
Molecular Hydrogen Improves Rice Storage Quality via Alleviating Lipid Deterioration and Maintaining Nutritional Values

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
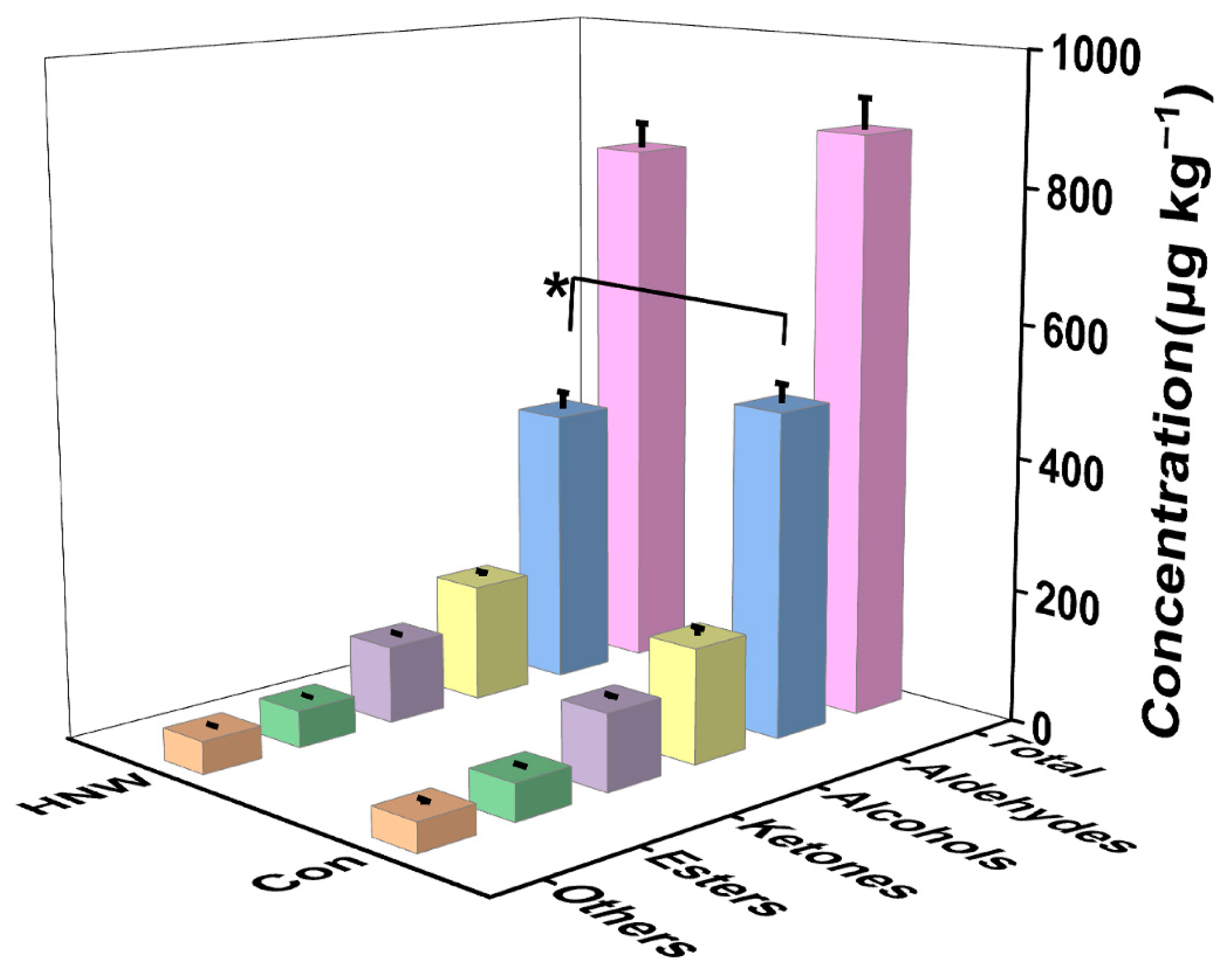
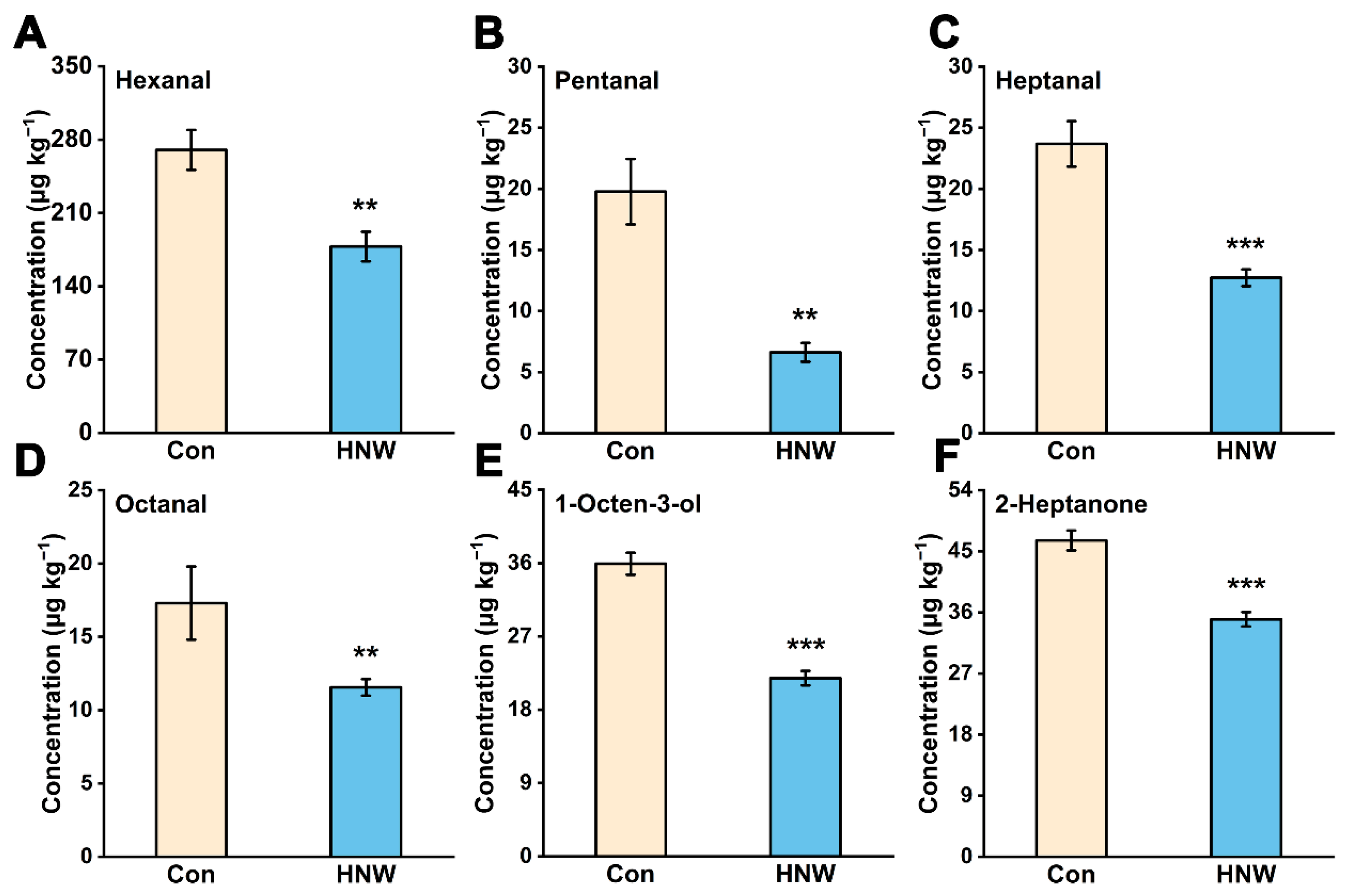
2.1. Preharvest HNW Treatment Alleviates the Generation of Off-Flavor during Rice Storage
