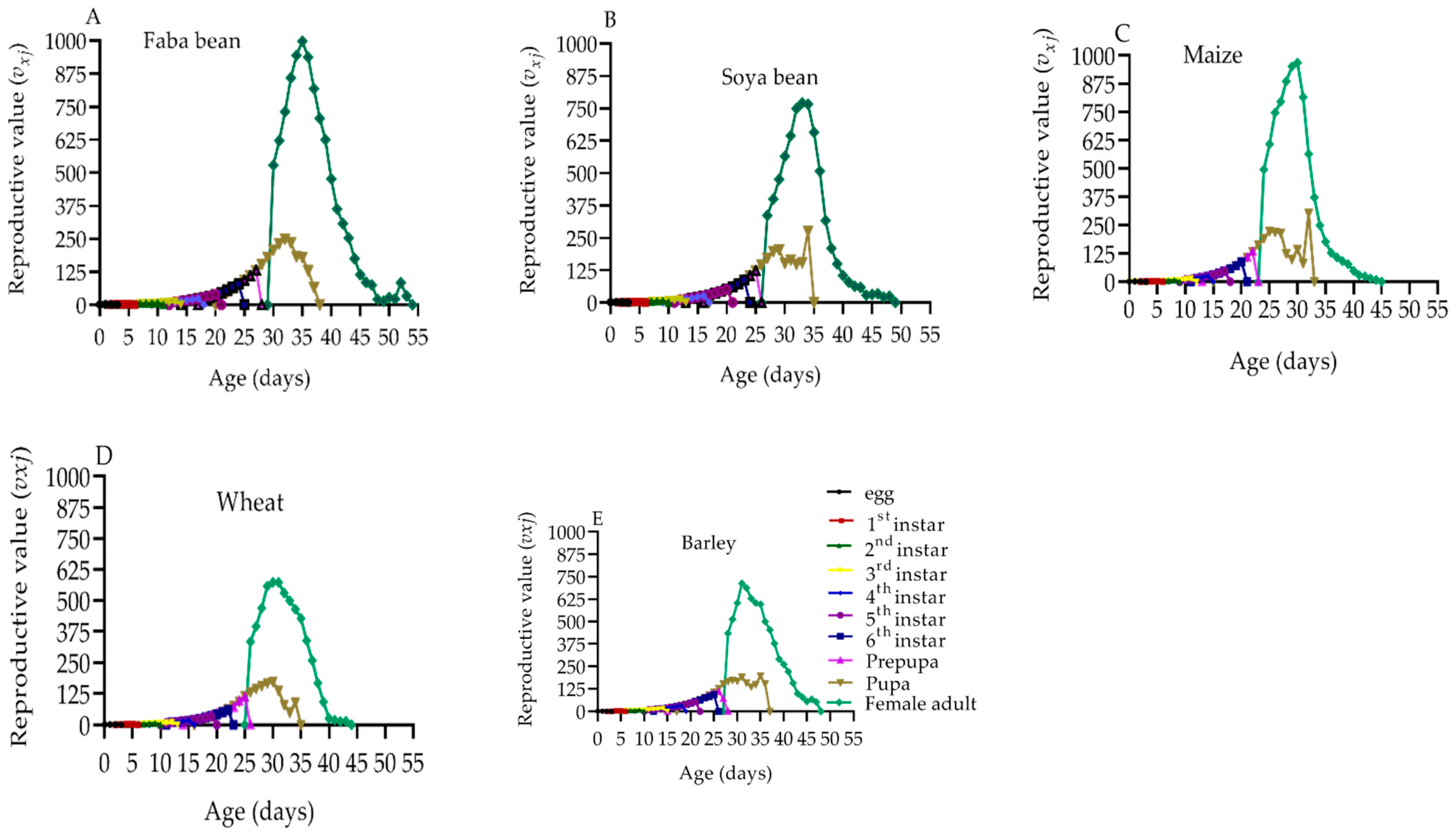
2.8. Reproduction Value of S. frugiperda
3. Discussion
4. Materials and Methods
4.1. Insects and Host Plants
4.2. Population Growth and Life Table of S. frugiperda
4.3. Life Table Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sparks, A.N. A Review of the Biology of the Fall Armyworm. Fla. Entomol. 1979, 62, 82. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Fleischer, S.; Meagher, R.L.; Hay-Roe, M.; Khan, A.; Murúa, M.G.; Silvie, P.; Vergara, C.; Westbrook, J. Fall armyworm migration across the Lesser Antilles and the potential for genetic exchanges between North and South American populations. PLoS ONE 2017, 12, e0171743. [Google Scholar]
- Jin, M.H.; Tao, J.H.; Qi, L.; Cheng, Y.; Sun, X.X.; Wu, K.M.; Xiao, Y.T. Genome editing of the SfABCC2 gene confers resistance to Cry1F toxin from Bacillus thuringiensis in Spodoptera frugiperda. J. Integr. Agric. 2021, 20, 815–820. [Google Scholar] [CrossRef]
- Yang, X.M.; Song, Y.F.; Sun, X.X.; Shen, X.J.; Wu, Q.L.; Zhang, H.W.; Zhang, D.M.; Zhao, S.Y.; Liang, G.M.; Wu, K.M. Population occurrence of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in the winter season of China. J. Integr. Agric. 2021, 20, 772–782. [Google Scholar] [CrossRef]
- Silva, D.M.D.; Bueno, A.D.F.; Andrade, K.; Stecca, C.D.S.; Neves, P.M.O.J.; Oliveira, M.S.N. Biology and nutrition of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on different food sources. Sci. Agric. 2017, 74, 18–31. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Zhi, J.; Ye, J.; Zhou, Y.; Li, C.; Liang, Y.; Yue, W.; Li, D.; Zeng, G.; Hu, C. Age-stage, two-sex life table analysis of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) reared on maize and kidney bean. Chem. Biol. Technol. Agric. 2021, 8, 1–8. [Google Scholar] [CrossRef]
- Deshmukh, S.S.; Prasanna, B.; Kalleshwaraswamy, C.; Jaba, J.; Choudhary, B. Fall Armyworm (Spodoptera frugiperda) Polyphagous Pests of Crops; Springer: Berlin/Heidelberg, Germany, 2021; pp. 349–372. [Google Scholar]
- Wu, M.F.; Qi, G.J.; Chen, H.; Ma, J.; Liu, J.; Jiang, Y.Y.; Lee, G.S.; Otuka, A.; Hu, G. Overseas immigration of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), invading Korea and Japan in 2019. Insect Sci. 2022, 29, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Huang, C.; Li, C.Y.; Zhou, H.X.; Ren, Y.I.; Li, Z.Y.; Xing, L.S.; Zhang, B.; Xi, Q.; Bo, L. Biology, invasion and management of the agricultural invader: Fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Integr. Agric. 2021, 20, 646–663. [Google Scholar]
