
Mobile Host mRNAs Are Translated to Protein in the Associated Parasitic Plant Cuscuta campestris


1
School of Plant and Environmental Sciences, Virginia Tech, Blacksburg, VA 24061, USA
2
Graduate School of Life and Environmental Sciences, Osaka Prefecture University, 1-1 Gakuen-cho, Naka-ku, Sakai 599-8531, Japan
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
‡
Present addresses: Division of Plant Science, University of Missouri, Columbia, MO 65211, USA.
Plants 2022, 11(1), 93; https://doi.org/10.3390/plants11010093
Submission received: 17 November 2021
/
Revised: 23 December 2021
/
Accepted: 23 December 2021
/
Published: 28 December 2021
(This article belongs to the Special Issue Parasitic Weeds: Biology and Control)
Abstract
:Cuscuta spp. are obligate parasites that connect to host vascular tissue using a haustorium. In addition to water, nutrients, and metabolites, a large number of mRNAs are bidirectionally exchanged between Cuscuta spp. and their hosts. This trans-specific movement of mRNAs raises questions about whether these molecules function in the recipient species. To address the possibility that mobile mRNAs are ultimately translated, we built upon recent studies that demonstrate a role for transfer RNA (tRNA)-like structures (TLSs) in enhancing mRNA systemic movement. C. campestris was grown on Arabidopsis that expressed a β-glucuronidase (GUS) reporter transgene either alone or in GUS-tRNA fusions. Histochemical staining revealed localization in tissue of C. campestris grown on Arabidopsis with GUS-tRNA fusions, but not in C. campestris grown on Arabidopsis with GUS alone. This corresponded with detection of GUS transcripts in Cuscuta on Arabidopsis with GUS-tRNA, but not in C. campestris on Arabidopsis with GUS alone. Similar results were obtained with Arabidopsis host plants expressing the same constructs containing an endoplasmic reticulum localization signal. In C. campestris, GUS activity was localized in the companion cells or phloem parenchyma cells adjacent to sieve tubes. We conclude that host-derived GUS mRNAs are translated in C. campestris and that the TLS fusion enhances RNA mobility in the host-parasite interactions.
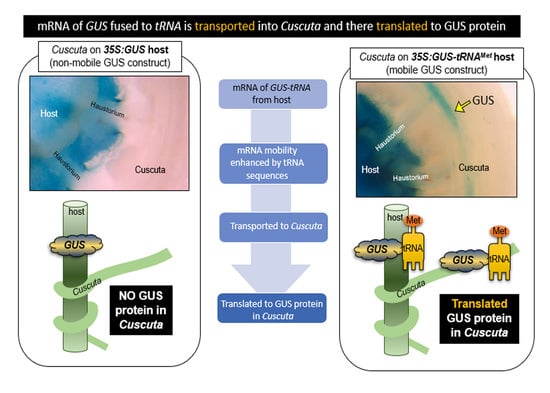
1. Introduction
Cuscuta spp. (dodders) are holoparasitic plants that attack a broad range of hosts, and are capable of causing substantial agricultural losses [1]. Cuscuta plants typically consist of yellow or orange stems, lacking roots or developed leaves. They connect by coiling around host stems, petioles, and leaves, and at these points of contact they develop haustoria, which are unique structures that grow invasively into the host to form a continuum with the host’s xylem and phloem tissues [2]. The haustorium functions to feed the parasite though uptake of water, sugars, and other nutrients, but is also capable of facilitating exchange of macromolecules including proteins [3], mRNAs [4], microRNAs [5], and possibly even DNAs, as implicated by horizontal gene transfer [6]. Movement of each of these classes of macromolecules raises many questions regarding the exchange of signals between host and parasite, but the least understood are arguably mRNAs, for which little is known about their mechanisms of movement, fate, and function in the plant-plant interaction. In particular, it is important to understand whether mobile mRNAs from the host are able to be translated into protein after arriving in the parasite, as this would provide a powerful mechanism for transmission of proteins that otherwise would be unable to move between the organisms.
Plants have evolved the ability to transport RNAs over long distances in the phloem. These non-cell-autonomous mRNAs are thought to function in coordinating plant development and response to stress [7]. Several mobile mRNAs have been demonstrated to affect the phenotype of the destination tissue, including Flowering locus T (FT), for which mobile protein and mRNA move from leaf phloem into shoot apical meristem to promote flowering [8]. Other well-characterized long-distance mobile mRNAs associated with phenotypes are a fusion of pyrophosphate-dependent phosphofructokinase with LeT6 in tomato [9], the BEL5 transcription factor from potato [10], and Gibberellic-Acid insensitive [11], among others [12,13]. Recent studies have identified large numbers of mobile cellular mRNAs through hetero-grafting combined with high-throughput sequencing technologies [14,15,16]. The large-scale exchange of mRNAs between Cuscuta plants and their hosts suggests that they are able to tap into this system, although the biological significance is not yet clear [17].
Another unsolved mystery is the mechanism(s) by which the cell-to-cell movement of mRNAs is regulated in plants. Studies have indicated multiple factors that contribute to mRNA mobility, including sequences of the 3′ and 5′ untranslated regions (UTRs) [18], and the presence of methylated cytosine bases in the mRNA coding sequence, or UTRs [19]. Furthermore, transfer RNA (tRNA) sequences or tRNA-like structures (TLSs) in the 3′ UTR of an mRNA were found to increase systemic mobility of associated mRNAs in plants [13,20]. In the latter work, Zhang et al. [20] added tRNA sequences to the β-glucuronidase (GUS) protein coding sequence and showed that they were sufficient to promote GUS mRNA mobility across Arabidopsis graft junctions. GUS enzyme activity was detected in the recipient tissue; in this case, wild type roots grafted to shoots expressing the 35S:GUS-tRNA transgene. They also demonstrated that the mobile GUS-tRNA mRNA was translated to protein in the roots. Not all tRNAs conferred mobility to associated mRNAs, so there is specificity in the system. For example, the tRNAs for methionine (tRNAMet) and glycine (tRNAGly) conferred mobility, while the isoleucine tRNA (tRNAIle) did not. The three-dimensional structure of the TLSs was shown to be important, as indicated by the finding that certain mutations of the hairpin loop structures affect mobility, as deletion of A and T loops of tRNAMet (tRNAMet-dAT) abolished movement, while deletion of D and T loops (tRNAMet-dDT) retained mobility.