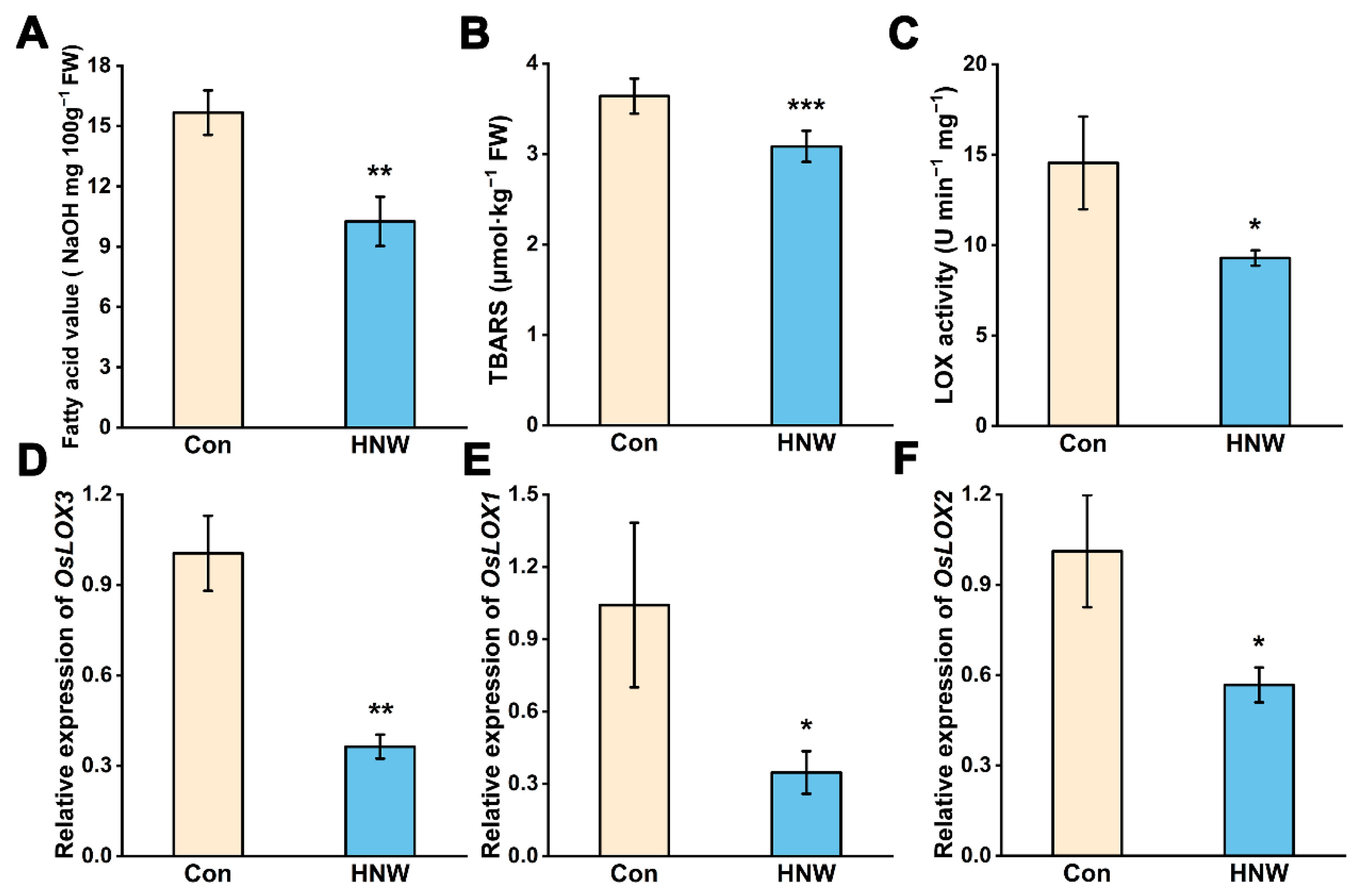
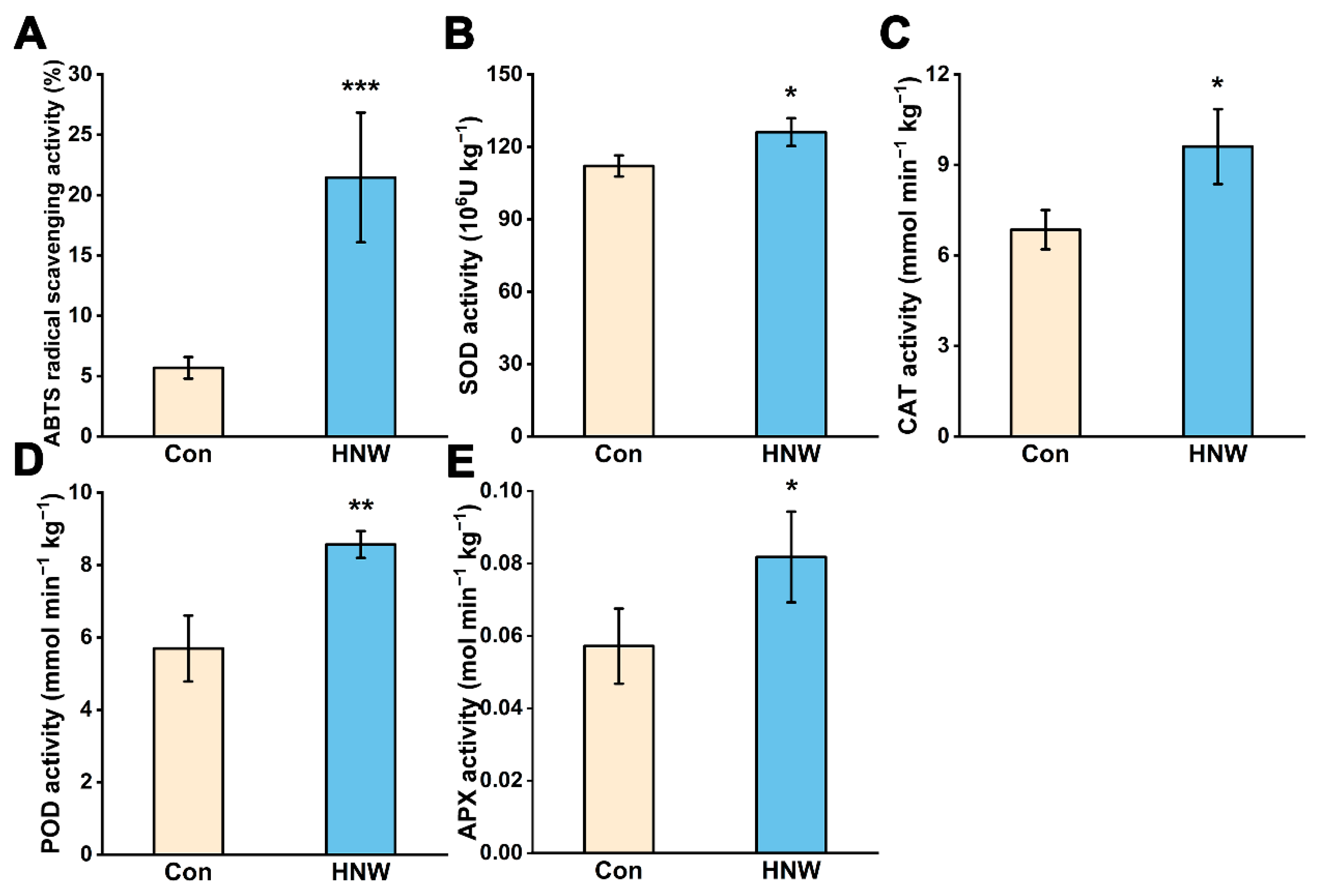
2.2. Alleviation in Lipid Oxidation and Increased Antioxidant Machinery in Stored Rice
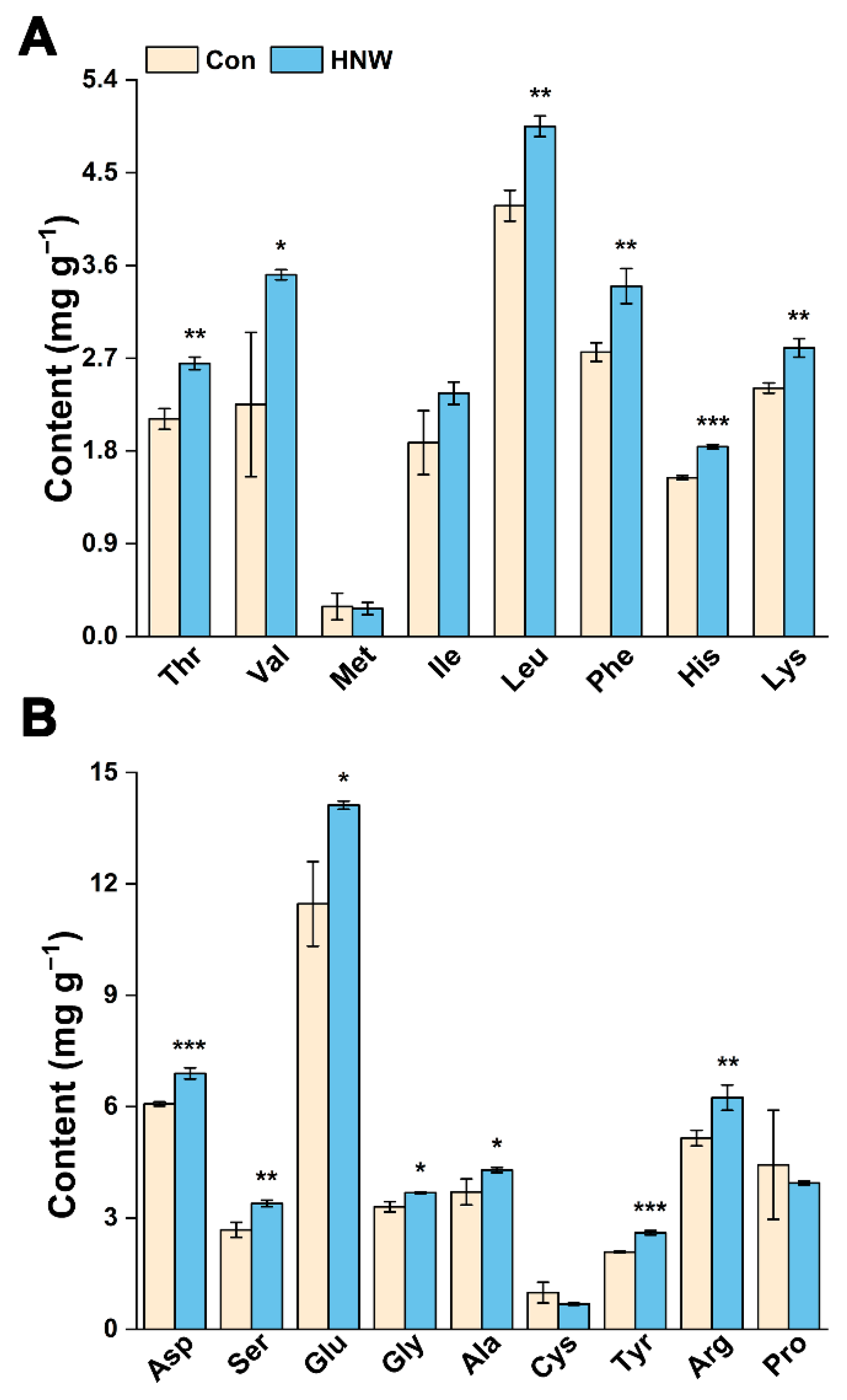
2.3. Preharvest HNW Treatment Maintains High Levels of Amino Acid Contents
