
The Effect of Frankia and Hebeloma crustiliniforme on Alnus alnobetula subsp. Crispa Growing in Saline Soil


Department of Biological Sciences, University of Manitoba, Winnipeg, MB R3T 2N2, Canada
*
Authors to whom correspondence should be addressed.
Plants 2022, 11(14), 1860; https://doi.org/10.3390/plants11141860
Submission received: 3 June 2022
/
Revised: 11 July 2022
/
Accepted: 13 July 2022
/
Published: 16 July 2022
(This article belongs to the Special Issue Abiotic Stress Responses in Woody Plants)
Abstract
:The mining of the oil sands region of Canada’s boreal forest creates disturbed land with elevated levels of salts. Understanding how native plants respond to salt stress is critical in reclaiming these lands. The native species, Alnus alnobetula subsp. crispa forms nitrogen-fixing nodules with Frankia, and ectomycorrhizae with a number of fungal species. These relationships may make the plant particularly well suited for restoring disturbed land. We inoculated A. alnobetula subsp. crispa with Frankia and Hebeloma crustiliniforme and exposed the plants to 0, 50, or 100 mM NaCl for seven weeks. Frankia-inoculated plants had increased biomass regardless of salt exposure, even though salt exposure reduced nitrogen fixation and reduced the efficiency of nitrogen-fixing nodules. The nitrogen-fixing symbiosis also decreased leaf stress and increased root phosphatase levels. This suggests that N-fixing plants not only have increased nitrogen nutrition but also have increased access to soil phosphorus. Mycorrhizae did not affect plant growth but did reduce nodule numbers and nodule efficiency. These results suggest that the nitrogen-fixing trait is more critical than mycorrhizae. While salt stress inhibits nitrogen-fixing symbiosis, plants still benefit from nitrogen fixation when exposed to salt.
1. Introduction
Salinity is one of the most globally important abiotic plant stressors. About 400 million hectares of land are salt-affected (http://www.fao.org/soils-portal/soil-management, accessed on 1 May 2022). In cultivated lands, salinity stress is commonly associated with irrigation [1]. In uncultivated land, salinity stress can also result from industrial activities. The extraction of bitumen from oil sands in the boreal region of western Canada results in the production of saline tailings material [2,3]. Approximately 70,000 ha of land have been disturbed due to oil sands development [4]. Revegetating areas with this tailings waste is a challenge, in part due to its salt levels.
Salt acts as an osmotic stress, restricting plant uptake of water, and an ionic stress, interfering with plant ion balance [5]. Both of these stresses can affect whole plant performance, reducing the mass of roots and shoots, plant height, leaf expansion, and basic physiological functions [6]. Most research on salt stress in plants has been conducted on crop species in temperate or arid regions. We know much less about plant responses to salt stress in the boreal region. This is especially true of nitrogen-fixing species, which could play a vital role in restoring industrially disturbed land in boreal ecosystems [7].
Actinorhizal plants form nitrogen-fixing nodules in association with the actinomycete Frankia. Many actinorhizal plants grow in low fertility soil and play an important role as pioneer plants early in plant community development [8]. Alnus is one of the most widely distributed actinorhizal genera [9,10]. In areas with a low availability of inorganic nitrogen, Alnus species rely heavily on N fixation, which not only meets their N requirement but also increases the fertility of the soil [11]. Alnus species, therefore, have the potential to be used in the reclamation of disturbed areas [4,12,13]. Some Alnus species are tolerant of soil salinity [14]. However, although Frankia can tolerate high salinity levels in vitro, nitrogenase activity is inhibited both in vitro [15] and in plant nodules [7]. Therefore, the benefit of using Alnus species in the reclamation of saline soils needs to be investigated.
Green alder (Alnus alnobetula subsp. crispa (Aiton) Raus) is a native woody plant widely distributed across the boreal region of North America. Research on its use for the revegetation of disturbed areas has mostly focused on its nitrogen-fixing ability [12]. One field study demonstrated that Frankia-inoculated green alder on oil sands tailings capped with soil substantially increased soil quality, reducing soil pH from 7.5 to 6.6, plant-available sodium content by 70%, and increasing soil organic matter content after two growing seasons [16]. Another study found that nodulated green alder biomass was greater than non-nodulated plants when grown in pure oil sands mixed with sand [4], confirming the effect of the nitrogen-fixing symbiosis on plant performance.
In addition to forming a nitrogen-fixing symbiosis, Alnus species can also be colonized by ectomycorrhizal fungi (ECMF) [9]. Plant association with ECMF can play an important role in increasing nutrient uptake via phosphatase and protease production [17,18]. In nitrogen-fixing plants such as Alnus, symbiosis with ECMF can increase both plant growth and nitrogen fixation [19]. ECMF are also known to alleviate different types of abiotic stress on plants, making them valuable tools in land restoration [20]. Markham [21] showed the dual symbiosis of Alnus incana subsp. rugosa with ECMF (Paxillus involutus) and Frankia increased plant mass when plants grew in a mixture of peat and gold mine tailings with elevated levels of heavy metals. However, the mechanism by which mycorrhizae benefit plants exposed to mine tailings remains unclear. Several reports have described the use of ECMF to reduce salt stress in plants. Nguyen et al. [22] found that NaCl-treated Picea glauca and Picea mariana had reduced shoot chloride levels when inoculated with Hebeloma crustuliniforme or Laccaria bicolor. On the other hand, these ECMF species did not reduce shoot chloride in Pinus banksiana under the same conditions. In salt-stressed aspen (Populus tremuloides), association with H. crustuliniforme resulted in higher plant biomass and root hydraulic conductance than in non-mycorrhizal seedlings [23]. It has been suggested that ECMF could play an important role in preventing Na+ and Cl− translocation to plant tissues and could enhance nutrient uptake, making ECMF a good choice for the remediation of tailings contaminated soil [22,24].