- Lu, H.; Tang, J.; Lyu, B.; Ma, Z.; He, X.; Chen, Q.; Su, H. Recent advances in biological control and invasion risk of Spodoptera frugiperda. Chin. J. Trop. Crops 2019, 40, 1237. [Google Scholar]
- Montezano, D.G.; Sosa-Gómez, D.; Specht, A.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.D.; Peterson, J.A.; Hunt, T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.I.; Shen, X.I.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm, Spodoptera frugiperda invading into China. J. Integr. Agric. 2021, 20, 664–672. [Google Scholar] [CrossRef]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.A.; Day, R.; Desneux, N.; Harrison, R.D.; Kriticos, D.; Rwomushana, I.; van den Berg, J. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen. 2022, 1–55. [Google Scholar] [CrossRef]
- Prasanna, B.; Huesing, J.; Eddy, R.; Peschke, V. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; CIMMYT: Ciudad de México, Mexico, 2018. [Google Scholar]
- Huang, Y.; Dong, Y.; Huang, W.; Ren, B.; Deng, Q.; Shi, Y.; Bai, J.; Ren, Y.; Geng, Y.; Ma, H. Overwintering distribution of fall armyworm (Spodoptera frugiperda) in Yunnan, China, and influencing environmental factors. Insects 2020, 11, 805. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Liu, J.; Xie, M.; Li, Y.; Yang, J.; Zhang, M.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Prot. 2019, 45, 10–19. [Google Scholar]
- Chen, Z.W.; Yang, Y.C.; Zhang, J.F.; Jin, M.H.; Liu, K.Y. Susceptibility and tissue specificity of Spodoptera frugiperda to Junonia coenia densovirus. J. Integr. Agric. 2021, 20, 840–849. [Google Scholar] [CrossRef]
- Yang, X.; Sun, X.; Zhao, S.; Li, J.; Chi, X.; Jiang, Y.; Wu, K. Population occurrence, spatial distribution and sampling technique of fall armyworm Spodoptera frugiperda in wheat fields. Plant Prot. 2020, 46, 10–16. [Google Scholar]
- Jiang, W.; Cheng, Q.; Lu, C.; Chen, W.; Zhao, D.; He, Y. Different host plants distinctly influence the adaptability of Myzus persicae (Hemiptera: Aphididae). Agriculture 2022, 12, 2162. [Google Scholar] [CrossRef]
- Wang, W.; He, P.; Zhang, Y.; Liu, T.; Jing, X.; Zhang, S. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. Insects 2020, 11, 639. [Google Scholar] [CrossRef]
- Ali, M.Y.; Naseem, T.; Arshad, M.; Ashraf, I.; Rizwan, M.; Tahir, M.; Rizwan, M.; Sayed, S.; Ullah, M.I.; Khan, R.R. Host-plant variations affect the biotic potential, survival, and population projection of Myzus persicae (Hemiptera: Aphididae). Insects 2021, 12, 375. [Google Scholar] [CrossRef]
