
Distribution of Phenolic Compounds and Antioxidant Activity in Plant Parts and Populations of Seven Underutilized Wild Achillea Species

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Phenolic Profiles of Leaves, Inflorescences, and Stems across Achillea Species
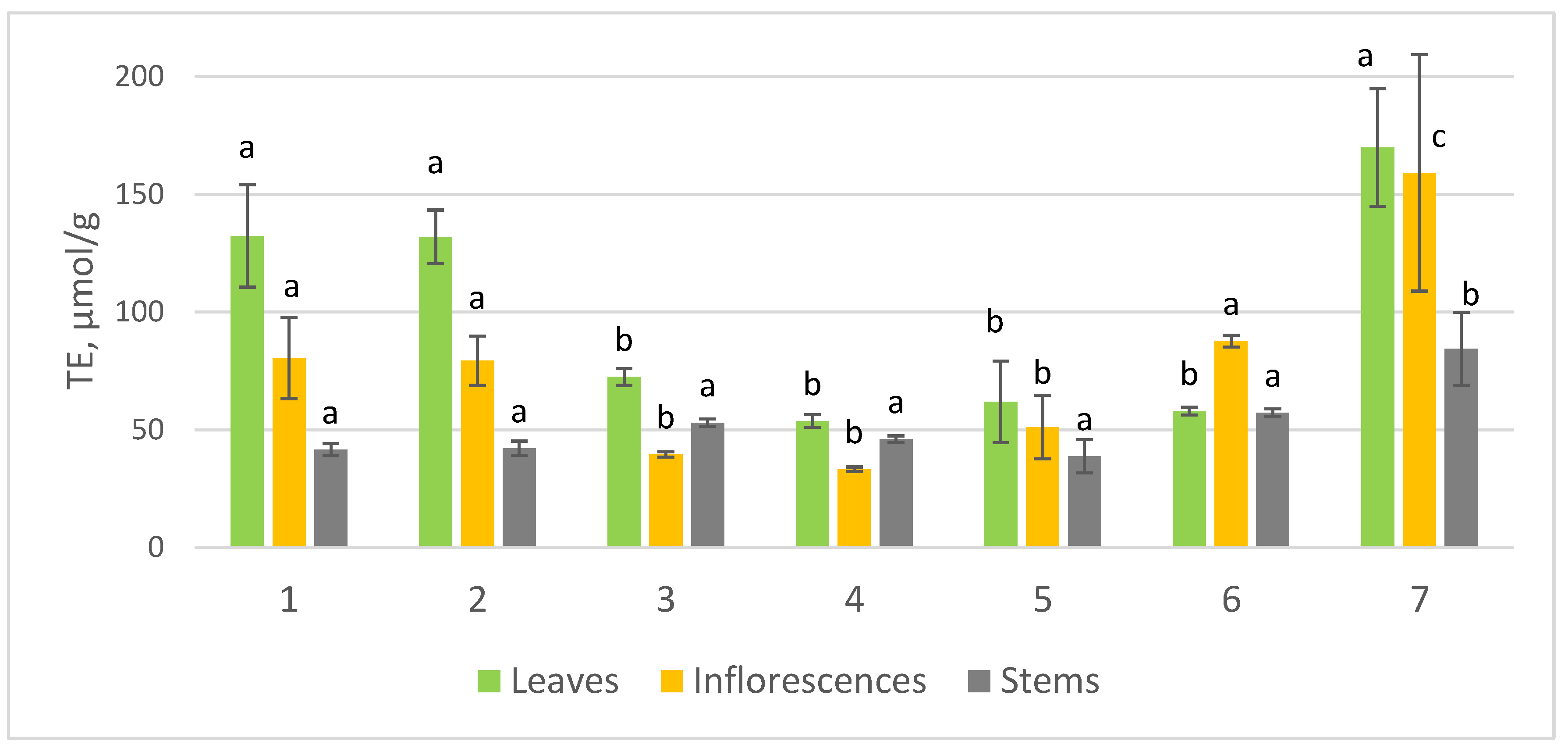
2.2. Antioxidant Activity of Leaves, Inflorescences, and Stems of Wild Achillea Species
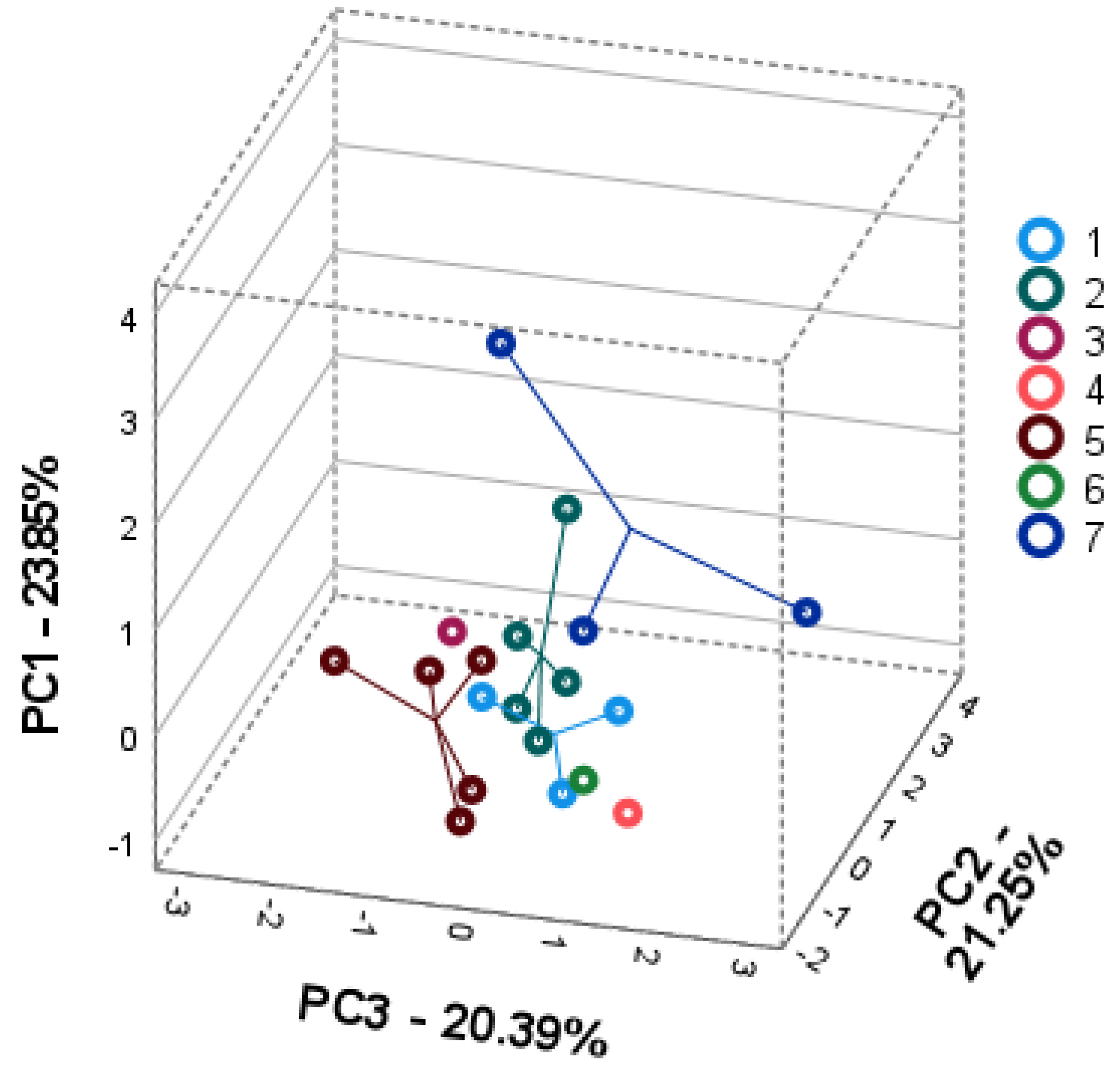
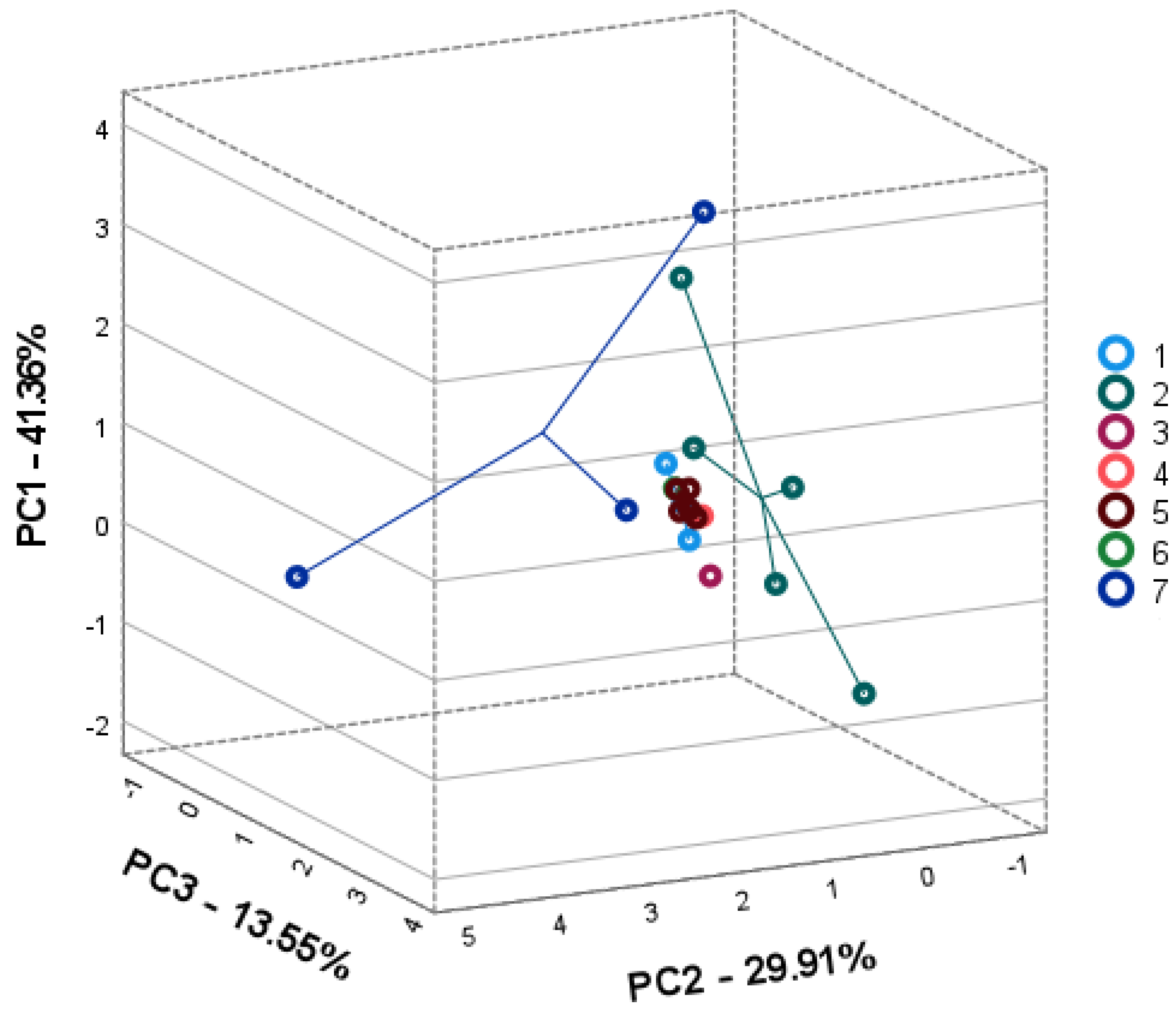
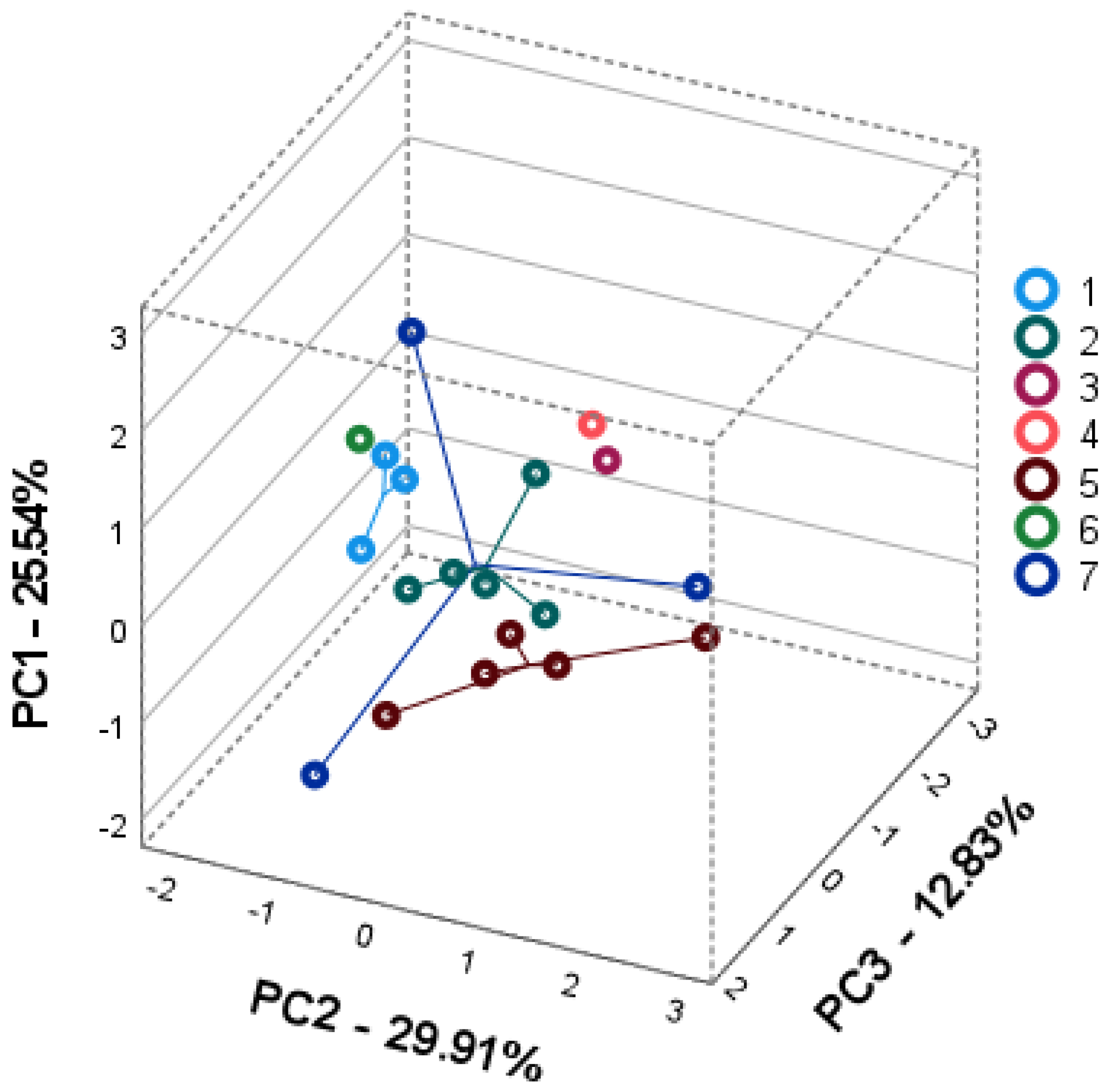
2.3. Intra- and Interspecific Differences; Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material Collection and Identification
4.2. Chemicals
4.3. Sample Preparation
4.4. Antioxidant Activity Assay
4.5. HPLC Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leaman, D.J. Sustainable wild collection of medicinal and aromatic plants Development of an international standard. In Medicinal and Aromatic Plants; Bogers, R.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 97–107. [Google Scholar]
- Ehrendorfer, F.; Guo, Y.-P. Multidisciplinary studies on Achillea sensu lato (Compositae-Anthemideae): New data on systematics and phylogeography. Willdenowia 2006, 36, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Güner, A.; Aslan, S.; Ekim, T.; Vural, M.; Babaç, M. Turkey Plant List (tracheophyta); Türkiye Bitk: Istanbul, Turkey, 2012. [Google Scholar]