The aim of this study was to determine the effect of both Frankia and ECMF on the performance of green alder under the saline conditions typical of oil sand industrial activity. We predicted that Frankia and ECMF inoculation would have a synergistic effect on plant growth by alleviating the nutrient deficiency associated with salt stress.
2. Results
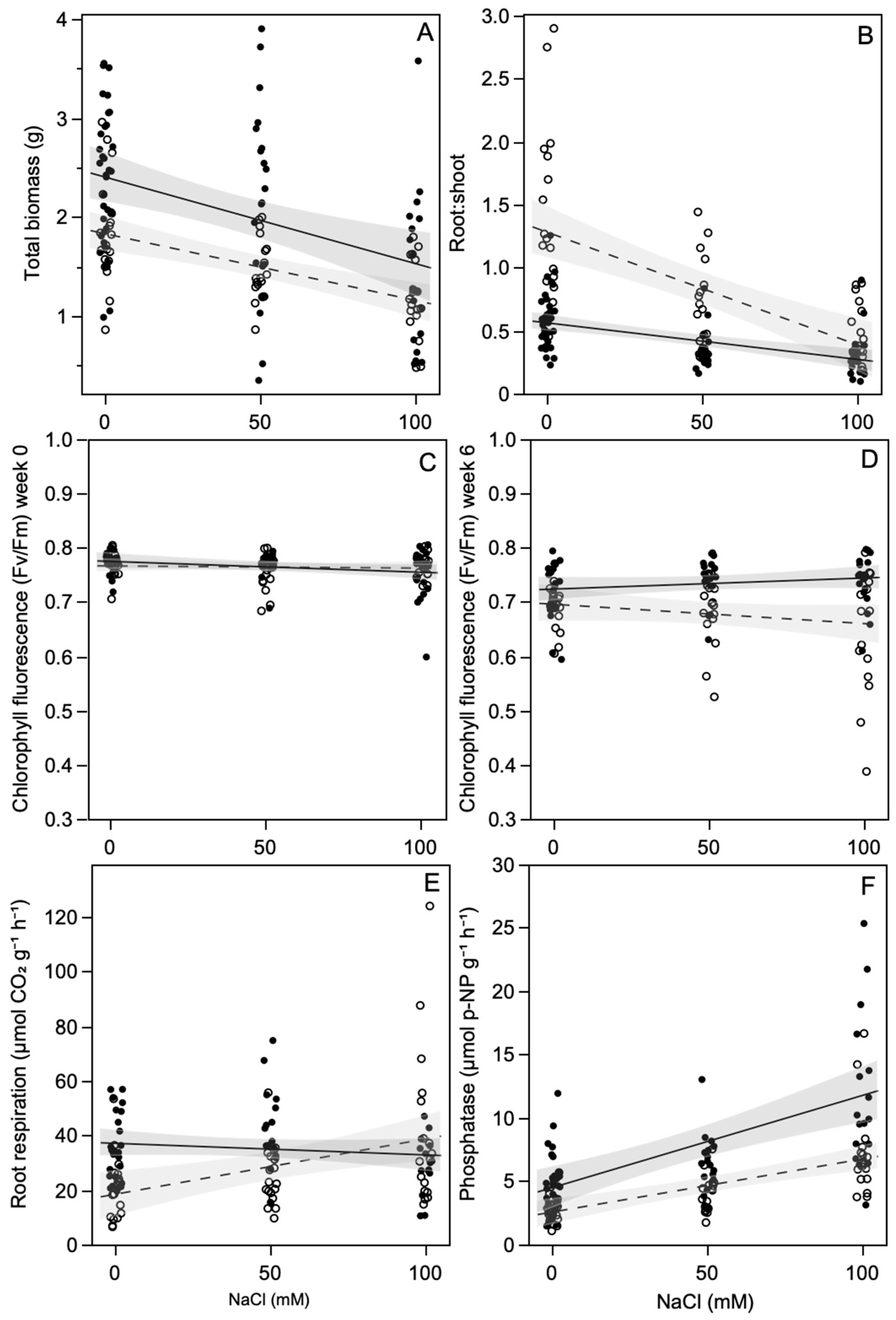
All but one of the ECMF-inoculated plants formed Hartig nets, and all but one of the Frankia-inoculated plants formed nodules visible to the naked eye. Frankia inoculation significantly increased total plant biomass by 37% compared with non-inoculated plants, regardless of mycorrhizal inoculation or salt level, i.e., there was no interaction between these treatments (Figure 1A, Table 1). Increasing NaCl significantly decreased total plant biomass, such that plants exposed to 100 mM NaCl were 40% smaller than plants exposed to no NaCl, regardless of Frankia or ECMF inoculation. Salt exposure had a stronger effect on the root:shoot ratio of non-Frankia-inoculated plants than Frankia-inoculated plants (Figure 1B). In the non-Frankia-inoculated plants, the root:shoot ratio was reduced by 66% in plants exposed to 100 mM NaCl compared to plants exposed to 0 mM NaCl. The Frankia-inoculated plants in the non-salt treatment had a lower root:shoot ratio than the non-inoculated plants, and it was reduced by 33% when the plants were exposed to 100 mM NaCl. Prior to the salt treatments, the chlorophyll fluorescence data showed that the plants were not stressed, and there was no difference between the treatments (Figure 1C). By week six, the maximum quantum yield decreased in all treatments, but the non-Frankia-inoculated plants had a lower maximum quantum yield, which decreased as the salt level increased. In the non-inoculated plants, root respiration (excluding nodules) increased by 42% for plants exposed to 100 mM NaCl compared to plants grown at 0 mM NaCl (Figure 1E). Salt exposure did not affect the respiration of Frankia-inoculated plants, and at 0 mM NaCl, the inoculated plants had a higher rate of root respiration than the non-inoculated plants. Root extracellular acid phosphatase activity increased with increasing salt exposure, with plants exposed to 100 mM NaCl having a 2.3 times higher level than plants not exposed to salt (Figure 1F). Frankia inoculation also increased root phosphatase by 1.5 times compared to non-inoculated plants, regardless of salt exposure. ECMF inoculation did not affect plant mass, root:shoot ratio, root respiration, or root phosphatase levels.