Our long-term objective is to understand the mechanisms by which Cuscuta spp. interact with their hosts, and specifically the role of RNAs in the interaction. Recent work by Liu et al. [3] suggested that protein movement between hosts and C. australis takes place primarily by direct protein movement, without need for an mRNA intermediary. In this paper, we address two central questions: (1) Does a tRNA fusion system that confers cell-cell mobility on GUS gene mRNAs in Arabidopsis also enable it to traffic into C. campestris? (2) Is such a mobile GUS mRNA translated into protein in C. campestris? Indeed, we have found that tRNA fused to the GUS gene facilitates the movement of GUS mRNA and results in GUS enzyme activity in C. campestris haustoria, stems, floral organs, phloem, and apical termini of sieve tubes. These results support the idea that the transported GUS-tRNA mRNA from Arabidopsis host plants is translated in C. campestris cells.
2. Results
2.1. tRNA Fusions Influence Mobility of GUS Activity
We used transgenic Arabidopsis plants expressing GUS either with or without tRNA fusions and assayed the movement of GUS activity into attached C. campestris. For this experiment, C. campestris stems parasitizing Arabidopsis floral shoots were sectioned as a unit and stained to reveal GUS activity (Figure 1A). As a negative control, we examined C. campestris growing on nontransgenic Arabidopsis because the related species C. pentagona has been reported to have endogenous GUS activity [21]. Unlike C. pentagona, no GUS activity was detected in wild type C. campestris (Figure 1B). C. campestris was then grown on Arabidopsis with 35S:GUS or 35S:GUS-tRNAMet transgenes and again sectioned and stained to reveal GUS activity. No GUS activity was detected in C. campestris expressing GUS without the tRNA sequence (Figure 1C,D) but was evident in C. campestris parasitizing hosts expressing GUS-tRNAMet (Figure 1E,F). These results indicate that the presence of tRNA motif promotes mobility of GUS activity from host to C. campestris, similar to its function in Arabidopsis grafting experiments [20].
Considering the open exchange of materials between Cuscuta spp. and their hosts, it is important to use extra caution in judging whether GUS moves as a protein, as opposed to an mRNA that is subsequently translated into protein. Although GUS has been considered to be a non-mobile protein, having been used for decades as a cell- and tissue-specific indicator of gene expression [22], it has been proposed to be mobile from host plants to C. australis [3]. Therefore, to further restrict GUS protein mobility, we fused a sequence encoding the endoplasmic reticulum (ER) signal peptide to the GUS gene construct. Previous studies have shown that ER targeting peptides are sufficient to block GFP protein movement [23,24]. Additionally, we used tRNA variants that were shown to differ in ability to confer mobility on mRNAs in grafted Arabidopsis [20]. Thus, in addition to using the non-ER localized GUS constructs, we generated transgenic Arabidopsis expressing 35S:ER-GUS, 35S:ER-GUS-tRNAMet and 35S:ER-GUS-tRNAMet-dDT, as well as others derived from the constructs reported by Zhang et al. (2016). Transgenic Arabidopsis plants were confirmed to show strong GUS activity using the fluorescent 4-MUG assay, while wild type plants had negligible activity (Supplemental Table S1).
C. campestris was grown on the Arabidopsis plants expressing GUS with or without tRNA fusions and with or without ER localization signals. The parasite stem was removed from the host and the haustoria regions were sectioned longitudinally and transversely before staining to detect GUS activity. No GUS activity was detected in C. campestris parasitizing hosts with 35S:GUS or 35S:ER-GUS (Figure 2A,B,G,H). However, the blue dye indicative of GUS activity was evident in C. campestris parasitizing hosts with tRNA fusions to the GUS gene: 35S:GUS-tRNAMet, 35S:GUS-tRNAMet-dDT, 35S:ER-GUS-tRNAMet, and 35S:ER-GUS-tRNAMet-dDT (Figure 2C–F,I–L). This pattern was confirmed by counting the number of haustoria showing GUS activity on these and additional transgenic lines. Haustoria from negative controls (wild type Col-0, 35S:empty, 35S:GUS, and 35S:ER-GUS) never showed GUS enzyme activity (Table 1). In contrast, 30% to 80% of C. campestris haustorial regions parasitizing Arabidopsis GUS lines with tRNAMet, tRNAMet-dDT, tRNAGly, and tRNAIle fusions showed GUS activity. Furthermore, 30% to 39% of C. campestris haustoria growing on hosts with ER-GUS-tRNAs showed GUS enzyme activity. The one exception was a lack of GUS enzyme activity in C. campestris growing on Arabidopsis expressing 35S:GUS-tRNAMet-dAT, although this is consistent with a lack of mobility reported for this construct in the Arabidopsis grafting assay [20].
2.2. GUS mRNA in C. campestris Is Associated with GUS-tRNA Fusions
We investigated the mobility of GUS mRNA from Arabidopsis plants expressing GUS with or without tRNA sequences. To avoid any possibility of contamination from parasite tissues in close contact with the host, total RNA was extracted from C. campestris stem more than 1 cm away from the haustoria. RT-PCR was used to detect mRNAs from the GUS gene constructs and C. campestris actin gene (CcActin8) as a positive control. While CcActin8 was amplified from all samples, GUS mRNAs were only amplified from parasite tissues where Cuscuta was growing on Arabidopsis expressing tRNA fusions: GUS-tRNAMet, GUS-tRNAMet-dDT, ER-GUS-tRNAMet, and ER-GUS-tRNAMet-dDT (Figure 2M).
