
Genome-Wide Characterization of Sedum plumbizincicola HMA Gene Family Provides Functional Implications in Cadmium Response

 and
and
Abstract
:1. Introduction
2. Results
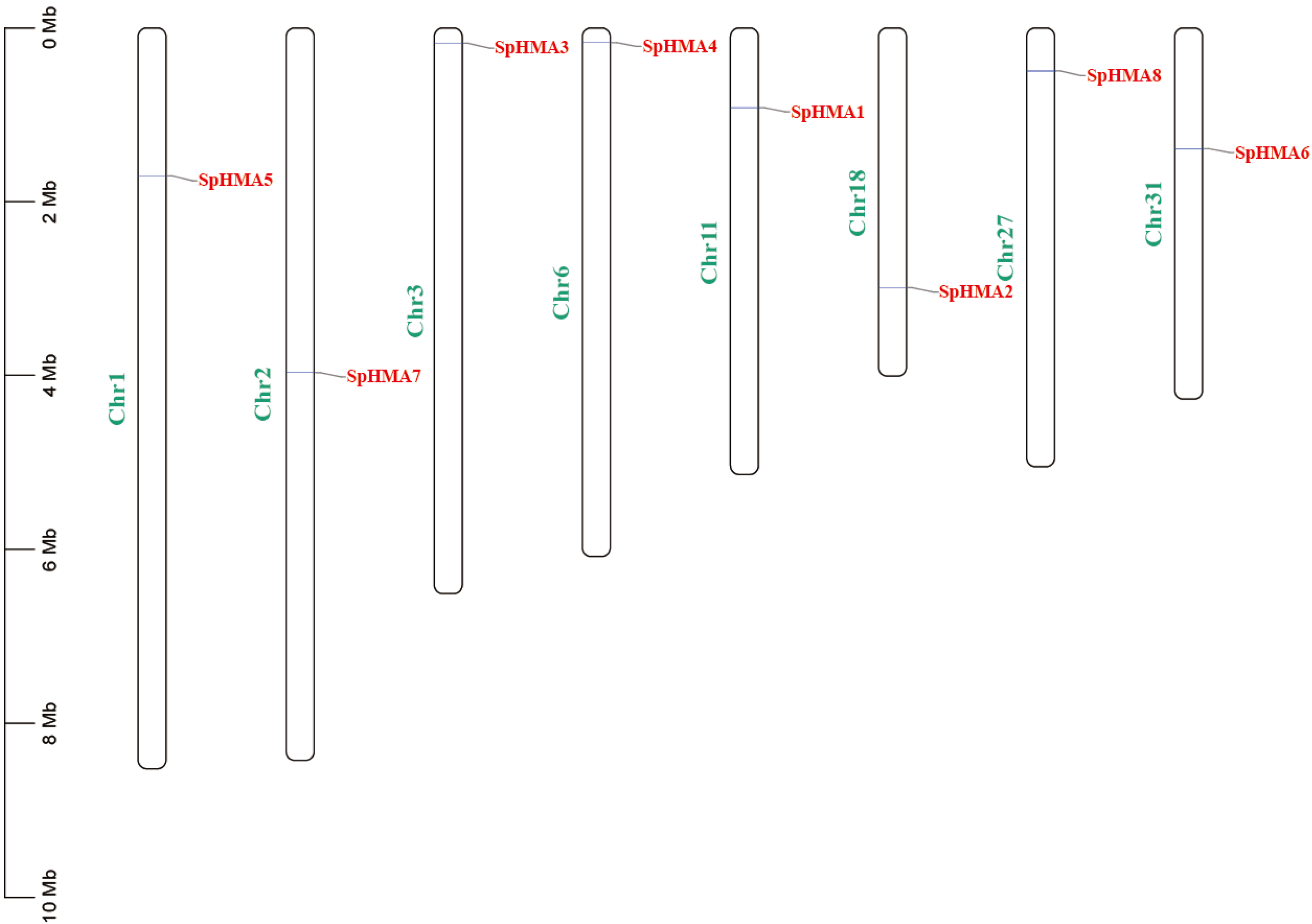
2.1. Genome-Wide Analysis of the SpHMAs
2.2. Phylogenetic and Classification of the HMA Gene Family
2.3. Conserved Motifs and Gene Structure of SpHMAs
2.4. Cis-Acting Elements in the Promoter Regions of SpHMA Genes
2.5. Co-Expression Analysis of the HMA Genes in S. plumbizincicola
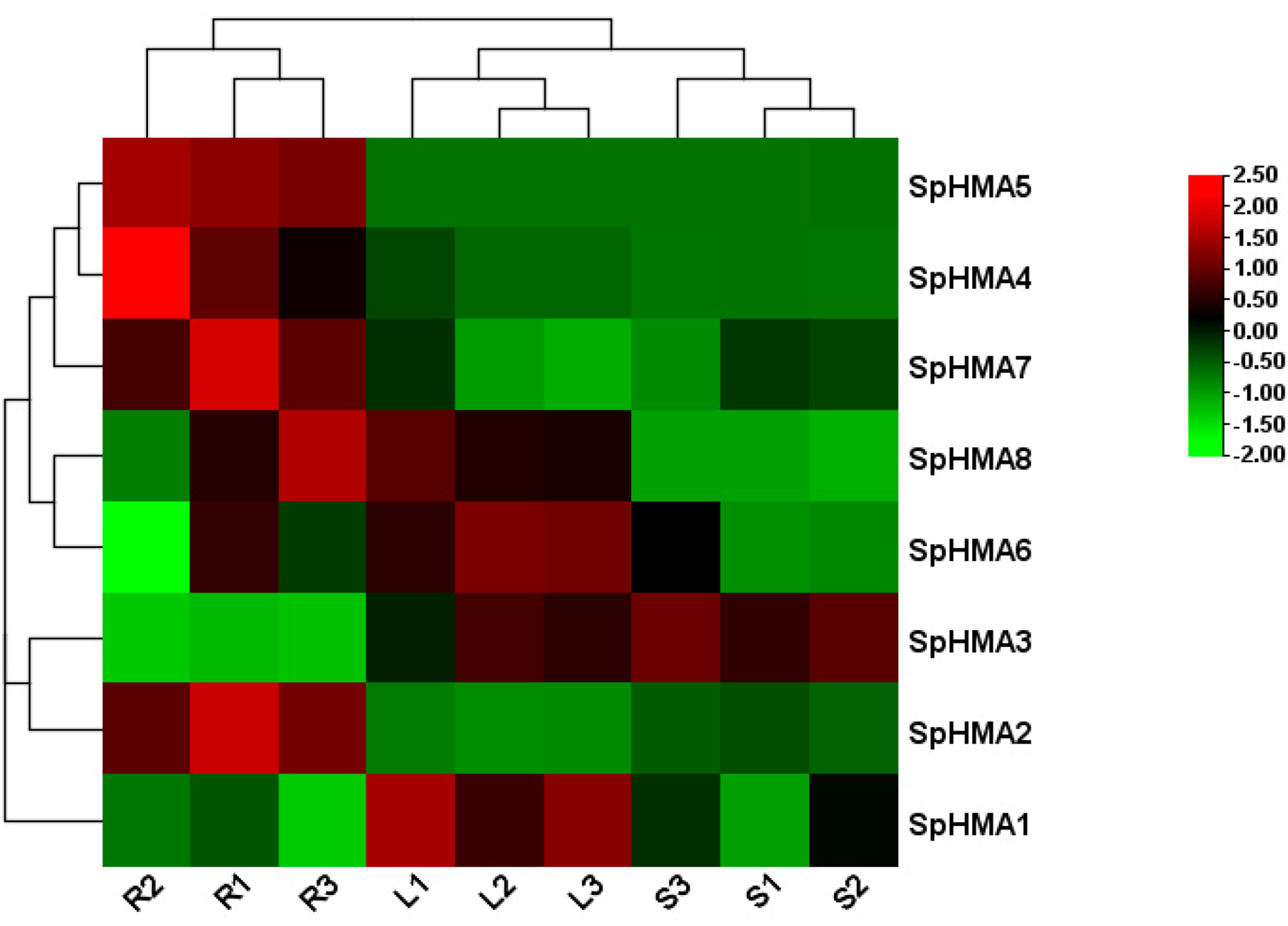
2.6. Expression Profiles of SpHMA Genes in Various Tissues
2.7. qRT-PCR Expression Analysis of the SpHMAs
2.8. Heterologous Expression of SpHMA7 in Yeast
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials and Treatments
5.2. Identification and Sequence Analysis of HMA Genes in S. plumbizincicola
5.3. Exon-Intron Structure, Motif Analysis and Promoter Element Analysis
5.4. Phylogenetic Analysis
5.5. RNA Isolation and Expression Analysis
5.6. Co-Expression Network Construction
5.7. Cloning and Construction of Expression Vectors
5.8. Yeast Expression Experiment
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Adimalla, N.; Chen, J.; Qian, H. Spatial characteristics of heavy metal contamination and potential human health risk assessment of urban soils: A case study from an urban region of South India. Ecotoxicol. Environ. Saf. 2020, 194, 110406. [Google Scholar] [CrossRef]
- Baker, A.J.; Brooks, R. Terrestrial higher plants which hyperaccumulate metallic elements. A review of their distribution, ecology and phytochemistry. Biorecovery 1989, 1, 81–126. [Google Scholar]
- Pedron, T.; Segura, F.R.; da Silva, F.F.; de Souza, A.L.; Maltez, H.F.; Batista, B.L. Essential and non-essential elements in Brazilian infant food and other rice-based products frequently consumed by children and celiac population. J. Food Compos. Anal. 2016, 49, 78–86. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E. Trophic transfer, bioaccumulation, and biomagnification of non-essential hazardous heavy metals and metalloids in food chains/webs—Concepts and implications for wildlife and human health. Hum. Ecol. Risk Assess. 2019, 25, 1353–1376. [Google Scholar] [CrossRef]