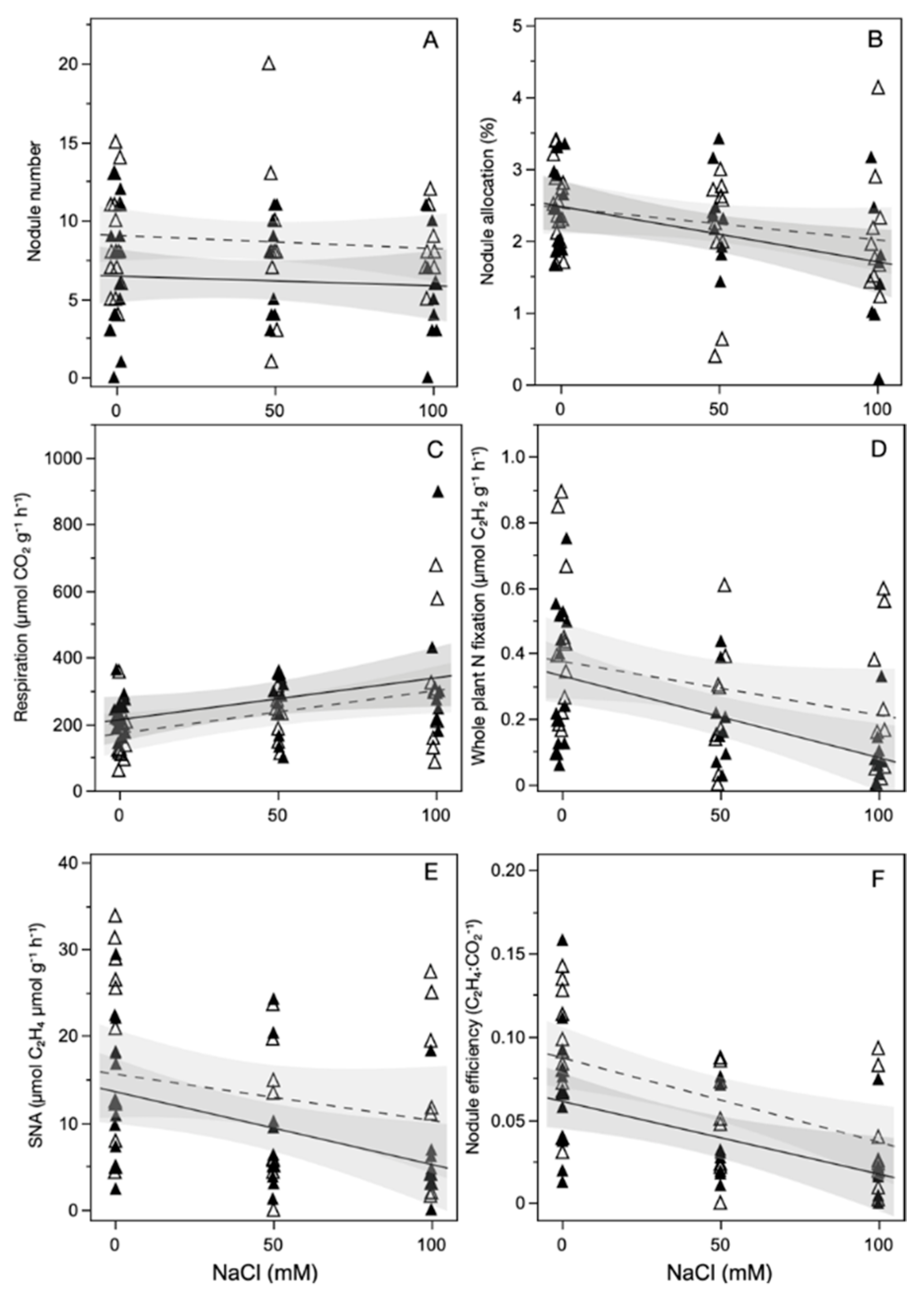
Salt exposure did not affect the number of nodules on a plant at the time of harvest (Figure 2A, Table 2). However, salt exposure reduced all other measures of the nitrogen fixing benefits of Frankia inoculation. Compared to non-salt exposed plants, plants exposed to 100 mM NaCl had a 24% reduction in biomass allocation to nodules, a 64% increase in nodule respiration, and a 42% decrease in specific nodule activity. These measures combined reduced nodule efficiency by 56% and the amount of total nitrogenase activity per plant mass by 54% when plants were exposed to 100 mM NaCl. ECMF inoculation reduced nodule number by 28% and nodule efficiency by 33%, compared to non-inoculated plants, regardless of salt exposure level.