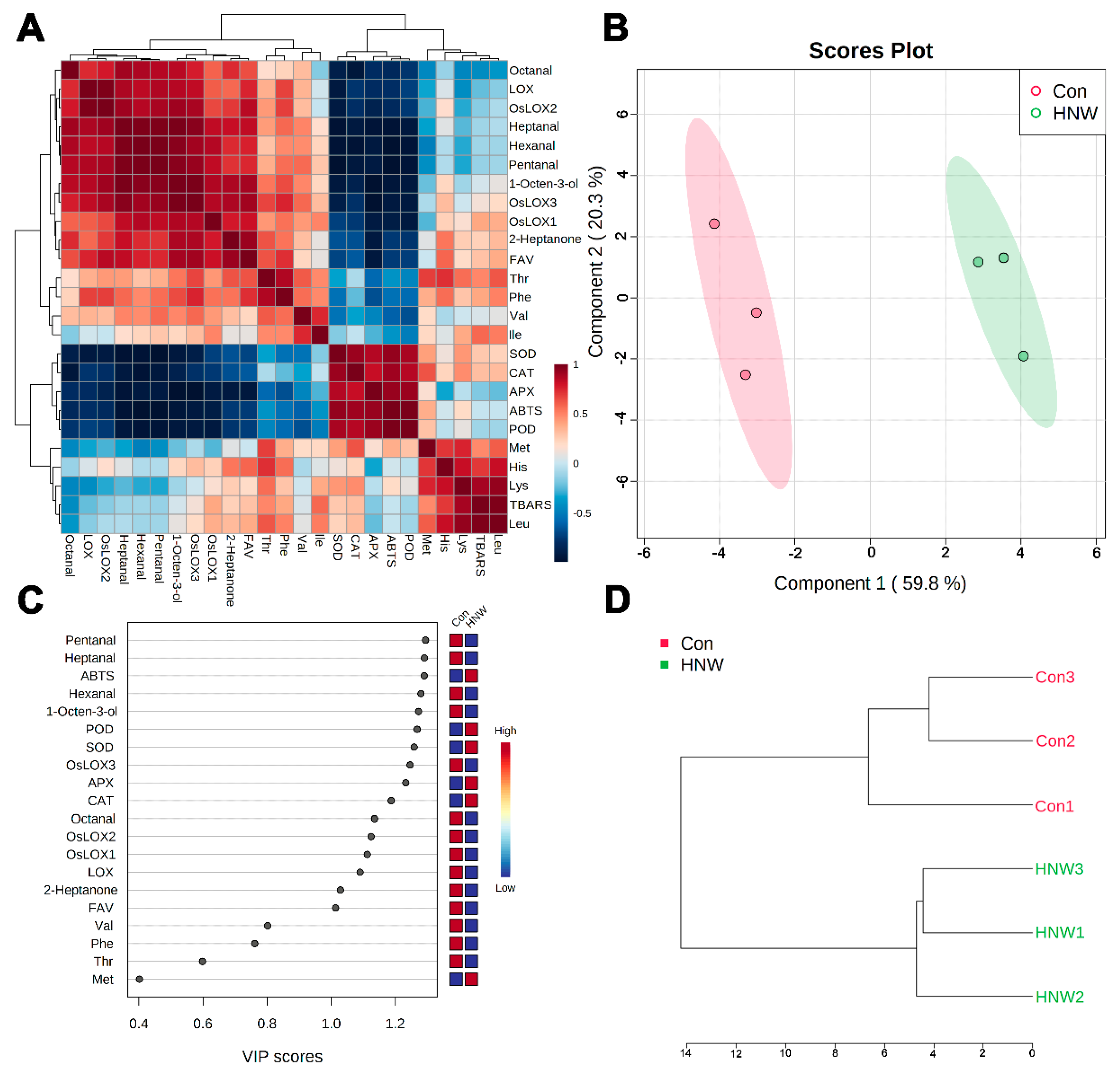
2.4. The Influence of HNW on Rice Storage Characters by Correlation Analysis and PLS-DA
3. Materials and Methods
3.1. Plant Material and Treatments
3.2. Extraction and Analysis of Volatiles
3.3. Quantification of Fatty Acid Value (FAV) and Thiobarbituric Acid Reactive Substances (TBARS)
3.4. Determination of 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic Acid (ABTS) Radical Scavenging Rate
3.5. Assay of Antioxidant Enzymes and Lipoxygenase (LOX) Activities
3.6. Transcriptional Analysis of Lipoxygenase Genes
3.7. Analysis of Hydrolysis Amino Acid Content
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tong, C.; Gao, H.; Luo, S.; Liu, L.; Bao, J. Impact of postharvest operations on rice grain quality: A review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 626–640. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data (accessed on 31 May 2022).