2.3. GUS mRNA Moves Long Distances in C. campestris and GUS Activity Is Localized in Phloem Cells
To investigate the distribution of GUS protein in C. campestris, stems of the parasite were sectioned at increasing distances from the haustorial region (Supplemental Figure S1A). GUS enzyme activity was strongly expressed in the Arabidopsis 35S:GUS-tRNAMet host stems (Supplemental Figure S1B,C,E,F). GUS activities were detected in C. campestris stems near the haustoria regions (Supplemental Figure S1C,F), as well as from 0.7 cm to 12 cm away (Supplemental Figure S1D,G–I). Quantitative RT-PCR Analyses of mRNAs from the same experiment indicated the presence of mobile GUS-tRNAMet and GUS-tRNA-Met-dDT transcripts from the entire length of the C. campestris stem (Supplemental Figure S1K,L).
To further localize the presence of the GUS enzyme, we assayed flowers of C. campestris grown on 35S:GUS-tRNAMet. GUS activity was detected at the base of floral buds located 4 to 6 cm away from the haustoria (Figure 3A). Specifically, GUS was observed in the peduncle and the base of, but not inside, the C. campestris ovary (Figure 3B,C). GUS activity was also detected in the vascular tissues at the base of the apical tip of C. campestris grown on 35S:GUS-tRNAMet and 35S:GUS-tRNAMet-dDT expressing Arabidopsis (Figure 3E,F). As in the flower, GUS activity was not detected in the meristematic region. Longitudinal (Figure 3H,I) and transverse (Figure 3J,K) sections showed that GUS activity was not co-localized with xylem. In further support of this observation, sequential staining for GUS activity, followed by phloroglucinol-HCl staining of lignin in xylem cells [25], indicated that for C. campestris growing on Arabidopsis 35S:GUS-tRNAMet plants the GUS signals were detected more centrally in the C. campestris stem than the lignin staining (Supplemental Figure S2).
To test whether GUS activity was localized in the phloem, we performed double staining for GUS activity and callose deposition that is indicative of sieve plates. GUS signals were detected first in C. campestris grown on Arabidopsis 35S:GUS-tRNAMet (Figure 4A); then the same sections were transferred to a confocal microscopy to identify GUS-stained cells by transmission image (Figure 4B) and stained with aniline blue to visualize callose deposition on the sieve plates of sieve tubes (Figure 4C). GUS activity was localized in the array of cells next to sieve tubes containing aniline blue-stained sieve plates (Figure 4D). Essentially, the same localization patterns of GUS activity and sieve tubes were obtained in C. campestris grown on Arabidopsis 35S:GUS-tRNAMet-dDT and 35S:GUS-tRNAGly (Supplemental Figure S3). These results suggest that GUS proteins were localized in the companion cells or phloem parenchyma cells adjacent to sieve tubes.
3. Discussion
The fate and function of mobile mRNAs in plants has been the subject of speculation and research since the earliest reports of systemically trafficked mRNAs in plants [26,27]. These issues are all the more intriguing when they occur in the context of host-parasite trans-species interactions. Recent breakthroughs have contributed to understanding how the mobility of mRNAs is regulated in plants and have shown that mobile mRNAs may be translated into proteins in their destination cells [19,20], but the subject has yet to be resolved in parasitic plant interactions. We used TLS-mediated mRNA mobility to simultaneously investigate mechanisms regulating mRNA transfer and translation of the mRNA in C. campestris feeding on transgenic plants.
The fusion of tRNA sequences to the GUS gene conferred mobility on GUS mRNA from Arabidopsis into attached C. campestris (Figure 2; Supplementary Figure S1). Subsequent translation to protein resulted in consistent detection of GUS enzyme activity in these C. campestris shoots (Figure 1 and Figure 2). Our results were consistent for two tRNAIle constructs (with or without an ER localization signal peptide) and independently verified in two different laboratories (Japan and the U.S.A.). These data confirm a lack of mobility for GUS encoded by constructs missing the tRNAs or for GUS fused to tRNAMet-dAT in host-parasite systems. Our findings are largely consistent with the graft transmissibility of GUS-tRNA fusions reported by Zhang et al. [20], who also demonstrated the mRNA mobility of GUS fused to tRNAMet, tRNAMet-dDT, and tRNAGly (compare to Table 1). One discrepancy between the Arabidopsis graft studies and our host-Cuscuta data is the mobility of GUS-tRNAIle into C. campestris, whereas no graft transmissibility of this tRNA fusion was seen [28]. Taken together, these results suggest that the regulation of mRNA movement across the C. campestris haustorial connection is similar, but not identical to, an Arabidopsis graft junction.
The presence of a TLS element associated with mRNA is just one of the mechanisms currently known to facilitate phloem mobility, but we wondered whether it could account for the large number of mobile mRNAs in C. campestris parasitizing Arabidopsis. To test this, we evaluated 492 of the most abundant mobile Arabidopsis mRNAs from a list of nearly 8000 previously reported host-to-Cuscuta mobile mRNAs [17]. Of these genes, 392 (79.6%) are reported as also being cell-to-cell mobile mRNAs in Arabidopsis (www.arabidopsis.org). We searched these 392 genes for a TLS structure and found that 35 genes (8.9%) had a TLS. This is consistent with a previous report that 11.4% of Arabidopsis mobile mRNAs identified from a grafting study have a TLS [20]. We conclude that the TLS motif is likely just one of several mechanisms to regulate host-Cuscuta mobility of mRNAs [19,29], yet this is an important finding in that it illustrates a simple mechanism for engineering mRNA mobility in a gene that otherwise may not be mobile. This will be a useful experimental tool for further investigations of host-Cuscuta interactions.