- Halder, D.; Saha, J.K.; Biswas, A. Accumulation of essential and non-essential trace elements in rice grain: Possible health impacts on rice consumers in West Bengal, India. Sci. Total Environ. 2020, 706, 135944. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Hao, X.; Lei, M.; Tie, B. The remediation technology and remediation practice of heavy metals-contaminated soil. J. Agro-Environ. Sci. 2013, 32, 409–417. [Google Scholar]
- Cunningham, S.D.; Ow, D.W. Promises and Prospects of Phytoremediation. Plant Physiol. 1996, 110, 715–719. [Google Scholar] [CrossRef] [PubMed]
- Awa, S.H.; Hadibarata, T. Removal of heavy metals in contaminated soil by phytoremediation mechanism: A review. Water Air Soil Pollut. 2020, 231, 1–15. [Google Scholar] [CrossRef]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation in hyperaccumulator plants. Physiol. Plant. 2021, 173, 148–166. [Google Scholar] [CrossRef]
- Verbruggen, N.; Juraniec, M.; Baliardini, C.; Meyer, C.-L. Tolerance to cadmium in plants: The special case of hyperaccumulators. BioMetals 2013, 26, 633–638. [Google Scholar] [CrossRef]
- Wu, L.H.; Liu, Y.J.; Zhou, S.B.; Guo, F.G.; Bi, D.; Guo, X.H.; Baker, A.J.M.; Smith, J.A.C.; Luo, Y.M. Sedum plumbizincicola X.H. Guo et S.B. Zhou ex L.H. Wu (Crassulaceae): A new species from Zhejiang Province, China. Plant Syst. Evol. 2013, 299, 487–498. [Google Scholar] [CrossRef]
- Song, W.; Wang, J.; Zhai, L.; Ge, L.; Hao, S.; Shi, L.; Lian, C.; Chen, C.; Shen, Z.; Chen, Y. A meta-analysis about the accumulation of heavy metals uptake by Sedum alfredii and Sedum plumbizincicola in contaminated soil. Int. J. Phytoremed. 2021. [Google Scholar] [CrossRef]
- Sun, L.; Cao, X.; Tan, C.; Deng, Y.; Cai, R.; Peng, X.; Bai, J. Analysis of the effect of cadmium stress on root exudates of Sedum plumbizincicola based on metabolomics. Ecotoxicol. Environ. Saf. 2020, 205, 111152. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Dong, M.; Mao, P.; Zhuang, P.; Paz-Ferreiro, J.; Li, Y.; Li, Y.; Hu, X.; Netherway, P.; Li, Z. Evaluation of phytoremediation potential of five Cd (hyper) accumulators in two Cd contaminated soils. Sci. Total Environ. 2020, 721, 137581. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef]