3. Discussion
Our results show that the growth benefit of the nitrogen-fixing symbiosis in A. alnobetula subsp. crispa is not affected by salt exposure (i.e., there was no interaction between the Frankia inoculation and salt treatments on biomass. Frankia inoculation also reduced salt stress in leaves as indicated by the maintenance of the maximum quantum yield of photosynthesis in presence of salts. The increased growth of nitrogen-fixing plants occurred even though salt exposure decreased nitrogen fixation per plant mass and nodule mass and decreased the efficiency of nitrogen fixation in nodules. The decrease in nodule function reported here is consistent with our earlier study on salt exposure in Eleagnus commutata [7], in which we found a decrease in the formation of nitrogen-fixing vesicles in nodules of plants exposed to 100 mM NaCl. This decrease in vesicles could explain the reduced efficiency of nodules that we found in the current study. The lack of an effect of NaCl on nodule number in our plants was expected since plants were inoculated well before salt exposure, such that the plants would have had enough time to form nodules before experiencing any salt stress. Tailings from oil sand extraction have been shown to decrease nodule numbers in A. alnobetula subsp. crispa but not nodule allocation [4]. While nodulated plants benefit from a source of fixed N, they do not need to invest as much of their biomass in the production of roots for N absorption, allowing them to produce relatively more photosynthetic tissue [25]. The increase in root respiration resulting from salt exposure that we found has been observed in many species and has been attributed to the increased ion flux in roots [26]. This increased metabolic cost further reduces the ability of plants with a high root: shoot ratio to grow. Therefore, having N-fixing nodules not only provides plants with a source of N but also reduces root exposure to salt by decreasing the area of roots in contact with the soil.
The higher acid phosphatase activity of Frankia-inoculated plants supports the prediction that N fixation can aid in P acquisition [27,28]. Phosphatase enzymes are N rich, and increased access to N can increase their production. Chodak and Niklinska [29] found soil under Alnus glutinosa had elevated acid phosphatase activity. Conifer-dominated stands with Alnus rubra were also shown to have increased soil P availability and plant tissue P [30]. However, increased phosphatase activity does not seem to be universal among N fixing plants. Png et al. [31] found that an actinorhizal plant (Allocasuarina lehmannian) did not increase phosphatase activity in response to decreasing soil P, but Acacia species did. They concluded that phosphatase activity in response to lower soil P was a phylogenetically constrained trait and not a result of having the nitrogen fixation trait. Our results show that even though salt stress reduced whole plant and nodule nitrogen fixation and the efficiency of nodules, nodulated plants increased root phosphatase activity. Similar to our results, previous studies have reported that salt and water stress can increase phosphatase activity in roots and shoots, presumably due to the reduced nutrient uptake associated with salt or water stress [32,33,34].
The lack of a mycorrhizal effect on plant performance was unexpected, given the low fertility of the soil used in this experiment. Studies on the effect of mycorrhizae on the performance of boreal forest plants under salt stress have been inconsistent. Muhsin and Zwiazek [35] found that white spruce in symbiosis with H. crustuliniforme and exposed to 25 mM NaCl had reduced accumulation of sodium in shoots and roots, increased N and P uptake in mycorrhizal roots, and increased ability for shoots to maintain high rates of transpiration under salt stress. Bois et al. [36] tested three ectomycorrhizal fungal species that were salt tolerant in vitro on greenhouse-grown conifers. The authors found that jack pine (Pinus banksiana L.) inoculated with H. crustuliniforme showed the greatest tolerance to salt stress (200 mM NaCl), while white spruce (Picea glauca) seedlings inoculated with Suillus tomentosus had the best growth even at 200 mM NaCl. However, Nguyen et al. [22] found little effect of H. crustulimiforme in altering the growth of P. glauca, Picea mariana, and P. banksiana when exposed to salt stress. Langenfeld-Heyser et al. [37] found no effect of mycorrhizal on the growth of hybrid poplar exposed to up to 500 mM NaCl.
Similar to our findings, Ba et al. [38] found that mycorrhizae inhibited nodule formation in a legume species (Acacia holosericea). This likely results from ectomycorrhizae reducing root hair density, the site nitrogen-fixing bacterial infection. On the other hand, inoculating Alnus tenufolia with ECMF has been shown to increase nodule mass but not affect nitrogen fixation [19].
Our results suggest that A. alnobetula subsp. crispa is moderately salt tolerant, having a 40% reduction in growth at 100 mM NaCl. A number of actinorhizal plants have been found to have a high level of salt tolerance. Some Casuarina species are halophytes and can grow in 600 mM NaCl [39,40]. In Casuarina equisetifolia, nodule formation is not affected by NaCl up to a concentration of 150 mM [41]. Shepherdia argentea possesses high salt tolerance of up to 400 mM NaCl [42]. Some Elaeagnus species can persist in up to 300 mM NaCl [43,44]. Elaeagnus and Shepherdia have species found in boreal regions and warrant more study for their use in restoration. While it has been argued that N fixing plants could be particularly useful in restoring areas with saline soil, the reduction in N fixation in salt-stressed plants may compromise their utility. However, using N fixing plants may solve the problem of needing to supply nitrogen to the soil without causing nitrogen runoff. Our results also indicate that N fixing plants can potentially improve the P fertility of soils.