- Chiriboga Morales, X.; Tamiru, A.; Sobhy, I.S.; Bruce, T.J.; Midega, C.A.; Khan, Z. Evaluation of African maize cultivars for resistance to fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) larvae. Plants 2021, 10, 392. [Google Scholar] [CrossRef]
- Saeed, S.; Sayyed, A.H.; Ahmad, I. Effect of host plants on life-history traits of Spodoptera exigua (Lepidoptera: Noctuidae). J. Pest Sci. 2010, 83, 165–172. [Google Scholar] [CrossRef]
- Altaf, N.; Idrees, A.; Ullah, M.I.; Arshad, M.; Afzal, A.; Afzal, M.; Rizwan, M.; Li, J. Biotic potential induced by different host plants in the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 921. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Jiang, X.; Ling, Y.; Jiang, T.; Chen, Y.; Long, D.; Long, L. A comparative study on growth, development and reproduction of Mythimna separata in four host plants. China Plant Prot. 2018, 38, 5–10. [Google Scholar]
- Zhou, J.; Zhang, J.; Tregenza, T.; Pan, Y.; Wang, Q.; Shi, H.; Liu, X. Larval host preference and suitability for the sawfly Mesoneura rufonota among five cinnamomun tree species. Insects 2020, 11, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, C.; Auad, A.; Mendes, S.; Frizzas, M. Crop losses and the economic impact of insect pests on Brazilian agriculture. Crop Prot. 2014, 56, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Wu, F.; Fan, J.; Zhang, R. Potential economic impact of invasive fall armyworm on mainly affected crops in China. J. Pest Sci. 2021, 94, 1065–1073. [Google Scholar] [CrossRef]
- Acharya, R.; Malekera, M.J.; Dhungana, S.K.; Sharma, S.R.; Lee, K.Y. Impact of rice and potato Host plants is higher on the reproduction than growth of corn strain fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 256. [Google Scholar] [CrossRef]
- Emden, V.H. Mechanisms of resistance: Antibiosis, antixenosis, tolerance, nutrition. In Encyclopedia of Pest Management; Marcel Dekker, Inc.: New York, NY, USA, 2002; pp. 483–600. [Google Scholar]
- Baldin, E.L.; Stamm, M.D.; Bentivenha, J.P.; Koch, K.G.; Heng-Moss, T.M.; Hunt, T.E. Feeding behavior of Aphis glycines (Hemiptera: Aphididae) on soybeans exhibiting antibiosis, antixenosis, and tolerance resistance. Fla. Entomol. 2018, 101, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Cheng, Y.; Wang, Q.; Liu, X.; Fu, Y.; Zhang, Y.; Chen, J. Push–pull plants in wheat intercropping system to manage Spodoptera frugiperda. J. Pest Sci. 2022, 3, 1–15. [Google Scholar] [CrossRef]