- Cao, D.; Zhang, H.; Wang, Y.; Zheng, L. Accumulation and Distribution Characteristics of Zinc and Cadmium in the Hyperaccumulator Plant Sedum plumbizincicola. Bull. Environ. Contam. Toxicol. 2014, 93, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhao, H.; Wu, L.; Liu, A.; Zhao, F.J.; Xu, W. Heavy metal ATPase 3 (HMA3) confers cadmium hypertolerance on the cadmium/zinc hyperaccumulator Sedum plumbizincicola. New Phytol. 2017, 215, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Li, L.-Z.; Tu, C.; Wu, L.-H.; Peijnenburg, W.J.G.M.; Ebbs, S.; Luo, Y.-M. Pathways of root uptake and membrane transport of Cd2+ in the zinc/cadmium hyperaccumulating plant Sedum plumbizincicola. Environ. Toxicol. Chem. 2017, 36, 1038–1046. [Google Scholar] [CrossRef]
- Fasani, E.; Manara, A.; Martini, F.; Furini, A.; DalCorso, G. The potential of genetic engineering of plants for the remediation of soils contaminated with heavy metals. Plant Cell Environ. 2018, 41, 1201–1232. [Google Scholar] [CrossRef] [PubMed]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef]
- Schat, H.; Llugany, M.; Bernhard, R. Metal-specific patterns of tolerance, uptake, and transport of heavy metals in hyperaccumulating and nonhyperaccumulating metallophytes. In Phytoremediation of Contaminated Soil and Water; CRC Press: Boca Raton, FL, USA, 2020; pp. 171–188. [Google Scholar]
- Zeng, F.; Chen, G.; Chen, X.; Li, Q.; Wu, X.; Zhang, G.-P. Cadmium accumulation in plants: Insights from physiological/molecular mechanisms to evolutionary biology. Authorea Preprint 2020. [Google Scholar] [CrossRef]
- Williams, L.E.; Mills, R.F. P1B-ATPases—An ancient family of transition metal pumps with diverse functions in plants. Trends Plant Sci. 2005, 10, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Axelsen, K.B.; Palmgren, M.G. Evolution of substrate specificities in the P-type ATPase superfamily. J. Mol. Evol. 1998, 46, 84–101. [Google Scholar] [CrossRef]
- Smith, A.T.; Smith, K.P.; Rosenzweig, A.C. Diversity of the metal-transporting P 1B-type ATPases. J. Biol. Inorg. Chem. 2014, 19, 947–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelsen, K.B.; Palmgren, M.G. Inventory of the superfamily of P-type ion pumps in Arabidopsis. Plant Physiol. 