- Applequist, W.L.; Moerman, D.E. Yarrow (Achillea millefolium L.): A Neglected Panacea? A Review of Ethnobotany, Bioactivity, and Biomedical Research1. Econ. Bot. 2011, 65, 209–225. [Google Scholar] [CrossRef]
- Honda, G.; Yeşilada, E.; Tabata, M.; Sezik, E.; Fujita, T.; Takeda, Y.; Takaishi, Y.; Tanaka, T. Traditional medicine in Turkey VI. Folk medicine in West Anatolia: Afyon, Kütahya, Denizli, Muğla, Aydin provinces. J. Ethnopharmacol. 1996, 53, 75–87. [Google Scholar] [CrossRef] [PubMed]
- European Pharmacopoeia 10.3. 2021. Available online: https://pheur.edqm.eu/subhome/10-3 (accessed on 13 January 2022).
- Zsuzsanna, R.; Biró-Sándor, Z. Assessment Report on Achillea millefolium L.; Committee on Herbal Medicinal Products (HMPC): Amsterdam, The Netherlands, 2009. [Google Scholar]
- Ali, S.I.; Gopalakrishnan, B.; Venkatesalu, V. Pharmacognosy, Phytochemistry and Pharmacological Properties of Achillea millefolium L.: A Review. Phytother. Res. 2017, 31, 1140–1161. [Google Scholar] [CrossRef] [PubMed]
- Valant-Vetschera, K.M. On the identity of five species of Achillea sect. Millefolium subsect. Filipendulinae (Compositae, Anthemideae). Willdenowia 1999, 29, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Demirel, M.A.; Suntar, I.; Ilhan, M.; Keles, H.; Kupeli Akkol, E. Experimental endometriosis remission in rats treated with Achillea biebersteinii Afan.: Histopathological evaluation and determination of cytokine levels. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 175, 172–177. [Google Scholar] [CrossRef]
- Akkol, E.K.; Koca, U.; Pesin, I.; Yilmazer, D. Evaluation of the Wound Healing Potential of Achillea biebersteinii Afan. (Asteraceae) by In Vivo Excision and Incision Models. Evid. Based. Complement. Alternat. Med. 2011, 2011, 39. [Google Scholar] [CrossRef] [Green Version]
- Hormozi, M.; Baharvand, P. Achillea biebersteinni Afan may inhibit scar formation: In vitro study. Mol. Genet. Genom. Med. 2019, 7, 640. [Google Scholar] [CrossRef] [Green Version]
- Erdogan, M.; Aǧca, C.; Aşkin, H. Achillea biebersteinii extracts suppress angiogenesis and enhance sensitivity to 5-fluorouracil of human colon cancer cells via the PTEN/AKT/mTOR pathway in vitro. Asian Pac. J. Trop. Biomed. 2020, 10, 505. [Google Scholar] [CrossRef]
- Gaweł-Bȩben, K.; Strzȩpek-Gomółka, M.; Czop, M.; Sakipova, Z.; Głowniak, K.; Kukula-Koch, W. Achillea millefolium L. and Achillea biebersteinii Afan. Hydroglycolic Extracts-Bioactive Ingredients for Cosmetic Use. Molecules 2020, 25, 3368. [Google Scholar] [CrossRef]
- The Plant List. Available online: http://www.theplantlist.org (accessed on 1 April 2021).
