
Phytochemical Profile and Antioxidant Capacity of Viscum album L. Subsp. album and Effects on Its Host Trees
 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Degree of Mistletoe Infestation in the Area under Study
2.2. Macroscopic and Microscopic Evaluation of Mistletoe Leaves
2.3. The Photosynthetic Pigments of Mistletoe and Its Host Leaves
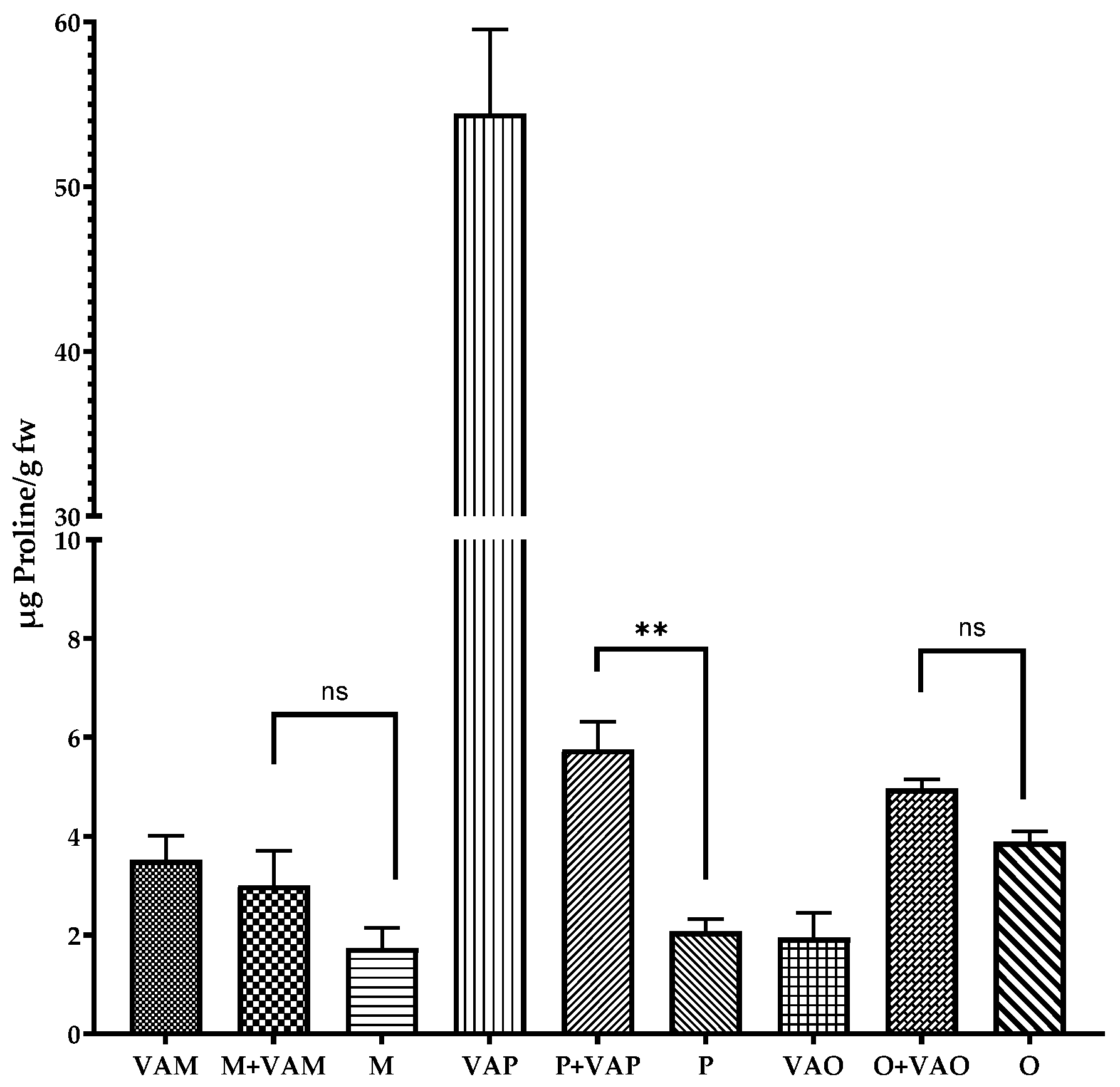
2.4. Proline Content of Leaves
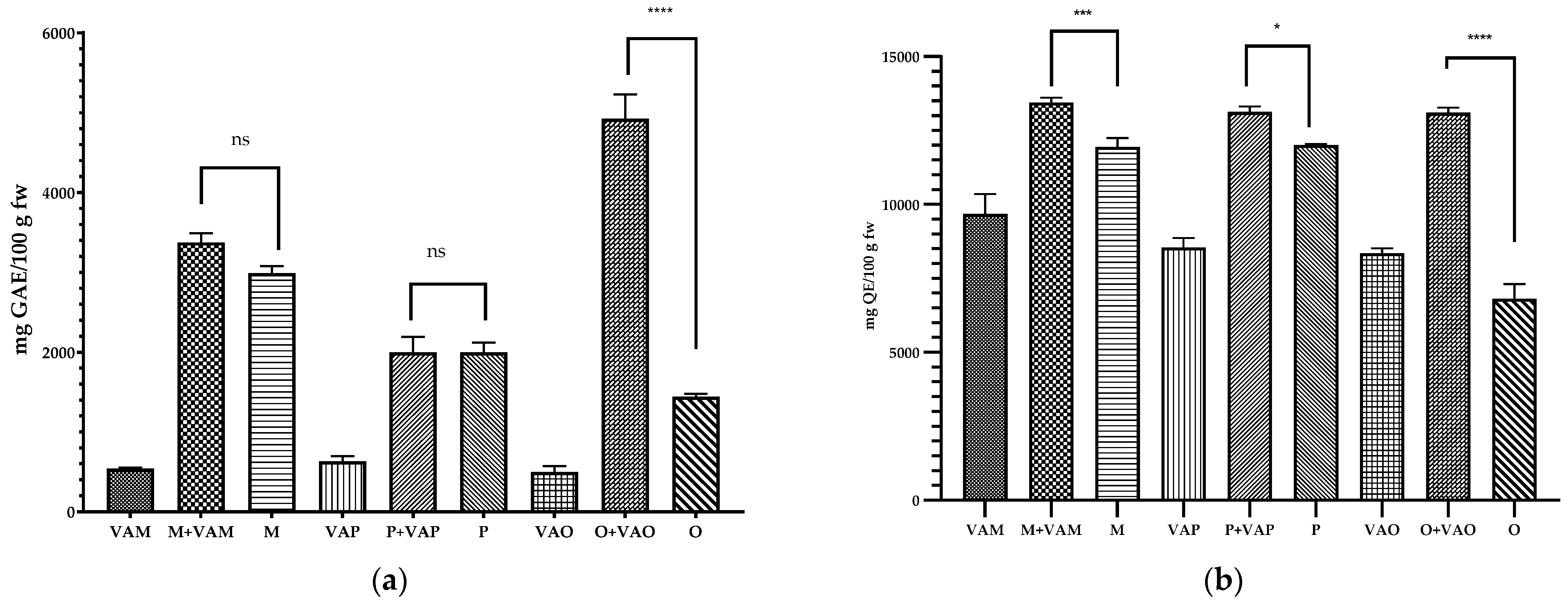
2.5. Phenols Content of Mistletoe Leaves and Host Tree Leaves
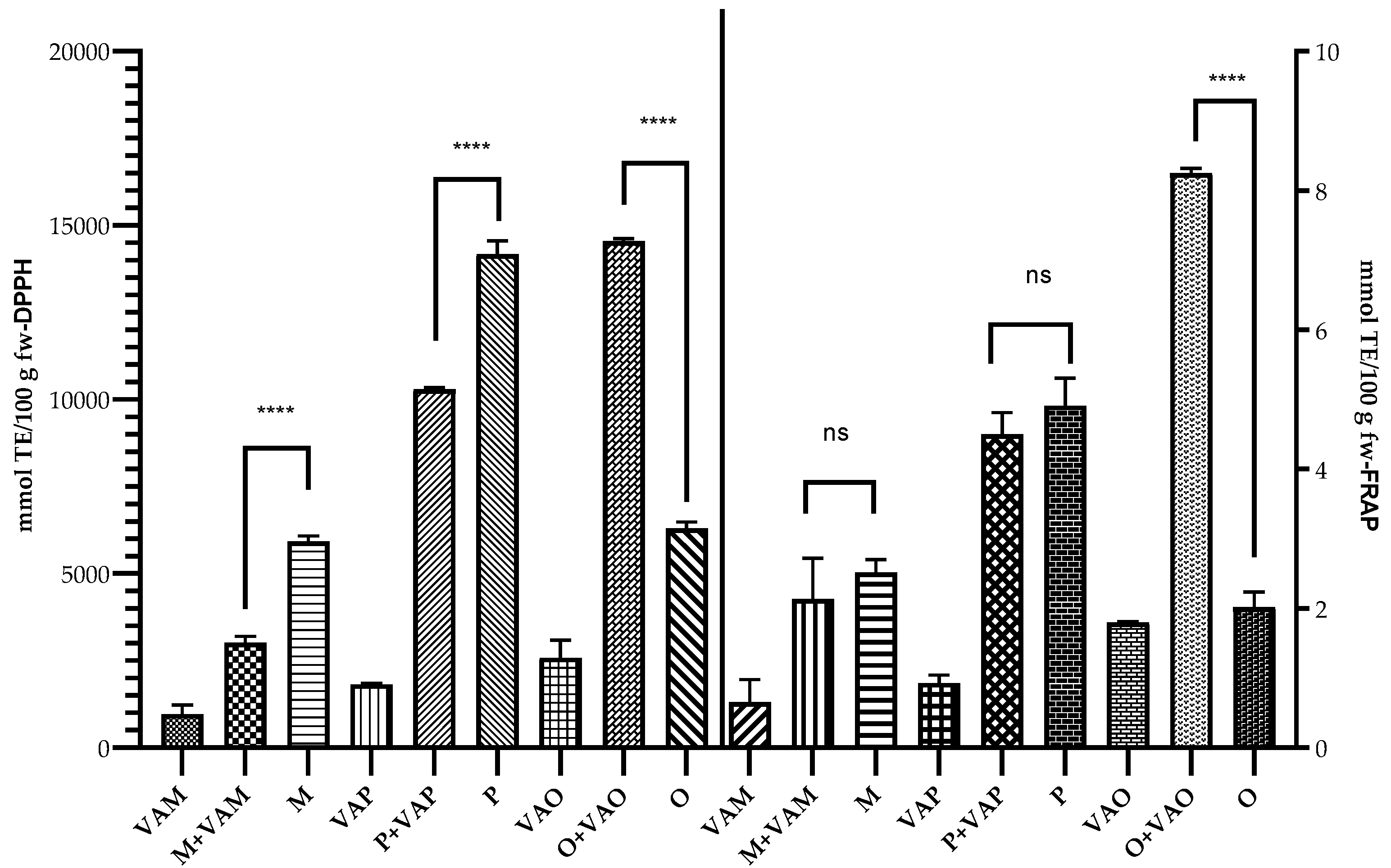
2.6. Antioxidant Capacity of Mistletoe Leaves and Their Host Trees
3. Materials and Methods
3.1. Biological Material
3.2. Determining the Degree of Mistletoe Infestation
3.3. Macroscopic and Microscopic Characterization of Mistletoe Leaves (V. album L. Subsp. Album)
3.4. Spectrophotometric Determination of Photosynthetic Pigments Content
3.5. HPLC-DAD-MS-ESI + Analysis Phenolic Compounds of Mistletoe Leaves
3.5.1. Sample Preparation
3.5.2. Chromatographic Condition
3.6. Spectrophotometric Determination of Total Phenols and Flavonoids Content
3.7. Proline Determination
3.8. Determination of Antioxidant Capacity
3.8.1. DPPH (2,2-Diphenyl-1-picryl-hydrazyl-hydrate) Assay
3.8.2. FRAP (Ferric-Reducing Antioxidant Power) Assay
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szmidla, H.; Tkaczyk, M.; Plewa, R.; Tarwacki, G.; Sierota, Z. Impact of Common Mistletoe (Viscum album L.) on Scots Pine Forests—A Call for Action. Forests 2019, 10, 847. [Google Scholar] [CrossRef] [Green Version]
- Stypiński, P.T. Biologia i Ekologia Jemioły Pospolitej (Viscum album, Viscaceae) w Polsce Biology and Ecology of the European Mistletoe (Viscum album, Viscaceae) in Poland; Fragmenta Floristica et Geobotanica; Instytut Botaniki im. W. Szafera, PAN: Kraków, Poland, 1997; ISBN 978-83-85444-50-3. [Google Scholar]
- Barney, C.; Hawksworth, F.; Geils, B. Hosts of Viscum album. Eur. J. For. Pathol. 2007, 28, 187–208. [Google Scholar] [CrossRef]
- Okubamichael, D.Y.; Griffiths, M.E.; Ward, D. Host Specificity in Parasitic Plants-Perspectives from Mistletoes. AoB Plants 2016, 8, plw069. [Google Scholar] [CrossRef] [PubMed]
- Kleszken, E.; Adrian, T.; Memete, A.; Miere, F.; Vicaş, S. On Overview of Bioactive Compounds, Biological and Pharmacological Effects of Mistletoe (Viscum album L). Pharmacophore 2022, 13, 10–26. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Erbilgin, N. Dwarf Mistletoe Infection in Jack Pine Alters Growth-Defense Relationships. Tree Physiol. 2018, 38, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Kleszken, E.; Laslo, V.; Vicas, S.I. Spectrophotometric Quantification of Green Pigments and Total Carotenoids from Mistletoe Grown on Different Host Trees. Nat. Resour. Sustain. Dev. 2019, 9, 27–33. [Google Scholar] [CrossRef]