- Nation Food and Strategic Reserves Administration. Available online: http://www.lswz.gov.cn/html/zt/kjhdz2021/2021-06/09/content_266101.shtml (accessed on 31 May 2022).
- Hu, X.; Lu, L.; Guo, Z.; Zhu, Z. Volatile compounds, affecting factors and evaluation methods for rice aroma: A review. Trends Food Sci. Technol. 2020, 97, 136–146. [Google Scholar] [CrossRef]
- Shin, M.G.; Yoon, S.H.; Rhee, J.S.; Kwon, T.W. Correlation between oxidative deterioration of unsaturated lipid and n-hexanal during storage of brown rice. J. Food Sci. 1986, 51, 460–463. [Google Scholar] [CrossRef]
- Monsoor, M.A.; Proctor, A. Volatile component analysis of commercially milled head and broken rice. J. Food Sci. 2004, 69, 632–636. [Google Scholar] [CrossRef]
- Belitz, H.D.; Grosch, W.; Schieberle, P. Lipids. In Food Chemistry, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2009; p. 158. ISBN 978-3-540-69934-7. [Google Scholar]
- Griglione, A.; Liberto, E.; Cordero, C.; Bressanello, D.; Cagliero, C.; Rubiolo, P.; Bicchi, C.; Sgorbini, B. High-Quality Italian rice cultivars: Chemical indices of ageing and aroma quality. Food Chem. 2015, 172, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Ise, K.; Li, C.; Honda, I.; Iwai, Y.; Matsukura, U. Volatile components in stored rice [Oryza sativa (L.)] of varieties with and without lipoxygenase-3 in seeds. J. Agric. Food Chem. 1999, 47, 1119–1124. [Google Scholar] [CrossRef]
- Schwab, W.; Davidovich-Rikanati, R.; Lewinsohn, E. Biosynthesis of plant-derived flavor compounds. Plant J. 2008, 54, 712–732. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Z.; Lu, Y.; Wang, Y.; She, D.; Song, M.; Wu, Y. Effect of the absence of lipoxygenase isoenzymes on the storage characteristics of rice grains. J. Stored Prod. Res. 2007, 43, 87–91. [Google Scholar] [CrossRef]
- Nice, D.J.; Robinson, D.S. Inhibition of lipid autoxidation by bovine superoxide dismutase. Food Chem. 1992, 45, 99–103. [Google Scholar] [CrossRef]
- Guo, F.X.; Zhang, M.X.; Chen, Y.; Zhang, W.H.; Xu, S.J.; Wang, J.H.; An, L.Z. Relation of several antioxidant enzymes to rapid freezing resistance in suspension cultured cells from alpine Chorispora Bungeana. Cryobiology 2006, 52, 241–250. [Google Scholar] [CrossRef]
- Ngoh, S.K.; Njomo, D. An overview of hydrogen gas production from solar energy. Renew. Sustain. Energy Rev. 2012, 16, 6782–6792. [Google Scholar] [CrossRef]
- Sharma, S.; Agarwal, S.; Jain, A. Significance of hydrogen as economic and environmentally friendly fuel. Energies 2021, 14, 7389. [Google Scholar] [CrossRef]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.I.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Kuang, Y.; Feng, L.; Liu, Y.; Wang, S.; Du, H.; Shen, W. Molecular hydrogen maintains the storage quality of Chinese chive through improving antioxidant capacity. Plants 2021, 10, 1095. [Google Scholar] [CrossRef]
- Li, L.; Liu, Y.; Wang, S.; Zou, J.; Ding, W.; Shen, W. Magnesium hydride-mediated sustainable hydrogen supply prolongs the vase life of cut carnation flowers via hydrogen sulfide. Front. Plant Sci. 2020, 11, 595376. [Google Scholar] [CrossRef]
- Bulut, M.; Sezer, Y.Ç.; Ceylan, M.M.; Alwazeer, D.; Koyuncu, M. Hydrogen-Rich water can reduce the formation of biogenic amines in butter. Food Chem. 2022, 384, 132613. [Google Scholar] [CrossRef]
- Hu, H.; Li, P.; Shen, W. Preharvest application of hydrogen-rich water not only affects daylily bud yield but also contributes to the alleviation of bud browning. Sci. Hortic. 2021, 287, 110267. [Google Scholar] [CrossRef]