- Carvalho, I.F.; Machado, L.L.; Neitzke, C.G.; Erdmann, L.L.; Oliveira, L.T.; Bernardi, D.; da Rosa, A.P.S.A. Biological parameters and fertility life table of Spodoptera frugiperda in different host plants. J. Agric. Sci. 2022, 14, 10. [Google Scholar] [CrossRef]
- Gopalakrishnan, R.; Kalia, V.K. Biology and biometric characteristics of Spodoptera frugiperda (Lepidoptera: Noctuidae) reared on different host plants with regard to diet. Pest Manag. Sci. 2022, 78, 2043–2051. [Google Scholar] [CrossRef] [PubMed]
- Boregas, K.G.B.; Mendes, S.M.; Waquil, J.M.; Fernandes, G.W. Estádio de adaptação de Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) em hospedeiros alternativos. Bragantia 2013, 72, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Mason, C.J.; Ray, S.; Davidson-Lowe, E.; Ali, J.G.; Luthe, D.S.; Felton, G. Plant nutrition influences resistant Maize Defense Responses to the Fall Armyworm (Spodoptera frugiperda). Front. Ecol. Evol. 2022, 10, 844274. [Google Scholar] [CrossRef]
- Gebretsadik, K.G.; Zhang, Y.; Chen, J. Screening and evaluation for antibiosis resistance of the spring wheat accessions to the grain aphid, Sitobion miscanthi (Takahashi) (Hemiptera: Aphididae). J. Integr. Agric. 2022, 21, 2329–2344. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, J.; Gao, Z.; He, K.; Bai, S.; Zhang, T.; Wang, Z. Performance of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on six host plants: Potential risks to mid-high latitude crops in China. J. Agric. Sci. 2020, 12, 16. [Google Scholar] [CrossRef]
- Pascacio-Villafán, C.; Williams, T.; Birke, A.; Aluja, M. Nutritional and non-nutritional food components modulate phenotypic variation but not physiological trade-offs in an insect. Sci. Rep. 2016, 6, 29413. [Google Scholar] [CrossRef] [Green Version]
- Tayal, M.; Somavat, P.; Rodriguez, I.; Thomas, T.; Christoffersen, B.; Kariyat, R. Polyphenol-rich purple corn pericarp extract adversely impacts herbivore growth and development. Insects 2020, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- He, L.M.; Wu, Q.L.; Gao, X.W.; Wu, K.M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Ashok, K.; Balasubramani, V.; Kennedy, J.S.; Geethalakshmi, V.; Jeyakumar, P.; Sathiah, N. Evaluating artificial diets for the fall armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) through nutritional indices and an age-stage, two-sex life table approach. Afr. Entomol. 2021, 29, 620–634. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2015. Available online: http://140.120.197.173/Ecology/Download/00-How%20to%20use%20TWOSEX.pdf (accessed on 2 December 2022).