- Muche, M.; Muasya, A.; Tsegay, B. Biology and Resource Acquisition of Mistletoes, and the Defense Responses of Host Plants. Ecol. Process. 2022, 11, 24. [Google Scholar] [CrossRef]
- Saucet, S.B.; Shirasu, K. Molecular Parasitic Plant–Host Interactions. PLoS Pathog. 2016, 12, e1005978. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, S.; Ilhan, V.; Turkoglu, H.I. Mistletoe (Viscum album) Infestation in the Scots Pine Stimulates Drought-Dependent Oxidative Damage in Summer. Tree Physiol. 2016, 36, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rigling, A.; Eilmann, B.; Koechli, R.; Dobbertin, M. Mistletoe-Induced Crown Degradation in Scots Pine in a Xeric Environment. Tree Physiol. 2010, 30, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Barbu, C. The Incidence and Distribution of White Mistletoe (Viscum album ssp. Abietis) on Silver Fir (Abies alba Mill.) Stands from Eastern Carpathians. Ann. For. Res. 2010, 53, 27–36. [Google Scholar]
- Catal, Y.; Carus, S. Effect of Pine Mistletoe on Radial Growth of Crimean Pine (Pinus nigra) in Turkey. J. Environ. Biol. 2011, 32, 263–270. [Google Scholar]
- Zweifel, R.; Bangerter, S.; Rigling, A.; Sterck, F.J. Pine and Mistletoes: How to Live with a Leak in the Water Flow and Storage System? J. Exp. Bot. 2012, 63, 2565–2578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilichowski, S.; Filip, R.; Kościelska, A.; Żaroffe, G.; Żyźniewska, A.; Iszkuło, G. Influence of Viscum album ssp. Austriacum (Wiesb.) Vollm. on Tree Radial Growth of Pinus sylvestris L. Wpływ Viscum album Ssp. Austriacum (Wiesb.) Vollm. Na Przyrost Radialny Pinus sylvestris L. Sylwan 2018, 162, 452–459. [Google Scholar]
- Sangüesa-Barreda, G.; Linares, J.C.; Julio Camarero, J. Drought and Mistletoe Reduce Growth and Water-Use Efficiency of Scots Pine. For. Ecol. Manag. 2013, 296, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Tsopelas, P.; Angelopoulos, A.; Economou, A.; Soulioti, N. Mistletoe (Viscum album) in the Fir Forest of Mount Parnis, Greece. For. Ecol. Manag. 2004, 202, 59–65. [Google Scholar] [CrossRef]
- Üstüner, T. The Effects of Mistletoe (Viscum album L.) on the Physiological Properties of Some Drupe Trees in Turkey. Turk. J. Agric. For. 2019, 43, 485–491. [Google Scholar] [CrossRef]
- Jäger, T.; Holandino, C.; Melo, M.; Condori Penaloza, E.; Passos Oliveira, A.; Garrett, R.; Glauser, G.; Grazi, M.; Ramm, H.; Urech, K.; et al. Metabolomics by UHPLC-Q-TOF Reveals Host Tree-Dependent Phytochemical Variation in Viscum album L. Plants 2021, 10, 1726. [Google Scholar] [CrossRef]
- Dash, S.P.; Dixit, S.; Sahoo, S. Phytochemical and Biochemical Characterizations from Leaf Extracts from Azadirachta Indica: An Important Medicinal Plant. Biochem. Anal. Biochem. 2017, 6, 1000323. [Google Scholar] [CrossRef]