Cuscuta spp. are known to take proteins directly from their hosts. This has been shown for phloem-expressed, soluble GFP [30,31] and phosphinothricin acetyl transferase [32]. Recently, large-scale movement of proteins from Arabidopsis and soybean hosts to C. australis has been described, including direct mobility of a GUS protein [3]. This stands in contrast to our work in which no evidence of GUS protein movement was detected. The work with C. australis did not include extra sequences with the GUS gene construct to enhance mobility, and the case for mobility was made based on detection of GUS activity in the absence of successful amplification of GUS mRNA from the same tissues. It is difficult to reconcile the difference in our two studies, although slightly different methodologies were used. The simplest answer may lie in potential differences in haustorial function between C. campestris and C. australis, and this subject warrants further investigation. It is likely that both mechanisms operate, and Liu et al. [3] concede that in their system some amount of host-encoded protein may arrive in the parasite through the translation of mobile mRNA. The larger question may revolve around the relative contributions of direct movement of mature proteins as compared to mRNA intermediates.
Localization of GUS expression in the parasite suggests that GUS mRNA moves long distances in the parasite and is imported into companion cells or phloem parenchyma cells of C. campestris (Figure 3 and Figure 4). The GUS activity was observed near shoot apices and floral organs, although it was not detected inside these structures. The pattern of staining of specific cells or groups of cells may be an artifact of the sectioning and staining methodology, or may reflect the uptake and translation of mobile mRNAs by specific cells, as suggested by targeted the synthesis and translation of mobile mRNA in specific phloem companion cells [33,34].
Taken together, the appearance of functional host protein in the parasite raises intriguing possibilities for novel organismal interactions. There is little doubt that direct protein exchange occurs between parasitic plants and their hosts, but mobile mRNAs encoding proteins that are membrane bound or too large to easily translocate would provide another avenue of plant-plant interaction. Just as recent studies of C. campestris microRNAs have demonstrated a role for these molecules in suppressing expression of specific host genes [35], mobile mRNAs may provide an additional means of host manipulation. It will be interesting to investigate the functional significance of this process.
4. Material and Methods
4.1. Plant Material and Growth Conditions
Experiments were conducted in two locations, with consistent results despite minor differences in growth conditions. In Japan, C. campestris seeds were harvested from lab-grown plants parasitizing Nicotiana tabacum hosts grown at 25 °C with 16 h light and 8 h dark cycles. Experimental growth conditions of C. campestris and Arabidopsis were described previously [36]. In the US, seedlings of a lab-growth line of C. campestris [17] were inoculated on beets (Beta vulgaris) and grown for one month at 25 °C with 14 h light and 10 h dark cycles. Pieces of C. campestris shoot tip (around 5 cm long) growing on beets were inoculated on the middle of Arabidopsis flowering stems (around 7 cm long). To promote coiling, plants were grown under a 65W Spot-Gro Plant Light (Sylvania) with 14 h light and 10 h dark cycles for two weeks.
Arabidopsis seeds were stratified in water at 4 °C for a day and then sown onto Sungro Professional Growing Mix. Plants were grown in a Conviron (Controlled Environments, Inc.) growth chamber with 9 h light and 15 h dark cycles for 6 to 8 weeks before inoculation with C. campestris.
4.2. Arabidopsis Plants Expressing ER-GUS with tRNAs
For cloning endoplasmic reticulum (ER) signal peptides fused to GUS-tRNA constructs, gDNAs were first extracted from transgenic Arabidopsis lines expressing GUS-tRNAMet, GUS-tRNAMet-dDT, GUS-tRNAGly, and GUS-tRNAIle [20]. These gDNAs were used as templates for cloning to insert the ER signal sequence into ER-GUS-tRNA constructs. ER-GUS with different tRNAs were cloned into pEarleyGate100 using the forward primer (with 23 amino acid ER targeting signal peptide from AT1G21270) and tRNA specific reverse primers (Supplemental Table S2) [37]. Transgenic Arabidopsis plants were generated by floral dipping [38], and at least five individual T2 lines were tested in this study.
4.3. Histochemical and Quantitative GUS Assays
Haustorial regions of two-week-old C. campestris attachments on various Arabidopsis transgenic lines were collected and embedded in 5% agarose. Using a VT1200 S fully automated vibrating blade microtome (Leica), agarose blocks with plant tissues were sectioned with 400 µm thickness and 0.8 mm/sec speed. Sliced tissues were collected into 48 well plates for further analysis. For the GUS staining, sectioned samples were stained with X-gluc solution for 2 h and destained in 70% EtOH for 10 h.
4.4. Paraffin Embedding
GUS-stained Cuscuta stems were fixed with 4% (w/v) paraformaldehyde phosphate buffer solution (FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) at room temperature for 24 h. Fixed samples were dehydrated and embedded in paraffin (Paraplast, Leica Biosystems, Wetzlar, Germany) as described previously [36]. Paraffin blocks were cut into 20 µm-thick sections by using a microtome (PR-50, Yamato Kohki, Asaka, Japan). Sections were extended with water on MAS-coated slide glass (Matsunami Glass Ind., Ltd., Kishiwada, Japan) and dewaxed as described previously [36]. Samples were observed by a BX53 Upright Microscope (Olympus, Tokyo, Japan, https://www.olympus-lifescience.com/, accessed on 16 November 2021).
For the histochemical GUS staining, sectioned samples were stained by GUS staining solution with 5-bromo-4- chloro-3-indolyl-BD-glucuronide (X-gluc) (Fisher) for 3 h in accordance with the guidelines of the manufacturer and photographed using a stereo-zoom microscope (Discovery V12, Carl Zeiss, Jena, Germany).
The fluorescent β-galactosidase assay with 4-MUG (Fisher) was conducted to detect GUS activity under high liquid treatment. Plant samples from Arabidopsis and C. campestris were collected, and total proteins were extracted in accordance with the guidelines of the manufacturer. Concentrations of total proteins were quantified by Bradford assay (Bio-Rad, Hercules, CA, USA) using bovine serum albumin (Promega, Madison, WI, USA) as a standard. For the 4-MUG assay, fluorescence was detected at the excitation/emission wavelengths of 365 nm/455 nm by a plate reader machine (Biotek Synergy HT). The GUS enzyme activity was expressed as picomoles of 4-methylumbelliferone (MU) (Sigma) produced per milligram protein per minute. Based on standard curves, the results of the 4-MUG assay were calculated.