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H.; You, M.; Atlihan, R.; Smith, C.L.; Kavousi, A.; Ozgokce, M.S.; Guncan, A.; Tuan, S.J.; Fu, J.W.; Xu, Y.Y. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Gen. 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Goodman, D. Optimal life histories, optimal notation, and the value of reproductive value. Am. Nat. 1982, 119, 803–823. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Huang, H.W.; Chi, H.; Smith, C.L. Linking demography and consumption of Henosepilachna vigintioctopunctata (Coleoptera: Coccinellidae) fed on Solanum photeinocarpum (Solanales: Solanaceae): With a new method to project the uncertainty of population growth and consumption. J. Econ. Entomol. 2018, 111, 1–9. [Google Scholar] [CrossRef]
- Akkopru, E.P.; Atlıhan, R.; Okut, H.; Chi, H. Demographic assessment of plant cultivar resistance to insect pests: A case study of the dusky-veined walnut aphid (Hemiptera: Callaphididae) on five walnut cultivars. J. Econ. Entomol. 2015, 108, 378–387. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Development Stage, d | Host Plants | |||||
|---|---|---|---|---|---|---|
| Maize | Faba Beans | Soya Beans | Wheat | Barley | p-Value | |
| Egg | 3.00 ± 0.00 | 3.00 ± 0.00 | 3.00 ± 0.00 | 3.31 ± 0.31 | 3.00 ± 0.0 | NS |
| 1st instar | 2.26 ± 0.05 c | 2.76 ± 0.04 a | 2.52 ± 0.05 b | 2.32 ± 0.05 c | 2.60 ± 0.06 b | <0.0001 |
| 2nd instar | 2.11 ± 0.04 c | 2.87 ± 0.08 a | 2.53 ± 0.06 b | 2.79 ± 0.05 a | 2.79 ± 0.68 a | <0.0001 |
| 3rd instar | 2.15 ± 0.05 c | 3.07 ± 0.08 a | 2.81 ± 0.05 b | 2.86 ± 0.05 b | 2.89 ± 0.77 ab | <0.0001 |
| 4th instar | 2.25 ± 0.05 d | 3.45 ± 0.06 a | 2.70 ± 0.07 c | 2.83 ± 0.07 c | 3.14 ± 0.63 b | <0.0001 |
| 5th instar | 2.41 ± 0.07 d | 3.50 ± 0.07 a | 2.65 ± 0.07 c | 2.54 ± 0.07 cd | 3.17 ± 0.60 b | <0.0001 |
| 6th instar | 2.54 ± 0.07 b | 3.24 ± 0.08 a | 2.50 ± 0.06 b | 2.63 ± 0.07 b | 3.24 ± 0.11 a | <0.0001 |
| 1st to 6th instar | 13.72 ± 0.18 d | 18.94 ± 0.19 a | 15.64 ± 0.15 c | 15.93 ± 0.21 c | 17.86 ± 0.31 b | <0.0001 |
| Pre-pupa | 1.40 ± 0.06 c | 2.05 ± 0.04 b | 1.97 ± 0.04 b | 1.97 ± 0.05 b | 2.27 ± 0.63 a | <0.0001 |
| Pupa | 9.69 ± 0.11 c | 10.75 ± 0.12 a | 10.32 ± 0.10 b | 10.37 ± 0.14 b | 9.95 ± 0.89 c | <0.0001 |
| Pre-adult | 27.83 ± 0.22 d | 34.79 ± 0.26 a | 30.92 ± 0.20 c | 30.97 ± 0.28 c | 33.05 ± 0.35 b | <0.0001 |
| Female adult | 11.94 ± 0.42 bc | 12.86 ± 0.45 ab | 13.51 ± 0.47 a | 11.21 ± 0.39 c | 11.15 ± 0.46 c | 0.0002 |
| Male adult | 10.58 ± 0.54 c | 16.07 ± 0.42 a | 13.66 ± 0.47 b | 9.94 ± 0.45 c | 10.04 ± 0.63 c | <0.0001 |
All Adult ( + +  ) ) | 11.26 ± 0.35 c | 14.59 ± 0.36 a | 13.58 ± 0.33 b | 10.62 ± 0.30 c | 10.59 ± 0.38 c | <0.0001 |
| Total longevity () | 39.13 ± 0.59 d | 46.69 ± 0.58 a | 43.78 ± 0.58 b | 41.26 ± 0.53 c | 43.35 ± 0.63 b | <0.0001 |
| Total longevity () | 39.00 ± 0.67 d | 51.68 ± 0.47 a | 45.34 ± 0.62 b | 41.97 ± 0.51 c | 44.29+0.49 b | <0.0001 |
| Total longevity ( + ) | 39.06 ± 0.44 d | 49.38 ± 0.47 a | 44.50 ± 0.43 b | 41.59 ± 0.37 c | 43.74 ± 0.42 b | <0.0001 |