- Luczkiewicz, M.; Cisowski, W.; Kaiser, P.; Ochocka, R.; Piotrowski, A. Comparative Analysis of Phenolic Acids in Mistletoe Plants from Various Hosts. Acta Pol. Pharm. 2001, 58, 373–379. [Google Scholar]
- Vicas, S.I.; Rugina, D.; Leopold, L.; Pintea, A.; Socaciu, C. HPLC Fingerprint of Bioactive Compounds and Antioxidant Activities of Viscum album from Different Host Trees. Not. Bot. Horti Agrobot. 2011, 1, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Turkkan, A.; Savas, H.B.; Yavuz, B.; Yigit, A.; Uz, E.; Bayram, N.A.; Kale, B. The Prophylactic Effect of Viscum album in Streptozotocin-Induced Diabetic Rats. North Clin. Istanb. 2016, 3, 83–89. [Google Scholar] [CrossRef]
- Kleinsimon, S.; Longmuss, E.; Rolff, J.; Jäger, S.; Eggert, A.; Delebinski, C.; Seifert, G. GADD45A and CDKN1A Are Involved in Apoptosis and Cell Cycle Modulatory Effects of Viscum TT with Further Inactivation of the STAT3 Pathway. Sci. Rep. 2018, 8, 5750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felenda, J.E.; Turek, C.; Stintzing, F.C. Antiproliferative Potential from Aqueous Viscum album L. Preparations and Their Main Constituents in Comparison with Ricin and Purothionin on Human Cancer Cells. J. Ethnopharmacol. 2019, 236, 100–107. [Google Scholar] [CrossRef]
- Oei, S.L.; Thronicke, A.; Kröz, M.; von Trott, P.; Schad, F.; Matthes, H. Impact of Oncological Therapy and Viscum album L Treatment on Cancer-Related Fatigue and Internal Coherence in Nonmetastasized Breast Cancer Patients. Integr. Cancer Ther. 2020, 19, 1534735420917211. [Google Scholar] [CrossRef]
- Stan, R.L.; Hangan, A.C.; Dican, L.; Sevastre, B.; Hanganu, D.; Catoi, C.; Sarpataki, O.; Ionescu, C.M. Comparative Study Concerning Mistletoe Viscotoxins Antitumor Activity. Biol. Futura 2013, 64, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Vicaş, S.; Rugină, D.; Socaciu, C. Comparative Study about Antioxidant Activities of Viscum album from Different Host Trees, Harvested in Different Seasons. J. Med. Plants Res. 2011, 5, 2237–2244. [Google Scholar]
- Pietrzak, W.; Nowak, R. Impact of Harvest Conditions and Host Tree Species on Chemical Composition and Antioxidant Activity of Extracts from Viscum album L. Molecules 2021, 26, 3741. [Google Scholar] [CrossRef]
- Karagöz, A.; Kesici, S.; Vural, A.; Usta, M.; Tezcan, B.; Semerci, T.; Teker, E. Cardioprotective Effects of Viscum album L. ssp. Album (Loranthaceae) on Isoproterenol-Induced Heart Failure via Regulation of the Nitric Oxide Pathway in Rats. Anatol. J. Cardiol. 2016, 16, 923–930. [Google Scholar] [CrossRef]
- Segneanu, A.-E.; Marin, C.N.; Herea, D.D.; Stanusoiu, I.; Muntean, C.; Grozescu, I. Romanian Viscum album, L.-Untargeted Low-Molecular Metabolomic Approach to Engineered Viscum-AuNPs Carrier Assembly. Plants 2022, 11, 1820. [Google Scholar] [CrossRef]
- Roxburgh, L.; Nicolson, S. Differential Dispersal and Survival of an African Mistletoe: Does Host Size Matter? Plant Ecol. 2008, 195, 21–31. [Google Scholar] [CrossRef]