- Sadeer, N.B.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety—chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Koushki, M.; Farrokhi Yekta, R.; Amiri-Dashatan, N.; Dadpay, M.; Goshadrou, F. Therapeutic effects of hydro-alcoholic extract of Achillea wilhelmsii C. Koch on indomethacin-induced gastric ulcer in rats: A proteomic and metabolomic approach. BMC Complement. Altern. Med. 2019, 19, 2623. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Naderi, G.H.; Sarrafzadegan, N.; Mohammadifard, N.; Mostafavi, S.; Vakili, R. Antihypertensive and antihyperlipidemic effects of Achillea wilhelmsii. Drugs Exp. Clin. Res. 2000, 26, 89–93. [Google Scholar]
- Ashtiani, M.; Nabatchian, F.; Galavi, H.R.; Saravani, R.; Farajian-Mashhadi, F.; Salimi, S. Effect of Achillea wilhelmsii extract on expression of the human telomerase reverse transcriptase mRNA in the PC3 prostate cancer cell line. Biomed. Rep. 2017, 7, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gevrenova, R.; Zengin, G.; Sinan, K.I.; Yıldıztugay, E.; Zheleva-Dimitrova, D.; Picot-Allain, C.; Mahomoodally, M.F.; Imran, M.; Dall’acqua, S. UHPLC-MS Characterization and Biological Insights of Different Solvent Extracts of Two Achillea Species (A. aleppica and A. santolinoides) from Turkey. Antioxidants 2021, 10, 1180. [Google Scholar] [CrossRef]
- Albayrak, S.; Silahtarlıoğlu, N. Determination of biological activities of essential oil and extract obtained from Achillea coarctata Poir. Adv. Tradit. Med. 2020, 20, 77–88. [Google Scholar] [CrossRef]
- Smoylovska, G.P.; Mazulin, O.V.; Abramov, A.V.; Bukhtiyarova, N.V. Pharmacologic and toxicologic properties of lyophilic extract Achillea setacea Waldst. et Kit. Zaporozhye Med. J. 2017, 11, 5308. [Google Scholar] [CrossRef] [Green Version]
- Ertaş, A.; Boǧa, M.; Haşimi, N.; Yeşil, Y.; Gören, A.C.; Topçu, G.; Kolak, U. Antioxidant, anticholinesterase, and antimicrobial activities and fatty acid constituents of Achillea cappadocica Hausskn. et Bornm. Turk. J. Chem. 2014, 38, 592–599. [Google Scholar] [CrossRef]
- Yener, I.; Yilmaz, M.A.; Olmez, O.T.; Akdeniz, M.; Tekin, F.; Hasimi, N.; Alkan, M.H.; Ozturk, M.; Ertas, A. A Detailed Biological and Chemical Investigation of Sixteen Achillea Species’ Essential Oils via Chemometric Approach. Chem. Biodivers. 2020, 17, e1900484. [Google Scholar] [CrossRef] [PubMed]
- Saeidi, K.; Moosavi, M.; Lorigooini, Z.; Maggi, F. Chemical characterization of the essential oil compositions and antioxidant activity from Iranian populations of Achillea wilhelmsii K.Koch. Ind. Crops Prod. 2018, 112, 274–280. [Google Scholar] [CrossRef]
- Bessada, S.M.F.; Barreira, J.C.M.; Oliveira, M.B.P.P. Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind. Crops Prod. 2015, 76, 604–615. [Google Scholar] [CrossRef]
- Verma, R.S.; Joshi, N.; Padalia, R.C.; Goswami, P.; Singh, V.R.; Chauhan, A.; Verma, S.K.; Iqbal, H.; Verma, R.K.; Chanda, D.; et al. Chemical composition and allelopathic, antibacterial, antifungal and in vitro acetylcholinesterase inhibitory activities of yarrow (Achillea millefolium L.) native to India. Ind. Crops Prod. 2017, 104, 144–155. [Google Scholar] [CrossRef]
- Yilmaz, M.A.; Ertas, A.; Yener, I.; Akdeniz, M.; Cakir, O.; Altun, M.; Demirtas, I.; Boga, M.; Temel, H. A comprehensive LC–MS/MS method validation for the quantitative investigation of 37 fingerprint phytochemicals in Achillea species: A detailed examination of A. coarctata and A. monocephala. J. Pharm. Biomed. Anal. 2018, 154, 413–424. [Google Scholar] [CrossRef]
- Varasteh-kojourian, M.; Abrishamchi, P.; Matin, M.M.; Asili, J.; Ejtehadi, H.; Khosravitabar, F.; Antioxidant, K.F. Antioxidant, cytotoxic and DNA protective properties of Achillea eriophora DC. and Achillea biebersteinii Afan. extracts: A comparative study. Avicenna J. Phytomed. 2017, 7, 157. [Google Scholar] [CrossRef]
- Şabanoğlu, S.; Gökbulut, A.; Altun, M.L. Characterization of phenolic compounds, total phenolic content and antioxidant activity of three Achillea species. Marmara Pharm. J. 2019, 23, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Farhadi, N.; Babaei, K.; Farsaraei, S.; Moghaddam, M.; Ghasemi Pirbalouti, A. Changes in essential oil compositions, total phenol, flavonoids and antioxidant capacity of Achillea millefolium at different growth stages. Ind. Crops Prod. 2020, 152, 2750. [Google Scholar] [CrossRef]