4. Materials and Methods
Alnus alnobetula subsp. crispa seeds collected from the Sandilands Provincial Forest (Latitude: 49.3 N, Longitude: 96.1 W) were stratified at 4 °C for two weeks and then sown in sterilized peat moss and perlite (3:1, v/v) and grown under artificial light (T5 HP fluorescent lamps) with a 16-h light: 8-h dark photoperiod. The relative humidity (RH) ranged from 30% to 50%, and the temperature was 25 °C/20 °C (day/night). At four months, we transferred seedlings to 6.25 cm diameter × 25 cm deep pots (D40H Deepots, Stuewe & Sons, Tangent, OR, USA) filled with soil from the forest where we collected the seeds. The soil is 99.8% sand by mass and has an inorganic N content of 10.2 ± 0.6 mg kg−1 (mean ± standard deviation) and phosphate level of 0.98 ± 0.33 mg kg−1 [45]. The soil was autoclaved for one hour. Seedlings were inoculated with Frankia and/or the ECMF species, Hebeloma crustiliniforme, with control plants receiving no inoculation. This resulted in four invocation treatments: 1. no Frankia, no ECMF, 2. Frankia, no ECMF, 3. No Frankia, ECMF and, 4. Frankia and ECMF. The Frankia inoculant consisted of surface-sterilized field-collected crushed nodules at 2 mg mL−1 of PBS Buffer. Each plant was inoculated by injecting 2 mL of the inoculant on the surface of the soil. An H. crustiliniforme culture (UAMH 6064) was obtained from the University of Alberta Mycological Herbarium and was originally isolated from a sporocarp collected under A. alnobetula subsp. crispa. The fungal strains were maintained at 25 °C in the dark for over 2 months on plates containing Modified Melin Norkrans (MMN) agar medium (composition: glucose 2.5 g, malt extract 2.0 g, yeast extract 1.0 g, potassium phosphate monobasic (KH2PO4) 0.5 g, ammonium phosphate dibasic ((NH4)2HPO4) 0.25 g, magnesium sulphate (MgSO4) 0.15 g, calcium chloride (CaCl2) 0.05 g, sodium chloride (NaCl) 0.025 g, ferric chloride (FeCl3) 0.012 g, agar 15 g, per liter distilled water) agar plates [45]. For the ECMF inoculation, three 3.9 mm diameter by ca. 7 mm height MMN agar plugs with active growing hyphae were placed next to the upper roots of the plant. Six weeks after inoculating the plants, the salt treatments were started. The salt concentrations (0, 50 and 100 mM NaCl) selected correspond to the range of concentrations found in the areas impacted by the oil sand industry. Ten plants in each combination of inoculation treatments received one of three salt treatments: 1. no NaCl (25 mL water), 2. two 25 mL doses of 50 mM NaCl (146 mg per pot), and 3. four 25 mL doses of 50 mM NaCl (292 mg per pot), hereafter referred to as 0, 50, or 100 mM salt treatments, respectively. Salt doses were applied once daily until the appropriate salt level was reached. Plants were then watered with distilled water to about saturation every two days for the remainder of the experiment. Each pot had a small cup underneath to collect the leaching solution from the soil, which we poured back onto the soil surface. Plants were exposed to NaCl conditions for seven weeks and then harvested and freeze dried to determine their biomass.
Chlorophyll fluorescence was measured using an Opti-Sciences OS30p chlorophyll fluorometer (Hudson, NH, USA) the week prior to and six weeks after applying the salt treatments. Leaves were dark acclimated for 30 min prior to taking measurements. For the data collected at six weeks, measurements were taken on each plant on two different days, and the values averaged. We used the maximum quantum yield (fv/fm) as an index of plant stress. At the time of harvest, roots with attached nodules were cleaned of soil with room temperature water. We measured whole root respiration (with nodules attached) by sealing the roots in a 50 mL cylinder, pumping air through the cylinder at 0.2 L min−1, and then through a desiccant tube before measuring the CO2 concentration using a CO2 analyzer (Qubit S151 CO2 Analyzer, Kingston, ON, Canada). We then removed the nodules from the plant, and their respiration rate was measured using the same procedure. The difference in CO2 concentrations between the whole root system and nodules was used to calculate root respiration in the absence of nodules. Samples were then freeze dried, and respiration rates were expressed on a dry mass basis.
To examine mycorrhizal formation, pieces of fresh roots were weighed and stored in 70% ethanol. The dry weight of these pieces was estimated using the dry to fresh weight ratio of the remaining roots. Mycorrhizae formation was confirmed by examining up to 10 sections of fine roots at 400× magnification for the presence of a Hartig net. Root sections were pre-selected for microscopic examination by the presence of brown flexuous fine roots or succulent fibrous root tips [46].
Acetylene reduction assays were performed on the nodules following [25]. Nodules were sealed in 50 mL containers with an atmosphere of 10% acetylene for one hour. Ethylene in the headspace was then measured using a gas chromatograph (Varian 4500, Edmonton, AB, Canada) with a gas sampling valve and a Haysep T column connected to an FID. The specific nodule activity (SNA) was expressed as the ethylene production rate per nodule dry mass. Nodule allocation was calculated as the nodule mass as a percentage of the total dry mass. We measured root acid phosphatase activity according to [47]. Briefly, root samples without nodules were incubated in the dark for one hour with 3.5 mM para-nitrophenyl phosphate (p-NPP) with a 100 mM modified universal buffer. The reaction was then stopped with 1M NaOH, and the absorbance of the filtered solution at 410 nm compared to a standard curve of para-nitrophenyl (p-NP). The phosphatase enzyme activity was expressed as µmol p-NP per hour per gram of dried root.
The data were analyzed using a three-factor least squares model with Frankia and ECMF inoculation as nominal effects and NaCl level as a continuous effect, with ten replicates per treatment. The nodule number per plant data was analyzed using a GLM model with a Poisson distribution. When there was a significant interaction between NaCl and the other factors, the effect of salt on the response variable was analyzed using linear regressions for each level of the interacting factor.
Author Contributions
This work was part of a graduate thesis project by H.C., supervised by J.M and S.R. All authors conceived and carried out the study. H.C. wrote the first draft of the manuscript, which was edited by J.M. and S.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed by a University of Manitoba Entrance Scholarship to H.C. and Natural Sciences and Engineering Research Council Discovery Grants S.R. and J.M.