- Ko, S.M.; Kwon, Y.K.; Kim, J.H.; Song, I.-J.; Lee, H.-Y.; Choi, D.-W.; Liu, J.R.; Kim, S.W. Transcriptome Analysis of Mistletoe (Viscum album) Haustorium Development. Hortic. Environ. Biotechnol. 2014, 55, 352–361. [Google Scholar] [CrossRef]
- Pennings, S.C.; Callaway, R.M. Parasitic Plants: Parallels and Contrasts with Herbivores. Oecologia 2002, 131, 479–489. [Google Scholar] [CrossRef]
- Mellado, A.; Zamora, R. Parasites Structuring Ecological Communities: The Mistletoe Footprint in Mediterranean Pine Forests. Funct. Ecol. 2017, 31, 2167–2176. [Google Scholar] [CrossRef] [Green Version]
- Bussing, A. Mistletoe: The Genus Viscum, 1st ed.; CRC Press: London, UK, 2000; ISBN 978-0-429-21986-3. [Google Scholar]
- Vicas, S.I.; Rugină, D.; Pantea, S.; Socaciu, C. The Morphological Features and UV-VIS Analysis of Some Taxonomic Markers of Genus Viscum. Bull. Univ. Agric. Sci. Vet. Med. Agric. 2009, 66, 193–200. [Google Scholar]
- Esteban, R.; Barrutia, O.; Artetxe, U.; Fernández-Marín, B.; Hernández, A.; García-Plazaola, J.I. Internal and External Factors Affecting Photosynthetic Pigment Composition in Plants: A Meta-Analytical Approach. New Phytol. 2015, 206, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.K.; Gautam, M. Biochemical Parameters of Plants as Indicators of Air Pollution. J. Environ. Biol. 2007, 28, 127–132. [Google Scholar]
- Vicaș, S.I.; Laslo, V.; Pantea, S.; Bandici, G.E. Chlorophyll and Carotenoids Pigments from Mistletoe (Viscum album) Leaves Using Different Solvents. An. Univ. Oradea Fasc. Biol. 2010, 17, 213–218. [Google Scholar]
- Skrypnik, L.; Maslennikov, P.; Feduraev, P.; Pungin, A.; Belov, N. Changes in Antioxidative Compounds and Enzymes in Small-Leaved Linden (Tilia cordata Mill.) in Response to Mistletoe (Viscum album L.) Infestation. Plants 2021, 10, 1871. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Ghosh, S.K. Alterations in Biochemical Components in Mesta Plants Infected with Yellow Vein Mosaic Disease. Braz. J. Plant Physiol. 2008, 20, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Üstüner, T.; Düzenli, S. Determination of the Chemical Compositions of Viscum album Ssp. Album and Soft Core Hosts. J. Chem. Metrol. 2017, 11, 68–77. [Google Scholar] [CrossRef]
- Abrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for Determination of Proline in Plants. Methods Mol. Biol. 2010, 639, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Signorelli, S.; Coitiño, E.L.; Borsani, O.; Monza, J. Molecular Mechanisms for the Reaction Between •OH Radicals and Proline: Insights on the Role as Reactive Oxygen Species Scavenger in Plant Stress. J. Phys. Chem. B 2014, 118, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Guerra, T.; Pizo, M.; Silva, W. Host Specificity and Aggregation for a Widespread Mistletoe in Campo Rupestre Vegetation. Flora Morphol. Distrib. Funct. Ecol. Plants 2017, 238, 148–154. [Google Scholar] [CrossRef] [Green Version]