- Apel, L.; Lorenz, P.; Urban, S.; Sauer, S.; Spring, O.; Stintzing, F.C.; Kammerer, D.R. Phytochemical characterization of different yarrow species (Achillea sp.) and investigations into their antimicrobial activity. Z. Fur. Naturforsch.-Sect. C J. Biosci. 2021, 76, 55–65. [Google Scholar] [CrossRef]
- Agar, O.T.; Dikmen, M.; Ozturk, N.; Yilmaz, M.A.; Temel, H.; Turkmenoglu, F.P. Comparative Studies on Phenolic Composition, Antioxidant, Wound Healing and Cytotoxic Activities of Selected Achillea L. Species Growing in Turkey. Molecules 2015, 20, 17976. [Google Scholar] [CrossRef] [Green Version]
- Zengin, G.; Aktumsek, A.; Ceylan, R.; Uysal, S.; Mocan, A.; Guler, G.O.; Mahomoodally, M.F.; Glamočlija, J.; Ćirić, A.; Soković, M. Shedding light on the biological and chemical fingerprints of three Achillea species (A. biebersteinii, A. millefolium and A. teretifolia). Food Funct. 2017, 8, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Huyut, Z.; Beydemir, Ş.; Gülçin, I. Antioxidant and antiradical properties of selected flavonoids and phenolic compounds. Biochem. Res. Int. 2017, 2017, 6791. [Google Scholar] [CrossRef] [PubMed]
- Alcázar Magaña, A.; Kamimura, N.; Soumyanath, A.; Stevens, J.F.; Maier, C.S. Caffeoylquinic acids: Chemistry, biosynthesis, occurrence, analytical challenges, and bioactivity. Plant J. 2021, 107, 1299–1319. [Google Scholar] [CrossRef]
- Strzępek-Gomółka, M.; Gaweł-Bęben, K.; Angelis, A.; Antosiewicz, B.; Sakipova, Z.; Kozhanova, K.; Głowniak, K.; Kukula-Koch, W. Identification of Mushroom and Murine Tyrosinase Inhibitors from Achillea biebersteinii Afan. Extract. Molecules 2021, 26, 964. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, K.; Xie, H.; Xie, Y.; Li, Y.; Zhao, X.; Jiang, X.; Chen, D. Antioxidant and cytoprotective effects of the Di-O-Caffeoylquinic acid family: The mechanism, structure–activity relationship, and conformational effect. Molecules 2018, 23, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salarbashi, D.; Fazly Bazzaz, B.S.; Sahebkar, A.; Karimkhani, M.M.; Ahmadi, A. Investigation of optimal extraction, antioxidant, and antimicrobial activities of Achillea biebersteinii and A. wilhelmsii. Pharm. Biol. 2012, 50, 1168–1176. [Google Scholar] [CrossRef]
- Salarbashi, D.; Fazly Bazzaz, B.S.; Karimkhani, M.M.; Sabeti Noghabi, Z.; Khanzadeh, F.; Sahebkar, A. Oil stability index and biological activities of Achillea biebersteinii and Achillea wilhelmsii extracts as influenced by various ultrasound intensities. Ind. Crops Prod. 2014, 55, 163–172. [Google Scholar] [CrossRef]
- Bariş, D.; Kizil, M.; Aytekin, C.; Kizil, G.; Yavuz, M.; Çeken, B.; Ertekin, A.S. In Vitro Antimicrobial and Antioxidant Activity of Ethanol Extract of Three Hypericum and Three Achillea Species From Turkey. Int. J. Food Prop. 2011, 14, 339–355. [Google Scholar] [CrossRef]
- Tsimogiannis, D.I.; Oreopoulou, V. Free radical scavenging and antioxidant activity of 5,7,3′,4′-hydroxy-substituted flavonoids. Innov. Food Sci. Emerg. Technol. 2004, 5, 523–528. [Google Scholar] [CrossRef]
- Rutkowska, M.; Olszewska, M.A.; Kolodziejczyk-Czepas, J.; Nowak, P.; Owczarek, A. Sorbus domestica Leaf Extracts and Their Activity Markers: Antioxidant potential and synergy effects in scavenging assays of multiple oxidants. Molecules 2019, 24, 2289. [Google Scholar] [CrossRef] [Green Version]
- Abd-Alla, H.I.; Shalaby, N.M.M.; Hamed, M.A.; El-Rigal, N.S.; Al-Ghamdi, S.N.; Bouajila, J. Phytochemical composition, protective and therapeutic effect on gastric ulcer and α-amylase inhibitory activity of Achillea biebersteinii Afan. Arch. Pharm. Res. 2016, 39, 10–20. [Google Scholar] [CrossRef]
- Omosa, L.K.; Amugune, B.; Ndunda, B.; Milugo, T.K.; Heydenreich, M.; Yenesew, A.; Midiwo, J.O. Antimicrobial flavonoids and diterpenoids from Dodonaea angustifolia. South Afr. J. Bot. 2014, 91, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Williams, C.A.; Greenham, J.; Harborne, J.B. The role of lipophilic and polar flavonoids in the classification of temperate members of the Anthemideae. Biochem. Syst. Ecol. 2001, 29, 929–945. [Google Scholar] [CrossRef]
- Zidorn, C. Guidelines for consistent characterisation and documentation of plant source materials for studies in phytochemistry and phytopharmacology. Phytochemistry 2017, 139, 56–59. [Google Scholar] [CrossRef]
- Padilla-González, G.F.; Frey, M.; Gómez-Zeledón, J.; Da Costa, F.B.; Spring, O. Metabolomic and gene expression approaches reveal the developmental and environmental regulation of the secondary metabolism of yacón (Smallanthus sonchifolius, Asteraceae). Sci. Rep. 2019, 9, 9246. [Google Scholar] [CrossRef]