2001, 126, 696–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.Y.; Choi, H.; Segami, S.; Cho, H.T.; Martinoia, E.; Maeshima, M.; Lee, Y. AtHMA1 contributes to the detoxification of excess Zn (II) in Arabidopsis. Plant J. 2009, 58, 737–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, M.; Crouzet, J.; Gravot, A.; Auroy, P.; Leonhardt, N.; Vavasseur, A.; Richaud, P. AtHMA3, a P1B-ATPase allowing Cd/Zn/co/Pb vacuolar storage in Arabidopsis. Plant Physiol. 2009, 149, 894–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, R.F.; Francini, A.; da Rocha, P.S.F.; Baccarini, P.J.; Aylett, M.; Krijger, G.C.; Williams, L.E. The plant P1B-type ATPase AtHMA4 transports Zn and Cd and plays a role in detoxification of transition metals supplied at elevated levels. FEBS Lett. 2005, 579, 783–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verret, F.; Gravot, A.; Auroy, P.; Leonhardt, N.; David, P.; Nussaume, L.; Vavasseur, A.; Richaud, P. Overexpression of AtHMA4 enhances root-to-shoot translocation of zinc and cadmium and plant metal tolerance. FEBS Lett. 2004, 576, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Ueno, D.; Yamaji, N.; Kono, I.; Huang, C.F.; Ando, T.; Yano, M.; Ma, J.F. Gene limiting cadmium accumulation in rice. Proc. Natl. Acad. Sci. USA 2010, 107, 16500–16505. [Google Scholar] [CrossRef] [Green Version]
- Miyadate, H.; Adachi, S.; Hiraizumi, A.; Tezuka, K.; Nakazawa, N.; Kawamoto, T.; Katou, K.; Kodama, I.; Sakurai, K.; Takahashi, H. OsHMA3, a P1B-type of ATPase affects root-to-shoot cadmium translocation in rice by mediating efflux into vacuoles. New Phytol. 2011, 189, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Wang, L.; Deng, X.; Wang, P.; Ma, Q.; Nian, H.; Wang, Y.; Yang, C. Genome-wide characterization of soybean P 1B-ATPases gene family provides functional implications in cadmium responses. BMC Genom. 2016, 17, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Xu, X.; Hu, X.; Liu, Q.; Wang, Z.; Zhang, H.; Wang, H.; Wei, M.; Wang, H.; Liu, H. Genome-wide analysis and heavy metal-induced expression profiling of the HMA gene family in Populus trichocarpa. Front. Plant Sci. 2015, 6, 1149. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xiao, H.; Sun, J.; Wang, S.; Wang, J.; Chang, P.; Zhou, X.; Lei, B.; Lu, K.; Luo, F. Genome-wide analysis and expression profiling of the HMA gene family in Brassica napus under cd stress. Plant Soil 2018, 426, 365–381. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, L.; Zhao, F.J.; Wu, L.; Liu, A.; Xu, W. SpHMA1 is a chloroplast cadmium exporter protecting photochemical reactions in the Cd hyperaccumulator Sedum plumbizincicola. Plant Cell Environ. 2019, 42, 1112–1124. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Qiu, W.; Li, Y.; Tan, J.; Han, X.; Wu, L.; Jiang, Y.; Deng, Z.; Wu, C.; Zhuo, R. Quantitative proteome analysis reveals changes of membrane transport proteins in Sedum plumbizincicola under cadmium stress. Chemosphere 2022, 287, 132302. [Google Scholar] [CrossRef]
- Van Dam, S.; Vosa, U.; van der Graaf, A.; Franke, L.; de Magalhaes, J.P. Gene co-expression analysis for functional classification and gene–disease predictions. Brief Bioinform. 2018, 19, 575–592. [Google Scholar] [CrossRef]
- Nosek, M.; Kaczmarczyk, A.; Jędrzejczyk, R.J.; Supel, P.; Kaszycki, P.; Miszalski, Z. Expression of genes involved in heavy metal trafficking in plants exposed to salinity stress and elevated Cd concentrations. Plants 2020, 9, 475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Qiu, W.; Liu, W.; Han, X.; Wu, L.; Yu, M.; Qiu, X.; He, Z.; Li, H.; Zhuo, R. Genome-wide characterization of the hyperaccumulator Sedum alfredii F-box family under cadmium stress. Sci. Rep. 2021, 11, 3023. [Google Scholar] [CrossRef]
- Bal, N.; Wu, C.C.; Catty, P.; Guillain, F.; Mintz, E. Cd2+ and the N-terminal metal-binding domain protect the putative membranous CPC motif of the Cd2+-ATPase of Listeria monocytogenes. Biochem. J. 2003, 369, 681–685. [Google Scholar] [CrossRef]
- Yu, C.-S.; Chen, Y.-C.; Lu, C.-H.; Hwang, J.-K. Prediction of protein subcellular localization. Proteins 2006, 64, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Martinoia, E.; Lee, Y. Vacuolar transporters for cadmium and arsenic in plants and their applications in phytoremediation and crop development. Plant Cell Physiol. 2018, 59, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Hu, P.; Li, Z.; Xu, W.; Zhou, T.; Zhong, D.; Luo, Y. Element case studies: Cadmium and zinc. In Agromining: Farming for Metals; Springer: Berlin/Heidelberg, Germany, 2021; pp. 453–469. [Google Scholar]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef] [PubMed]