- Moreira, F.; Teixeira-Costa, L.; Ceccantini, G.; Furlan, C.M. Mistletoe Effects on the Host Tree Tapirira Guianensis: Insights from Primary and Secondary Metabolites. Chemoecology 2019, 29, 11–24. [Google Scholar] [CrossRef]
- Marvibaigi, M.; Supriyanto, E.; Amini, N.; Abdul Majid, F.A.; Jaganathan, S.K. Preclinical and Clinical Effects of Mistletoe against Breast Cancer. Biomed. Res. Int. 2014, 2014, 785479. [Google Scholar] [CrossRef] [Green Version]
- Szurpnicka, A.; Zjawiony, J.K.; Szterk, A. Therapeutic Potential of Mistletoe in CNS-Related Neurological Disorders and the Chemical Composition of Viscum Species. J. Ethnopharmacol. 2019, 231, 241–252. [Google Scholar] [CrossRef]
- Song, C.; Wei, X.-Y.; Qiu, Z.-D.; Gong, L.; Chen, Z.-Y.; Ma, Y.; Shen, Y.; Zhao, Y.-J.; Wang, W.-H.; Lai, C.-J.-S.; et al. Exploring the Resources of the Genus Viscum for Potential Therapeutic Applications. J. Ethnopharmacol. 2021, 277, 114233. [Google Scholar] [CrossRef]
- Holandino, C.; de Oliveira Melo, M.N.; Oliveira, A.P.; da Costa Batista, J.V.; Capella, M.A.M.; Garrett, R.; Grazi, M.; Ramm, H.; Torre, C.D.; Schaller, G.; et al. Phytochemical Analysis and in Vitro Anti-Proliferative Activity of Viscum album Ethanolic Extracts. BMC Complement. Med. Ther. 2020, 20, 215. [Google Scholar] [CrossRef]
- Yang, P.; Jiang, Y.; Pan, Y.; Ding, X.; Rhea, P.; Ding, J.; Hawke, D.H.; Felsher, D.; Narla, G.; Lu, Z.; et al. Mistletoe Extract Fraxini Inhibits the Proliferation of Liver Cancer by Down-Regulating c-Myc Expression. Sci. Rep. 2019, 9, 6428. [Google Scholar] [CrossRef] [Green Version]
- Menke, K.; Schwermer, M.; Eisenbraun, J.; Schramm, A.; Zuzak, T.J. Anticancer Effects of Viscum album Fraxini Extract on Medulloblastoma Cells in Vitro. Complement. Med. Res. 2021, 28, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Peñaloza, E.; Holandino, C.; Scherr, C.; de Araujo, P.I.P.; Borges, R.M.; Urech, K.; Baumgartner, S.; Garrett, R. Comprehensive Metabolome Analysis of Fermented Aqueous Extracts of Viscum album L. by Liquid Chromatography−High Resolution Tandem Mass Spectrometry. Molecules 2020, 25, 4006. [Google Scholar] [CrossRef] [PubMed]
- Stefanucci, A.; Zengin, G.; Llorent-Martinez, E.J.; Dimmito, M.P.; Della Valle, A.; Pieretti, S.; Ak, G.; Sinan, K.I.; Mollica, A. Viscum album L. Homogenizer-Assisted and Ultrasound-Assisted Extracts as Potential Sources of Bioactive Compounds. J. Food Biochem. 2020, 44, e13377. [Google Scholar] [CrossRef]
- Nazaruk, J.; Orlikowski, P. Phytochemical Profile and Therapeutic Potential of Viscum album L. Nat. Prod. Res. 2016, 30, 373–385. [Google Scholar] [CrossRef]