4.5. Phloroglucinol-HCl (Wiesner) Staining
Phloroglucinol (3%) (Sigma) dissolved in ethanol was mixed with concentrated HCl (Sigma) to make the phloroglucinol-HCl (Wiesner) staining solution [39]. Sectioned tissues were dipped into the solution for 5 min and directly observed under a stereo-zoom microscope (Discovery V12, Carl Zeiss).
4.6. Reverse Transcriptase (RT) PCR and Quantitative PCR
Total RNAs were extracted from at least five independent biological replicates of Arabidopsis or C. campestris stems using the Trizol reagent and in accordance with the protocol of the manufacturer (Invitrogen). Equal amounts of extracted total RNAs were reverse transcribed using random primers and M-MLV in accordance with the protocol of the manufacturer (High Capacity cDNA Reverse Transcription Kit, ABI).
Gene-specific primers (Table S2) were used in RT-PCR with iProof High-Fidelity DNA Polymerase (Bio-rad) to amplify genes of interest. GUS-plus primers were used to measure the GUS mRNA movement from host plants into C. campestris stems. CcActin 8 was a positive control to check the equal amount of RNA.
For quantitative RT-PCR (qRT-PCR), C. campestris stems (approximately 12 cm-long from the parasite haustorial site to the apical tip) were divided into six segments (2 cm each). Total RNAs were extracted from three biological replicates of C. campestris stems by using RNeasy Plant Mini Kit (QIAGEN, Hilden, Germany). First cDNA strand synthesis was performed by using oligio(dT) primer and ReverTra Ace (TOYOBO, Osaka, Japan). qRT-PCR of GUS transcript was performed by using gene specific primers (Table S2), Fast SYBRTM Green Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), and the StepOnePlus Real-Time PCR System (Thermo Fisher Scientific, https://corporate.thermofisher.com/, accessed on 16 November 2021). Standard curves were generated by using partial GUS sequence cloned in a plasmid pCR™-Blunt II-TOPO® (Thermo Fisher Scientific) as a template.
4.7. Aniline-Blue Staining
Dewaxed paraffin sections, 20 µm-thickness, of C. campestris stems were stained for 45 min with 1% (w/v) aniline blue solution dissolved in 50mM NaPO4 buffer, pH 7.0, and washed by sterile water twice. Fluorescence were observed by using BX53 Upright Microscope (Olympus) and a laser-scanning confocal microscope (Leica TCS SP8, Leica Biosystems).
4.8. Searching tRNA-like Structure (TLS) Motif in Mobile Host Genes
A database, containing 492 mRNAs that had been found to be mobile from Arabidopsis to Cuscuta [17], was screened to determine the presence of a TLS motif. The full-length sequences of each mobile mRNA were obtained from TAIR (arabidopsis.org). PlaMoM (Plant Mobile Macromolecules) (http://www.systembioinfo.org/plamom/, accessed on 16 November 2021) provides a search tool to predict a TLS element [40] and was used to analyze the mobile Arabidopsis genes to identify presence of any TLS motif.
5. Conclusions
We have addressed the question of whether host-encoded mRNA could be translated into a functional protein following translocation into the parasitic plant C. campestris. As part of this work, we used tRNA gene sequences as signals for long-distance trafficking of mRNAs [20]. We observed that GUS-tRNA fusions expressed in Arabidopsis hosts resulted in detection of both GUS mRNA and GUS enzymatic activity in associated C. campestris shoots. Furthermore, this GUS expression appeared in C. campestris tissues near the haustorial connections as well as in shoots and floral organs located distantly from the point of host attachment. GUS expression was associated with the parasite vascular system, suggesting that mobile mRNAs are translated in companion cells or phloem parenchyma. The fact that functional GUS enzyme was produced in the parasite raises the possibility that mobile mRNAs lead to exchange of proteins that may affect the physiology of one or both plants in the parasite-host interaction. Considering the breadth of diversity in mobile mRNAs [17], it is interesting to consider a potential role for mRNAs in parasitic plant communication.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants11010093/s1, Figure S1: GUS staining and quantification of GUS transcript levels using Cuscuta on 35S:GUS-tRNAMet and 35S:GUS-tRNAMet-dDT Arabidopsis.; Figure S2: Histochemical localization of β-glucuronidase and xylem.; Figure S3: GUS activity detected in the cells adjacent to aniline-blue-stained sieve tube.; Table S1: β-Glucosidase activities in host Arabidopsis plants.; Table S2: List of primers used in this study.