- Haque, A.M.; Gohari, G.; EI-Shehawi, A.M.; Dutta, A.K.; Elseehy, M.M.; Kabir, A.H. Genome wide identification, characterization and expression profiles of heavy metal ATPase3 (HMA3) in plants. J. King Saud Univ. Sci. 2022, 34, 101730. [Google Scholar] [CrossRef]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 transporter is involved in root-to-shoot translocation of Zn and Cd in rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Chaudhary, K.; Jan, S.; Khan, S. Heavy metal ATPase (HMA2, HMA3, and HMA4) genes in hyperaccumulation mechanism of heavy metals. In Plant Metal Interaction; Elsevier: Amsterdam, The Netherlands, 2016; pp. 545–556. [Google Scholar]
- Abdel-Ghany, S.E.; Müller-Moulé, P.; Niyogi, K.K.; Pilon, M.; Shikanai, T. Two P-type ATPases are required for copper delivery in Arabidopsis thaliana chloroplasts. Plant Cell 2005, 17, 1233–1251. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 357–359. [Google Scholar]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Yin, H.; Song, X.; Zhang, Y.; Liu, M.; Sang, J.; Jiang, J.; Li, J.; Zhuo, R. Integration of small RNA s, degradome and transcriptome sequencing in hyperaccumulator Sedum alfredii uncovers a complex regulatory network and provides insights into cadmium phytoremediation. Plant Biotechnol. J. 2016, 14, 1470–1483. [Google Scholar] [CrossRef] [Green Version]
- Szczypka, M.S.; Wemmie, J.A.; Moye-Rowley, W.S.; Thiele, D.J. A yeast metal resistance protein similar to human cystic fibrosis transmembrane conductance regulator (CFTR) and multidrug resistance-associated protein. J. Biol. Chem. 1994, 269, 22853–22857. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.N. | Gene Name | Gene ID | Chr. No. | Protein Length (AA) | MW (KDa) | PI | Predict Location | Motif | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | SpHMA1 | evm.model.000029F.120 | Chr11 | 841 | 91.75 | 8.67 | Chloroplast | TGE | SPC | DKTGT | HP | PEDK | GDGINDAP |
| 2 | SpHMA2 | evm.model.000008F.259 | Chr18 | 771 | 82.22 | 7.17 | Cell membrane | TGE | CPC | DKTGT | HP | PEDK | GDGINDAP |
| 3 | SpHMA3 | evm.model.000098F.40 | Chr03 | 805 | 86.87 | 5.7 | tonoplast | TGE | CPC | DKTGT | HP | PEDK | GDGINDAP |
| 4 | SpHMA4 | evm.model.000053F.243.6 | Chr06 | 995 | 107.58 | 5.61 | Cell membrane | TGE | CPC | DKTGT | HP | PLGK | GDGINDSP |
| 5 | SpHMA5 | evm.model.000076F.84 | Chr01 | 959 | 103.56 | 5.55 | Cell membrane | TGE | CPC | DKTGT | HP | PDQK | GDGINDSP |