- Onay-Uçar, E.; Karagöz, A.; Arda, N. Antioxidant Activity of Viscum album Ssp. Album. Fitoterapia 2006, 77, 556–560. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhou, X.-Z.; Ye, L.; Yuan, Q.; Freeberg, S.; Shi, C.; Zhu, P.-W.; Bao, J.; Jiang, N.; Shao, Y. Rhamnazin Attenuates Inflammation and Inhibits Alkali Burn-Induced Corneal Neovascularization in Rats. RSC Adv. 2018, 8, 26696–26706. [Google Scholar] [CrossRef] [Green Version]
- Papuc, C.; Crivineanu, M.; Goran, G.; Nicorescu, V.; Durdun, C. Free Radicals Scavenging and Antioxidant Activity of European Mistletoe (Viscum album) and European Birthwort (Aristolochia clematitis). Rev. Chim. 2010, 61, 619–622. [Google Scholar]
- Pelzer, F.; Tröger, W. Complementary Treatment with Mistletoe Extracts During Chemotherapy: Safety, Neutropenia, Fever, and Quality of Life Assessed in a Randomized Study. J. Altern. Complement. Med. 2018, 24, 954–961. [Google Scholar] [CrossRef]
- Lech, P.; Żółciak, A.; Hildebrand, R. Occurrence of European Mistletoe (Viscum album L.) on Forest Trees in Poland and Its Dynamics of Spread in the Period 2008–2018. Forests 2020, 11, 83. [Google Scholar] [CrossRef]
- Nayek, S.; Choudhury, I.; Haque, J.; Nishika, J.; Roy, S. Spectrophotometric Analysis of Chlorophylls and Carotenoids from Commonly Grown Fern Species by Using Various Extracting Solvents. Res. J. Chem. Sci. 2014, 4, 2231–2606. [Google Scholar] [CrossRef] [Green Version]
- Aleixandre-Tudo, J.L.; du Toit, W. The Role of UV-Visible Spectroscopy for Phenolic Compounds Quantification in Winemaking; IntechOpen: London, UK, 2018; ISBN 978-1-78985-496-1. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Oxidants and Antioxidants Part A.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 152–178. [Google Scholar]
- Pallag, A.; Bungau, S.; Tit, D.M.; Tünde, J.; Sirbu, V.; Honiges, A.; Horhogea, C. Comparative Study of Polyphenols, Flavonoids and Chlorophylls in Equisetum arvense L. Populations. Rev. Chim. 2016, 67, 530–533. [Google Scholar]
- Miere Groza, F.; Teușdea, A.C.; Laslo, V.; Cavalu, S.; Fritea, L.; Dobjanschi, L.; Zdrinca, M.; Zdrinca, M.; Ganea, M.; Pașc, P.; et al. Evaluation of In Vitro Wound-Healing Potential, Antioxidant Capacity, and Antimicrobial Activity of Stellaria media (L.) Vill. Appl. Sci. 2021, 11, 11526. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Budau, R.; Memete, A.; Adrian Ioan, T.; Vicaş, S. Phytochemical Screening and Antioxidant Capacity of Two Berry Cultivars, ‘Ruben’ and ‘Duke’, Depending on Their Harvesting Time. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Food Sci. Technol. 2022, 1, 27–35. [Google Scholar] [CrossRef]
- Memete, A.R.; Teusdea, A.C.; Timar, A.V.; Vuscan, A.N.; Mintaș, O.S.; Cavalu, S.; Vicas, S.I. Effects of Different Edible Coatings on the Shelf Life of Fresh Black Mulberry Fruits (Morus nigra L.). Agriculture 2022, 12, 1068. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trees | F (%) | Infested Trees | Number of Mistletoes on Host Trees | Height of Infested Host Trees (m) | Height of the Uninfested Trees (m) | Circumference of the Trunk of Infested Host Trees (m) | Circumference of the Trunk of Uninfested Host Trees (m) |