- Iwanycki Ahlstrand, N.; Havskov Reghev, N.; Markussen, B.; Bruun Hansen, H.C.; Eiriksson, F.F.; Thorsteinsdóttir, M.; Rønsted, N.; Barnes, C.J. Untargeted metabolic profiling reveals geography as the strongest predictor of metabolic phenotypes of a cosmopolitan weed. Ecol. Evol. 2018, 8, 6812–6826. [Google Scholar] [CrossRef]
- Moore, B.D.; Andrew, R.L.; Külheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Zidorn, C. Plant chemophenetics—A new term for plant chemosystematics/plant chemotaxonomy in the macro-molecular era. Phytochemistry 2019, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Arabaci, T. Revision of Achillea L. (Asteraceae) Genus Grown in Turkey; Nonu University, Institute of Sciences and Technology: Malatya, Turkey, 2006. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Raudone, L.; Vilkickyte, G.; Pitkauskaite, L.; Raudonis, R.; Vainoriene, R.; Motiekaityte, V.; Raudone, L.; Vilkickyte, G.; Pitkauskaite, L.; Raudonis, R.; et al. Antioxidant Activities of Vaccinium vitis-idaea L. Leaves within Cultivars and Their Phenolic Compounds. Molecules 2019, 24, 844. [Google Scholar] [CrossRef] [Green Version]
- Vilkickyte, G.; Raudone, L.; Petrikaite, V. Phenolic fractions from vaccinium vitis-idaea l. And their antioxidant and anticancer activities assessment. Antioxidants 2020, 9, 1261. [Google Scholar] [CrossRef]
- EMEA. ICH Topic Q2 (R1), Validation of Analytical Procedures: Text and Methodology; European Medicines Agency: London, UK, 2005. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | A. cappadocica | A. setacea | A. aleppica | A. coarctata | A.santolinoides subsp. wilhelmsii | A. santolinoides | A. arabica | p-Value |
|---|---|---|---|---|---|---|---|---|
| Neochlorogenic acid | 1608.73 ± 156.58 | 728.7 ± 111.15 | 1683.2 ± 84.16 | 378 ± 18.9 | 935.4 ± 216.89 | 519.4 ± 14.99 | 1824.28 ± 1013.49 | 0.21 |
| Chlorogenic acid | 3347.2 ± 944.48 ab1 | 8062.76 ± 1279.67 abc | 10,020.3 ± 501.02 bc | 1973.4 ± 98.67 a | 4871.02 ± 1666.55 ab | 4038.2 ± 116.57 ab | 12,884.98 ± 4288.21 c | <0.05 |
| 4-caffeoylquinic acid | 193.47 ± 46.78 ab | 996.4 ± 368.42 b | 873.1 ± 43.66 ab | 139.6 ± 6.98 a | 508.56 ± 134.31 ab | 231.8 ± 6.69 ab | 619.5 ± 282.72 ab | <0.05 |
| 3,4-dicaffeoylquinic acid | 2020.63 ± 627.73 ab | 3399.46 ± 1211 b | 683.2 ± 34.16 a | 1722.2 ± 86.11 ab | 570.72 ± 177.14 a | 2328.8 ± 67.23 ab | 787.68 ± 583.73 a | <0.05 |
| 3,5-dicaffeoylquinic acid | 7564.07 ± 2067.07 ab | 11,570.64 ± 2836.29 b | 2758.8 ± 137.94 a | 6615.3 ± 330.77 ab | 2451.38 ± 853.18 a | 5012.2 ± 144.69 ab | 4662.28 ± 3081.53 ab | <0.05 |
| 1,5-dicaffeoylquinic acid | 396.03 ± 79.37 b | 21.54 ± 21.54 a | 0 ± 0 a | 750.4 ± 37.52 c | 25.98 ± 16.93 a | 404.7 ± 11.68 b | 114.28 ± 90.28 a | <0.001 |
| 4,5-dicaffeoylquinic acid | 1232.77 ± 469.86 a | 1062.88 ± 454.86 a | 378.8 ± 18.94 a | 6615.3 ± 330.77 b | 299.82 ± 90.24 a | 1210.6 ± 34.95 a | 245.5 ± 141.07 a | <0.001 |
| Caffeic acid | 34.47 ± 6.73 b | 33.56 ± 11.64 b | 0.7 ± 0.04 a | 21.1 ± 1.06 ab | 24.86 ± 6.44 ab | 83.6 ± 2.41 c | 11.58 ± 6.84 ab | <0.001 |
| Quercitrin | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 8904.9 ± 5126.48 b | <0.05 |
| Rutin | 1785.07 ± 476.65 abc | 932.56 ± 531.04 ab | 0 ± 0 a | 3137.4 ± 156.87 c | 130.08 ± 109.22 ab | 1019.3 ± 29.42 ab | 2115.63 ± 1284.52 bc | <0.05 |
| Quercetin | 15.8 ± 7.9 a | 23.34 ± 5.97 ab | 0 ± 0 a | 0 ± 0 a | 19.86 ± 0.66 ab | 59.2 ± 1.71 c | 45.85 ± 15.87 bc | <0.05 |
| Isoquercitrin | 1541.5 ± 783.89 a | 0 ± 0 b | 80.5 ± 4.03 b | 199.8 ± 9.99 bc | 135.72 ± 80.37 bc | 112.1 ± 3.24 bc | 1732.24 ± 1629.62 a | <0.05 |
| Luteolin | 86.43 ± 43.6 b | 129.96 ± 4.9 bc | 0 ± 0 a | 183.3 ± 9.17 c | 114 ± 1.81 bc | 151.8 ± 4.38 bc | 115.68 ± 39.38 bc | <0.001 |
| Luteolin-7-glucoside | 63.4 ± 29.33 a | 127.98 ± 35.1 b | 703.3 ± 35.17 b | 33.8 ± 1.69 a | 453.58 ± 215.59 b | 39.8 ± 1.15 a | 3763.95 ± 3487.18 c | <0.05 |
| Luteolin-7-rutinoside | 40.07 ± 20.03 a | 88.36 ± 37.22 a | 0 ± 0 b | 37.8 ± 1.89 ab | 271.56 ± 137.1 c | 0 ± 0 b | 83.5 ± 57.68 a | <0.05 |
| Luteolin-3,7-diglucoside | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | - |
| Apigenin | 1.23 ± 1.23 | 0 ± 0 | 4 ± 0.2 | 3.3 ± 0.17 | 2.78 ± 2.47 | 0 ± 0 | 1.73 ± 1.73 | 0.45 |
| Apigenin-7-glucoside | 0 ± 0 a | 0 ± 0 a | 216.5 ± 10.83 b | 0 ± 0 a | 158.7 ± 59.39 b | 0 ± 0 a | 0 ± 0 a | <0.001 |