Author Contributions
K.A. and J.H.W. independently conceived the experiments; S.-Y.P. and K.S. conducted assays of GUS activity and mRNA movement; S.-Y.P. generated ER-tagged constructs and plants; K.S. conducted high resolution GUS staining experiments; J.B. analyzed TLS sequences and provided technical support; S.-Y.P. and K.S. analyzed the data; S.-Y.P. drafted the manuscript with additional help from J.H.W., K.A. and K.S.; K.A. and J.H.W. share equal responsibility and agree to serve as co-corresponding authors. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support was provided by US National Science Foundation award IOS-1645027 and US Department of Agriculture National Institute of Food and Agriculture award (VA-160111) to J.H.W.; Grant-in-Aid for JSPS Fellows (19J14848, JSPS) to K.S.; and Grant-in-Aid for Scientific Research (18H03950 and 19H00944, JSPS) to K.A.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We thank Friedrich Kragler (Max Planck Institute) for GUS-tRNA fusion plants and Junpei Takano (Osaka Prefecture University) for the help in confocal laser scanning microscopy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Costea, M.; Tardif, F.J. The biology of Canadian weeds. 133. Cuscuta campestris Yuncker, C. gronovii Willd. ex Schult., C. umbrosa Beyr. ex Hook., C. epithymum (L.) L. and C. epilinum Weihe. Can. J. Plant Sci. 2006, 86, 293–316. [Google Scholar] [CrossRef]
- Shimizu, K.; Aoki, K. Development of parasitic organs of a stem holoparasitic plant in genus Cuscuta. Front. Plant Sci. 2019, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Shen, G.; Xu, Y.; Liu, H.; Zhang, J.; Li, S.; Li, J.; Zhang, C.; Qi, J.; Wang, L.; et al. Extensive inter-plant protein transfer between Cuscuta parasites and their host plants. Mol. Plant 2019, 13, 573–585. [Google Scholar] [CrossRef]
- Kim, G.; Westwood, J.H. Macromolecule exchange in Cuscuta–host plant interactions. Curr. Opin. Plant Biol. 2015, 26, 20–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, N.R.; Axtell, M.J. Small RNA warfare: Exploring origins and function of trans-species microRNAs from the parasitic plant Cuscuta. Curr. Opin. Plant Biol. 2019, 50, 76–81. [Google Scholar] [CrossRef]
- Yang, Z.; Wafula, E.K.; Kim, G.; Shahid, S.; McNeal, J.R.; Ralph, P.E.; Timilsena, P.R.; Yu, W.-B.; Kelly, E.A.; Zhang, H.; et al. Convergent horizontal gene transfer and cross-talk of mobile nucleic acids in parasitic plants. Nat. Plants 2019, 2019. 5, 991–1001. [Google Scholar] [CrossRef]
- Kehr, J.; Buhtz, A. Long distance transport and movement of RNA through the phloem. J. Exp. Bot. 2007, 59, 85–92. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.-J.; Huang, N.-C.; Liu, Y.-S.; Lu, C.-A.; Yu, T.-S. Long-distance movement of Arabidopsis FLOWERING LOCUS T RNA participates in systemic floral regulation. RNA Biol. 2012, 9, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Canio, W.; Kessler, S.; Sinha, N. Developmental Changes Due to Long-Distance Movement of a Homeobox Fusion Transcript in Tomato. Science 2001, 293, 287–289. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Chatterjee, M.; Yu, Y.; Suh, S.-G.; Miller, W.A.; Hannapel, D.J. Dynamics of a mobile RNA of potato involved in a long-distance signaling pathway. Plant Cell 2006, 18, 3443–3457. [Google Scholar] [CrossRef] [Green Version]
- Haywood, V.; Yu, T.-S.; Huang, N.-C.; Lucas, W.J. Phloem long-distance trafficking of GIBBERELLIC ACID-INSENSITIVE RNA regulates leaf development. Plant J. 2005, 42, 49–68. [Google Scholar] [CrossRef] [PubMed]
- Hannapel, D.J. Long-Distance Signaling via Mobile RNAs. In Long-Distance Systemic Signaling and Communication in Plants; Baluška, F., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 53–70. [Google Scholar]
- Kehr, J.; Kragler, F. Long distance RNA movement. New Phytol. 2018, 218, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Thieme, C.J.; Rojas-Triana, M.; Stecyk, E.; Schudoma, C.; Zhang, W.; Yang, L.; Miñambres, M.; Walther, D.; Schulze, W.X.; Paz-Ares, J.; et al. Endogenous Arabidopsis messenger RNAs transported to distant tissues. Nat. Plants 2015, 1, 15025. [Google Scholar] [CrossRef]
- Yang, Y.Z.; Mao, L.Y.; Jittayasothorn, Y.; Kang, Y.M.; Jiao, C.; Fei, Z.J.; Zhong, G.-Y. Messenger RNA exchange between scions and rootstocks in grafted grapevines. BMC Plant Biol. 2015, 15, 251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, X.H.; Liu, W.Q.; Wang, T.; Zhang, J.L.; Li, X.J.; Zhang, W.N. Systemic long-distance signaling and communication between rootstock and scion in grafted vegetables. Front. Plant Sci. 2020, 11, 460. [Google Scholar] [CrossRef]
- Kim, G.; LeBlanc, M.L.; Wafula, E.K.; de Pamphilis, C.W.; Westwood, J.H. Genomic-scale exchange of mRNA between a parasitic plant and its hosts. Science 2014, 345, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.K.; Lin, T.; Hannapel, D.J. Untranslated regions of a mobile transcript mediate RNA metabolism. Plant Physiol. 2009, 151, 1831–1843. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Perrera, V.; Saplaoura, E.; Apelt, F.; Bahin, M.; Kramdi, A.; Olas, J.; Mueller-Roeber, B.; Sokolowska, E.; Zhang, W.; et al. m5C methylation guides systemic transport of messenger RNA over graft junctions in plants. Curr. Biol. 2019, 29, 2465–2476.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Thieme, C.J.; Kollwig, G.; Apelt, F.; Yang, L.; Winter, N.; Andresen, N.; Walther, D.; Kragler, F. tRNA-Related Sequences Trigger Systemic mRNA Transport in Plants. Plant Cell 2016, 28, 1237–1249. [Google Scholar] [CrossRef] [Green Version]