|---|---|---|---|---|---|---|---|
| Malus domestica Barkh. | 45.83 | 11 | 1–5 | 5.38 ± 1.35 c | 4.93 ± 1.80 c | 0.85 ± 0.24 b | 0.69 ± 0.18 b |
| Prunus domestica L. | 36.84 | 7 | 1–2 | 7.61 ± 1.85 b | 6.04 ± 1.72 b | 0.57 ± 0.15 c | 0.54 ± 0.13 c |
| Populus alba L. | 70.37 | 19 | 3–20 | 14.01 ± 2.43 a | 9.1 ± 0.73 a | 1.79 ± 0.68 a | 1.51 ± 0.63 a |
| Host Tree | VAM | VAP | VAO |
|---|---|---|---|
 |  |  | |
| Length leaves * (min–max) | 6.13 ± 1.27 b (0.8–8.6) | 7.18 ± 1.15 a (0.8–10) | 4.14 ± 1.18 c (0.6–4.4) |
| Samples | Chlorophyll a | Chlorophyll b | Total Carotenoids |
|---|---|---|---|
| VAM | 1.226 ± 0.005 g | 0.493 ± 0.045 g | 0.389 ± 0.008 g |
| M + VAM | 1.734 ± 0.016 e | 0.670 ± 0.038 e | 0.532 ± 0.007 e |
| M | 2.451 ± 0.141 c | 1.096 ± 0.086 c | 0.674 ± 0.008 c |
| VAP | 1.233 ± 0.012 g | 0.497 ± 0.006 g | 0.379 ± 0.001 g |
| P + VAP | 2.711 ± 0.006 b | 1.042 ± 0.003 b | 0.868 ± 0.003 a |
| P | 3.001 ± 0.046 a | 1.133 ± 0.109 a | 0.848 ± 0.007 a |
| VAO | 1.427 ± 0.023 f | 0.607 ± 0.033 f | 0.480 ± 0.000 f |
| O + VAO | 1.957 ± 0.059 d | 0.889 ± 0.161 d | 0.578 ± 0.026 d |
| O | 2.869 ± 0.005 a | 1.257 ± 0.011 a | 0.706 ± 0.003 b |
| Peak No. | Rt (min) | UV λmax (nm) | [M + H+] (m/z) | Compound | Subclass | VAM | VAP | VAO |
|---|---|---|---|---|---|---|---|---|
| 1 | 3.33 | 265 | 155 | Dihydroxybenzoic acid | Hydroxybenzoic acids | 2.107 ± 0.19 b | 2.695 ± 0.20 a | 3.055 ± 0.28 a |
| 2 | 10.85 | 323 | 355, 163 | 3-Caffeoylquinic acid (Neochlorogenic acid) | Hydroxycinnamic acids | 0.775 ± 0.09 a | 0.356 ± 0.03 b | 0.721 ± 0.06 a |
| 3 | 11.85 | 323 | 355, 163 | 4-Caffeoylquinic acid (Criptochlorogenic acid) | 0.313 ± 0.02 a | 0.181 ± 0.01 b | nd | |
| 4 | 12.47 | 323 | 355, 163 | 5-Caffeoylquinic acid (Chlorogenic acid) | 1.413 ± 0.11 a | 0.551 ± 0.04 c | 1.000 ± 0.07 b | |
| 5 | 13.21 | 330 | 387, 223 | Sinapic acid glucoside | 0.490 ± 0.04 a | 0.303 ± 0.02 b | 0.473 ± 0.04 a | |
| 6 | 13.85 | 330 | 475, 163 | Dicaffeoyl tartaric acid | 0.585 ± 0.05 | nd | nd | |
| 7 | 14.28 | 330 | 399, 223 | 3-Sinapoylquinic acid | 0.536 ± 0.04 a | 0.431 ± 0.03 b | 0.016 ± 0.00 c | |
| 8 | 14.84 | 330 | 399, 223 | 5-Sinapoylquinic acid | 1.044 ± 0.09 a | 0.584 ± 0.05 b | 0.663 ± 0.06 b | |
| 9 | 15.75 | 255, 360 | 611, 303 | Quercetin rutinoside (Rutin) | Flavonol glycoside | 0.171 ± 0.01 b | nd | 0.308 ± 0.02 a |
| 10 | 16.18 | 255, 360 | 465, 303 | Quercetin glucoside | 0.323 ± 0.02 a | 0.486 ± 0.04 b | 0.786 ± 0.06 c | |
| 11 | 16.77 | 330 | 225 | Sinapic acid | Hydroxycinnamic acid | 0.570 ± 0.05 b | 0.458 ± 0.03 b | 1.105 ± 0.09 a |