| 6 | SpHMA6 | evm.model.000022F.259 | Chr31 | 936 | 98.47 | 7.92 | Cell membrane | TGE | CPC | DKTGT | HP | PDEK | GDGINDAA |
| 7 | SpHMA7 | evm.model.000000F.676 | Chr02 | 694 | 74.55 | 7.03 | Cell membrane | TGE | CPC | DKTGT | HP | PAGK | GDGINDSP |
| 8 | SpHMA8 | evm.model.000024F.130 | Chr27 | 878 | 93.06 | 5.84 | Cell membrane | TGE | CPC | DKTGT | HP | PQQK | GDGINDAP |
| Primer Name | Primer Sequence (5′-3′) | Primer Length (bp) |
|---|---|---|
| SpHMA7-F | ATGCAGCTTCCAGTCATATTCA | 22 |
| SpHMA7-R | TTACTCTACAGTTATTTCAAGAATG | 25 |
| SpHMA7.pyes2-F | tcactatagggaatattaATGCAGCTTCCAGTCATATTCA | 40 |
| SpHMA7.pyes2-R | gatgcggccctctagTTACTCTACAGTTATTTCAAGAATG | 40 |
| SpUBC9-F | TGGCGTCGAAAAGGATTCTGA | 21 |
| SpUBC9-F | CCTTCGGTGGCTTGAATGGATA | 22 |
| qRT.SpHMA1-F | TGCTGCTGCTTGTCCTTACT | 20 |
| qRT.SpHMA1-R | AGACGCAAAGGCAGCCATAG | 22 |
| qRT.SpHMA2-F | GTGGCTGTAATCCCTGCTGT | 20 |
| qRT.SpHMA2-R | GATGTGGCCGCCTTTGAAAG | 21 |
| qRT.SpHMA3-F | TAAAATGGGTGGCTGTGGCT | 21 |
| qRT.SpHMA3-R | AACTGCCATGATTGCCAGGA | 21 |
| qRT.SpHMA4-F | TTATGGTGTCACCGCTGCAA | 20 |
| qRT.SpHMA4-R | TCAAACCCTGCATCCTCCAC | 20 |
| qRT.SpHMA5-F | GTTCGGATCGCTAACGTTGC | 20 |
| qRT.SpHMA5-R | TCAGCTCAGCTTCGAAACCA | 20 |
| qRT.SpHMA6-F | CAGAGGTGTTGCTGCTGGTA | 20 |
| qRT.SpHMA6-R | GCTGAAGACACTTGCGGTTG | 20 |
| qRT.SpHMA7-F | GTTGATCGAGCTTTCACCCG | 20 |
| qRT.SpHMA7-R | TGTCACCGGGTTGGATCAAT | 20 |
| qRT.SpHMA8-F | CGAAGAGACGTGTTTCGGGA | 20 |
| qRT.SpHMA8-R | CACAGCACAATGCCACCAAA | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Q.; Qiu, W.; Yu, M.; Li, S.; Lu, Z.; Zhu, Y.; Kan, X.; Zhuo, R. Genome-Wide Characterization of Sedum plumbizincicola HMA Gene Family Provides Functional Implications in Cadmium Response. Plants 2022, 11, 215. https://doi.org/10.3390/plants11020215
Huang Q, Qiu W, Yu M, Li S, Lu Z, Zhu Y, Kan X, Zhuo R. Genome-Wide Characterization of Sedum plumbizincicola HMA Gene Family Provides Functional Implications in Cadmium Response. Plants. 2022; 11(2):215. https://doi.org/10.3390/plants11020215
Chicago/Turabian StyleHuang, Qingyu, Wenmin Qiu, Miao Yu, Shaocui Li, Zhuchou Lu, Yue Zhu, Xianzhao Kan, and Renying Zhuo. 2022. "Genome-Wide Characterization of Sedum plumbizincicola HMA Gene Family Provides Functional Implications in Cadmium Response" Plants 11, no. 2: 215. https://doi.org/10.3390/plants11020215