- Schoenbeck, M.A.; Swanson, G.A.; Brommer, S.J. β-Glucuronidase activity in seedlings of the parasitic angiosperm Cusctua pentagona: Developmental impact of the β-glucuronidase inhibitor saccharic acid 1,4-lactone. Funct. Plant Biol. 2007, 34, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Waigmann, E.; Zambryski, P. Tobacco mosaic virus movement protein-mediated protein transport between trichome cells. Plant Cell 1995, 7, 2069–2079. [Google Scholar]
- Crawford, K.M.; Zambryski, P.C. Subcellular localization determines the availability of non-targeted proteins to plasmodesmatal transport. Curr. Biol. 2000, 10, 1032–1040. [Google Scholar] [CrossRef] [Green Version]
- Stadler, R.; Wright, K.M.; Lauterbach, C.; Amon, G.; Gahrtz, M.; Feuerstein, A.; Oparka, K.J.; Sauer, N. Expression of GFP-fusions in Arabidopsis companion cells reveals non-specific protein trafficking into sieve elements and identifies a novel post-phloem domain in roots. Plant J. 2005, 41, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Liljegren, S. Phloroglucinol stain for lignin. Cold Spring Harb. Protoc. 2010, 2010, pdb.prot4954. [Google Scholar] [CrossRef]
- Ruiz-Medrano, R.; Xoconostle-Cazares, B.; Lucas, W. Phloem long-distance transport of CmNACP mRNA: Implications for supracellular regulation in plants. Development 1999, 126, 4405–4419. [Google Scholar] [CrossRef]
- Lucas, W.J.; Yoo, B.-C.; Kragler, F. RNA as a long-distance information macromolecule in plants. Nat. Rev. Mol. Cell Biol. 2001, 2, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, L.; Kragler, F. The phloem-delivered RNA pool contains small noncoding RNAs and interferes withtranslation. Plant Physiol. 2009, 150, 378–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderwood, A.; Kopriva, S.; Morris, R.J. Transcript abundance axplains mRNA mobility data in Arabidopsis thaliana. Plant Cell 2016, 28, 610–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haupt, S.; Oparka, K.J.; Sauer, N.; Neumann, S. Macromolecular trafficking between Nicotiana tabacum and the holoparasite Cuscuta reflexa. J. Exp. Bot. 2001, 52, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Birschwilks, M.; Haupt, S.; Hofius, D.; Neumann, S. Transfer of phloem-mobile substances from the host plants to the holoparasite Cuscuta sp. J. Exp. Bot. 2006, 57, 911–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Qu, F.; Li, Z.; Doohan, D. Inter-species protein trafficking endows dodder (Cuscuta pentagona) with a host-specific herbicide-tolerant trait. New Phytol. 2013, 198, 1017–1022. [Google Scholar] [CrossRef]
- Chen, Q.; Payyavula, R.S.; Chen, L.; Zhang, J.; Zhang, C.; Turgeon, R. FLOWERING LOCUS T mRNA is synthesized in specialized companion cells in Arabidopsis and Maryland Mammoth tobacco leaf veins. Proc. Natl. Acad. Sci. USA 2018, 115, 2830–2835. [Google Scholar] [CrossRef] [Green Version]
- Slewinski, T.L.; Zhang, C.; Turgeon, R. Structural and functional heterogeneity in phloem loading and transport. Front. Plant Sci. 2013, 4, 244. [Google Scholar] [CrossRef] [Green Version]
- Shahid, S.; Kim, G.; Johnson, N.R.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; de Pamphilis, C.W.; Westwood, J.H.; et al. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature 2018, 553, 82–85. [Google Scholar] [CrossRef]
- Hozumi, A.; Bera, S.; Fujiwara, D.; Obayashi, T.; Yokoyama, R.; Nishitani, K.; Aoki, K. Arabinogalactan proteins accumulate in the cell walls of searching hyphae of the stem parasitic plants, Cuscuta campestris and Cuscuta japonica. Plant Cell Physiol. 2017, 58, 1868–1877. [Google Scholar] [CrossRef]
- Lao, J.; Oikawa, A.; Bromley, J.R.; McInerney, P.; Suttangkakul, A.; Smith-Moritz, A.M.; Plahar, H.; Chiu, T.-Y.; Fernández-Niño, S.M.G.; Ebert, B.; et al. The plant glycosyltransferase clone collection for functional genomics. Plant J. 2014, 79, 517–529. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Mitra, P.P.; Loqué, D. Histochemical Staining of Arabidopsis thaliana Secondary Cell Wall Elements. J. Vis. Exp. JoVE 2014, 87, 51381. [Google Scholar] [CrossRef] [Green Version]
- Guan, D.; Yan, B.; Thieme, C.; Hua, J.; Zhu, H.; Boheler, K.R.; Zhao, Z.; Kragler, F.; Xia, Y.; Zhang, S. PlaMoM: A comprehensive database compiles plant mobile macromolecules. Nucleic. Acids Res. 2017, 45, D1021–D1028. [Google Scholar] [CrossRef]
Figure 1.
Histochemical localization of β-glucuronidase. Haustoria between Arabidopsis and Cuscuta campestris were transversely cross-sectioned (as indicated by the red arrow) (A). C. campestris was inoculated on stems of 3-week-old Arabidopsis plants; wild type (WT) (B), 35S:GUS (C,D), and 35S:GUS-tRNAMet (E,F). D and F are high-magnification images of C and E, respectively. (B–F). The blue color of GUS activity in C. campestris is indicated by yellow arrows (E,F). Asterisks indicate haustoria. Scale bar: 500 μm.
Figure 1.
Histochemical localization of β-glucuronidase. Haustoria between Arabidopsis and Cuscuta campestris were transversely cross-sectioned (as indicated by the red arrow) (A). C. campestris was inoculated on stems of 3-week-old Arabidopsis plants; wild type (WT) (B), 35S:GUS (C,D), and 35S:GUS-tRNAMet (E,F). D and F are high-magnification images of C and E, respectively. (B–F). The blue color of GUS activity in C. campestris is indicated by yellow arrows (E,F). Asterisks indicate haustoria. Scale bar: 500 μm.

Figure 2.
GUS mRNA movement. Cuscuta campestris was inoculated on stems of three-week-old Arabidopsis; 35:GUS (A,B), 35S:GUS-tRNAMet (C,D), 35S:GUS-tRNAMet-dDT (E,F), 35:ER-GUS (G,H), 35S:ER-GUS-tRNAMet (I,J), and 35S:ER-GUS-tRNAMet-dDT (K,L). Haustoria between Arabidopsis and C. campestris were longitudinally (A,C,E,G,I,K) and transversely (B,D,F,H,J,L) cross-sectioned. Asterisks indicate haustoria. GUS mRNA was detected in C. campestris stems on the 35S:GUS-tRNA-Met, 35S:GUS-tRNA-Met-dDT, 35S:ER-GUS-tRNA-Met, and 35S:ER-GUS-tRNA-Met-dDT. C. campestris Actin8 (CcActin8) was used as a reference gene (M). Scale bar: 500 μm.
Figure 2.