| 12 | 17.01 | 255, 360 | 609, 303 | Quercetin-O-[hydroxymethylglutaryl] hexoside (Quercetin derivative) | Flavonol | 0.392 ± 0.03 a | 0.374 ± 0.02 a | 0.432 ± 0.03 a |
| 13 | 17.69 | 240, 350 | 479, 317 | Isorhamnetin glucoside | 0.270 ± 0.01 b | 0.232 ± 0.02 b | 0.356 ± 0.03 a | |
| 14 | 18.11 | 240, 350 | 623, 317 | Isorhamnetin-O-[hydroxymethylglutaryl] hexoside (Isorhamnetin derivative) | 0.458 ± 0.04 b | 0.444 ± 0.04 b | 0.830 ± 0.07 a | |
| 15 | 18.77 | 240, 350 | 493, 317 | Isorhamnetin-glucuronide | 0.445 ± 0.03 b | 0.390 ± 0.03 b | 0.695 ± 0.06 a | |
| 16 | 19.51 | 240, 350 | 755, 317 | Isorhamnein-(dirhamnosyl)-rhamnoside | 0.354 ± 0.03 b | 0.368 ± 0.02 b | 0.659 ± 0.05 a | |
| 17 | 19.81 | 240, 350 | 625, 317 | Isorhamnein-glucosyl-rhamnoside | 0.249 ± 0.01 a | 0.317 ± 0.02 b | 0.503 ± 0.04 c | |
| 18 | 20.35 | 245, 350 | 493, 331 | Rhamnazin glucoside | 0.478 ± 0.04 b | 0.514 ± 0.04 b | 1.025 ± 0.08 a | |
| 19 | 20.71 | 245, 350 | 639, 331 | Rhamnazin rutinoside | 0.279 ± 0.01 b | 0.372 ± 0.03 b | 0.653 ± 0.06 a | |
| 20 | 21.64 | 255, 360 | 303 | Quercetin | 0.137 ± 0.01 b | 0.200 ± 0.01 b | 0.677 ± 0.06 a | |
| 21 | 24.03 | 240, 350 | 317 | Isorhamnetin | 0.042 ± 0.003 b | 0.210 ± 0.02 a | 0.212 ± 0.01 a |
| Code | Explanation of Coding |
|---|---|
| VAM | Leaves of V. album L. subsp. album parasitizing the apple (M. domestica Barkh.) |
| VAP | Leaves of V. album L. subsp. album parasitizing the plum (P. domestica L.) |
| VAO | Leaves of V. album L. subsp. album parasitizing the poplar (P. alba L.) |
| M + VAM | Apple leaves from the tree that is infested with mistletoe |
| P + VAP | Plum leaves from the tree that is infested with mistletoe |
| O + VAO | Poplar leaves from the tree that is infested with mistletoe |
| M | Leaves from an apple tree not infested with mistletoe |
| P | Leaves from a plum tree not infested with mistletoe |
| O | Leaves from a poplar tree not infested with mistletoe |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kleszken, E.; Purcarea, C.; Pallag, A.; Ranga, F.; Memete, A.R.; Miere, F.; Vicas, S.I. Phytochemical Profile and Antioxidant Capacity of Viscum album L. Subsp. album and Effects on Its Host Trees. Plants 2022, 11, 3021. https://doi.org/10.3390/plants11223021
Kleszken E, Purcarea C, Pallag A, Ranga F, Memete AR, Miere F, Vicas SI. Phytochemical Profile and Antioxidant Capacity of Viscum album L. Subsp. album and Effects on Its Host Trees. Plants. 2022; 11(22):3021. https://doi.org/10.3390/plants11223021
Chicago/Turabian StyleKleszken, Eva, Cornelia Purcarea, Annamaria Pallag, Floricuta Ranga, Adriana Ramona Memete, Florina Miere (Groza), and Simona Ioana Vicas. 2022. "Phytochemical Profile and Antioxidant Capacity of Viscum album L. Subsp. album and Effects on Its Host Trees" Plants 11, no. 22: 3021. https://doi.org/10.3390/plants11223021