| Santin | 234.7 ± 13.66 ab | 784.02 ± 287.25 b | 0 ± 0 a | 0 ± 0 a | 459.94 ± 141.44 ab | 0 ± 0 a | 267.03 ± 107.43 ab | <0.05 |
| Compounds | A. cappadocica | A. setacea | A. aleppica | A. coarctata | A.santolinoides subsp. wilhelmsii | A. santolinoides | A. arabica | p Value |
|---|---|---|---|---|---|---|---|---|
| Neochlorogenic acid | 1203.93 ± 19.58 | 289.58 ± 37.88 | 230.1 ± 6.64 | 152.1 ± 4.39 | 207.96 ± 33.98 | 228 ± 6.58 | 682.7 ± 422.66 | 0.41 |
| Chlorogenic acid | 1049 ± 176.38 ab | 3249.96 ± 1109.51 b | 1405.8 ± 40.58 ab | 672.4 ± 19.41 a | 839.7 ± 270.28 a | 942.1 ± 27.2 ab | 2438.85 ± 837.56 ab | <0.05 |
| 4-caffeoylquinic acid | 99.5 ± 27.15 a | 621.26 ± 291.17 b | 140.3 ± 4.05 ab | 69.1 ± 1.99 a | 141.6 ± 33.1 ab | 253 ± 7.3 ab | 573.05 ± 423.09 c | <0.05 |
| 3.4-dicaffeoylquinic acid | 589.2 ± 52.04 ab | 1822.62 ± 483.73 b | 456.7 ± 13.18 a | 764.4 ± 22.07 ab | 299.58 ± 56.53 a | 708.4 ± 20.45 sb | 839.8 ± 639.54 ab | <0.05 |
| 3.5-dicaffeoylquinic acid | 2273.47 ± 302.5 ab | 5330.06 ± 1314.69 b | 2102.7 ± 60.7 ab | 3097.6 ± 89.42 ab | 1266.26 ± 329.77 a | 1361.4 ± 39.3 a | 2125.35 ± 1539.41 ab | <0.05 |
| 1.5-dicaffeoylquinic acid | 152.63 ± 77.05 a | 143.48 ± 51.16 a | 63.8 ± 1.84 a | 984.6 ± 28.42 b | 28.32 ± 19.59 a | 104.8 ± 3.03 a | 105.8 ± 90.06 a | <0.001 |
| 4.5-dicaffeoylquinic acid | 295.2 ± 12.65 ab | 456.88 ± 95.26 b | 292.3 ± 8.44 ab | 3097.6 ± 89.42 c | 207.94 ± 41.49 a | 209.7 ± 6.05 a | 242.2 ± 66.74 ab | <0.001 |
| Caffeic acid | 21.27 ± 9.02 b | 0 ± 0 a | 0.1 ± 0 a | 25.8 ± 0.74 b | 6 ± 4.37 a | 49.3 ± 1.42 c | 3.25 ± 3.25 a | <0.001 |
| Quercitrin | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 8487.21 ± 3445.28 b | <0.05 |
| Rutin | 155.93 ± 49.75 ab | 102.1 ± 49.38 a | 0 ± 0 a | 93.6 ± 2.7 a | 10.1 ± 10.1 a | 63.1 ± 1.82 a | 265.08 ± 85.94 b | <0.05 |
| Quercetin | 60.2 ± 16.18 a | 35.58 ± 5.07 a | 40 ± 1.15 a | 40.2 ± 1.16 a | 42.46 ± 8.34 a | 54.6 ± 1.58 a | 203.7 ± 134.08 a | <0.05 |
| Isoquercitrin | 192.63 ± 90.01 ab | 513.58 ± 174.94 b | 149.3 ± 4.31 ab | 24.9 ± 0.72 a | 54.14 ± 22.35 a | 51.6 ± 1.49 a | 182.09 ± 171.89 ab | <0.05 |
| Luteolin | 284.27 ± 167.68 ab | 723.6 ± 176.02 b | 0 ± 0 c | 694.7 ± 20.05 b | 245 ± 34.84 ab | 725.7 ± 20.95 b | 560.05 ± 360.27 ab | <0.05 |
| Luteolin-7-glucoside | 732.4 ± 253.58 a | 1171.72 ± 238.2 a | 75.8 ± 2.19 a | 181.2 ± 5.23 a | 520.06 ± 154.57 a | 978.5 ± 28.25 a | 5646.13 ± 3185.87 b | <0.05 |
| Luteolin-7-rutinoside | 95.77 ± 33.1 a | 156.86 ± 68.58 a | 37.8 ± 1.09 a | 0 ± 0 a | 170.38 ± 46.78 a | 76.9 ± 2.22 a | 217.98 ± 169.76 a | <0.05 |
| Luteolin-3.7-diglucoside | 0 ± 0 a | 531.26 ± 260.77 b | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 596.8 ± 596.8 b | <0.05 |
| Apigenin | 43.6 ± 26.55 a | 47.72 ± 30.84 a | 0 ± 0 b | 2.3 ± 0.07 b | 0 ± 0 b | 29.2 ± 0.84 ab | 6.88 ± 4.88 b | <0.05 |
| Apigenin-7-glucoside | 138 ± 97.09 a | 137.66 ± 80.47 a | 18.7 ± 0.54 b | 0 ± 0 b | 41.3 ± 12.09 b | 99.3 ± 2.87 ab | 13.28 ± 13.28 b | <0.05 |
| Santin | 229.07 ± 10.36 a | 548.8 ± 190.9 b | 0 ± 0 c | 0 ± 0 c | 228.32 ± 16.21 a | 0 ± 0 c | 321.7 ± 90.89 ab | <0.05 |
| Compounds | A. cappadocica | A. setacea | A. aleppica | A. coarctata | A.santolinoides subsp. wilhelmsii | A. santolinoides | A. arabica | p Value |
|---|---|---|---|---|---|---|---|---|
| Neochlorogenic acid | 1198.13 ± 15.83 a | 277.6 ± 56.41 ab | 430.5 ± 12.43 b | 169.6 ± 4.9 a | 264.88 ± 56.6 ab | 223 ± 6.44 ab | 347.18 ± 122.04 ab | <0.05 |
| Chlorogenic acid | 670.97 ± 121.5 a | 1554.64 ± 340.63 b | 1901.4 ± 54.89 b | 554.9 ± 16.02 a | 2044.22 ± 1086.49 b | 1001.4 ± 28.91 b | 1992.4 ± 709.1 b | <0.05 |
| 4-caffeoylquinic acid | 55.03 ± 4.08 a | 354.58 ± 149.37 b | 280.6 ± 8.1 b | 51.1 ± 1.48 a | 252.74 ± 86.63 b | 87.7 ± 2.53 a | 269.23 ± 197.21 b | <0.05 |
| 3.4-dicaffeoylquinic acid | 388.8 ± 94.75 a | 664.18 ± 206.38 b | 361.8 ± 10.44 a | 441.8 ± 12.75 ab | 219.84 ± 63.56 a | 409.4 ± 11.82 ab | 318.25 ± 226.39 a | <0.05 |
| 3.5-dicaffeoylquinic acid | 1557.77 ± 152.48 | 1583.34 ± 360.82 | 1167.4 ± 33.7 | 1916.9 ± 55.34 | 1175.4 ± 423.71 | 1210.6 ± 34.95 | 1165.95 ± 745.73 | 0.84 |
| 1.5-dicaffeoylquinic acid | 169.8 ± 17.51 b | 85.2 ± 10.66 a | 40.2 ± 1.16 a | 524 ± 15.13 c | 102.58 ± 30.18 ab | 105.6 ± 3.05 ab | 54 ± 36.29 a | <0.001 |
| 4.5-dicaffeoylquinic acid | 273.07 ± 30.27 bc | 177.18 ± 42.64 abc | 433.6 ± 12.52 c | 296.4 ± 8.56 bc | 122.96 ± 28.15 a | 211.5 ± 6.11 abc | 131.95 ± 79.11 ab | <0.001 |