Data Availability Statement
Data are available from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant. Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, S. Tamarack response to salinity: Effects of sodium chloride on growth and ion, pigment, and soluble carbohydrate levels. Can. J. For. Res. 2005, 35, 2806–2812. [Google Scholar] [CrossRef]
- Renault, S.; Qualizza, C.; MacKinnon, M. Suitability of altai wildrye (Elymus angustus) and slender wheatgrass (Agropyron trachycaulum) for initial reclamation of saline composite tailings of oil sands. Environ. Pollut. 2004, 128, 339–349. [Google Scholar] [CrossRef]
- Bissonnette, C.; Fahlman, B.; Peru, K.M.; Khasa, D.P.; Greer, C.W.; Headley, J.V.; Roy, S. Symbiosis with Frankia sp. benefits the establishment of Alnus viridis ssp. crispa and Alnus incana ssp. rugosa in tailings sand from the Canadian oil sands industry. Ecol. Eng. 2014, 68, 167–175. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotox. Environ. Safe. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Shao, J.; Renault, S.; Markham, J. Nitrogen fixation symbiosis and salt tolerance of the boreal woody species Elaeagnus commutata. Physiol. Plant. 2020, 42, 100. [Google Scholar] [CrossRef]
- Chapin III, S.F.; Walker, L.R.; Fastie, C.L.; Sharman, L.C. Mechanisms of primary succession following deglaciation at Glacier Bay, Alaska. Ecol. Monogr. 1994, 64, 149–175. [Google Scholar] [CrossRef]
- Põlme, S.; Bahram, M.; Yamanaka, T.; Nara, K.; Cheng Dai, Y.; Grebenc, T.; Kraigher, H.; Toivonen, M.; Wang, P.-H.; Matsuda, Y.; et al. Biogeography of ectomycorrhizal fungi associated with alders (Alnus spp.) in relation to biotic and abiotic variables at the global scale. New Phytol. 2013, 198, 1239–1249. [Google Scholar] [CrossRef] [Green Version]
- Schwencke, J.; Carú, M. Advances in actinorhizal symbiosis: Host plant-Frankia interactions, biology, and applications in arid land reclamation. Arid Land Res. Manag. 2001, 15, 285–327. [Google Scholar] [CrossRef]
- Vogel, J.G.; Gower, S.T. Carbon and nitrogen dynamics of boreal jack pine stands with and without a green alder understory. Ecosystems 1998, 1, 386–400. [Google Scholar] [CrossRef]
- Perinet, P.; Brouillette, J.G.; Fortin, J.A.; Lalonde, M. Large scale inoculation of actinorhizal plants with Frankia. Plant Soil 1985, 87, 175–183. [Google Scholar] [CrossRef]
- Santi, C.; Bogusz, D.; Franche, C. Biological nitrogen fixation in non-legume plants. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sera, B. Salt-tolerant trees usable for Central European cities—A review. Hortic. Sci. 2017, 44, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Oshone, R.; Mansour, S.; Tisa, L. Effect of salt stress on the physiology of Frankia sp strain CcI6. J. Biosci. 2013, 38, 699–702. [Google Scholar] [CrossRef]
- Lefrancois, E.; Quoreshi, A.; Khasa, D.; Fung, M.; Whyte, L.G.; Roy, S.; Greer, C.W. Field performance of alder-Frankia symbionts for the reclamation of oil sands sites. Appl. Soil Ecol. 2010, 46, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Landeweert, R.; Hoffland, E.; Finlay, R.D.; Kuyper, T.W.; Van Breemen, N. Linking plants to rocks: Ectomycorrhizal fungi mobilize nutrients from minerals. Trends Ecol. Evol. 2001, 16, 248–254. [Google Scholar] [CrossRef]
- Diagne, N.; Thioulouse, J.; Sanguin, H.; Prin, Y.; Krasova-Wade, T.; Sylla, S.; Galiana, A.; Baudoin, E.; Neyra, M.; Svistoonoff, S.; et al. Ectomycorrhizal diversity enhances growth and nitrogen fixation of Acacia mangium seedlings. Soil Biol. Biochem. 2013, 57, 468–476. [Google Scholar] [CrossRef]
- Yamanaka, T.; Li, C.Y.; Bormann, B.T.; Okabe, H. Tripartite associations in an alder: Effects of Frankia and Alpova diplophloeus on the growth, nitrogen fixation and mineral acquisition of Alnus tenuifolia. Plant Soil 2003, 254, 179–186. [Google Scholar] [CrossRef]
- Roy, S.; Khasa, D.; Greer, C.W. Combining alders, frankiae, and mycorrhizae for the revegetation and remediation of contaminated ecosystems. Can. J. Bot. 2007, 85, 237–251. [Google Scholar] [CrossRef]
- Markham, J.H. The effect of Frankia and Paxillus involutus on the performance of Alnus incana spp. rugosa in mine tailings. Can. J. Bot. 2005, 83, 1384–1390. [Google Scholar] [CrossRef]
- Nguyen, H.; Polanco, M.; Zwiazek, J.J. Gas Exchange and Growth Responses of Ectomycorrhizal Picea mariana, Picea glauca, and Pinus banksiana Seedlings to NaCl and Na2SO4. Plant. Biol. 2008, 8, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Polanco, C.; MacKinnon, M.; Zwiazek, J.J. Responses of ectomycorrhizal Populus tremuloides and Betula papyrifera seedlings to salinity. Exp. Bot. 2008, 62, 357–363. [Google Scholar] [CrossRef]
- Meharg, A.A.; Cairney, J.W.G. Ectomycorrhizas—Extending the capabilities of rhizosphere remediation? Soil Biol. Biochem. 2000, 32, 1475–1484. [Google Scholar] [CrossRef]
- Markham, J.H.; Zekveld, C. Nitrogen fixation makes allocation to roots independent of nitrogen supply. Can. J. Bot. 2007, 85, 787–793. [Google Scholar] [CrossRef]
- Bloom, A.; Epstein, E. Varietal differences in salt- induced respiration in barley. Plant. Sci. Lett. 1984, 35, 1–3. [Google Scholar] [CrossRef]