= Male, = Female. The larval development stages, pupal, pre adult and adult durations, and total longevity were analyzed using age-stage, two-sex life table. Mean values M ± SE in the same row followed by different letters were significantly different (p < 0.05) (paired bootstrap test).| Survival Rate, % | Host Plants | ||||
|---|---|---|---|---|---|
| Maize | Faba Beans | Soya Beans | Wheat | Barley | |
| Egg | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| 1st instar | 96.05 | 86.91 | 90.47 | 98.97 | 83.14 |
| 2nd instar | 96.05 | 79.43 | 86.66 | 98.97 | 78.65 |
| 3rd instar | 94.74 | 78.50 | 80.00 | 92.78 | 74.15 |
| 4th instar | 88.16 | 78.50 | 77.14 | 90.72 | 73.03 |
| 5th instar | 86.86 | 78.50 | 75.24 | 89.70 | 73.03 |
| 6th instar | 85.53 | 78.50 | 74.29 | 88.66 | 70.78 |
| Prepupa | 85.53 | 78.50 | 73.33 | 84.53 | 67.42 |
| Pupa | 84.21 | 71.03 | 72.30 | 73.20 | 65.16 |
| Adult | 84.21 | 71.03 | 72.30 | 73.20 | 65.16 |
| Host Plants | Biological Parameters | ||||
|---|---|---|---|---|---|
| APOP (d) | TPOP (d) | Oviposition (d) | Fecundity | Female Ratio | |
| Maize | 2.84 ± 0.22 c | 30.02 ± 03 d | 6.16 ± 0.34 a | 1705.45 ± 125.84 a | 48.44% |
| Faba beans | 3.80 ± 0.37 ab | 37.63 ± 0.50 a | 6.09 ± 0.29 a | 1706.40 ± 123.19 a | 46.05% |
| Soya beans | 4.08 ± 0.24 a | 34.30 ± 0.34 bc | 5.61 ± 0.34 ab | 1315.59 ± 95.78 b | 53.95% |
| Wheat | 3.57 ± 0.20 abc | 33.54 ± 2.55 c | 4.58 ± 0.25 c | 1015.29 ± 89.98 c | 36.54% |
| Barley | 3.06 ± 0.17 bc | 35.26 ± 2.91 b | 4.97 ± 0.28 bc | 1160.74 ± 59.05 bc | 30.49% |
| p-value | <0.0102 | <0.0001 | <0.0007 | <0.0001 | |
| Host Plants | Population Parameters | |||
|---|---|---|---|---|
| Ro | r (d−1) | λ (d−1) | T (d) | |
| Maize | 695.64 ± 108.34 a | 0.205 ± 0.0065 a | 1.228 ± 0.0074 a | 31.877 ± 0.44 c |
| Faba beans | 558.17 ± 86.85 ab | 0.162 ± 0.0049 b | 1.176 ± 0.0057 b | 39.079 ± 0.45 a |
| Soya beans | 513.70 ± 72.83 ab | 0.173 ± 0.0046 b | 1.188 ± 0.0054 b | 36.167 ± 0.34 bc |
| Wheat | 443.43 ± 63.66 b | 0.172 ± 0.0054 b | 1.187 ± 0.0063 | 34.92 ± 0.47 c |
| Barley | 397.74 ± 61.11 b | 0.165 ± 0.0047 b | 1.179 ± 0.0053 b | 36.93 ± 0.52 b |
| p-value | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gebretsadik, K.G.; Liu, Y.; Yin, Y.; Zhao, X.; Li, X.; Chen, F.; Zhang, Y.; Chen, J.; Chen, A. Population Growth of Fall Armyworm, Spodoptera frugiperda Fed on Cereal and Pulse Host Plants Cultivated in Yunnan Province, China. Plants 2023, 12, 950. https://doi.org/10.3390/plants12040950
Gebretsadik KG, Liu Y, Yin Y, Zhao X, Li X, Chen F, Zhang Y, Chen J, Chen A. Population Growth of Fall Armyworm, Spodoptera frugiperda Fed on Cereal and Pulse Host Plants Cultivated in Yunnan Province, China. Plants. 2023; 12(4):950. https://doi.org/10.3390/plants12040950
Chicago/Turabian StyleGebretsadik, Kifle Gebreegziabiher, Ying Liu, Yanqiong Yin, Xueqing Zhao, Xiangyong Li, Fushou Chen, Yong Zhang, Julian Chen, and Aidong Chen. 2023. "Population Growth of Fall Armyworm, Spodoptera frugiperda Fed on Cereal and Pulse Host Plants Cultivated in Yunnan Province, China" Plants 12, no. 4: 950. https://doi.org/10.3390/plants12040950