- Li, L.; Lou, W.; Kong, L.; Shen, W. Hydrogen commonly applicable from medicine to agriculture: From molecular mechanisms to the field. Curr. Pharm. Des. 2021, 27, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Wang, J.; Zhao, Z.; Kong, L.; Lou, W.; Zhang, T.; Jing, D.; Yu, J.; Shu, Z.; Huang, L.; et al. Molecular hydrogen increases quantitative and qualitative traits of rice grain in field trials. Plants 2021, 10, 2331. [Google Scholar] [CrossRef]
- Zhao, Q.; Yousaf, L.; Xue, Y.; Shen, Q. Changes in flavor of fragrant rice during storage under different conditions. J. Sci. Food Agric. 2020, 100, 3435–3444. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, B.; Dong, J.; Tian, Y.; Lin, Y.; Fang, G.; Wang, S. Identification of mouldy rice using an electronic nose combined with SPME-GC/MS. J. Stored Prod. Res. 2022, 95, 101921. [Google Scholar] [CrossRef]
- Li, L.; Wang, J.; Jiang, K.; Kuang, Y.; Zeng, Y.; Cheng, X.; Liu, Y.; Wang, S.; Shen, W. Preharvest application of hydrogen nanobubble water enhances strawberry flavor and consumer preferences. Food Chem. 2022, 377, 131953. [Google Scholar] [CrossRef]
- Kechkin, I.A.; Ermolaev, V.A.; Ivanov, M.V.; Romanenko, A.I.; Gurkovskaya, E.A. Dependence of fat acidity value on wheat grain storage conditions. BIO Web Conf. 2020, 17, 00107. [Google Scholar] [CrossRef]
- Shi, J.; Zhang, T.; Wang, T.; Wu, M. Effects of glutelin and lipid oxidation on the physicochemical properties of rice starch. Cereal Chem. 2021, 98, 683–692. [Google Scholar] [CrossRef]
- Birla, D.S.; Malik, K.; Sainger, M.; Chaudhary, D.; Jaiwal, R.; Jaiwal, P.K. Progress and challenges in improving the nutritional quality of rice (Oryza sativa L.). Crit. Rev. Food Sci. Nutr. 2017, 57, 245–2481. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Gao, C.; Fang, P.; Lin, G.; Shen, W. Alleviation of cadmium toxicity in Medicago sativa by hydrogen-rich water. J. Hazard. Mater. 2013, 260, 715–724. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Kuang, Y.; Shen, W. Packaging with hydrogen gas modified atmosphere can extend chicken egg storage. J. Sci. Food Agric. 2021, 102, 976–983. [Google Scholar] [CrossRef]
- Jin, Z.; Zhao, P.; Gong, W.; Ding, W.; He, Q. Fe-Porphyrin: A redox-related biosensor of hydrogen molecule. Nano Res. 2022, 1–6. [Google Scholar] [CrossRef]
- Kubota, R.; Asayama, S.; Kawakami, H. A bioinspired polymer-bound Mn-porphyrin as an artificial active center of catalase. Chem. Commun. 2014, 50, 15909–15912. [Google Scholar] [CrossRef]
- Wang, Q.; Lei, J.; Deng, S.; Zhang, L.; Ju, H. Graphene-Supported ferric porphyrin as a peroxidase mimic for electrochemical DNA biosensing. Chem. Commun. 2013, 49, 916. [Google Scholar] [CrossRef]
- Pappa, H.; Patterson, W.R.; Poulos, T.L. The homologous tryptophan critical for cytochrome c peroxidase function is not essential for ascorbate peroxidase activity. J. Biol. Inorg. Chem. 1996, 1, 61–66. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, J.; Hao, H.; Feng, Z.; Chen, M.; Wang, H.; Ye, M. Hydrogen-Rich water increases postharvest quality by enhancing antioxidant capacity in Hypsizygus marmoreus. AMB Expr. 2017, 7, 221. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.; Li, X.; Li, C.; Tang, Q.; Duan, B.; Cheng, Y.; Hou, J.; Li, J. Effect of sodium nitroprusside on antioxidative enzymes and the phenylpropanoid pathway in blueberry fruit. Food Chem. 2019, 295, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Zhu, D.; Yao, Q.; Tang, H.; Ding, X. Hydrogen-Rich water treatment maintains the quality of Rosa sterilis fruit by regulating antioxidant capacity and energy metabolism. LWT 2022, 161, 113361. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, Y.; Cheng, H.; Zhang, G.; Lyu, W. Biochemical composition distribution in different grain layers is associated with the edible quality of rice cultivars. Food Chem. 2019, 311, 125896. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Wang, E.; Li, C.; Cai, M.; Cheng, B.; Cao, C.; Jiang, Y. Use of protein content, amylose content, and RVA parameters to evaluate the taste quality of rice. Front. Nutr. 2022, 8, 758547. [Google Scholar] [CrossRef]