- Houlton, B.Z.; Wang, Y.-P.; Vitousek, P.; Field, C.B. A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 2008, 454, 327–330. [Google Scholar] [CrossRef]
- Wang, Y.-P.; Houlton, B.Z.; Field, C.B. A model of biogeochemical cycles of carbon, nitrogen, and phosphorus including symbiotic nitrogen fixation and phosphatase production. Glob. Biogeochem. Cycles 2007, 21, gb1018. [Google Scholar] [CrossRef]
- Chodak, M.; Niklinska, M. The effect of different tree species on the chemical and microbial properties of reclaimed mine soils. Biol. Fertl. Soils 2010, 46, 555–566. [Google Scholar] [CrossRef]
- Giardina, C.P.; Huffman, S.; Binkley, D.; Caldwell, B.A. Alders increase soil phosphorus availability in a Douglas-fir plantation. Can. J. For. Res. 1995, 25, 1652–1657. [Google Scholar] [CrossRef]
- Png, G.; Turner, B.L.; Albornoz, F.E.; Hayes, P.E.; Lambers, H.; Laliberté, E. Greater root phosphatase activity in nitrogen-fixing rhizobial but not actinorhizal plants with declining phosphorus availability. J. Ecol. 2017, 105, 1246–1255. [Google Scholar] [CrossRef] [Green Version]
- Flasinski, S.; Zamorski, R.; Kotowska, U. The effect of water and salt stresses on the phosphorus content and acid phosphatase activity in oilseed rape. Acta Soc. Bot. Pol. 1989, 58, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Barrett-Lennard, E.G.; Robin, E.G.; Greenway, H. Effect of phosphorus deficiency and water deficit on phosphatase activities from wheat leaves. J. Exp. Bot. 1982, 33, 682–693. [Google Scholar] [CrossRef]
- Sharma, A.; Thakur, M.; Rana, M.; Singh, K. Effect of plant growth hormones and abiotic stresses on germination, growth and phosphatase activities in Sorghum bicolor (L.) Moench seeds. Afr. J. Biotechnol. 2004, 3, 308–312. [Google Scholar] [CrossRef]
- Mushin, T.; Zwiazek, J. Colonization with Hebeloma crustuliniforme increases water conductance and limits shoot sodium uptake in white spruce (Picea glauca) seedlings. Plant Soil 2002, 238, 217–225. [Google Scholar] [CrossRef]
- Bois, G.; Bigras, F.J.; Bertrand, A.; Piche, Y.; Fung, M.Y.P.; Khasa, D.P. Ectomycorrhizal fungi affect the physiological responses of Picea glauca and Pinus banksiana seedlings exposed to an NaCl gradient. Tree Physiol. 2006, 26, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- Langenfeld-Heyser, R.; Gao, J.; Ducic, T.; Tachd, P.; Lu, C.F.; Fritz, E.; Gafur, A.; Polle, A. Paxillus involutus mycorrhiza attenuate NaCl-stress responses in the salt-sensitive hybrid poplar Populus × canescens. Mycorrhiza 2007, 17, 121–131. [Google Scholar] [CrossRef]
- Ba, A.M.; Balaji, B.; Piche, Y. Effect of time of inoculation on in vitro ectomycorrhizal colonization and nodule initiation in Acacia holosericea seedlings. Mycorrhiza 1994, 4, 109–119. [Google Scholar] [CrossRef]
- Ribeiro-Barros, A.I.; Pawlowski, K.; Ramalho, J.C. Mechanisms of salt stress tolerance in Casuarina: A review of recent research. J. For. Res. 2022, 27, 113–116. [Google Scholar] [CrossRef]
- Batista-Santos, P.; Duro, N.; Rodrigues, A.P.; Semedo, J.N.; Alves, P.; da Costa, M.; Graça, I.; Pais, I.P.; Scotti-Campos, P.; Lidon, F.; et al. Is salt stress tolerance in Casuarina glauca Sieb. ex Spreng. associated with its nitrogen- fixing root- nodule symbiosis? An analysis at the photosynthetic level. Plant Physiol. Biochem. 2015, 96, 97–109. [Google Scholar] [CrossRef]
- Tani, C.; Sasakawa, H. Salt tolerance of Casuarina equisetifolia and Frankia Ceq1 strain isolated from the root nodules of C. equisetifolia. Soil Sci. Plant Nutr. 2003, 49, 215–222. [Google Scholar] [CrossRef]
- Qin, J.; Dong, W.; He, K.; Chen, J.; Liu, J.; Wang, Z. Physiological responses to salinity in silver buffaloberry (Shepherdia argentea) introduced to Qinghai high- cold and saline area, China. Photosynthetica 2010, 48, 51–58. [Google Scholar] [CrossRef]
- Khamzina, A.; Lamers, L.P.M.; Vlek, P.L.G. Nitrogen fixation by Elaeagnus angustifolia in the reclamation of degraded croplands of central Asia. Tree Physiol. 2009, 29, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maimmaiti, A.; Yunus, Q.; Iwanaga, F.; Mori, F.; Tanaka, K.; Yamanaka, N. Effects of salinity on growth, photosynthesis, inorganic and organic osmolyte accumulation in Elaeagnus oxycarpa seedlings. Acta Physiol. Plant. 2014, 36, 881–892. [Google Scholar] [CrossRef]
- Chen, H.; Renault, S.; Markham, J. The effect of Frankia and multiple ectomycorrhizal fungal species on Alnus growing in low fertility soil. Symbiosis 2020, 80, 207–215. [Google Scholar] [CrossRef]
- Molina, R.; Myrold, D.D.; Li, C.Y. Root symbioses of red alder: Technological opportunities for enhanced regeneration and soil improvement. In The Biology and Management of Red Alder; Schwintzer, C.R., Tjepkema, J.D., Eds.; Oregon State University Press: Corvallis, OR, USA, 1994; pp. 23–46. [Google Scholar]
- Treseder, K.K.; Vitousek, P. Effects of soil nutrient availability on investment in acquisition of N and P in Hawaiian rain forests. Ecology 2001, 82, 946–954. [Google Scholar] [CrossRef]
Figure 1.