| Caffeic acid | 2.47 ± 1.18 a | 8.12 ± 1.01 a | 25.8 ± 0.74 b | 2.6 ± 0.08 a | 8.24 ± 2.26 a | 31.5 ± 0.91 b | 10.85 ± 9.06 a | <0.001 |
| Quercitrin | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 2442.38 ± 1407.88 b | <0.05 |
| Rutin | 204.03 ± 16.38 a | 366.86 ± 219.79 a | 0 ± 0 b | 1732.2 ± 50 c | 46.1 ± 28.38 b | 63.5 ± 1.83 b | 355.8 ± 213.21 a | <0.001 |
| Quercetin | 54.57 ± 5.93 b | 29.84 ± 2.37 a | 24.1 ± 0.7 a | 24.8 ± 0.72 a | 21 ± 0.7 a | 76.6 ± 2.21 c | 47.7 ± 8.31 b | <0.001 |
| Isoquercitrin | 476.67 ± 111.16 bc | 0 ± 0 a | 0 ± 0 a | 0 ± 0 a | 35.2 ± 23.64 a | 720.5 ± 20.8 c | 303.23 ± 303.23 ab | <0.001 |
| Luteolin | 74.03 ± 37.07 | 45.4 ± 27.81 | 129.6 ± 3.74 | 111.7 ± 3.22 | 48.58 ± 30 | 118.5 ± 3.42 | 100.93 ± 34.4 | 0.21 |
| Luteolin-7-glucoside | 6.9 ± 3.86 a | 18.96 ± 4.86 a | 86.4 ± 2.49 ab | 13.5 ± 0.39 a | 197.36 ± 74.13 b | 5.1 ± 0.15 a | 198.55 ± 114.2 b | <0.05 |
| Luteolin-7-rutinoside | 203.87 ± 93.88 a | 135 ± 14.8 ab | 105.1 ± 3.03 ab | 32.9 ± 0.95 b | 46.58 ± 18.63 b | 179.9 ± 5.19 ab | 162.93 ± 90.55 ab | 0.02 |
| Luteolin-3.7-diglucoside | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | - |
| Apigenin | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | - |
| Apigenin-7-glucoside | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | - |
| Santin | 214.9 ± 6.37 a | 247.48 ± 15.54 a | 0 ± 0 b | 0 ± 0 b | 243.72 ± 22.41 a | 0 ± 0 b | 223.08 ± 5.52 a | <0.001 |
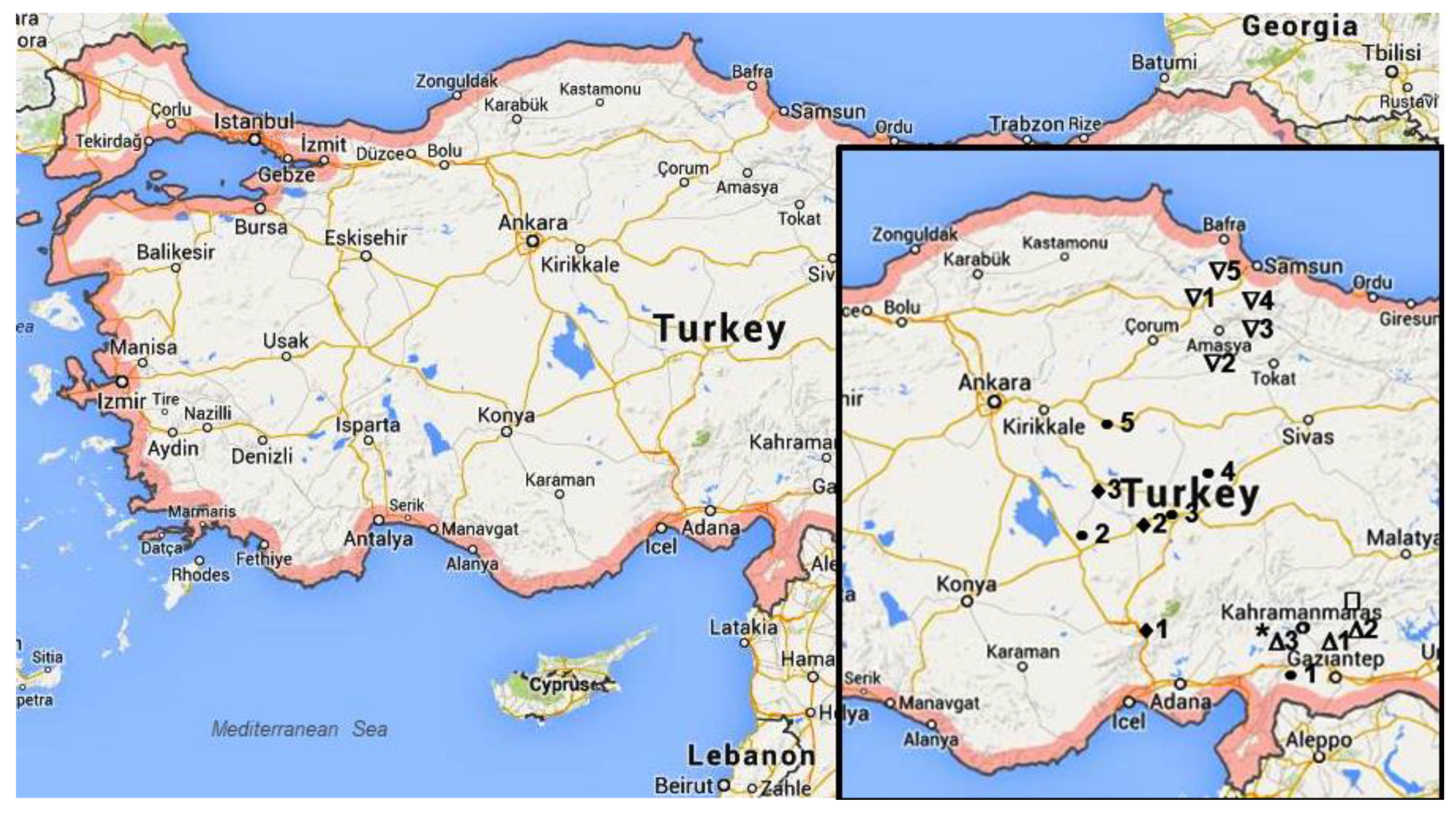
| Species | Population | Voucher Number | Province, Geographical Region | Latitude (°N) | Longitude (°E) | Elevation (m a.s.l.) | Habitat |
|---|---|---|---|---|---|---|---|
| A. cappadocica | 1 | BMYO #AC1 | Niğde, Southern Anatolia | 37. 54 | 34.54 | 2057 | Conifer woodland |
| 2 | BMYO #AC2 | Nevşehir, Central Anatolia | 38.28 | 34.40 | 1596 | Highland meadow | |
| 3 | BMYO #AC3 | Nevşehir, Central Anatolia | 38.39 | 34.29 | 937 | Highland meadow | |
| A. setacea | 1 | BMYO #AS1 | Çorum, Black Sea | 40.53 | 35.13 | 1012 | Barren mountain slope |
| 2 | BMYO #AS2 | Amasya, Black Sea | 40.47 | 35.25 | 914 | Barren mountain slope | |
| 3 | BMYO #AS3 | Çorum, Black Sea | 40.56 | 35.39 | 1197 | Barren mountain slope | |
| 4 | BMYO #AS4 | Amasya, Black Sea | 41.04 | 42.13 | 1117 | Stony roadside | |
| 5 | BMYO #AS5 | Samsun, Black Sea | 41.09 | 35.11 | 664 | Stony roadside | |
| A. aleppica | 1 | BMYO #Aa1 | Gaziantep, Southeastern Anatolia | 37.09 | 37.24 | 668 | Calcareous mountainside |
| A. coarctata | 1 | BMYO#Aa2 | Gaziantep, Southeastern Anatolia | 37.16 | 37.46 | 446 | Stony roadside |
| A.santolinoides subsp. wilhelmsii | 1 | BMYO #AW1 | Gaziantep, Southeastern Anatolia | 36.52 | 36.59 | 1054 | High altitude stony land |
| 2 | BMYO #AW2 | Niğde, Central Anatolia | 38.21 | 34.22 | 1384 | Stony calcareous areas | |
| 3 | BMYO #AW3 | Niğde, Central Anatolia | 38.25 | 34.51 | 1726 | Stony calcareous area | |
| 4 | BMYO #AW4 | Niğde, Central Anatolia | 38.27 | 34°.59 | 1661 | Calcareous stony areas | |
| 5 | BMYO #AW5 | Nevşehir, Central Anatolia | 39.38 | 35.54 | 1139 | Calcareous stony area | |
| A. santolinoides | 1 | BMYO #Aa3 | Gaziantep, Southeastern Anatolia | 37.20 | 37.03 | 520 | Stony roadside |
| A. arabica | 1 | BMYO * #AA54 | Kahramanmaraş, Southeastern Anatolia | 36.58 | 37.24 | 975 | Conifer woodland |
| 2 | BMYO # AA55 | Gaziantep, South-eastern Anatolia | 37.06 | 37.38 | 682 | Conifer woodland | |
| 3 | BMYO # AA56 | Gaziantep, Southeastern Anatolia | 37.01 | 37.06 | 1276 | Conifer woodland |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raudone, L.; Radušiene, J.; Seyis, F.; Yayla, F.; Vilkickyte, G.; Marksa, M.; Ivanauskas, L.; Cırak, C. Distribution of Phenolic Compounds and Antioxidant Activity in Plant Parts and Populations of Seven Underutilized Wild Achillea Species. Plants 2022, 11, 447. https://doi.org/10.3390/plants11030447
Raudone L, Radušiene J, Seyis F, Yayla F, Vilkickyte G, Marksa M, Ivanauskas L, Cırak C. Distribution of Phenolic Compounds and Antioxidant Activity in Plant Parts and Populations of Seven Underutilized Wild Achillea Species. Plants. 2022; 11(3):447. https://doi.org/10.3390/plants11030447
Chicago/Turabian StyleRaudone, Lina, Jolita Radušiene, Fatih Seyis, Fatih Yayla, Gabrielė Vilkickyte, Mindaugas Marksa, Liudas Ivanauskas, and Cüneyt Cırak. 2022. "Distribution of Phenolic Compounds and Antioxidant Activity in Plant Parts and Populations of Seven Underutilized Wild Achillea Species" Plants 11, no. 3: 447. https://doi.org/10.3390/plants11030447