- Chen, H.; Chen, D.; He, L.; Wang, T.; Lu, H.; Yang, F.; Deng, F.; Chen, Y.; Tao, Y.; Li, M.; et al. Correlation of taste values with chemical compositions and Rapid Visco Analyser profiles of 36 indica rice (Oryza sativa L.) varieties. Food Chem. 2021, 349, 129176. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Chen, S.; Chen, F.; Chen, F. Concentration-Dependent emission of floral scent terpenoids from diverse cultivars of Chrysanthemum morifolium and their wild relatives. Plant Sci. 2021, 309, 110959. [Google Scholar] [CrossRef]
- Choi, S.; Seo, H.S.; Lee, K.R.; Lee, S.; Lee, J.; Lee, J. Effect of milling and long-term storage on volatiles of black rice (Oryza sativa L.) determined by headspace solid-phase microextraction with gas chromatography—Mass spectrometry. Food Chem. 2019, 276, 572–582. [Google Scholar] [CrossRef]
- Gayen, D.; Ali, N.; Sarker, S.N.; Datta, S.K.; Datta, K. Down-Regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta 2015, 242, 353–363. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, Y.; Liu, X.; Jiang, L.; Chen, L.; Han, X.; Jin, M.; Liu, S.; Liu, F.; Lv, J.; et al. OsRab5a regulates endomembrane organization and storage protein trafficking in rice endosperm cells. Plant J. 2010, 64, 812–824. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Z.; Zhang, X.H.; Yue, D.M. Alternation of antioxidative enzyme gene expression in rice seedlings exposed to methylene blue. Environ. Sci. Pollut. Res. 2014, 21, 14014–14022. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, R.P.; Singh, P.K.; Awasthi, S.; Chakrabarty, D.; Trivedi, P.K.; Tripathi, R.D. Selenium ameliorates arsenic induced oxidative stress through modulation of antioxidant enzymes and thiols in rice (Oryza sativa L.). Ecotoxicology 2014, 23, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.; Manneberg, M.; Juranville, J.F.; Lahm, H.W.; Fountoulakis, M. Effect of the hydrolysis method on the determination of the amino acid composition of proteins. J. Chromatogr. A 1998, 795, 263–275. [Google Scholar] [CrossRef]
- Yang, J.; Wang, Y.; Liu, L.; Liu, L.; Wang, C.; Wang, C.; Li, C. Effects of exogenous salicylic acid and pH on pathogenicity of biotrophy-associated secreted protein 1 (BAS1)-overexpressing strain, Magnaporthe oryzae. Environ. Sci. Pollut. Res. 2018, 26, 13725–13737. [Google Scholar] [CrossRef]
- Xu, H.; Wei, Y.; Zhu, Y.; Lian, L.; Xie, H.; Cai, Q.; Chen, Q.; Lin, Z.; Wang, Z.; Xie, H.; et al. Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity. Plant Biotechnol. J. 2014, 13, 526–539. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, C.; Zhao, Z.; Zhang, Y.; Li, M.; Li, L.; Cheng, P.; Shen, W. Molecular Hydrogen Improves Rice Storage Quality via Alleviating Lipid Deterioration and Maintaining Nutritional Values. Plants 2022, 11, 2588. https://doi.org/10.3390/plants11192588
Cai C, Zhao Z, Zhang Y, Li M, Li L, Cheng P, Shen W. Molecular Hydrogen Improves Rice Storage Quality via Alleviating Lipid Deterioration and Maintaining Nutritional Values. Plants. 2022; 11(19):2588. https://doi.org/10.3390/plants11192588
Chicago/Turabian StyleCai, Chenxu, Zhushan Zhao, Yingying Zhang, Min Li, Longna Li, Pengfei Cheng, and Wenbiao Shen. 2022. "Molecular Hydrogen Improves Rice Storage Quality via Alleviating Lipid Deterioration and Maintaining Nutritional Values" Plants 11, no. 19: 2588. https://doi.org/10.3390/plants11192588