Effect of salt exposure on: (A) total plant mass, (B) root:shoot ratio, leaf chlorophyll fluorescence prior to (C) and six weeks after (D) salt exposure, (E) root respiration excluding nodules and, (F) root extracellular acid phosphatase activity, of Frankia-inoculated (closed symbols, solid line) and non-inoculated plants (open symbols, dashed line) averaged across ECMF inoculation treatments. Lines are least squares fits with 95% confidence intervals.
Figure 1.
Effect of salt exposure on: (A) total plant mass, (B) root:shoot ratio, leaf chlorophyll fluorescence prior to (C) and six weeks after (D) salt exposure, (E) root respiration excluding nodules and, (F) root extracellular acid phosphatase activity, of Frankia-inoculated (closed symbols, solid line) and non-inoculated plants (open symbols, dashed line) averaged across ECMF inoculation treatments. Lines are least squares fits with 95% confidence intervals.

Figure 2.
Effect of salt exposure on: (A) nodule number; (B) total plant biomass allocated to nodules; (C) nodule respiration; (D) nitrogenase activity relative to whole plant mass; (E) specific nodule activity; (F) the efficiency of nitrogenase activity, for plants that were inoculated with both Frankia and ECMF (closed symbols) or Frankia (open symbols).
Figure 2.
Effect of salt exposure on: (A) nodule number; (B) total plant biomass allocated to nodules; (C) nodule respiration; (D) nitrogenase activity relative to whole plant mass; (E) specific nodule activity; (F) the efficiency of nitrogenase activity, for plants that were inoculated with both Frankia and ECMF (closed symbols) or Frankia (open symbols).


{kind=link}
{kind=link}
Table 1.
p values from least squares models of the effect of Frankia, ECMF, and salt exposure on plant performance.
Table 1.
p values from least squares models of the effect of Frankia, ECMF, and salt exposure on plant performance.
| Total Mass | Root: Shoot | Extracellular Phosphatase * | Root Respiration | Chlorophyll Fluorescence | ||
|---|---|---|---|---|---|---|
| Week 0 | Week 6 | |||||
| Frankia | <0.0001 | <0.0001 | <0.0001 | 0.0142 | 0.9729 | <0.0001 |
| ECMF | 0.5392 | 0.5599 | 0.3161 | 0.6995 | 0.2407 | 0.1390 |
| NaCl | <0.0001 | <0.0001 | <0.0001 | 0.0258 | 0.0808 | 0.7787 |
| Frankia × ECMF | 0.5384 | 0.7750 | 0.6225 | 0.6394 | 0.4793 | 0.3789 |
| Frankia × NaCl | 0.4615 | 0.0001 | 0.7563 | 0.0017 | 0.2561 | 0.0268 |
| ECMF × NaCl | 0.9535 | 0.6063 | 0.6772 | 0.3803 | 0.1492 | 0.0603 |
| Frankia × ECMF × NaCl | 0.1458 | 0.2164 | 0.1330 | 0.3955 | 0.2882 | 0.0522 |
* Analysis performed on log transformed data.
Table 2.
p values from least squares models of the effect of ECMF and salt exposure on nodule number, biomass allocation to nodules, nodule respiration, specific nodule activity (SNA), and nodule efficiency.
Table 2.
p values from least squares models of the effect of ECMF and salt exposure on nodule number, biomass allocation to nodules, nodule respiration, specific nodule activity (SNA), and nodule efficiency.
| Nodule Number | Nodule Allocation | Nodule Respiration | SNA | Nodule Efficiency | |
|---|---|---|---|---|---|
| ECMF | <0.0001 | 0.5198 | 0.2023 | 0.1389 | 0.0100 |
| NaCl | 0.3617 | 0.0065 | 0.0015 | 0.0122 | <0.00001 |
| ECMF × NaCl | 0.9911 | 0.4512 | 0.9695 | 0.5681 | 0.7450 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, H.; Renault, S.; Markham, J. The Effect of Frankia and Hebeloma crustiliniforme on Alnus alnobetula subsp. Crispa Growing in Saline Soil. Plants 2022, 11, 1860. https://doi.org/10.3390/plants11141860
AMA Style
Chen H, Renault S, Markham J. The Effect of Frankia and Hebeloma crustiliniforme on Alnus alnobetula subsp. Crispa Growing in Saline Soil. Plants. 2022; 11(14):1860. https://doi.org/10.3390/plants11141860
Chicago/Turabian StyleChen, Haoran, Sylvie Renault, and John Markham. 2022. "The Effect of Frankia and Hebeloma crustiliniforme on Alnus alnobetula subsp. Crispa Growing in Saline Soil" Plants 11, no. 14: 1860. https://doi.org/10.3390/plants11141860
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.