GUS mRNA movement. Cuscuta campestris was inoculated on stems of three-week-old Arabidopsis; 35:GUS (A,B), 35S:GUS-tRNAMet (C,D), 35S:GUS-tRNAMet-dDT (E,F), 35:ER-GUS (G,H), 35S:ER-GUS-tRNAMet (I,J), and 35S:ER-GUS-tRNAMet-dDT (K,L). Haustoria between Arabidopsis and C. campestris were longitudinally (A,C,E,G,I,K) and transversely (B,D,F,H,J,L) cross-sectioned. Asterisks indicate haustoria. GUS mRNA was detected in C. campestris stems on the 35S:GUS-tRNA-Met, 35S:GUS-tRNA-Met-dDT, 35S:ER-GUS-tRNA-Met, and 35S:ER-GUS-tRNA-Met-dDT. C. campestris Actin8 (CcActin8) was used as a reference gene (M). Scale bar: 500 μm.

Figure 3.
Localization of GUS activity in C. campestris stems on Arabidopsis. 35S:GUS-tRNAMet (A–C,E,H,J), 35S:GUS-tRNAMet-dDT (F,I,K), and wild type (WT) (D,G). (A–C) Longitudinally sectioned C. campestris flowers from plants on a 35S:GUS-tRNAMet host. (B,C) High-magnification images of (A). White arrows indicate GUS signals in flower and peduncle. (D–F) C. campestris apices (segment 1–12 cm from host) and (G–K) stems (segment 0–2 cm) were GUS stained, embedded in paraffin, and (G,I) longitudinally, or (J,K) transversely, sectioned in 20 μm-thickness. Black arrows indicate the apical termini of sieve tube. PX, parasite xylem. Scale bars: 100 μm.
Figure 3.
Localization of GUS activity in C. campestris stems on Arabidopsis. 35S:GUS-tRNAMet (A–C,E,H,J), 35S:GUS-tRNAMet-dDT (F,I,K), and wild type (WT) (D,G). (A–C) Longitudinally sectioned C. campestris flowers from plants on a 35S:GUS-tRNAMet host. (B,C) High-magnification images of (A). White arrows indicate GUS signals in flower and peduncle. (D–F) C. campestris apices (segment 1–12 cm from host) and (G–K) stems (segment 0–2 cm) were GUS stained, embedded in paraffin, and (G,I) longitudinally, or (J,K) transversely, sectioned in 20 μm-thickness. Black arrows indicate the apical termini of sieve tube. PX, parasite xylem. Scale bars: 100 μm.

Figure 4.
GUS activity detected in the cells adjacent to the aniline-blue-stained sieve tube. A 20 μm-thick paraffin section of Cuscuta campestris stem on an Arabidopsis 35S:GUS-tRNAMet host was stained with X-gluc for 24 h and aniline blue for 45 min. (A) Bright field image by upright microscope. (B) Transmission image by confocal laser scanning microscopy. (C) Fluorescent image of aniline blue-stained sieve plates by confocal laser scanning microscopy. (D) Overlay image of (B,C). GUS activity (red arrows) was detected in the cells adjacent to the aniline-blue-stained sieve tube (yellow arrows). Scale bar: 100 μm.
Figure 4.
GUS activity detected in the cells adjacent to the aniline-blue-stained sieve tube. A 20 μm-thick paraffin section of Cuscuta campestris stem on an Arabidopsis 35S:GUS-tRNAMet host was stained with X-gluc for 24 h and aniline blue for 45 min. (A) Bright field image by upright microscope. (B) Transmission image by confocal laser scanning microscopy. (C) Fluorescent image of aniline blue-stained sieve plates by confocal laser scanning microscopy. (D) Overlay image of (B,C). GUS activity (red arrows) was detected in the cells adjacent to the aniline-blue-stained sieve tube (yellow arrows). Scale bar: 100 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Percent of Cuscuta campestris haustoria showing GUS enzyme activity.
| Arabidopsis Lines | Number of Cuscuta Haustoria | Total Number of Samples | % with GUS Detection | |
|---|---|---|---|---|
| GUS Detected | No GUS | |||
| Wild type Col-0 | 0 | 13 | 13 | 0 |
| 35S:empty (pEarleygate100) | 0 | 10 | 10 | 0 |
| 35S:GUS | 0 | 35 | 35 | 0 |
| 35S:GUS-tRNAMET | 39 | 10 | 49 | 80 |
| 35S:GUS-tRNAMET dDT | 14 | 20 | 34 | 41 |
| 35S:GUS-tRNAGly | 12 | 20 | 32 | 38 |
| 35S:GUS-tRNAIle | 11 | 26 | 37 | 30 |
| 35S:GUS-tRNAMET dAT | 0 | 12 | 12 | 0 |
| 35S:ER-GUS | 0 | 12 | 12 | 0 |
| 35S:ER-GUS-tRNAMET | 9 | 17 | 26 | 35 |
| 35S:ER-GUS-tRNAMET dDT | 13 | 20 | 33 | 39 |
| 35S:ER-GUS-tRNAGly | 11 | 17 | 28 | 39 |
| 35S:ER-GUS-tRNAIle | 8 | 19 | 27 | 30 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, S.-Y.; Shimizu, K.; Brown, J.; Aoki, K.; Westwood, J.H. Mobile Host mRNAs Are Translated to Protein in the Associated Parasitic Plant Cuscuta campestris. Plants 2022, 11, 93. https://doi.org/10.3390/plants11010093
AMA Style
Park S-Y, Shimizu K, Brown J, Aoki K, Westwood JH. Mobile Host mRNAs Are Translated to Protein in the Associated Parasitic Plant Cuscuta campestris. Plants. 2022; 11(1):93. https://doi.org/10.3390/plants11010093
Chicago/Turabian StylePark, So-Yon, Kohki Shimizu, Jocelyn Brown, Koh Aoki, and James H. Westwood. 2022. "Mobile Host mRNAs Are Translated to Protein in the Associated Parasitic Plant Cuscuta campestris" Plants 11, no. 1: 93. https://doi.org/10.3390/plants11010093
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.